Planted or Natural Pine Forests, Which One Will Better Recover after Drought? Insights from Tree Growth and Stable C and H Isotopes

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites and Tree Species
2.2. Climate Data
2.3. Field Sampling and Dendrochronological Data
2.4. Processing Tree-Ring Width Data
2.5. Isotopic Analyses
2.6. Statistical Analysis
3. Results
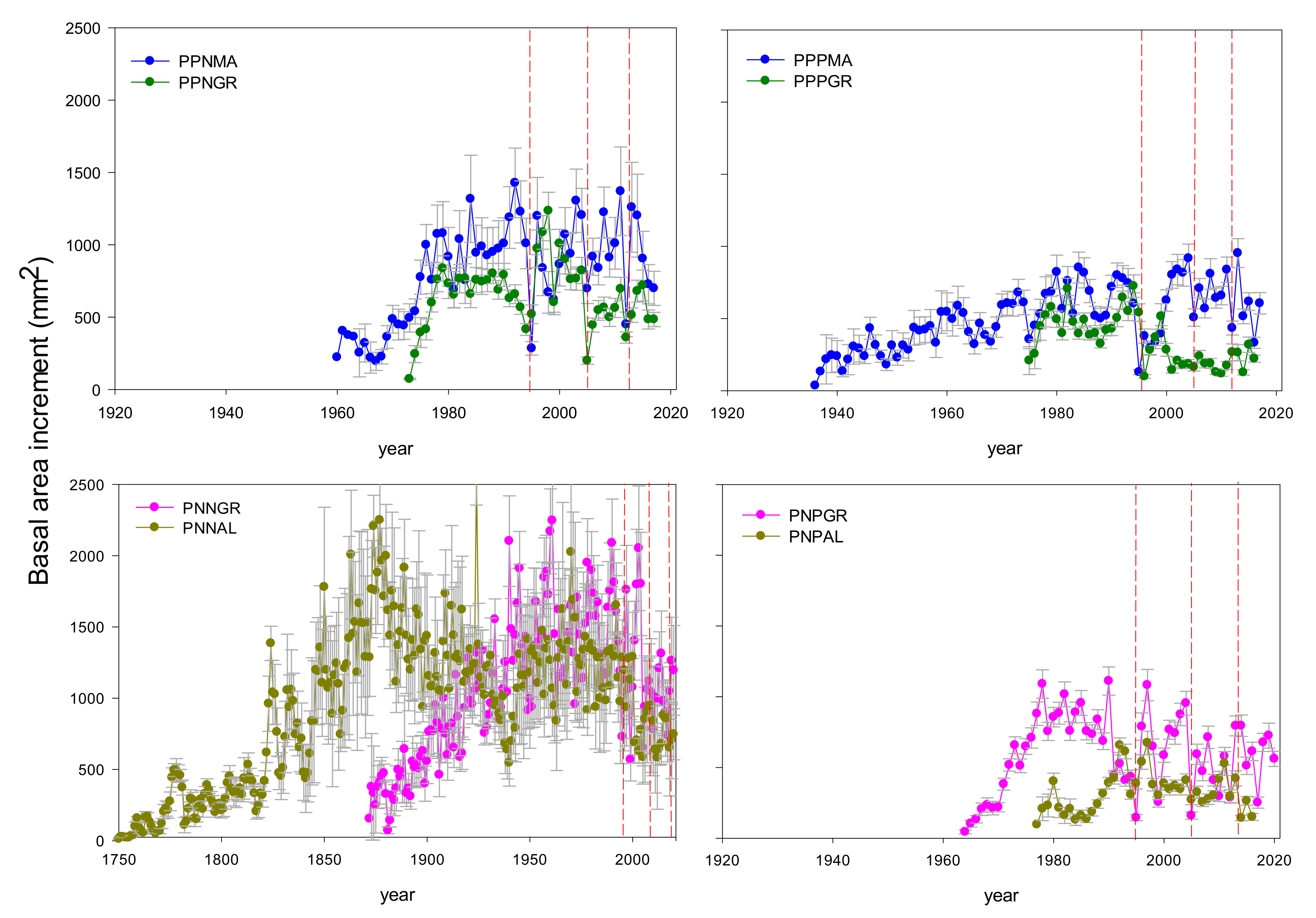
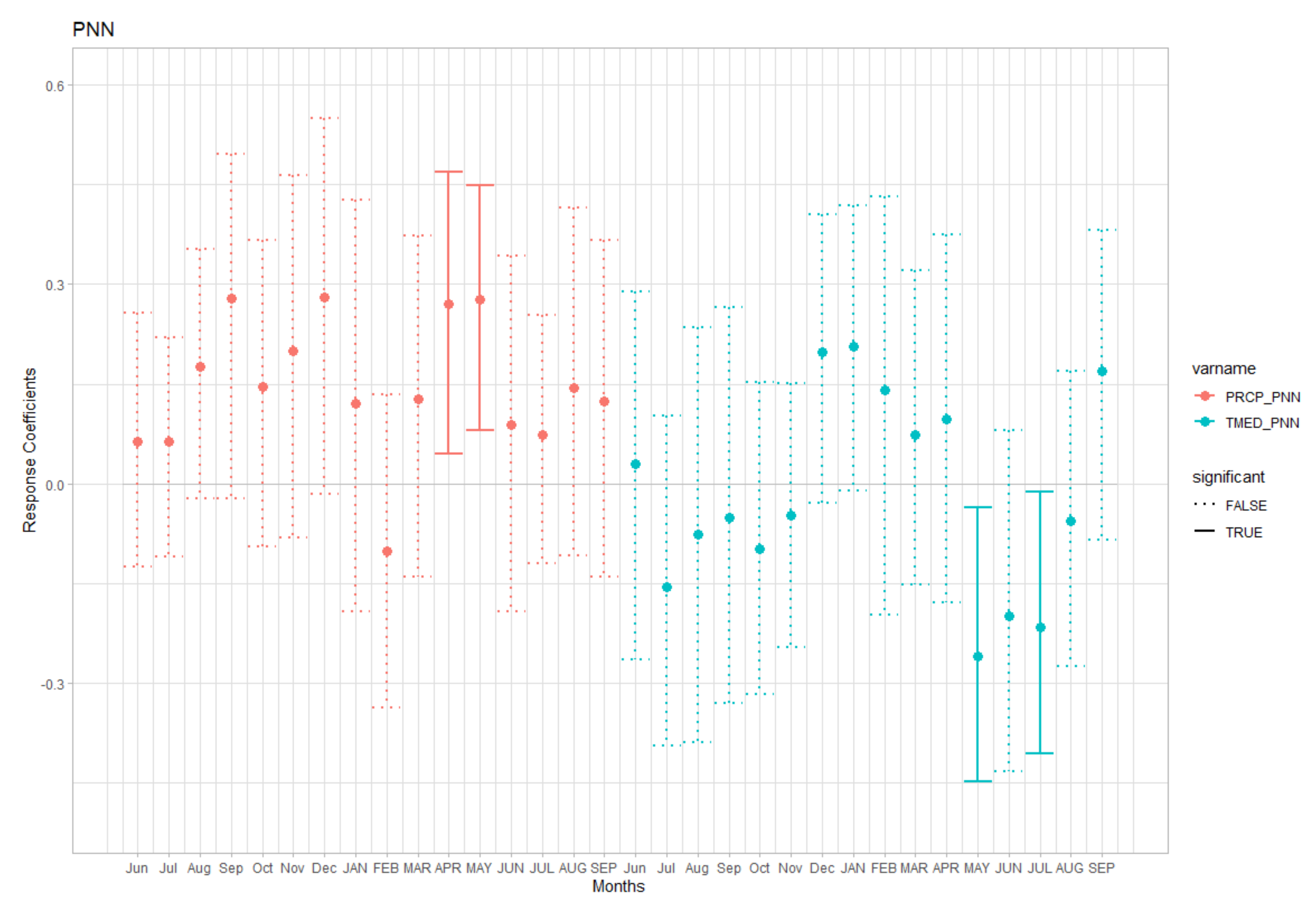
3.1. Growth Patterns and Climate-Growth Relationships
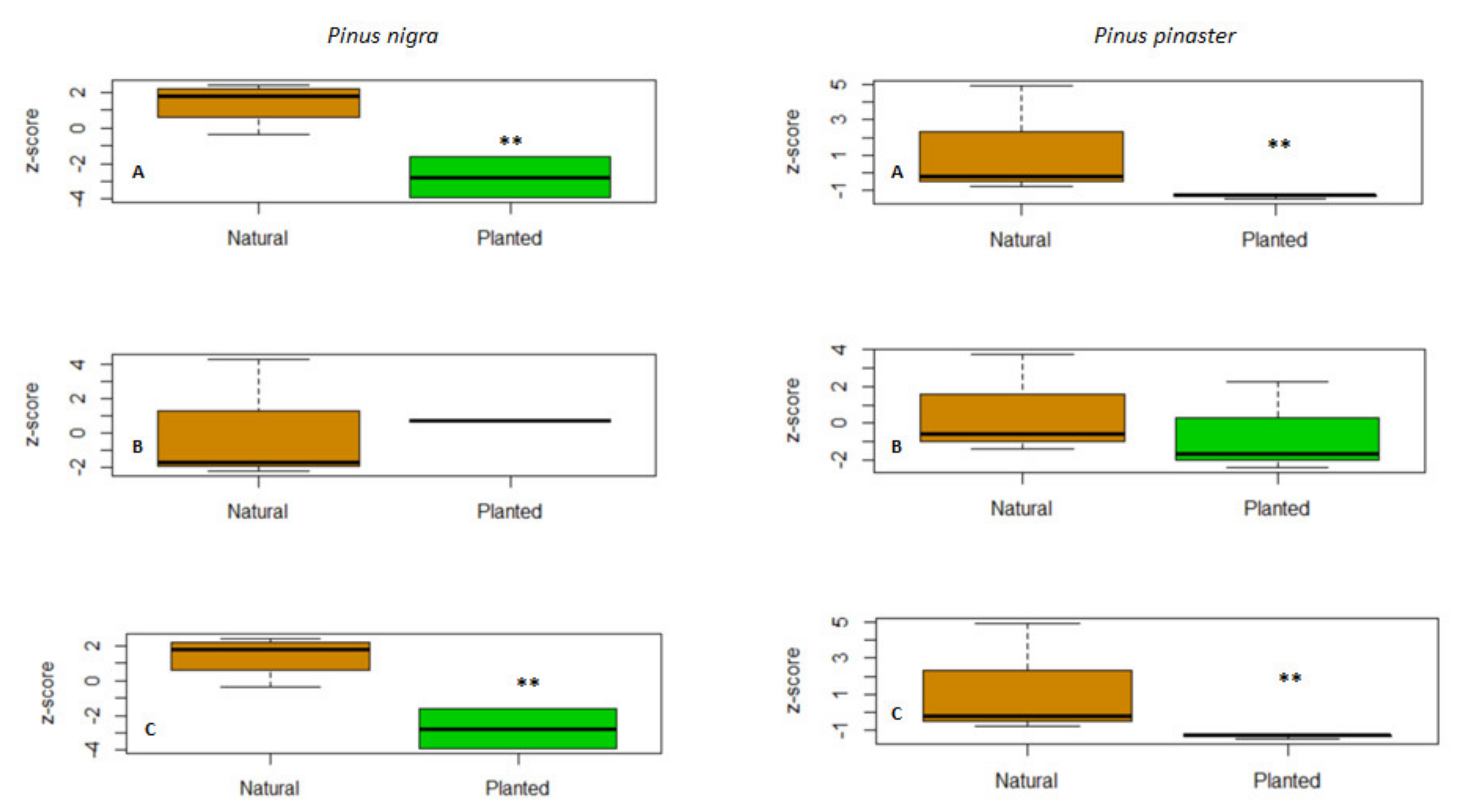
3.2. Growth Resilience Indices
3.3. Stable Isotopes (δ13C, δ2H)
4. Discussion
4.1. Growth Patterns and Responses to Climate in Planted and Natural Forests
4.2. Improved Growth Resilience in Natural Forests
4.3. Forest Type and Drought Effects on Isotopic Signals (δ13C and δ2H)
4.4. Implications for Forest Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early-warning signals of dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; Lopez-Moreno, J.I.; Azorin-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Navarro-Cerrillo, R.M.; Camarero, J.J.; Fernández-Cancio, A. Selective drought-induced decline of pine species in southeastern Spain. Clim. Chang. 2012, 113, 767–785. [Google Scholar] [CrossRef]
- Rodriguez-Vallejo, C.; Navarro-Cerrillo, R.M. Contrasting Response to Drought and Climate of Planted and Natural Pinus pinaster Aiton Forests in Southern Spain. Forests 2019, 10, 603. [Google Scholar] [CrossRef]
- MAGRAMA. La Restauración forestall en España: 75 años de una Ilusión; Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente Secretaría General Técnica Centro de Publicaciones: Madrid, Spain, 2017. [Google Scholar]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Hendrey, G.; Niu, S.; McDowell, N.; Allen, C.D. Tree mortality in a warming world: Causes, patterns, and implications. Environ. Res. Lett. 2022, 17, 030201. [Google Scholar] [CrossRef]
- Rodriguez-Vallejo, C.; Navarro-Cerrillo, R.M.; Manzanedo, R.D.; Rodriguez, G.P.; Gazol, A.; Camarero, J.J. High resilience, but low viability, of pine plantations in the face of a shift towards a drier climate. For. Ecol. Manag. 2021, 479, 118537. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Navarro, R.M.; Camarero, J.J.; Fernández-Cancio, A. Drought-induced growth decline of Aleppo and maritime pine forests in south-eastern Spain. For. Syst. 2010, 19, 458. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Sánchez-Salguero, R.; Rodriguez, C.; Lazo, J.D.; Moreno-Rojas, J.M.; Palacios-Rodriguez, G.; Camarero, J.J. Is thinning an alternative when trees could die in response to drought? The case of planted Pinus nigra and P. Sylvestris stands in southern Spain. For. Ecol. Manag. 2019, 433, 313–324. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Camarero, J.J.; Dobbertin, M.; Fernández-Cancio, A.; Vilà-Cabrera, A.; Manzanedo, R.D.; Zavala, M.A.; Navarro-Cerrillo, R.M. Contrasting vulnerability and resilience to drought-induced decline of densely planted vs. natural rear-edge Pinus nigra forests. For. Ecol. Manag. 2013, 310, 956–967. [Google Scholar] [CrossRef]
- Camarero, J.J. The drought-dieback-death conundrum in trees and forests. Plant Ecol. Divers. 2021, 14, 1–12. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Sánchez-Salguero, R.; Zavala, M.A.; Serra-Maluquer, X.; Gutiérrez, E.; de Luis, M.; Sangüesa-Barreda, G.; Novak, K.; Rozas, V.; et al. Tree growth response to drought partially explains regional-scale growth and mortality patterns in Iberian forests. Ecol. Appl. 2022, 32, e2589. [Google Scholar] [CrossRef]
- Gessler, A.; Ferrio, J.P.; Hommel, R.; Treydte, K.; Werner, R.A.; Monson, R.K. Stable isotopes in tree rings: Towards a mechanistic understanding of isotope fractionation and mixing processes from the leaves to the wood. Tree Physiol. 2014, 34, 796–818. [Google Scholar] [CrossRef] [PubMed]
- Gessler, A.; Cailleret, M.; Joseph, J.; Schönbeck, L.; Schaub, M.; Lehmann, M.; Treydte, K.; Rigling, A.; Timofeeva, G.; Saurer, M. Drought induced tree mortality-a tree-ring isotope based conceptual model to assess mechanisms and predispositions. New Phytol. 2018, 219, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Cherubini, P.; Battipaglia, G.; Innes, J.L. Tree vitality and forest health: Can tree-ring stable isotopes be used as indicators? Curr. For. Rep. 2021, 7, 69–80. [Google Scholar] [CrossRef]
- Salazar-Tortosa, D.; Castro, J.; de Casas, R.R.; Viñegla, B.; Sánchez-Cañete, E.P.; Villar-Salvador, P. Gas exchange at whole plant level shows that a less conservative water use is linked to a higher performance in three ecologically distinct pine species. Environ. Res. Lett. 2018, 13, 045004. [Google Scholar] [CrossRef]
- Maseyk, K.; Hemming, D.; Angert, A.; Leavitt, S.W.; Yakir, D. Increase in water-use efficiency and underlying processes in pine forests across a precipitation gradient in the dry Mediterranean region over the past 30 years. Oecologia 2011, 167, 573–585. [Google Scholar] [CrossRef]
- Marshall, J.D.; Monserud, R.A. Co-occurring species differ in tree-ring δ18O trends. Tree Physiol. 2006, 26, 1055–1066. [Google Scholar] [CrossRef]
- Shestakova, T.A.; Camarero, J.J.; Ferrio, J.P.; Knorre, A.A.; Gutiérrez, E.; Voltas, J. Increasing drought effects on five european pines modulate δ13C-growth coupling along a Mediterranean altitudinal gradient. Funct. Ecol. 2017, 31, 1359–1370. [Google Scholar] [CrossRef]
- Battipaglia, G.; Cherubini, P. Stable Isotopes in Tree Rings of Mediterranean Forests. In Stable Isotopes in Tree Rings; Siegwolf, R.T.W., Brooks, J.R., Roden, J., Saurer, M., Eds.; Tree Physiology 8; Springer: Cham, Switzerland, 2022; pp. 605–629. [Google Scholar] [CrossRef]
- De La Serrana, R.G.; Vilagrosa, A.; Alloza, J.A. Pine mortality in southeast Spain after an extreme dry and warm year: Interactions among drought stress, carbohydrates and bark beetle attack. Trees 2015, 29, 1791–1804. [Google Scholar] [CrossRef]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Linares, J.-C.; Delgado-Huertas, A.; Camarero, J.J.; Merino, J.; Carreira, J.A. Competition and drought limit the response of water-use efficiency to rising atmospheric carbon dioxide in the Mediterranean fir Abies pinsapo. Oecologia 2009, 161, 611–624. [Google Scholar] [CrossRef]
- Moreno-Gutiérrez, C.; Battipaglia, G.; Cherubini, P.; Saurer, M.; Nicolás, E.; Contreras, S.; Querejeta, J.I. Stand structure modulates the long-term vulnerability of Pinus halepensis to climatic drought in a semiarid Mediterranean ecosystem. Plant Cell Environ. 2012, 35, 1026–1039. [Google Scholar] [CrossRef] [PubMed]
- Kannenberg, S.A.; Cabon, A.; Babst, F.; Belmecheri, S.; Delpierre, N.; Guerrieri, R.; Maxwell, J.T.; Meinzer, F.C.; Moore, D.J.; Pappas, C.; et al. Drought-induced decoupling between carbon uptake and tree growth impacts forest carbon turnover time. Agric. For. Meteorol. 2022, 322, 108996. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Rodriguez-Vallejo, C.; Silveiro, E.; Hortal, A.; Palacios-Rodríguez, G.; Duque-Lazo, J.; Camarero, J.J. Cumulative Drought Stress Leads to a Loss of Growth Resilience and Explains Higher Mortality in Planted than in Naturally Regenerated Pinus pinaster Stands. Forests 2018, 9, 358. [Google Scholar] [CrossRef]
- Bogino, S.M.; Bravo, F. Growth response of Pinus pinaster Ait. to climatic variables in central Spanish forests. Ann. For. Sci. 2008, 65, 506. [Google Scholar] [CrossRef]
- Caminero, L.; Génova, M.; Camarero, J.J.; Sánchez-Salguero, R. Growth responses to climate and drought at the southernmost European limit of Mediterranean Pinus pinaster forests. Dendrochronologia 2018, 48, 20–29. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.; Sánchez-Salguero, R.; Herrera, R.; Ruiz, C.C.; Moreno-Rojas, J.; Manzanedo, R.; López-Quintanilla, J. Contrasting growth and water use efficiency after thinning in mixed Abies pinsapo-Pinus pinaster-Pinus sylvestris forests. J. For. Sci. 2016, 62, 53–64. [Google Scholar] [CrossRef]
- USDA Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; Soil Survey Staff; Agr. Handbook 436; USDA-NRCS: Washington, DC, USA, 1999; 869p.
- Lucas-Borja, M.E.; Bose, A.K.; Andivia, E.; Candel-Pérez, D.; Plaza-Álvarez, P.; Linares, J.C. Assessing tree drought resistance and climate-growth relationships under different tree age classes in a Pinus nigra Arn. ssp. salzmannii forest. Forests 2021, 12, 1161. [Google Scholar] [CrossRef]
- De Cáceres, M.; Martin-StPaul, N.; Turco, M.; Cabon, A.; Granda, V. Estimating daily meteorological data and downscaling climate models over landscapes. Environ. Model. Softw. 2018, 108, 186–196. [Google Scholar] [CrossRef]
- Hargreaves, G.H.; Samani, Z.A. Estimating Potential Evapotranspiration. J. Irrig. Drain. Div. 1982, 108, 225–230. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Biondi, F.; Qeadan, F. A Theory-Driven Approach to Tree-Ring Standardization: Defining the Biological Trend from Expected Basal Area Increment. Tree-Ring Res. 2008, 64, 81–96. [Google Scholar] [CrossRef]
- Bunn, A.G. Statistical and visual crossdating in R using the dplR library. Dendrochronologia 2010, 28, 251–258. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Jiménez-Morillo, N.T.; Palma, V.; Garcia, R.; Dias, C.B.; Cabrita, M.J. Combination of Stable Isotope Analysis and Chemometrics to Discriminate Geoclimatically and Temporally the Virgin Olive Oils from Three Mediterranean Countries. Foods 2020, 9, 1855. [Google Scholar] [CrossRef]
- González-Pérez, J.A.; Jiménez Morillo, N.T.; Rosa Arranz, J.M.; Knicker, H.; González-Vila, F.J. Pyrolysis-compound specific isotope analysis (Py-CSIA). Technique and applications. In Proceedings of the 21st International Symposium on Analytical and Applied. Pyrolysis, Nancy, France, 9–12 May 2016. [Google Scholar]
- San-Emeterio, L.M.; López-Núñez, R.; González-Vila, F.J.; González-Pérez, J.A. Evolution of composting process in maize biomass revealed by analytical pyrolysis (Py-GC/MS) and pyrolysis compound specific isotope analysis (Py-CSIA). Appl. Sci. 2021, 11, 6684. [Google Scholar] [CrossRef]
- Schimmelmann, A.; Qi, H.; Coplen, T.B.; Brand, W.A.; Fong, J.; Meier-Augenstein, W.; Kemp, H.F.; Toman, B.; Ackermann, A.; Assonov, S.; et al. Organic Reference Materials for Hydrogen, Carbon, and Nitrogen Stable Isotope-Ratio Measurements: Caffeines, n-Alkanes, Fatty Acid Methyl Esters, Glycines, l-Valines, Polyethylenes, and Oils. Anal. Chem. 2016, 88, 4294–4302. [Google Scholar] [CrossRef]
- Coplen, T.B. Guidelines and recommended terms for expression of stable-isotope-ratio and gas-ratio measurement results. Rapid Commun. Mass Spectrom. 2011, 25, 2538–2560. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Leary, M.H.; Berry, J.A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust. J. Plant Physiol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Belmecheri, S.; Lavergne, A. Compiled records of atmospheric CO2 concentrations and stable carbon isotopes to reconstruct climate and derive plant ecophysiological indices from tree rings. Dendrochronologia 2020, 63, 125748. [Google Scholar] [CrossRef]
- Pontius, J.; Hallett, R. Comprehensive Methods for Earlier Detection and Monitoring of Forest Decline. For. Sci. 2014, 60, 1156–1163. [Google Scholar] [CrossRef]
- R Core Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Zang, C.; Biondi, F. treeclim: An R package for the numerical calibration of proxy-climate relationships. Ecography 2014, 38, 431–436. [Google Scholar] [CrossRef]
- Rubio-Cuadrado, Á.; López, R.; Rodríguez-Calcerrada, J.; Gil, L. Stress and tree mortality in Mediterranean pine forests: Anthropogenic influences. In Pines and Their Mixed Forest Ecosystems in the Mediterranean Basin; Springer: Cham, Switzerland, 2021; pp. 141–181. [Google Scholar] [CrossRef]
- Fernández-de-Uña, L.; McDowell, N.G.; Canellas, I.; Gea-Izquierdo, G. Disentangling the effect of competition, CO2 and climate on intrinsic water-use efficiency and tree growth. J. Ecol. 2016, 104, 678–690. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Tardif, J.C.; Conciatori, F. Attributing forest responses to global-change drivers: Limited evidence of a CO2-fertilization effect in Iberian pine growth. J. Biogeogr. 2015, 42, 2220–2233. [Google Scholar] [CrossRef]
- Valeriano, C.; Gazol, A.; Colangelo, M.; Camarero, J.J. Drought Drives Growth and Mortality Rates in Three Pine Species under Mediterranean Conditions. Forests 2021, 12, 1700. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sánchez-Salguero, R. Effects of global change on tree growth and vigor of Mediterranean pines. In Pines and Their Mixed Forest Ecosystems in the Mediterranean Basin 2021; Springer: Cham, Switzerland, 2021; pp. 237–249. [Google Scholar]
- Manrique-Alba, A.; Beguería, S.; Tomas-Burguera, M.; Camarero, J. Increased Post-Drought Growth after Thinning in Pinus nigra Plantations. Forests 2021, 12, 985. [Google Scholar] [CrossRef]
- Xu, B.; Arain, M.A.; Black, T.A.; Law, B.E.; Pastorello, G.Z.; Chu, H. Seasonal variability of forest sensitivity to heat and drought stresses: A synthesis based on carbon fluxes from North American forest ecosystems. Glob. Chang. Biol. 2020, 26, 901–918. [Google Scholar] [CrossRef]
- Silvério, E.; Duque-Lazo, J.; Navarro-Cerrillo, R.M.; Pereña, F.; Palacios-Rodríguez, G. Resilience or Vulnerability of the Rear-Edge Distributions of Pinus halepensis and Pinus pinaster Plantations Versus that of Natural Populations, under Climate-Change Scenarios. For. Sci. 2020, 66, 178–190. [Google Scholar] [CrossRef]
- Frank, D.C.; Poulter, B.; Saurer, M.; Esper, J.; Huntingford, C.; Helle, G.; Treydte, K.; Zimmermann, N.E.; Schleser, G.H.; Ahlström, A.; et al. Water-use efficiency and transpiration across European forests during the Anthropocene. Nat. Clim. Chang. 2015, 5, 579–583. [Google Scholar] [CrossRef]
- Forrester, D.I. Transpiration and water-use efficiency in mixed-species forests versus monocultures: Effects of tree size, stand density and season. Tree Physiol. 2015, 35, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, T.J.; Del Campo, A.D.; Herrera, R.; Molina, A.J. Simultaneous assessment, through sap flow and stable isotopes, of water use efficiency (WUE) in thinned pines shows improvement in growth, tree-climate sensitivity and WUE, but not in WUEi. For. Ecol. Manag. 2016, 361, 298–308. [Google Scholar] [CrossRef]
- Linares, J.C.; Camarero, J.J. From pattern to process: Linking intrinsic water-use efficiency to drought-induced forest decline. Glob. Chang. Biol. 2012, 18, 1000–1015. [Google Scholar] [CrossRef]
- Voltas, J.; Lucabaugh, D.; Chambel, M.R.; Ferrio, J.P. Intraspecific variation in the use of water sources by the circum-Mediterranean conifer Pinus halepensis. New Phytol. 2015, 208, 1031–1041. [Google Scholar] [CrossRef]
- Volkmann, T.H.M.; Haberer, K.; Gessler, A.; Weiler, M. High-resolution isotope measurements resolve rapid ecohydrological dynamics at the soil–plant interface. New Phytol. 2016, 210, 839–849. [Google Scholar] [CrossRef]
- Cormier, M.-A.; Werner, R.A.; Sauer, P.E.; Gröcke, D.R.; Leuenberger, M.C.; Wieloch, T.; Schleucher, J.; Kahmen, A. 2H-fractionations during the biosynthesis of carbohydrates and lipids imprint a metabolic signal on the δ2H values of plant organic compounds. New Phytol. 2018, 218, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, M.M.; Vitali, V.; Schuler, P.; Leuenberger, M.; Saurer, M. More than climate: Hydrogen isotope ratios in tree rings as novel plant physiological indicator for stress conditions. Dendrochronologia 2020, 65, 125788. [Google Scholar] [CrossRef]
- Wang, J.; Fu, B.; Wang, L.; Lu, N.; Li, J. Water use characteristics of the common tree species in different plantation types in the Loess Plateau of China. Agric. For. Meteorol. 2020, 288–289, 108020. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Ubierna, N. Carbon isotope effects in relation to CO2 assimilation by tree canopies. In Stable Isotopes in Tree Rings; Siegwolf, R.T.W., Brooks, J.R., Roden, J., Saurer, M., Eds.; Tree Physiology 8; Springer: Cham, Switzerland, 2022; pp. 291–310. [Google Scholar]
- Lehmann, M.M.; Schuler, P.; Cormier, M.A.; Allen, S.T.; Leuenberger, M.; Voelker, S. The Stable Hydrogen Isotopic Signature: From Source Water to Tree Rings. In Stable Isotopes in Tree Rings; Siegwolf, R.T.W., Brooks, J.R., Roden, J., Saurer, M., Eds.; Tree Physiology 8; Springer: Cham, Switzerland, 2022; pp. 331–359. [Google Scholar] [CrossRef]
- Tyutkova, E.; Loskutov, S.; Petrov, I.; Dorzhiev, D. Seasonal biochemical changes in Betula tortuosa Ledeb. annual rings in Alpine forest-tundra of Kuznetsk Ala Tau Mountains. Wood Sci. Technol. 2022, 57, 289–306. [Google Scholar] [CrossRef]
- Moulin, J.C.; Ribeiro, D.D.S.; Vidaurre, G.B.; Mulin, L.B.; Moreira, S.I. Effect of drought stress on the formation and lignification of eucalyptus wood cells. IAWA J. 2022, 43, 263–275. [Google Scholar] [CrossRef]
- Meusburger, K.; Trotsiuk, V.; Schmidt-Walter, P.; Baltensweiler, A.; Brun, P.; Bernhard, F.; Walthert, L. Soil–plant interactions modulated water availability of Swiss forests during the 2015 and 2018 droughts. Glob. Change Biol. 2022, 28, 5928–5944. [Google Scholar] [CrossRef] [PubMed]
- Manrique-Alba, A.; Beguería, S.; Molina, A.J.; González-Sanchis, M.; Tomàs-Burguera, M.; del Campo, A.D.; Colangelo, M.; Camarero, J.J. Long-term thinning effects on tree growth, drought response and water use efficiency at two Aleppo pine plantations in Spain. Sci. Total. Environ. 2020, 728, 138536. [Google Scholar] [CrossRef] [PubMed]
- Sohn, J.A.; Saha, S.; Bauhus, J. Potential of forest thinning to mitigate drought stress: A meta-analysis. For. Ecol. Manag. 2016, 380, 261–273. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Stand Type | Site | Lat. (N) | Long. (W) | DBH (cm) | H (m) | G (m2 ha-1) | N (Trees ha-1) | Elev (m) | Exp | Slope (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pinus pinaster | N | PPNMA | 36.59 | 4.96 | 33.4(1.7) | 8.2(0.3) | 15.7 | 180 | 400 | SW | 35–65 |
| P | PPPMA | 36.49 | 5.12 | 32.2(3.2) | 8.8(0.4) | 10.6 | 130 | 260 | S | >65 | |
| N | PPNGR | 37.23 | 3.45 | 33.2(0.8) | 10.6 (0.6) | 20.7 | 240 | 1320 | NE | 35–65 | |
| P | PPPGR | 37.24 | 2.57 | 12.9(0.6) | 7.4(0.3) | 16.5 | 1250 | 1300 | NW | 35–65 | |
| Pinus nigra | N | PNNGR | 37.08 | 3.46 | 60.4(4.2) | 7.8(0.4) | 71.8 | 250 | 1700 | N | 35–65 |
| P | PNPGR | 37.04 | 3.55 | 27.4(1.1) | 13.1 (0.3) | 27.2 | 460 | 1425 | NW | 2 | |
| N | PNNAL | 37.26 | 2.52 | 81.2(5.9) | 9.5(0.3) | 41.5 | 80 | 1800 | NW | 35–65 | |
| P | PNPAL | 37.22 | 2.56 | 15.8(0.5) | 9.5(0.3) | 17.6 | 900 | 1725 | N | 35–65 |
| Species | Site | Timespan (No. Years) | TRW (cm) | BAI20 (cm2 Year−1) | MS | Rbar | AC1 | EPS |
|---|---|---|---|---|---|---|---|---|
| P. pinaster | PPNMA | 1960–2017 (57) | 2.90 (0.14) ** | 9.41 (1.60) *** | 0.332 | 0.594 | 0.668 | 0.957 |
| PPPMA | 1936–2017 (81) | 1.95 (0.09) | 6.44 (0.65) | 0.373 | 0.687 | 0.644 | 0.982 | |
| PPNGR | 1973–2017(45) | 2.56 (0.26) *** | 6.42 (0.51) *** | 0.312 | 0.476 | 0.868 | 0.948 | |
| PPPGR | 1975–2016(41) | 1.94 (0.31) | 2.28 (0.34) | 0.373 | 0.567 | 0.887 | 0.966 | |
| P. nigra | PNNGR | 1869–2020 (151) | 1.72 (0.05) *** | 11.65 (2.33) *** | 0.297 | 0.529 | 0.724 | 0.895 |
| PNPGR | 1964–2020 (56) | 2.07 (0.16) | 5.89 (0.52) | 0.451 | 0.618 | 0.784 | 0.973 | |
| PNNAL | 1747–2020 (273) | 1.05 (0.02) *** | 7.60 (2.43) ** | 0.345 | 0.503 | 0.741 | 0.853 | |
| PNPAL | 1977–2016 (39) | 2.12 (0.22) | 3.43 (0.36) | 0.338 | 0.510 | 0.796 | 0.949 |
| Species | Stand Type | Site | δ13C (‰) | δ2H (‰) | iWUE (μmol mol−1) |
|---|---|---|---|---|---|
| Pinus pinaster | Málaga | PPNMA | −25.11 (0.50). | −61.39 (1.24) * | 109.81 (5.83) |
| PPPMA | −24.43 (0.83) | −57.28 (3.27) | 116.97 (9.37) | ||
| Granada | PPNGR | −25.33 (0.47) * | −65.15 (1.80) | 110.05 (2.38) * | |
| PPPGR | −23.79 (0.31) | −68.56 (4.03) | 123.93 (3.78) | ||
| PPN | −25.24 (0.33) | −63.64 (1.29) | 108.95 (2.22) | ||
| PPP | −24.11 (0.43) | −60.93 (3.22) | 120.44 (4.93) | ||
| Pinus nigra | Granada | PNNGR | −24.51 (0.34) | −76.83 (6.04) * | 116.22 (3.88) |
| PNPGR | −23. 82 (0.57) | −66.69 (5.37) | 123.94 (6.27) | ||
| Almería | PNNAL | −25.65 (0.19) ** | −60.09 (1.22) * | 103.00 (2.11) * | |
| PNPAL | −24.28 (0.31) | −53.98 (1.75) | 118.86 (3.89) | ||
| PNN | −25.08 (0.25) | −69.29 (3.81) * | 109.61 (2.90) * | ||
| PNP | −24.05 (0.31) | −58.67 (2.23) | 121.40 (3.60) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-Cerrillo, R.M.; Cachinero-Vivar, A.M.; Ruiz-Gómez, F.J.; Camarero, J.J.; González-Pérez, J.A.; Pérez-Priego, Ó. Planted or Natural Pine Forests, Which One Will Better Recover after Drought? Insights from Tree Growth and Stable C and H Isotopes. Forests 2023, 14, 573. https://doi.org/10.3390/f14030573
Navarro-Cerrillo RM, Cachinero-Vivar AM, Ruiz-Gómez FJ, Camarero JJ, González-Pérez JA, Pérez-Priego Ó. Planted or Natural Pine Forests, Which One Will Better Recover after Drought? Insights from Tree Growth and Stable C and H Isotopes. Forests. 2023; 14(3):573. https://doi.org/10.3390/f14030573
Chicago/Turabian StyleNavarro-Cerrillo, Rafael Mª, Antonio M. Cachinero-Vivar, Francisco J. Ruiz-Gómez, J. Julio Camarero, José A. González-Pérez, and Óscar Pérez-Priego. 2023. "Planted or Natural Pine Forests, Which One Will Better Recover after Drought? Insights from Tree Growth and Stable C and H Isotopes" Forests 14, no. 3: 573. https://doi.org/10.3390/f14030573
APA StyleNavarro-Cerrillo, R. M., Cachinero-Vivar, A. M., Ruiz-Gómez, F. J., Camarero, J. J., González-Pérez, J. A., & Pérez-Priego, Ó. (2023). Planted or Natural Pine Forests, Which One Will Better Recover after Drought? Insights from Tree Growth and Stable C and H Isotopes. Forests, 14(3), 573. https://doi.org/10.3390/f14030573