
Spatial Habitat Structure Assembles Willow-Dependent Communities across the Primary Successional Watersheds of Mount St. Helens, USA

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
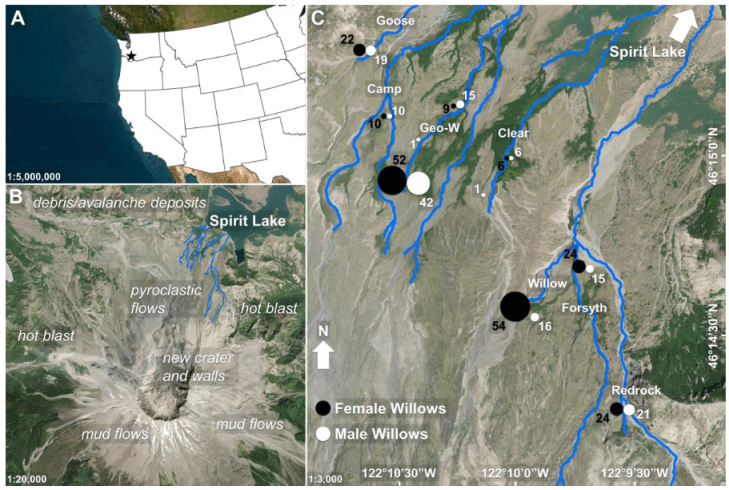
2.1. Site Description
2.2. Field Data Collection
2.3. Leaf Litter Chemistry
2.4. Leaf Area Measurements
2.5. Landscape Connectivity
2.6. Statistical Analyses
3. Results
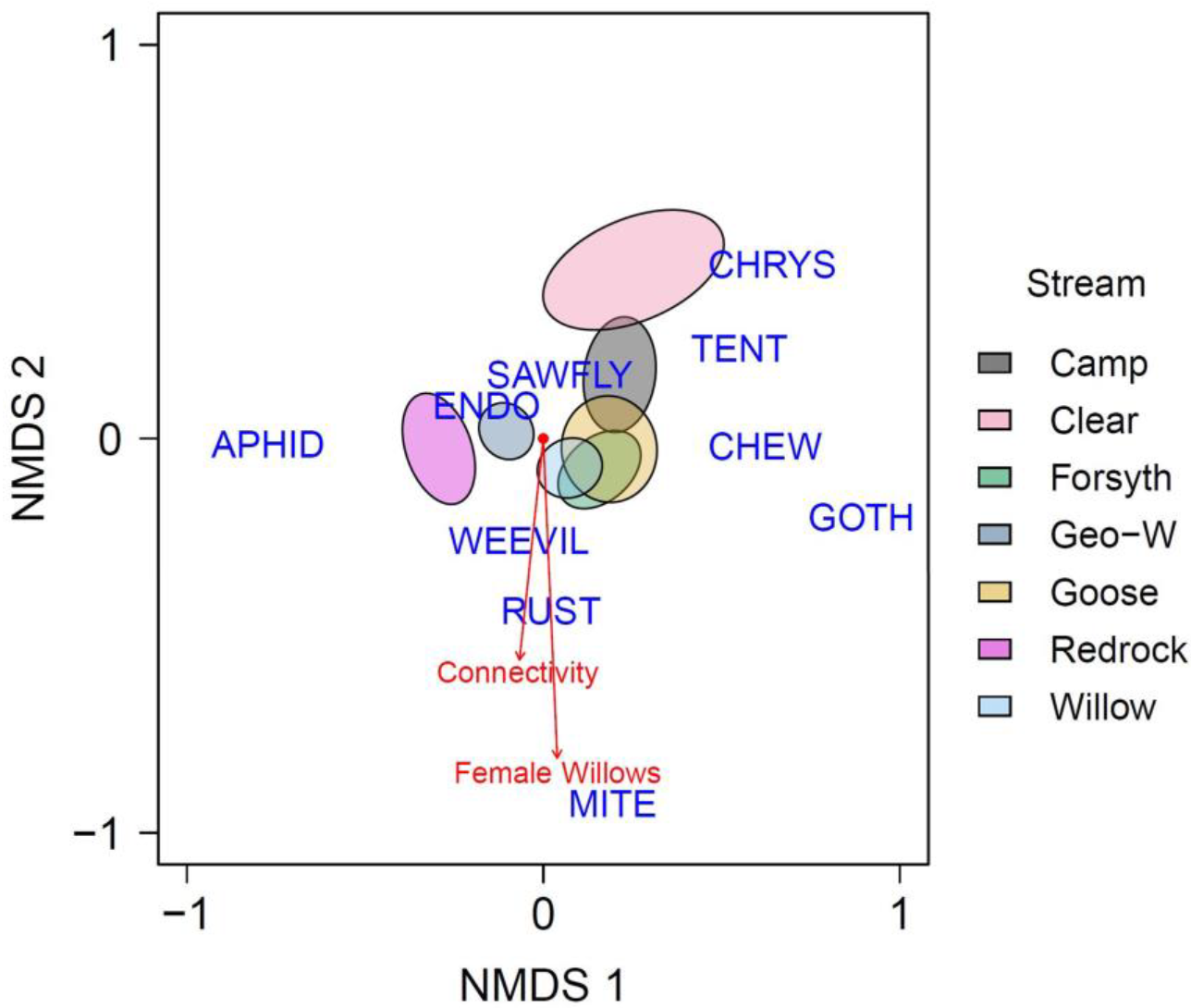
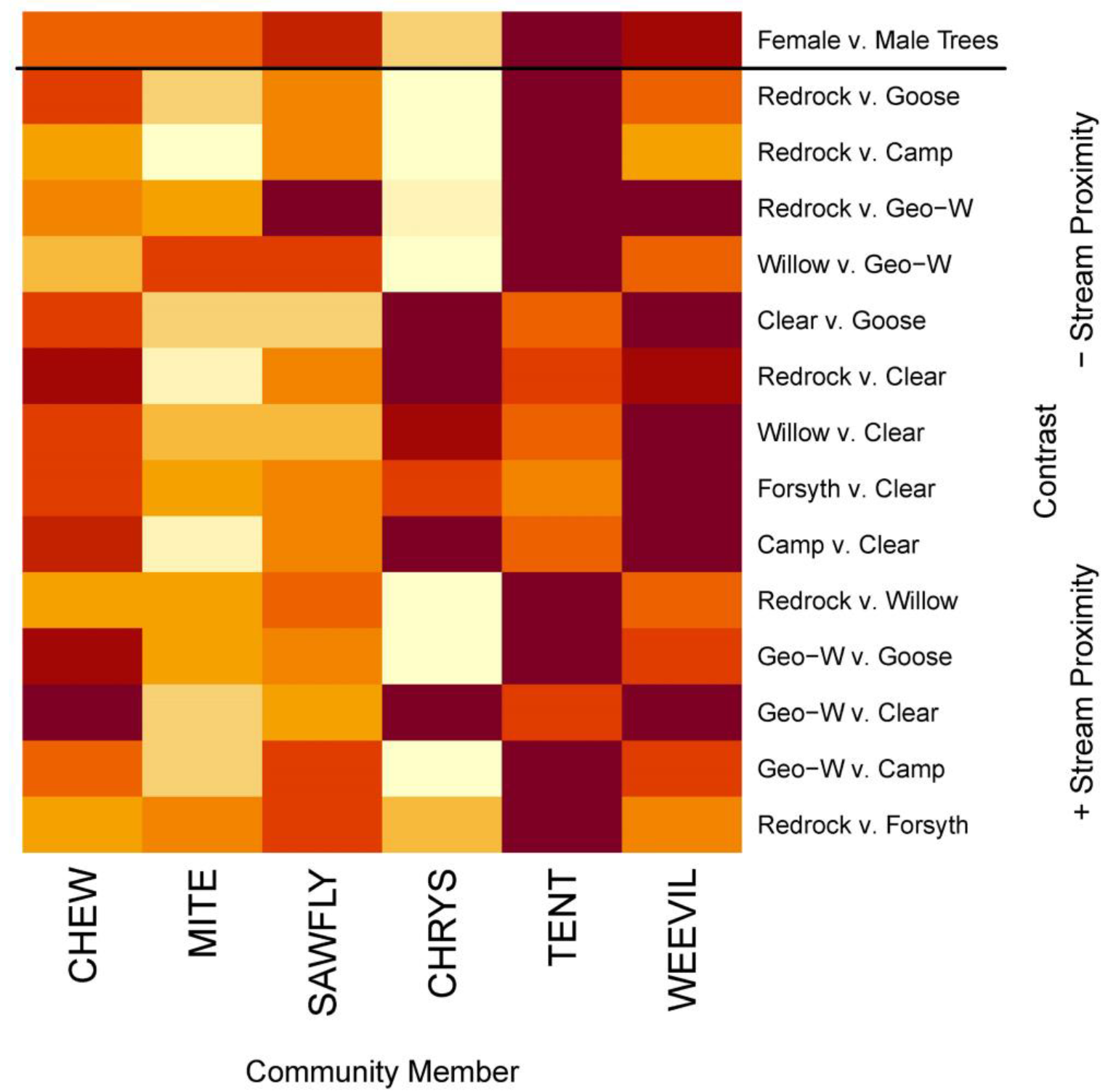
3.1. Landscape-Level Models
3.2. Leaf Litter Chemistry Models
3.3. Leaf Area Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaynor, K.M.; Hojnowski, C.E.; Carter, N.H.; Brashares, J.S. The influence of human disturbance on wildlife nocturnality. Science 2018, 360, 1232–1235. [Google Scholar] [CrossRef] [PubMed]
- Laurance, W.F.; Useche, D.C.; Rendeiro, J.; Kalka, M.; Bradshaw, C.J.A.; Sloan, S.P.; Laurance, S.G.; Campbell, M.; Abernethy, K.; Alvarez, P.; et al. Averting biodiversity collapse in tropical forest protected areas. Nature 2012, 489, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Lindenmayer, D.B.; Likens, G.E.; Franklin, J.F. Rapid responses to facilitate ecological discoveries from major disturbances. Front. Ecol. Environ. 2010, 8, 527–532. [Google Scholar] [CrossRef]
- Morcillo, D.O.; Steiner, U.K.; Grayson, K.L.; Ruiz-Lambides, A.V.; Hernández-Pacheco, R. Hurricane-induced demographic changes in a non-human primate population. R. Soc. Open Sci. 2020, 7, 200173. [Google Scholar] [CrossRef]
- Major, J.J.; Pierson, T.C.; Dinehart, R.L.; Costa, J.E. Sediment yield following severe volcanic disturbance—A two-decade perspective from Mount St. Helens. Geology 2000, 28, 819–822. [Google Scholar] [CrossRef]
- Zobel, D.B.; Antos, J.A.; Fischer, D.G. Secondary disturbance following a deposit of volcanic tephra: A 30-year record from old-growth forest understory. Can. J. For. Res. 2021, 51, 1541–1549. [Google Scholar] [CrossRef]
- Jentsch, A.; White, P. A theory of pulse dynamics and disturbance in ecology. Ecology 2019, 100, e02734. [Google Scholar] [CrossRef]
- Shackelford, N.; Starzomski, B.M.; Banning, N.C.; Battaglia, L.L.; Becker, A.; Bellingham, P.J.; Bestelmeyer, B.; Catford, J.A.; Dwyer, J.M.; Dynesius, M.; et al. Isolation predicts compositional change after discrete disturbances in a global meta-study. Ecography 2017, 40, 1256–1266. [Google Scholar] [CrossRef]
- Banks, S.C.; Cary, G.J.; Smith, A.L.; Davies, I.D.; Driscoll, D.A.; Gill, A.M.; Lindenmayer, D.B.; Peakall, R. How does ecological disturbance influence genetic diversity? Trends Ecol. Evol. 2013, 28, 670–679. [Google Scholar] [CrossRef]
- Holyoak, M.; Caspi, T.; Redosh, L.W. Integrating disturbance, seasonality, multi-year temporal dynamics, and dormancy into the dynamics and conservation of metacommunities. Front. Ecol. Evol. 2020, 8, 571130. [Google Scholar] [CrossRef]
- Urban, M.C. Disturbance heterogeneity determines freshwater metacommunity structure. Ecology 2004, 85, 2971–2978. [Google Scholar] [CrossRef]
- Vass, M.; Langenheder, S. The legacy of the past: Effects of historical processes on microbial metacommunities. Aquat. Microb. Ecol. 2017, 79, 13–19. [Google Scholar] [CrossRef]
- Rohal, C.B.; Cranney, C.; Kettenring, K.M. Abiotic and landscape factors constrain restoration outcomes across spatial scales of a widespread invasive plant. Front. Plant Sci. 2019, 10, 481. [Google Scholar] [CrossRef]
- Sharp, S.J.; Angelini, C. The role of landscape composition and disturbance type in mediating salt marsh resilience to feral hog invasion. Biol. Invasions 2019, 21, 2857–2869. [Google Scholar] [CrossRef]
- Biswas, S.R.; Mallik, A.U.; Braithwaite, N.T.; Biswas, P.L. Effects of disturbance type and microhabitat on species and functional diversity relationship in stream-bank plant communities. For. Ecol. Manag. 2019, 432, 812–822. [Google Scholar] [CrossRef]
- Newman, E.A. Disturbance ecology in the anthropocene. Front. Ecol. Evol. 2019, 7, 147. [Google Scholar] [CrossRef]
- Rodewald, A.D.; Yahner, R.H. Influence of landscape composition on avian community structure and associated mechanisms. Ecology 2001, 82, 3493–3504. [Google Scholar] [CrossRef]
- Barba-Escoto, L.; Ponce-Mendoza, A.; García-Romero, A.; Calvillo-Medina, R.P. Plant community strategies responses to recent eruptions of Popocatépetl volcano, Mexico. J. Veg. Sci. 2019, 30, 375–385. [Google Scholar] [CrossRef]
- Shackelford, N.; Standish, R.J.; Lindo, Z.; Starzomski, B.M. The role of landscape connectivity in resistance, resilience, and recovery of multi-trophic microarthropod communities. Ecology 2018, 99, 1164–1172. [Google Scholar] [CrossRef]
- Sferra, C.O.; Hart, J.L.; Howeth, J.G. Habitat age influences metacommunity assembly and species richness in successional pond ecosystems. Ecosphere 2017, 8, e01871. [Google Scholar] [CrossRef]
- Montoya, D. Challenges and directions toward a general theory of ecological recovery dynamics: A metacommunity perspective. One Earth 2021, 4, 1083–1094. [Google Scholar] [CrossRef]
- Vanschoenwinkel, B.; Buschke, F.; Brendonck, L. Disturbance regime alters the impact of dispersal on alpha and beta diversity in a natural metacommunity. Ecology 2013, 94, 2547–2557. [Google Scholar] [CrossRef]
- Uroy, L.; Ernoult, A.; Mony, C. Effect of landscape connectivity on plant communities: A review of response patterns. Landsc. Ecol. 2019, 34, 203–225. [Google Scholar] [CrossRef]
- Zeller, K.A.; Lewison, R.; Fletcher, R.J.; Tulbure, M.G.; Jennings, M.K. Understanding the importance of dynamic landscape connectivity. Land 2020, 9, 303. [Google Scholar] [CrossRef]
- Fahrig, L.; Paloheimo, J. Effect of spatial arrangement of habitat patches on local population size. Ecology 1988, 69, 468–475. [Google Scholar] [CrossRef]
- Mony, C.; Vannier, N.; Brunellière, P.; Biget, M.; Coudouel, S.; Vandenkoornhuyse, P. The influence of host-plant connectivity on fungal assemblages in the root microbiota of Brachypodium pinnatum. Ecology 2020, 101, e02976. [Google Scholar] [CrossRef]
- Rotchés-Ribalta, R.; Winsa, M.; Roberts, S.P.M.; Öckinger, E. Associations between plant and pollinator communities under grassland restoration respond mainly to landscape connectivity. J. Appl. Ecol. 2018, 55, 2822–2833. [Google Scholar] [CrossRef]
- Brudvig, L.A.; Damaschen, E.I.; Tewksbury, J.J.; Haddad, N.M.; Levey, D.J. Landscape connectivity promotes plant biodiversity spillover into non-target habitats. Proc. Natl. Acad. Sci. USA 2009, 106, 9328–9332. [Google Scholar] [CrossRef]
- Joern, A.; Laws, A.N. Ecological mechanisms underlying arthropod species diversity in grasslands. Annu. Rev. Entomol. 2013, 58, 19–36. [Google Scholar] [CrossRef]
- Laurance, W.F. Theory meets reality: How habitat fragmentation research has transcended island biogeographic theory. Biol. Conserv. 2008, 141, 1731–1744. [Google Scholar] [CrossRef]
- Allen, C.; Gonzales, R.; Parrott, L. Modelling the contribution of ephemeral wetlands to landscape connectivity. Ecol. Model. 2020, 419, 108944. [Google Scholar] [CrossRef]
- Bishop-Taylor, R.; Tulbure, M.G.; Broich, M. Surface-water dynamics and land use influence landscape connectivity across a major dryland region. Ecol. Appl. 2017, 27, 1124–1137. [Google Scholar] [CrossRef] [PubMed]
- Bishop-Taylor, R.; Tulbure, M.G.; Broich, M. Impact of hydroclimatic variability on regional-scale landscape connectivity across a dynamic dryland region. Ecol. Indic. 2018, 94, 142–150. [Google Scholar] [CrossRef]
- Mathys, A.; Coops, N.C.; Waring, R.H. Soil water availability effects on the distribution of 20 tree species in western North America. For. Ecol. Manag. 2014, 313, 144–152. [Google Scholar] [CrossRef]
- Andrus, R.A.; Harvey, B.J.; Rodman, K.C.; Hart, S.J.; Veblen, T.T. Moisture availability limits subalpine tree establishment. Ecology 2018, 99, 567–575. [Google Scholar] [CrossRef]
- López, B.C.; Holmgren, M.; Sabaté, S.; Gracia, C.A. Estimating annual rainfall threshold for establishment of tree species in water-limited ecosystems using tree-ring data. J. Arid Environ. 2008, 72, 602–611. [Google Scholar] [CrossRef]
- Del Arroyo, O.G.; Silver, W.L. Disentangling the long-term effects of disturbance on soil biogeochemistry in a wet tropical forest ecosystem. Glob. Change Biol. 2018, 24, 1673–1684. [Google Scholar] [CrossRef]
- Kemp, K.B.; Higuera, P.E.; Morgan, P.; Abatzoglou, J.T. Climate will increasingly determine post-fire tree regeneration success in low-elevation forests, Northern Rockies, USA. Ecosphere 2019, 10, e02568. [Google Scholar] [CrossRef]
- Romero, L.M.; Smith III, T.J.; Fourqurean, J.W. Changes in mass and nutrient content of wood during decomposition in a south Florida mangrove forest. J. Ecol. 2005, 93, 618–631. [Google Scholar] [CrossRef]
- Stevens-Rumann, C.S.; Morgan, P. Tree regeneration following wildfires in the western US: A review. Fire Ecol. 2019, 15, 15. [Google Scholar] [CrossRef]
- Ciccazzo, S.; Esposito, A.; Borruso, L.; Brusetti, L. Microbial communities and primary succession in high altitude mountain environments. Ann. Microbiol. 2016, 66, 43–60. [Google Scholar] [CrossRef]
- Na, X.; Yu, H.; Wang, P.; Zhu, W.; Niu, Y.; Huang, J. Vegetation biomass and soil moisture coregulate bacterial community succession under altered precipitation regimes in a desert steppe in northwestern China. Soil Biol. Biochem. 2019, 136, 107520. [Google Scholar] [CrossRef]
- Walker, L.R.; Chapin, F.S. Physiological controls over seedling growth in primary succession on an Alaskan floodplain. Ecology 1986, 67, 1508–1523. [Google Scholar] [CrossRef]
- Yurkewycz, R.P.; Bishop, J.G.; Crisafulli, C.M.; Harrison, J.A.; Gill, R.A. Gopher mounds decrease nutrient cycling rates and increase adjacent vegetation in volcanic primary succession. Oceologia 2014, 176, 1135–1150. [Google Scholar] [CrossRef] [PubMed]
- Mony, C.; Uroy, L.; Khalfallah, F.; Haddad, N.; Vandenkoornhuyse, P. Landscape connectivity for the invisibles. Ecography 2022, 2022, e06041. [Google Scholar] [CrossRef]
- Wang, B.; Tian, C.; Sun, J. Effects of landscape complexity and stand factors on arthropod communities in poplar forests. Ecol. Evol. 2019, 9, 7143–7156. [Google Scholar] [CrossRef]
- Frazier, A.E.; Kedron, P. Landscape metrics: Past progress and future directions. Curr. Landsc. Ecol. Rep. 2017, 2, 63–72. [Google Scholar] [CrossRef]
- McRae, B.H.; Dickson, B.G.; Keitt, T.H.; Shah, V.B. Using circuit theory to model connectivity in ecology, evolution, and conservation. Ecology 2008, 89, 2712–2724. [Google Scholar] [CrossRef]
- Minor, E.S.; Urban, D.L. A graph-theory framework for evaluating landscape connectivity and conservation planning. Conserv. Biol. 2008, 22, 297–307. [Google Scholar] [CrossRef]
- Baguette, M.; Dyck, H.V. Landscape connectivity and animal behavior: Functional grain as a key determinant for dispersal. Landsc. Ecol. 2007, 22, 1117–1129. [Google Scholar] [CrossRef]
- Diniz, M.F.; Cushman, S.A.; Machado, R.B.; Júnior, P.D.M. Landscape connectivity modeling from the perspective of animal dispersal. Landsc. Ecol. 2020, 35, 41–58. [Google Scholar] [CrossRef]
- Tischendorf, L.; Fahrig, L. On the usage and measurement of landscape connectivity. Oikos 2003, 90, 7–19. [Google Scholar] [CrossRef]
- McRae, B.H.; Beier, P. Circuit theory predicts gene flow in plant and animal populations. Proc. Natl. Acad. Sci. USA 2007, 104, 19885–19890. [Google Scholar] [CrossRef]
- McRae, B.H.; Hall, S.A.; Beier, P.; Theobald, D.M. Where to restore ecological connectivity? Detecting barriers and quantifying restoration benefits. PLoS ONE 2012, 7, e52604. [Google Scholar] [CrossRef]
- Laliberté, J.; St-Laurent, M.-H. Validation of functional connectivity modeling: The Achilles’ heel of landscape connectivity mapping. Landsc. Urban Plan. 2020, 202, 103878. [Google Scholar] [CrossRef]
- Hall, K.R.; Anantharaman, R.; Landau, V.A.; Clark, M.; Dickson, B.G.; Jones, A.; Platt, J.; Edelman, A.; Shah, V.B. Circuitscape in Julia: Empowering dynamic approaches to connectivity assessment. Land 2021, 10, 301. [Google Scholar] [CrossRef]
- Zeller, K.A.; Wattles, D.W.; Destefano, S. Evaluating methods for identifying large mammal road crossing locations: Black bears as a case study. Landsc. Ecol. 2020, 35, 1799–1808. [Google Scholar] [CrossRef]
- Koen, E.L.; Bowman, J.; Garroway, C.J.; Mills, S.C.; Wilson, P.J. Landscape resistance and American marten gene flow. Landsc. Ecol. 2012, 27, 29–43. [Google Scholar] [CrossRef]
- Lozier, J.D.; Strange, J.P.; Koch, J.B. Landscape heterogeneity predicts gene flow in a widespread polymorphic bumble bee, Bombus bifarius (Hymenoptera: Apidae). Conserv. Genet. 2013, 14, 1099–1110. [Google Scholar] [CrossRef]
- Sackett, L.C.; Cross, T.B.; Jones, R.T.; Johnson, W.C.; Ballare, K.; Ray, C.; Collinge, S.K.; Martin, A.P. Connectivity of prairie dog colonies in an altered landscape: Inferences from analysis of microsatellite DNA variation. Conserv. Genet. 2012, 13, 407–418. [Google Scholar] [CrossRef]
- Dickson, B.G.; Albano, C.M.; Anantharaman, R.; Beier, P.; Fargione, J.; Graves, T.A.; Gray, M.E.; Hall, K.R.; Lawler, J.J.; Leonard, P.B.; et al. Circuit-theory applications to connectivity science and conservation. Conserv. Biol. 2018, 33, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Naiman, R.J.; Bechtold, J.S.; Drake, D.C.; Latterell, J.J.; O’Keefe, T.C.; Balian, E.V. Origins, patterns, and importance of heterogeneity in riparian systems. In Ecosystem Function in Heterogeneous Landscapes; Lovett, G.M., Turner, M.G., Jones, C.G., Weathers, K.C., Eds.; Springer: New York, NY, USA, 2006. [Google Scholar]
- Wimp, G.M.; Young, W.P.; Woolbright, S.A.; Martinsen, G.D.; Keim, P.; Whitham, T.G. Conserving plant genetic diversity for dependent animal communities. Ecol. Lett. 2004, 7, 776–780. [Google Scholar] [CrossRef]
- Bangert, R.K.; Turek, R.J.; Rehill, B.; Wimp, G.M.; Schweitzer, J.A.; Allan, G.J.; Bailey, J.K.; Martinsen, G.D.; Keim, P.; Lindroth, R.L.; et al. A genetic similarity rule determines arthropod community structure. Mol. Ecol. 2006, 15, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Lamit, L.J.; Busby, P.E.; Lau, M.K.; Compson, Z.G.; Wojtowicz, T.; Keith, A.R.; Zinkgraf, M.S.; Schweitzer, J.A.; Shuster, S.M.; Gehring, C.A.; et al. Tree genotype mediates covariance among communities from microbes to lichens and arthropods. J. Ecol. 2015, 103, 840–850. [Google Scholar] [CrossRef]
- Cornelissen, T.; Stiling, P. Sex-biased herbivory: A meta-analysis of the effects of gender on plant-herbivore interactions. Oikos 2005, 111, 488–500. [Google Scholar] [CrossRef]
- Dickson, L.L.; Whitham, T.G. Genetically-based plant resistance traits affect arthropods, fungi, and birds. Oecologia 1996, 106, 400–406. [Google Scholar] [CrossRef]
- Donaldson, J.R.; Stevens, M.T.; Barnhill, H.R.; Lindroth, R.L. Age-related shifts in leaf chemistry of clonal aspen (Populus tremuloides). J. Chem. Ecol. 2006, 32, 1415–1429. [Google Scholar] [CrossRef]
- Lau, M.K.; Arnold, A.E.; Johnson, N.C. Factors influencing communities of foliar fungal endophytes in riparian woody plants. Fungal Ecol. 2013, 6, 365–378. [Google Scholar] [CrossRef]
- Durben, R.M.; Walker, F.M.; Holeski, L.; Keith, A.R.; Kovacs, Z.; Hurteau, S.R.; Lindroth, R.L.; Shuster, S.M.; Whitham, T.G. Beavers, bugs and chemistry: A mammalian herbivore changes chemistry composition and arthropod communities in foundation tree species. Forests 2021, 12, 877. [Google Scholar] [CrossRef]
- Martinsen, G.D.; Floate, K.D.; Waltz, A.M.; Wimp, G.M.; Whitham, T.G. Positive interactions between leafrollers and other arthropods enhance biodiversity on hybrid cottonwoods. Oecologia 2000, 123, 82–89. [Google Scholar] [CrossRef]
- Newcombe, G.; Fraser, S.J.; Ridout, M.; Busby, P.E. Leaf endophytes of Populus trichocarpa act as pathogens of neighboring plant species. Front. Microbiol. 2020, 11, 573056. [Google Scholar] [CrossRef]
- Wolfe, E.R.; Younginger, B.S.; LeRoy, C.J. Fungal endophyte-infected leaf litter alters in-stream microbial communities and negatively influences aquatic fungal sporulation. Oikos 2018, 128, 405–415. [Google Scholar] [CrossRef]
- Badri, D.V.; Zolla, G.; Bakker, M.G.; Manter, D.K.; Vivanco, J.M. Potential impact of soil microbiomes on the leaf metabolome and on herbivore feeding behavior. New Phytol. 2013, 198, 264–273. [Google Scholar] [CrossRef]
- Fargen, C.; Emery, S.M.; Carreiro, M.M. Influence of Lonicera maackii invasion on leaf litter decomposition and macroinvertebrate communities in an urban stream. Nat. Areas J. 2015, 35, 392–403. [Google Scholar] [CrossRef]
- Martinsen, G.D.; Driebe, E.M.; Whitham, T.G. Indirect interactions mediated by changing plant chemistry: Beaver browsing benefits beetles. Ecology 1998, 79, 192–200. [Google Scholar] [CrossRef]
- Chang, C.C.; HilleRisLambers, J. Trait and phylogenetic patterns reveal deterministic community assembly mechanisms on Mount St. Helens. Plant Ecol. 2019, 220, 675–698. [Google Scholar] [CrossRef]
- Jouval, F.; Bigot, L.; Bureau, S.; Quod, J.-P.; Penin, L.; Adjeroud, M. Diversity, structure and demography of coral assemblages on underwater lava flows of different ages at Reunion Island and implications for ecological succession hypotheses. Sci. Rep. 2020, 10, 20821. [Google Scholar] [CrossRef]
- Lopes, L.F.; Oliveira, S.C.; Neto, C.; Zêzere, J.L. Vegetation evolution by ecological succession as a potential bioindicator of landslides relative age in Southwestern Mediterranean region. Nat. Hazards 2020, 103, 599–622. [Google Scholar] [CrossRef]
- Végh, L.; Tsuyuzaki, S. Differences in canopy and understorey diversities after the eruptions of Mount Usu, northern Japan—Impacts of early forest management. For. Ecol. Manag. 2022, 510, 120106. [Google Scholar] [CrossRef]
- Che-Castaldo, C.; Crisafulli, C.M.; Bishop, J.G.; Fagan, W.F. What causes female bias in the secondary sex ratios of the dioecious woody shrub Salix sitchensis colonizing a primary successional landscape? Am. J. Bot. 2015, 102, 1309–1322. [Google Scholar] [CrossRef]
- LeRoy, C.J.; Ramstack Hobbs, J.M.; Claeson, S.M.; Moffet, J.; Garthwaite, I.J.; Criss, N.; Walker, L. Plant sex influences aquatic–terrestrial interactions. Ecosphere 2020, 11, e02994. [Google Scholar] [CrossRef]
- Che-Castaldo, C.; Crisafulli, C.M.; Bishop, J.G.; Zipkin, E.F.; Fagan, B.F. Disentangling herbivore impacts in primary succession by refocusing the plant stress and vigor hypotheses on phenology. Ecol. Monogr. 2019, 89, e01389. [Google Scholar] [CrossRef]
- Choudhury, D. Herbivore induced changes in leaf-litter resource quality: A neglected aspect of herbivory in ecosystem nutrient dynamics. Oikos 1988, 51, 389–393. [Google Scholar] [CrossRef]
- Xiao, L.; Carrillo, J.; Siemann, E.; Ding, J. Herbivore-specific induction of indirect and direct defensive responses in leaves and roots. AoB Plants 2019, 11, plz003. [Google Scholar] [CrossRef]
- Ramstack Hobbs, J.M.; Garthwaite, I.J.; Lancaster, L.; Moffett-Dobbs, J.A.; Johnson, K.; Criss, N.; McConathy, V.; James, C.A.; Gipe, A.; Claeson, S.M.; et al. The influence of weevil herbivory on leaf litter chemistry in dioecious willows. Ecol. Evol. 2022, 12, e9626. [Google Scholar] [CrossRef]
- Aide, T.M.; Londoño, E.C. The effects of rapid leaf expansion on the growth and survivorship of a Lepidopteran herbivore. Oikos 1989, 55, 66–70. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M. Do small leaves expand faster than large leaves, and do shorter expansion times reduce herbivore damage? Oikos 2003, 90, 517–524. [Google Scholar] [CrossRef]
- Ewers, R.M.; Didham, R.K. Confounding factors in the detection of species responses to habitat fragmentation. Biol. Rev. Camb. Philos. Soc. 2006, 81, 117–142. [Google Scholar] [CrossRef]
- Lipman, P.W.; Mullineaux, D.R. The 1980 eruptions of Mount St. Helens, Washington. In USGS Numbered Series; Professional Paper 1250; U.S. Government Printing Office: Washington, DC, USA, 1981. [Google Scholar] [CrossRef]
- Swanson, F.J.; Major, J.J. Physical events, environments, and geological—Ecological interactions at Mount St. Helens: March 1980–2004. In Ecological Responses to the 1980 Eruption of Mount St. Helens; Dale, V.H., Swanson, F.J., Crisafulli, C.M., Eds.; Springer: New York, NY, USA, 2005; pp. 27–44. [Google Scholar]
- Claeson, S.M.; LeRoy, C.J.; Finn, D.S.; Stancheva, R.H.; Wolfe, E.R. Variation in riparian and stream assemblages across the primary succession landscape of Mount St. Helens, U.S.A. Freshw. Biol. 2021, 66, 1002–1017. [Google Scholar] [CrossRef]
- Esri Inc. ArcGIS Pro (Version 3.0.0). 2022. Available online: https://www.esri.com/en-us/arcgis/products/arcgis-pro/overview (accessed on 12 August 2022).
- Fisher, M.J. The morphology and anatomy of the flowers of the Salicaceae I. Am. J. Bot. 1928, 15, 307–326. [Google Scholar] [CrossRef]
- Porter, L.J.; Foo, L.Y.; Furneaux, R.H. Isolation of three naturally occurring O-β-glucopyranosides of procyanidin polymers. Phytochemistry 1985, 24, 567–569. [Google Scholar] [CrossRef]
- Hagerman, A.E.; Butler, L.G. Choosing appropriate methods and standards for assaying tannin. J. Chem. Ecol. 1989, 15, 1795–1810. [Google Scholar] [CrossRef]
- Feinstein, L.M.; Blackwood, C.B. Taxa-area relationship and neutral dynamics influence the diversity of fungal communities on senesced tree leaves. Environ. Microbiol. 2012, 14, 1488–1499. [Google Scholar] [CrossRef]
- Tielens, E.K.; Gruner, D.S. Intraspecific variation in host plant traits mediates taxonomic and functional composition of local insect herbivore communities. Ecol. Entomol. 2020, 45, 1382–1395. [Google Scholar] [CrossRef]
- Abramoff, M.D.; Magalhaes, P.J.; Ram, S.J. Image Processing with ImageJ. Biophoton. Int. 2004, 11, 36–42. [Google Scholar]
- U.S. Geological Survey. 3D Elevation Program 1-Meter Resolution Digital Elevation Model (Published 20200606). 2019. Available online: https://www.usgs.gov/the-national-map-data-delivery (accessed on 15 August 2022).
- R Core Team. R Version 4.2.0: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 7 September 2022).
- Pinheiro, J.; Bates, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-160. 2022. Available online: https://CRAN.R-project.org/package=nlme (accessed on 10 September 2022).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 25 September 2022).
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of Southern Wisconsin. Ecol. Mongr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Akaike, H. Factor analysis and AIC. Psychometrika 1987, 52, 317–332. [Google Scholar] [CrossRef]
- Lenth, R.V.; Buerkner, P.; Giné-Vázquez, I.; Herve, M.; Jung, M.; Love, J.; Miguez, F.; Riebl, H.; Singmann, H. emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.8.3. 2022. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 25 September 2022).
- Lenth, R.V. Least-squares means: The R package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef]
- Martinez Arbizu, P. pairwiseAdonis: Pairwise Multilevel Comparison Using Adonis. R Package Version 0.4. 2020. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 25 September 2022).
- Aggemyr, E.; Auffret, A.G.; Jädergård, L.; Cousins, S.A.O. Species richness and composition differ in response to landscape and biogeography. Landsc. Ecol. 2018, 33, 2273–2284. [Google Scholar] [CrossRef]
- García, D.; Martínez, D. Species richness matters for the quality of ecosystem services: A test using seed dispersal by frugivorous birds. Proc. Biol. Sci. 2012, 279, 3106–3113. [Google Scholar] [CrossRef]
- Harrison, J.G.; Philbin, C.S.; Gompert, Z.; Forister, G.W.; Hernandez-Espinosa, L.; Sullivan, B.W.; Wallace, I.S.; Beltran, L.; Dodson, C.D.; Francis, J.S.; et al. Deconstruction of a plant-arthropod community reveals influential plant traits with nonlinear effects on arthropod assemblages. Funct. Ecol. 2018, 32, 1317–1328. [Google Scholar] [CrossRef]
- Marks, J.C.; Haden, G.A.; Harrop, B.L.; Reese, E.G.; Keams, J.L.; Watwood, M.E.; Whitham, T.G. Genetic and environmental controls of microbial communities on leaf litter in streams. Freshw. Biol. 2009, 54, 2616–2627. [Google Scholar] [CrossRef]
- Rinkes, Z.L.; DeForest, J.L.; Grandy, A.S.; Moorhead, D.L.; Weintraub, M.N. Interactions between leaf litter quality, particle size, and microbial community during the earliest stage of decay. Biogeochemistry 2014, 117, 153–168. [Google Scholar] [CrossRef]
- Fagan, W.F.; Denno, R.F. Stoichiometry of actual vs. potential predator–prey interactions: Insights into nitrogen limitation for arthropod predators. Ecol. Lett. 2004, 7, 876–883. [Google Scholar] [CrossRef]
- Wiesenborn, W.D. Biomasses of arthropod taxa differentially increase on nitrogen-fertilized willows and cottonwoods. Restor. Ecol. 2011, 19, 323–332. [Google Scholar] [CrossRef]
- Popescu, C.; Oprina-Pavelescu, M.; Dinu, V.; Cazacu, C.; Burdon, F.J.; Forio, M.A.E.; Kupilas, B.; Friberg, N.; Goethals, P.; McKie, B.G.; et al. Riparian vegetation structure influences terrestrial invertebrate communities in an agricultural landscape. Water 2021, 13, 188. [Google Scholar] [CrossRef]
- Rocha-Ortega, M.; Rodríguez, P.; Córdoba-Aguilar, A. Spatial and temporal effects of land use change as potential drivers of odonate community composition but not species richness. Biodivers. Conserv. 2019, 28, 451–466. [Google Scholar] [CrossRef]
- Helfer, S. Rust fungi and global change. New Phytol. 2013, 201, 770–780. [Google Scholar] [CrossRef]
- David, A.S.; Glueckert, J.S.; Enloe, S.F.; Cortes, A.C.; Abdel-Kader, A.A.; Lake, E.C. Eriophyid mite Floracarus perrepae readily colonizes recovering invasive vine Lygodium microphyllum following herbicide treatment. BioControl 2021, 66, 573–584. [Google Scholar] [CrossRef]
- Mukwevho, L.; Olckers, T.; Simelane, D.O. Establishment, dispersal and impact of the flower-galling mite Aceria lantanae (Acari: Trombidiformes: Eriophyidae) on Lantana camara (Verbenaceae) in South Africa. Biol. Control 2017, 107, 33–40. [Google Scholar] [CrossRef]
- Peng, M.-H.; Hung, Y.-C.; Liu, K.-L.; Neoh, K.-B. Landscape configuration and habitat complexity shape arthropod assemblage in urban parks. Sci. Rep. 2020, 10, 16043. [Google Scholar] [CrossRef]
- Gish, M.; Inbar, M. Standing on the shoulders of giants: Young aphids piggyback on adults when searching for a host plant. Front. Zool. 2018, 15, 49. [Google Scholar] [CrossRef]
- Wolfe, E.R.; Dove, R.; Webster, C.; Ballhorn, D.J. Culturable fungal endophyte communities of primary successional plants on Mount St. Helens, WA, USA. BMC Ecol. Evol. 2022, 22, 18. [Google Scholar] [CrossRef]
- Spanowicz, A.G.; Jaeger, J.A.G. Measuring landscape connectivity: On the importance of within-patch connectivity. Landsc. Ecol. 2019, 34, 2261–2278. [Google Scholar] [CrossRef]
- Allison, S.D.; Martiny, J.B.H. Resistance, resilience, and redundancy in microbial communities. Proc. Natl. Acad. Sci. USA 2008, 105, 11512–11519. [Google Scholar] [CrossRef]
- Borkenhagen, A.; Cooper, D.J. Tolerance of fen mosses to submergence, and the influence on moss community composition and ecosystem resilience. J. Veg. Sci. 2018, 29, 127–135. [Google Scholar] [CrossRef]
- Lavorel, S.; Garnier, E. Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail. Funct. Ecol. 2002, 16, 545–556. [Google Scholar] [CrossRef]
- Van der Plas, F. Biodiversity and ecosystem functioning in naturally assembled communities. Biol. Rev. 2019, 94, 1220–1245. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Portsmouth, A.; Tena, D.; Tobias, M.; Matesanz, S.; Valladares, F. Do we underestimate the importance of leaf size in plant economics? Disproportional scaling of support costs within the spectrum of leaf physiognomy. Ann. Bot. 2007, 100, 283–303. [Google Scholar] [CrossRef]
- Steppe, K.; Niinemets, Ü.; Teskey, R.O. Tree size- and age-related changes in leaf physiology and their influence on carbon gain. In Size- and Age-Related Changes in Tree Structure and Function. Tree Physiology; Meinzer, F., Lachenbruch, B., Dawson, T., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 4, pp. 235–253. [Google Scholar] [CrossRef]
- Májeková, M.; Hájek, T.; Albert, A.J.; de Bello, F.; Doležal, J.; Götzenberger, L.; Janeček, S.; Lepš, J.; Liancourt, P.; Mudrák, O. Weak coordination between leaf drought tolerance and proxy traits in herbaceous plants. Funct. Ecol. 2021, 35, 1299–1311. [Google Scholar] [CrossRef]
- Schreiber, S.G.; Hacke, U.G.; Chamberland, S.; Lowe, C.W.; Kamelchuk, D.; Bräutigam, K.; Campbell, M.M.; Thomas, B.R. Leaf size serves as a proxy for xylem vulnerability to cavitation in plantation trees. Plant Cell Environ. 2015, 39, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Moraga, A.D.; Martin, A.E.; Fahrig, L. The scale of effect of landscape context varies with the species’ response variable measured. Landsc. Ecol. 2019, 34, 703–715. [Google Scholar] [CrossRef]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, J.M.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Truchy, A.; Angeler, D.G.; Sponseller, R.A.; Johnson, R.K.; McKie, B.G. Chapter Two—Linking biodiversity, ecosystem functioning and services, and ecological resilience: Towards an integrative framework for improved management. Adv. Ecol. Res. 2015, 53, 55–96. [Google Scholar] [CrossRef]
- Oliver, T.H.; Heard, M.S.; Isaac, N.J.B.; Roy, D.B.; Procter, D.; Eigenbrod, F.; Freckleton, R.; Hector, A.; Orme, C.D.L.; Petchey, O.L.; et al. Biodiversity and resilience of ecosystem functions. Trends Ecol. Evol. 2015, 30, 673–684. [Google Scholar] [CrossRef]
- Friedman, J.M.; Lee, V.J. Extreme floods, channel change, and riparian forests along ephemeral streams. Ecol. Monogr. 2002, 72, 409–425. [Google Scholar] [CrossRef]
- Bloss, D.A.; Brotherson, J.D. Vegetation response to a moisture gradient on an ephemeral stream in central Arizona. Great Basin Nat. 1979, 39, 161–176. [Google Scholar]
- Mohammad, M.K.; Al-Rammahi, H.M.; Cogoni, D.; Fenu, G. Conservation need for a plant species with extremely small populations linked to ephemeral streams in adverse desert environments. Water 2022, 14, 2638. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | df | σ | t | p | |
|---|---|---|---|---|---|
| Landscape Model (n = 335) | |||||
| Connectivity | 1 | −0.718 | 0.263 | −2.731 | 0.007 |
| Stream-Clear | 1 | −0.188 | 0.394 | −0.477 | 0.633 |
| Stream-Forsyth | 1 | 0.423 | 0.301 | 1.406 | 0.161 |
| Stream-Geo-W | 1 | −0.808 | 0.264 | −3.059 | 0.002 |
| Stream-Goose | 1 | −0.488 | 0.304 | −1.608 | 0.109 |
| Stream-Redrock | 1 | −0.936 | 0.294 | −3.187 | 0.002 |
| Stream-Willow | 1 | −0.025 | 0.293 | −0.086 | 0.931 |
| Residuals | 327 | 0.114 | 1.065 | - | - |
| Litter Chemistry Model (n = 31) | |||||
| %N | 1 | 1.492 | 0.657 | 2.269 | 0.031 |
| %CT | 1 | 0.150 | 0.053 | 2.842 | 0.008 |
| Weevils | 1 | 0.985 | 0.996 | 0.988 | 0.332 |
| Residuals | 28 | 0.095 | 0.867 | - | - |
| Leaf Area Model (n = 109) | |||||
| SLA | 1 | >−0.001 | <0.001 | −5.417 | <0.001 |
| Willow Sex | 1 | −0.289 | 0.186 | −1.555 | 0.123 |
| Residuals | 107 | 0.054 | 0.969 | - | - |
| Factor | df | SS | R2 | F | p |
|---|---|---|---|---|---|
| Landscape Model (n = 335) | |||||
| Connectivity | 1 | 0.361 | 0.013 | 5.298 | 0.003 |
| Willow Sex | 1 | 0.386 | 0.014 | 5.670 | 0.003 |
| Stream | 6 | 5.600 | 0.196 | 13.716 | 0.001 |
| Residual | 326 | 22.183 | 0.778 | - | - |
| Litter Chemistry Model (n = 31) | |||||
| %N | 1 | 0.097 | 0.056 | 1.748 | 0.166 |
| %CT | 1 | 0.080 | 0.046 | 1.444 | 0.237 |
| Residual | 28 | 1.559 | 0.898 | - | - |
| Leaf Area Model (n = 109) | |||||
| SLA | 1 | 0.353 | 0.041 | 4.575 | 0.015 |
| Willow Sex | 1 | 0.064 | 0.007 | 0.825 | 0.501 |
| Residual | 107 | 8.179 | 0.952 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minsavage-Davis, C.D.; Garthwaite, I.J.; Fisher, M.D.; Leigh, A.; Ramstack Hobbs, J.M.; Claeson, S.M.; Wimp, G.M.; LeRoy, C.J. Spatial Habitat Structure Assembles Willow-Dependent Communities across the Primary Successional Watersheds of Mount St. Helens, USA. Forests 2023, 14, 322. https://doi.org/10.3390/f14020322
Minsavage-Davis CD, Garthwaite IJ, Fisher MD, Leigh A, Ramstack Hobbs JM, Claeson SM, Wimp GM, LeRoy CJ. Spatial Habitat Structure Assembles Willow-Dependent Communities across the Primary Successional Watersheds of Mount St. Helens, USA. Forests. 2023; 14(2):322. https://doi.org/10.3390/f14020322
Chicago/Turabian StyleMinsavage-Davis, Charli D., Iris J. Garthwaite, Marisa D. Fisher, Addison Leigh, Joy M. Ramstack Hobbs, Shannon M. Claeson, Gina M. Wimp, and Carri J. LeRoy. 2023. "Spatial Habitat Structure Assembles Willow-Dependent Communities across the Primary Successional Watersheds of Mount St. Helens, USA" Forests 14, no. 2: 322. https://doi.org/10.3390/f14020322
APA StyleMinsavage-Davis, C. D., Garthwaite, I. J., Fisher, M. D., Leigh, A., Ramstack Hobbs, J. M., Claeson, S. M., Wimp, G. M., & LeRoy, C. J. (2023). Spatial Habitat Structure Assembles Willow-Dependent Communities across the Primary Successional Watersheds of Mount St. Helens, USA. Forests, 14(2), 322. https://doi.org/10.3390/f14020322