
Changes in Soil Properties and Enzyme Stoichiometry in Three Different Forest Types Changed to Tea Plantations

Abstract
:1. Introduction
2. Methods
2.1. Study Area
2.2. Site Selection and Sampling
2.3. Soil Parameter Determination
2.4. Calculation of Enzyme Stoichiometry Ratio
| C/N enzyme activity ratio (EC/N) = ln(BG)/ln(NAG) C/P enzyme activity ratio (EC/P) = ln(BG)/ln(ACP) N/P enzyme activity ratio (EN/P) = ln(NAG)/ln(ACP) |
| VL = [(ln(BG)/ln(NAG))2 + (ln(BG)/ln(ACP))2]1/2 VA = Degrees{ATAN2[(ln(BG)/ln(ACP)), (ln(BG)/ln(NAG))]} |
2.5. Data Processing and Analysis
3. Results
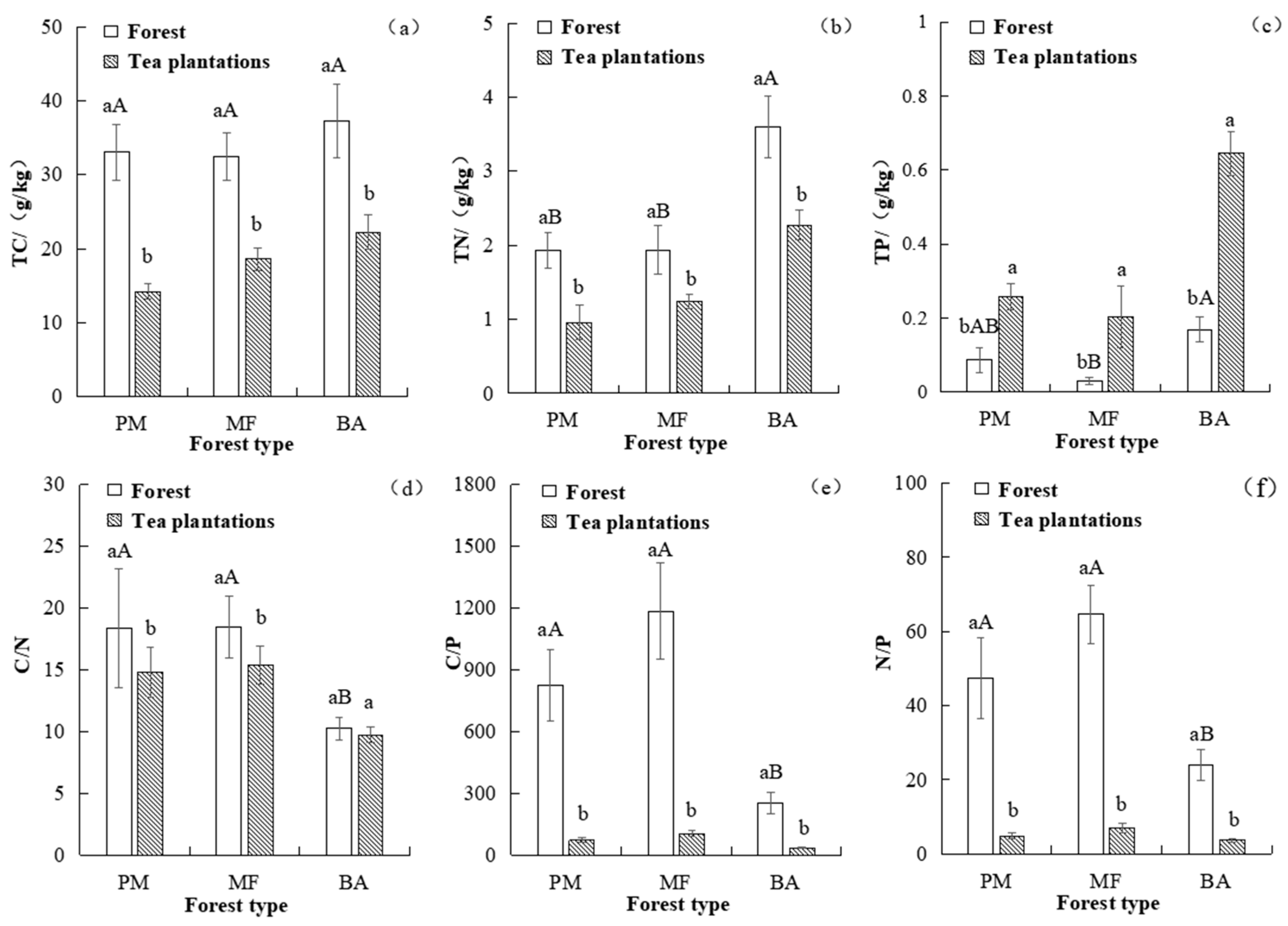
3.1. Effects of Converting Three Forest Types into Tea Plantations on Soil Carbon, Nitrogen, Phosphorus, and Their Stoichiometry
3.2. Impact of Converting Three Forest Types into Tea Plantations on Soil Enzyme Activity and Enzyme Metrics
3.2.1. Impact of Converting Three Forest Types into Tea Plantations on Soil Enzyme Activity
3.2.2. Impact of Converting Three Forest Types into Tea Plantations on Soil Enzyme Metrics
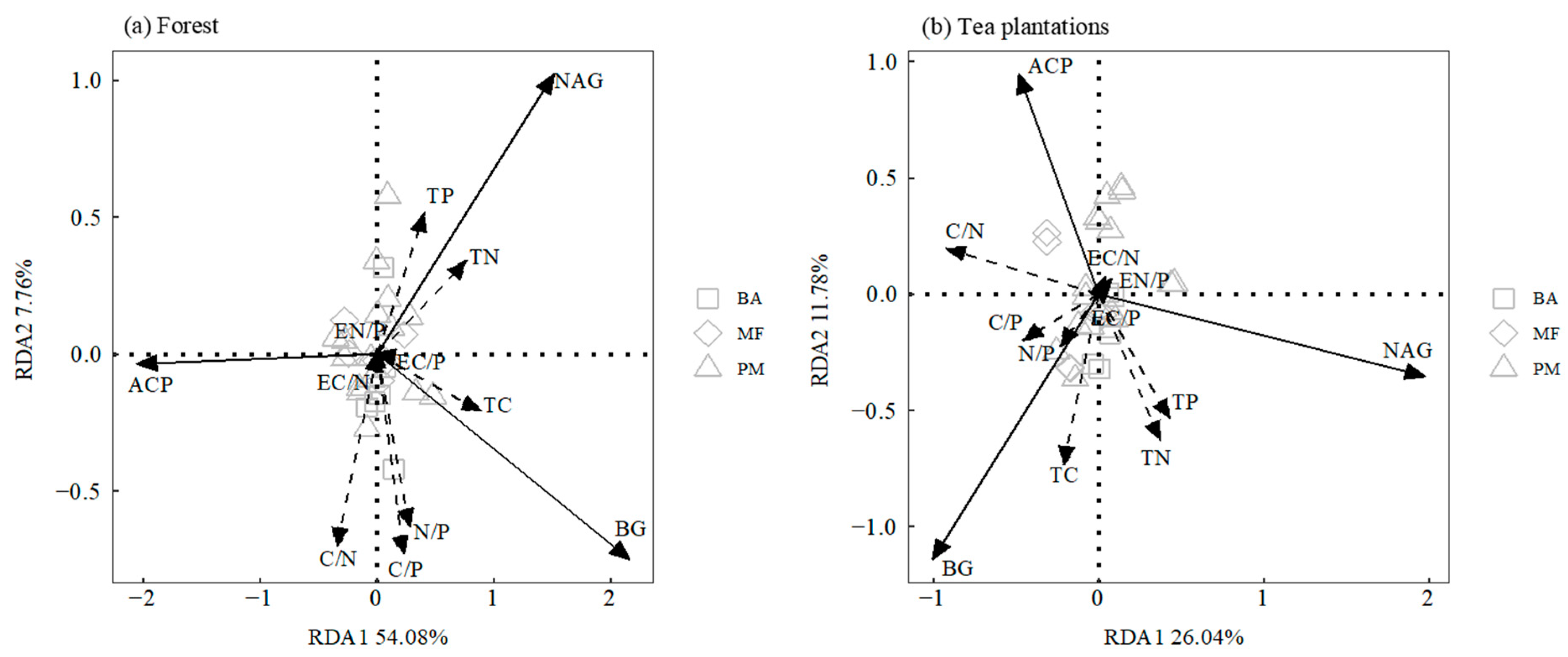
3.3. Relationship between Soil Enzyme Activity and Soil Nutrients
4. Discussion
4.1. The Impact of Forest-to-Tea Conversion on Soil Carbon, Nitrogen, Phosphorus, and Their Stoichiometry
4.2. The Effects of Converting Forest Land to Tea Plantation on Soil Enzyme Activity and Its Stoichiometric Ratios
4.3. Driving Factors of Changes in Soil Enzyme Activity during the Conversion from Forest Land to Tea Plantation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, S.Y. Carbon Balance of Tea Plantation Ecosystem in China. Ph.D Thesis, Zhejiang University, Hangzhou, China, 2010. [Google Scholar]
- Li, S.; Wu, X.; Xue, H.; Gu, B.; Cheng, H.; Zeng, J.; Peng, C.; Ge, Y.; Chang, J. Quantifying carbon storage for tea plantations in China. Agric. Ecosyst. Environ. 2011, 141, 390–398. [Google Scholar] [CrossRef]
- Lv, J.L.; Yan, M.J.; Song, B.L.; Guan, J.H.; Shi, W.Y.; Du, S. Ecological stoichiometry characteristics of soil carbon, nitrogen, and phosphorus in an oak forest and a black locust plantation in the Loess hilly region. Acta Ecol. Sin. 2017, 37, 3385–3393. (In Chinese) [Google Scholar]
- Yang, Y.H.; Fang, J.Y.; Guo, D.L.; Ji, C.J.; Ma, W.H. Vertical Patterns of Soil Carbon, Nitrogen and Carbon: Nitrogen Stoichiometry in Tibetan Grasslands. Biogeosci. Discuss. 2010, 7, 1–24. [Google Scholar] [CrossRef]
- Zhang, X.R.; Zhang, W.Q.; Wang, H.R.; Lu, X.X.; Chun, F.; Sai, X. Response of soil ecological stoichiometric characteristics to grazing intensity in Stipa kirschnii grassland. Acta Ecol. Sin. 2021, 41, 5309–5316. (In Chinese) [Google Scholar]
- Don, A.; Schumacher, J.; Freibauer, A. Impact of tropical land-use change on soil organic carbon stocks: A metaanalysis. Glob. Chang. Biol. 2011, 17, 1658–1670. [Google Scholar] [CrossRef]
- Guo, L.B.; Gifford, R.M. Soil carbon stocks and land use change: A meta analysis. Glob. Chang. Biol. 2002, 8, 345–360. [Google Scholar] [CrossRef]
- Liu, X.; Ma, J.; Ma, Z.W.; Li, L.H. Soil Nutrient Contents and Stoichiometry as Affected by Land-Use in an Agro-Pastoral Region of Northwest China. Catena 2017, 150, 146–153. [Google Scholar] [CrossRef]
- Yang, W.; Zhou, J.G.; Wang, M.H.; Han, Z.; Zhang, M.Y.; Li, Y.Y.; Lv, D.Q.; Wu, J.S. Spatial heterogeneity of soil carbon, nitrogen, and phosphorus ecological stoichiometry characteristics in a subtropical hilly watershed. Acta Pedol. Sin. 2015, 52, 1336–1344. (In Chinese) [Google Scholar]
- Alvarez, C.; Alvarez, C.R.; Costantini, A.; Basanta, M. Carbon and Nitrogen Sequestration in Soils under Different Management in the Semi-Arid Pampa (Argentina). Soil Tillage Res. 2014, 142, 25–31. [Google Scholar] [CrossRef]
- Sun, C.L.; Wang, Y.W.; Wang, C.J.; Li, Q.J.; Wu, Z.H.; Yuan, D.S.; Zhang, J.L. Effect of land used conversion on soil extracellular enzyme activity and its stoichiometry characteristics in karst mountainous areas. Acta Ecol. Sin. 2021, 41, 4140–4149. (In Chinese) [Google Scholar]
- Wang, C.; Dong, Y.Q.; Lu, Y.; Li, B.; Tang, X.; Qiu, J.C.; Hu, J.S. Effects of Transforming Forest Land into Terraced Land on the Characteristics of Soil Carbon, Nitrogen, Phosphorus and Their Stoichiometry in North Guangdong, China. Chin. J. Appl. Ecol. 2021, 32, 2440–2448. (In Chinese) [Google Scholar]
- Chen, L.; Gong, J.; Fu, B.; Huang, Z.; Huang, Y.; Gui, L. Effect of Land Use Conversion on Soil Organic Carbon Sequestration in the Loess Hilly Area, Loess Plateau of China. Ecol. Res. 2007, 22, 641–648. [Google Scholar] [CrossRef]
- Tian, H.; Chen, G.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Zhao, T.; Yan, H.; Jiang, Y.L.; Huang, Y.-M.; An, S. Effects of Vegetation Types on Soil Microbial Biomass C, N, P on the Loess Hilly Area. Acta Ecol. Sin. 2013, 33, 5615–5622. (In Chinese) [Google Scholar] [CrossRef]
- Huang, Y.R.; Gao, W.; Huang, S.D.; Lin, J.; Tan, F.L.; You, H.M.; Yang, L. Ecostoichiometric characteristics of carbon, nitrogen and phosphorus in Fujian evergreen broad-leaved forest. Acta Ecol. Sin. 2021, 41, 1991–2000. (In Chinese) [Google Scholar]
- Li, Z.B.; Zhou, B.; Ma, T.T.; Ke, H.C.; Xu, G.C.; Zhang, W.; Yu, K.X.; Cheng, Y.T. Effects of Ecological Management on Characteristics of Soil Carbon, Nitrogen, Phosphorus and Their Stoichiometry in Loess Hilly Region, China. Soil Water Conserv. 2017, 31, 313–317. (In Chinese) [Google Scholar]
- Wang, Z.; Zheng, F. Impact of vegetation succession on leaf-litter-soil C:N:P stoichiometry and their intrinsic relationship in the ziwuling area of China’s Loess Plateau. J. For. Res. 2021, 32, 697–711. [Google Scholar] [CrossRef]
- Guan, S.; Zhang, D.; Zhang, Z. Soil Enzymes and Their Research Methods; Agricultural Press: Beijing, China, 1986. [Google Scholar]
- Moreau, D.; Bardgett, R.D.; Finlay, R.D.; Jones, D.L.; Philippot, L. A Plant Perspective on Nitrogen Cycling in the Rhizosphere. Funct. Ecol. 2019, 33, 540–552. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Belnap, J.; Findlay, S.G.; Shah, J.J.F.; Hill, B.H.; Kuehn, K.A.; Kuske, C.R.; Litvak, M.E.; Martinez, N.G.; Moorhead, D.L.; et al. Extracellular enzyme kinetics scale with resource availability. Biogeochemistry 2014, 121, 287–304. [Google Scholar] [CrossRef]
- Bengtson, P.; Bengtsson, G. Rapid Turnover of DOC in Temperate forests accounts for increased CO2 production at elevated temperatures. Ecol. Lett. 2007, 10, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Allison, S.D. Cheaters, Diffusion and nutrients constrain decomposition by microbial enzymes in spatially structured environments. Ecol. Lett. 2005, 8, 626–635. [Google Scholar] [CrossRef]
- Kumar, S.; Nand, S.; Dubey, D.; Pratap, B.; Dutta, V. Variation in extracellular enzyme activities and their influence on the performance of surface-flow constructed wetland microcosms (CWMs). Chemosphere 2020, 251, 126377. [Google Scholar] [CrossRef] [PubMed]
- Merino, C.; Godoy, R.; Matus, F. Soil Enzymes and biological activity at different levels of organic matter stability. J. Soil Sci. Plant Nutr. 2016, 16, 14–30. [Google Scholar]
- Zhang, X.X.; Yang, L.M.; Chen, Z.; Li, Y.Q.; Lin, Y.Y.; Zheng, X.Z.; Chu, H.Y.; Yang, Y.S. Patterns of ecoenzymatic stoichiometry on types of forest soils form different parent materials in subtropical areas. Acta Ecol. Sin. 2018, 38, 5828–5836. (In Chinese) [Google Scholar]
- Feng, C.; Ma, Y.; Jin, X.; Wang, Z.; Ma, Y.; Fu, S.; Chen, H.Y.H. Soil enzyme activities increase following restoration of degraded subtropical forests. Geoderma 2019, 351, 180–187. [Google Scholar] [CrossRef]
- Hill, B.H.; Elonen, C.M.; Jicha, T.M.; Cotter, A.M.; Trebitz, A.S.; Danz, N.P. Sediment microbial enzyme activity as an indicator of nutrient limitation in great lakes coastal wetlands. Freshw. Biol. 2006, 51, 1670–1683. [Google Scholar] [CrossRef]
- Schimel, J.P.; Weintraub, M.N. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model. Soil Biol. Biochem. 2003, 35, 549–563. [Google Scholar] [CrossRef]
- Hill, B.H.; Elonen, C.M.; Jicha, T.M.; Bolgrien, D.W.; Moffett, M.F. Sediment microbial enzyme activity as an indicator of nutrient limitation in the great rivers of the Upper Mississippi River basin. Biogeochemistry 2010, 97, 195–209. [Google Scholar] [CrossRef]
- Hill, B.H.; Elonen, C.M.; Seifert, L.R.; May, A.A.; Tarquinio, E. Microbial enzyme stoichiometry and nutrient limitation in US streams and rivers. Ecol. Indic. 2012, 18, 540–551. [Google Scholar] [CrossRef]
- Insabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of Soil Enzyme Activity at Global Scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Hill, B.H.; Shah, J.J.F. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature 2009, 462, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.X.; Fang, L.C.; Guo, X.B.; Wang, X.; Zhang, Y.J.; Li, P.F.; Zhang, X.C. Ecoenzymatic stoichiometry and microbial nutrient limitation in rhizosphere soil in the arid area of the northern Loess Plateau, China. Soil Biol. Biochem. 2018, 116, 11–21. [Google Scholar] [CrossRef]
- Xu, Z.; Yu, G.; Zhang, X.; He, N.; Wang, Q.; Wang, S.; Wang, R.; Zhao, N.; Jia, Y.; Wang, C. Soil enzyme activity and stoichiometry in forest ecosystems along the North-South Transect in eastern China (NSTEC). Soil Biol. Biochem. 2017, 104, 152–163. [Google Scholar] [CrossRef]
- Chen, Y.Z.; Wang, F.; Wu, Z.D.; Liu, X.Y.; Liu, J. Effect of converting forestland to tea plantation on physiochemical properties of soil. Acta Tea Sin. 2018, 59, 205–210. (In Chinese) [Google Scholar]
- Xiao, T.X. Wuyi Tea Classics; Science Press: Beijing, China, 2008. (In Chinese) [Google Scholar]
- You, W.; Ji, Z.; Wu, L.; Deng, X.; Huang, D.; Chen, B.; Yu, J.; He, D. Modeling changes in land use patterns and ecosystem services to explore a potential solution for meeting the management needs of a heritage site at the landscape level. Ecol. Indic. 2017, 73, 68–78. [Google Scholar] [CrossRef]
- Xi, D.; Yu, Z.P.; Xiong, Y.; Liu, X.Y.; Liu, J. Altitudinal changes of soil organic carbon fractions of evergreen broadleaved forests in Guanshan Mountain, Jiangxi, China. Chin. J. Appl. Ecol. 2020, 31, 3349–3356. (In Chinese) [Google Scholar]
- Liu, M.H.; Xie, T.T.; Li, R.; Li, L.J.; Li, C.X. Carbon, nitrogen, and phosphorus ecological stoichiometric characteristics between Taxodium ascendens and soil in the water-level fluctuation zone of the Three Gorges Reservoir region. Acta Ecol. Sin. 2020, 40, 3072–3084. (In Chinese) [Google Scholar]
- Zeng, Q.X.; Zhang, Q.F.; Lin, K.M.; Zhou, J.C.; Yuan, X.C.; Mei, K.C.; Wu, Y.; Cui, J.Y.; Xu, J.G.; Chen, Y.M. Enzyme stoichiometry evidence revealed that five years nitrogen addition exacerbated the carbon and phosphorus limitation of soil microorganisms in a Phyllostachys pubescens forest. Chin. J. Appl. Ecol. 2021, 32, 521–528. (In Chinese) [Google Scholar]
- Hill, B.H.; Elonen, C.M.; Jicha, T.M.; Kolka, R.K.; Lehto, L.L.P.; Sebestyen, S.D.; Seifert-Monson, L.R. Ecoenzymatic stoichiometry and microbial processing of organic matter in northern bogs and fens reveals a common p-limitation between peatland types. Biogeochemistry 2014, 120, 203–224. [Google Scholar] [CrossRef]
- Lai, J.; Zou, Y.; Zhang, J.; Peres-Neto, P.R. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca.hp R package. Methods Ecol. Evol. 2022, 13, 782–788. [Google Scholar] [CrossRef]
- Wei, X.; Shao, M.; Fu, X.; Horton, R.; Li, Y.; Zhang, X. Distribution of soil organic C, N and P in three adjacent land use patterns in the northern Loess Plateau, China. Biogeochemistry 2009, 96, 149–162. [Google Scholar] [CrossRef]
- Liu, W.; Fu, S.; Yan, S.; Ren, C.; Wu, S.; Deng, J.; Li, B.; Han, X.; Yang, G. Responses of plant community to the linkages in plant-soil C:N:P stoichiometry during secondary succession of abandoned farmlands, China. J. Arid Land 2020, 12, 215–226. [Google Scholar] [CrossRef]
- Li, Y.; Han, H.Y.; Wang, W.J.; Yang, G.F.; Zhao, C.C. Effects of different land use types on soil organic carbon and microbial respiration in Huang-Huai-Hai Plain. Ecol. Environ. Sci. 2017, 26, 62–66. (In Chinese) [Google Scholar]
- Zhou, Z.; Liu, Y.; Zhang, L.M.; Xu, Y.N.; Su, L.L.; Liao, H. Soil Nutrient Status in Wuyi Tea Region and Its Effects on Tea Quality-Related Constituents. Sci. Agric. Sin. 2019, 52, 1425–1434. (In Chinese) [Google Scholar]
- Yang, Z.; Baoyin, T.; Minggagud, H.; Sun, H.; Li, F.Y. Recovery succession drives the convergence, and grazing versus fencing drives the divergence of plant and soil N/P stoichiometry in a semiarid steppe of inner Mongolia. Plant Soil 2017, 420, 303–314. [Google Scholar] [CrossRef]
- Tessier, J.T.; Raynal, D.J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. J. Appl. Ecol. 2003, 40, 523–534. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef]
- Wang, D.; Ma, F.Y.; Yao, X.F.; Xin, H.; Song, X.; Zhang, Z.X. Properties of soil microbes, nutrients and soil enzyme activities and their relationship in a degraded wetland of Yellow River Delta. Sci. Soil Water Conserv. 2012, 10, 94–98. (In Chinese) [Google Scholar]
- Fu, B.J.; Guo, X.D.; Chen, L.D.; Ma, K.M.; Li, J.R. Soil nutrient changes due to land use changes in Northern China: A case study in Zunhua County, Hebei Province. Soil Use Manag. 2001, 17, 294–296. [Google Scholar] [CrossRef]
- Ngo-Mbogba, M.; Yemefack, M.; Nyeck, B. Assessing soil quality under different land cover types within shifting agriculture in South Cameroon. Soil Tillage Res. 2015, 150, 124–131. [Google Scholar] [CrossRef]
- Tian, J.; Sheng, M.Y.; Wang, P.; Wen, P.C. Influence of Land Use Change on Litter and Soil C, N, P Stoichiometric Characteristics and Soil Enzyme Activity in Karst Ecosystem, Southwest China. Environ. Sci. 2019, 40, 4278–4286. (In Chinese) [Google Scholar]
- Ning, M.L.; Gao, H.H.; Huang, T.Y.; Yu, W.J.; Kang, H.Z. Effects of land use patterns on soil enzyme activity in Chongming Island. Chin. J. Ecol. 2017, 36, 1949–1956. (In Chinese) [Google Scholar]
- Sun, L.; Kominami, Y.; Yoshimura, K.; Kitayama, K. Root-exudate flux variations among four co-existing canopy species in a temperate forest, Japan. Ecol. Res. 2017, 32, 331–339. [Google Scholar] [CrossRef]
- Mo, X.L.; Dai, X.Q.; Wang, H.M.; Fu, X.L.; Kou, L. Rhizosphere effects of overstory tree and understory shrub species in central subtropical plantations—A case study at Qianyanzhou, Taihe, Jiangxi, China. Chin. J. Plant Ecol. 2018, 42, 723–733. (In Chinese) [Google Scholar]
- Zhang, H.Y.; Wang, K.Q.; Song, Y.L.; Zhao, Y.Y.; Chen, X. Distribution characteristics of soil activity organic carbon in difference land use types in JianShan river watershed in middle Yunnan province. Res. Soil Water Conserv. 2019, 26, 16–21. (In Chinese) [Google Scholar]
- Wei, L.; Razavi, B.S.; Wang, W.; Zhu, Z.; Liu, S.; Wu, J.; Kuzyakov, Y.; Ge, T. Labile carbon matters more than temperature for enzyme activity in paddy soil. Soil Biol. Biochem. 2019, 135, 134–143. [Google Scholar] [CrossRef]
- Peng, X.; Wang, W. Stoichiometry of soil extracellular enzyme activity along a climatic transect in temperate grasslands of northern China. Soil Biol. Biochem. 2016, 98, 74–84. [Google Scholar] [CrossRef]
- Zhou, J.H.; Gao, R.R.; Wei, Q.; Yuan, Y.H.; Pu, H.Y. Effect of difference land use patterns on enzyme activities and microbial diversity in upland red soil. Res. Soil Water Conserv. 2020, 34, 327–332. (In Chinese) [Google Scholar]
- Liu, J.B.; Chen, J.; Chen, G.S.; Gou, J.F.; Li, Y.Q. Enzyme stoichiometry indicates the variation of microbial nutrient requirements at different soil depths in subtropical forests. PLoS ONE 2020, 15, e0220599. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, C.; Wang, Y.; Xu, Z.; Han, H.; Li, L.; Wan, S. warming and increased precipitation have differential effects on soil extracellular enzyme activities in a temperate grassland. Sci. Total Environ. 2013, 444, 552–558. [Google Scholar] [CrossRef]
- Waring, B.G.; Weintraub, S.R.; Sinsabaugh, R.L. Ecoenzymatic stoichiometry of microbial nutrient acquisition in tropical soils. Biogeochemistry 2014, 117, 101–113. [Google Scholar] [CrossRef]
- Allison, S.D.; Vitousek, P.M. Responses of extracellular enzymes to simple and complex nutrient inputs. Soil Biol. Biochem. 2005, 37, 937–944. [Google Scholar] [CrossRef]
- Gao, Q.Y.; Dai, X.Q.; Wang, J.L.; Fu, X.L.; Kou, L.; Wang, H.M. Characteristics of soil enzymes stoichiometry in rhizosphere of understory vegetation in subtropical forest plantations. Chin. J. Plant Ecol. 2019, 43, 258–272. (In Chinese) [Google Scholar] [CrossRef]
- Kivlin, S.N.; Treseder, K.K. Soil Extracellular enzyme activities correspond with abiotic factors more than fungal community composition. Biogeochemistry 2014, 117, 23–37. [Google Scholar] [CrossRef]
- Zhao, F.; Xu, B.; Yang, X.; Jin, Y.; Li, J.; Xia, L.; Chen, S.; Ma, H. Remote sensing estimates of grassland aboveground biomass based on MODIS net primary productivity (NPP): A case study in the xilingol grassland of northern China. Remote Sens. 2014, 6, 5368–5386. [Google Scholar] [CrossRef]
- Huang, H.L.; Zong, N.; He, N.P.; Tian, J. Characteristics of soil enzyme stoichiometry along an altitude gradient on Qinghai-Tibet Plateau alpine meadow, China. Chin. J. Appl. Ecol. 2019, 30, 3689–3696. (In Chinese) [Google Scholar]
- Gu, X.N.; He, H.S.; Tao, Y.; Jin, Y.H.; Zhang, X.Y.; Xu, Z.W.; Wang, Y.T.; Song, X.X. Soil microbial community structure, enzyme activities, and their influencing factors along different altitudes of Changbai Mountain. Acta Ecol. Sin. 2017, 37, 8374–8384. (In Chinese) [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | pH | SWC (%) | Soil Type | Soil Temperature (℃) |
|---|---|---|---|---|
| PM | 4.85 ± 0.62 | 23.54 ± 0.07 | Red soil | 22.4 ± 1.23 |
| PM tea plantation | 4.54 ± 0.26 | 21.45 ± 0.06 | Red soil | 21.05 ± 0.99 |
| MF | 4.65 ± 0.15 | 29.94 ± 0.05 | Red soil | 22 ± 0.24 |
| MF tea plantation | 4.04 ± 0.12 | 27.97 ± 0.03 | Red soil | 21.5 ± 0.37 |
| BA | 5.14 ± 0.22 | 36.09 ± 0.06 | Red soil | 20.5 ± 0.69 |
| BA tea plantation | 4.24 ± 0.24 | 28.45 ± 0.01 | Red soil | 19.6 ± 0.45 |
| Type | ΔTC (%) | ΔTN (%) | ΔTP (%) | ΔC/N (%) | ΔC/P (%) | ΔN/P (%) |
|---|---|---|---|---|---|---|
| PM-Tea plantation | −138.9 ± 19.51A | −128.98 ± 16.86A | 65.9 ± 27.85A | −22.23 ± 4.17A | −1224.47 ± 238.41B | −1130.35 ± 320.62A |
| MF-Tea plantation | −90.61 ± 19.33AB | −63.37 ± 22.55B | 81.88 ± 16.30A | −25.33 ± 6.51A | −1892.32 ± 347.86A | −1159.5 ± 306.60A |
| BA-Tea plantation | −67.64 ± 19.17B | −58.02 ± 10.97B | 70.11 ± 20.47A | −4.30 ± 0.98B | −622.16 ± 154.43C | −513.83 ± 100.28B |
| Type | EC/N | EC/P | EN/P | Vector Length | Vector Angle |
|---|---|---|---|---|---|
| PM | 1.06 ± 0.04a | 0.85 ± 0.09a | 0.80 ± 0.07a | 1.36 ± 0.09a | 51.29 ± 2.54a |
| PM_tea plantation | 1.06 ± 0.09a | 0.86 ± 0.08a | 0.81 ± 0.03a | 1.37 ± 0.13a | 50.91 ± 0.91a |
| MF | 1.04 ± 0.15a | 0.83 ± 0.05b | 0.79 ± 0.06a | 1.33 ± 0.07b | 51.58 ± 1.92a |
| MF_tea plantation | 1.09 ± 0.09a | 0.87 ± 0.01a | 0.80 ± 0.06a | 1.40 ± 0.07a | 51.49 ± 2.27a |
| BA | 0.98 ± 0.05a | 0.85 ± 0.03b | 0.87 ± 0.02b | 1.30 ± 0.06a | 48.94 ± 0.73a |
| BA_tea plantation | 0.97 ± 0.08a | 0.87 ± 0.04a | 0.89 ± 0.04a | 1.30 ± 0.09a | 48.23 ± 1.10a |
| Soil Property | TC | TN | TP | C/N | C/P | N/P | |
|---|---|---|---|---|---|---|---|
| Enzyme | |||||||
| BG | 0.784 ** | 0.640 ** | −0.077 | 0.025 | 0.378 * | 0.406 ** | |
| NAG | 0.805 ** | 0.949 ** | 0.088 | −0.368 * | 0.108 | 0.172 | |
| ACP | 0.416 ** | 0.320 * | −0.391 * | 0.301 | 0.266 | 0.289 | |
| EC/N | 0.171 | −0.117 | −0.102 | 0.426 ** | 0.212 | 0.148 | |
| EC/P | 0.515 ** | 0.427 ** | 0.246 | −0.122 | 0.135 | 0.114 | |
| EN/P | 0.427 ** | 0.597 ** | 0.404 ** | −0.536 ** | −0.051 | −0.017 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhang, J.; Qiu, Q.; Zhou, Y.; You, W. Changes in Soil Properties and Enzyme Stoichiometry in Three Different Forest Types Changed to Tea Plantations. Forests 2023, 14, 2043. https://doi.org/10.3390/f14102043
Li Y, Zhang J, Qiu Q, Zhou Y, You W. Changes in Soil Properties and Enzyme Stoichiometry in Three Different Forest Types Changed to Tea Plantations. Forests. 2023; 14(10):2043. https://doi.org/10.3390/f14102043
Chicago/Turabian StyleLi, Ying, Jinlin Zhang, Qingyan Qiu, Yan Zhou, and Weibin You. 2023. "Changes in Soil Properties and Enzyme Stoichiometry in Three Different Forest Types Changed to Tea Plantations" Forests 14, no. 10: 2043. https://doi.org/10.3390/f14102043
APA StyleLi, Y., Zhang, J., Qiu, Q., Zhou, Y., & You, W. (2023). Changes in Soil Properties and Enzyme Stoichiometry in Three Different Forest Types Changed to Tea Plantations. Forests, 14(10), 2043. https://doi.org/10.3390/f14102043