

Variables Affecting the Pine Processionary Moth Flight: A Survey in the North-Western Italian Alps


Abstract
1. Introduction
2. Materials and Methods
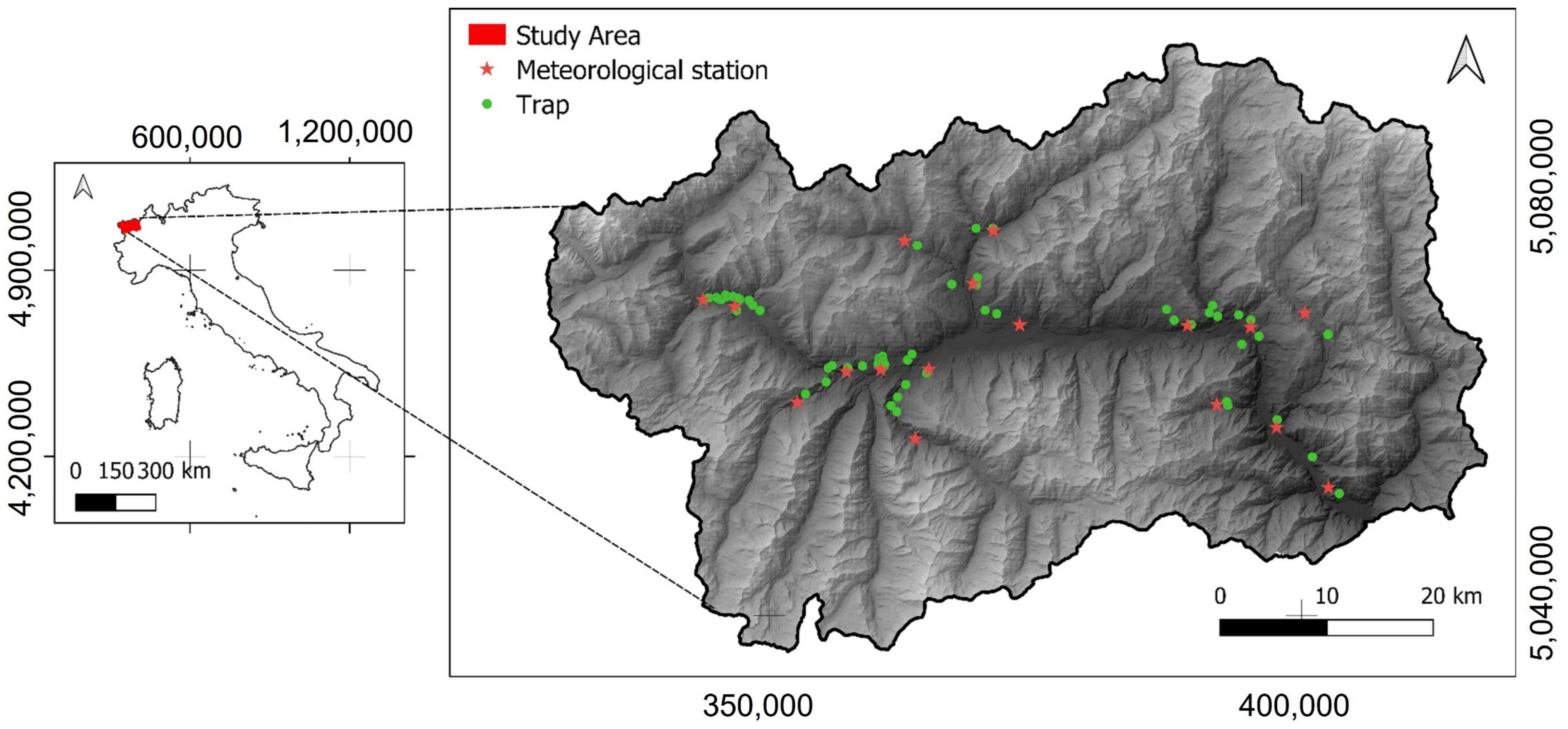
2.1. Study Area
2.2. Seasonal Flight Activity of the Pine Processionary Moth
2.3. Infestation Index
2.4. Meteorological Data
2.5. Data Collection and Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations Kyoto Protocol—Targets for the First Commitment Period. Available online: http://unfccc.int/kyoto_protocol/items/2830.php (accessed on 15 November 2022).
- Malamel, J. Seasonal dynamics on spider population in Pathiramanal island, Kerala, India: A case study. In Arthropods: Are They Beneficial for Mankind? Ranz, R.E.R., Ed.; IntechOpen: London, UK, 2021; p. 55. [Google Scholar]
- Pureswaran, D.S.; Roques, A.; Battisti, A. Forest insects and climate change. Curr. For. Rep. 2018, 4, 35–50. [Google Scholar] [CrossRef]
- Netherer, S.; Schopf, A. Potential effects of climate change on insect herbivores in European forests—General aspects and the pine processionary moth as specific example. For. Ecol. Manag. 2010, 259, 831–838. [Google Scholar] [CrossRef]
- Battisti, A.; Stastny, M.; Netherer, S.; Robinet, C.; Schopf, A.; Roques, A.; Larsson, S. Expansion of geographic range in the pine processionary moth caused by increased winter temperatures. Ecol. Appl. 2005, 15, 2084–2096. [Google Scholar] [CrossRef]
- Battisti, A.; Stastny, M.; Buffo, E.; Larsson, S. A rapid altitudinal range expansion in the pine processionary moth produced by the 2003 climatic anomaly. Glob. Chang. Biol. 2006, 12, 662–671. [Google Scholar] [CrossRef]
- Buffo, E.; Battisti, A.; Stastny, M.; Larsson, S. Temperature as a predictor of survival of the pine processionary moth in the Italian Alps. Agric. For. Entomol. 2007, 9, 65–72. [Google Scholar] [CrossRef]
- Jactel, H.; Menassieu, P.; Vétillard, F.; Barthélémy, B.; Piou, D.; Frérot, B.; Rousselet, J.; Goussard, F.; Branco, M.; Battisti, A. Population monitoring of the pine processionary moth (Lepidoptera: Thaumetopoeidae) with pheromone-baited traps. For. Ecol. Manag. 2006, 235, 96–106. [Google Scholar] [CrossRef]
- Robinet, C.; Rousselet, J.; Roques, A. Potential spread of the pine processionary moth in France: Preliminary results from a simulation model and future challenges. Ann. For. Sci. 2014, 71, 149–160. [Google Scholar] [CrossRef]
- Ferracini, C.; Pogolotti, C.; Alma, A. A mismatch in the emergence of Torymus sinensis may affect the effectiveness of this biocontrol agent? Biol. Control 2022, 174, 105029. [Google Scholar] [CrossRef]
- Forrest, J. Complex responses of insect phenology to climate change. Curr. Opin. Insect Sci. 2016, 17, 49–54. [Google Scholar] [CrossRef]
- Gordo, O.; Sanz, J.J. Phenology and climate change: A long-term study in a Mediterranean locality. Oecologia 2005, 146, 484–495. [Google Scholar] [CrossRef]
- Bryant, S.R.; Thomas, C.D.; Bale, J.S. Thermal ecology of gregarious and solitary nettle-feeding nymphalid butterfly larvae. Oecologia 2000, 122, 1–10. [Google Scholar] [CrossRef]
- Roy, D.B.; Sparks, T.H. Phenology of British butterflies and climate change. Glob. Chang. Biol. 2000, 6, 407–416. [Google Scholar] [CrossRef]
- Robinet, C.; Baier, P.; Pennerstorfer, J.; Schopf, A.; Roques, A. Modelling the effects of climate change on the potential feeding activity of Thaumetopoea pityocampa (Den. & Schiff.) (Lep., Notodontidae) in France. Glob. Ecol. Biogeogr. 2007, 16, 460–471. [Google Scholar] [CrossRef]
- Pimentel, C.; Calvão, T.; Ayres, M. Impact of climatic variation on populations of pine processionary moth Thaumetopoea pityocampa in a core area of its distribution. Agric. For. Entomol. 2011, 13, 273–281. [Google Scholar] [CrossRef]
- Camarero, J.; Tardif, J.; Gazol, A.; Conciatori, F. Pine processionary moth outbreaks cause longer growth legacies than drought and are linked to the North Atlantic Oscillation. Sci. Total Environ. 2022, 819, 153041. [Google Scholar] [CrossRef]
- Leather, S.; Walters, K.; Bale, J. The Ecology of Insect Overwintering; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Arnaldo, P.; Oliveira, I.; Santos, J.; Leite, S. Climate change and forest plagues: The case of the pine. For. Syst. 2011, 20, 508–515. [Google Scholar] [CrossRef]
- Huchon, H.; Démolin, G. La bioécologie de la Processionnaire du pin: Dispersion potentielle, dispersion actuelle. Rev. For. Française 1970, 22, 220–234. [Google Scholar] [CrossRef]
- Robinet, C.; Rousselet, J.; Pineau, P.; Miard, F.; Roques, A. Are heat waves susceptible to mitigate the expansion of a species progressing with global warming? Ecol. Evol. 2013, 3, 2947–2957. [Google Scholar] [CrossRef]
- Toïgo, M.; Barraquand, F.; Barnagaud, J.; Piou, D.; Jactel, H. Geographical variation in climatic drivers of the pine processionary moth population dynamics. For. Ecol. Manag. 2017, 404, 141–155. [Google Scholar] [CrossRef]
- Ferracini, C.; Saitta, V.; Pogolotti, C.; Rollet, I.; Vertui, F.; Dovigo, L. Monitoring and management of the pine processionary moth in the north-western Italian Alps. Forests 2020, 11, 1253. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Analysing Ecological Data; Statistics for Biology and Health; Springer: New York, NY, USA, 2007; Volume 680, ISBN 9780387459677. [Google Scholar]
- Haelewaters, D.; Hiller, T.; Ceryngier, P.; Eschen, R.; Gorczak, M.; Houston, M.L.; Kisło, K.; Knapp, M.; Landeka, N.; Pfliegler, W.P.; et al. Do biotic and abiotic factors influence the prevalence of a common parasite of the invasive alien ladybird Harmonia axyridis? Front. Ecol. Evol. 2022, 10, 773423. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Statistics for Biology and Health; Springer: New York, NY, USA, 2009; Volume 574, ISBN 9780387874579. [Google Scholar]
- Zhang, Y.; Xue, Y.; Xu, B.; Chongliang, Z.; Zhang, C.; Zan, X. Evaluating the effect of input variables on quantifying the spatial distribution of croaker Johnius belangerii in Haizhou Bay, China. J. Oceanol. Limnol. 2021, 39, 1570–1583. [Google Scholar] [CrossRef]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer Science & Business Media: Berlin/Heidelberg, Germany; Springer: New York, NY, USA, 2006; ISBN 9780387989570. [Google Scholar]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning: With Applications in R; Springer: New York, NY, USA, 2013; p. 426. [Google Scholar] [CrossRef]
- Kuhn, M. Caret: Classification and Regression Training. R Package Version 6.0-93. 2022. Available online: https://CRAN.R-project.org/package=caret (accessed on 15 November 2022).
- Lüdecke, D.; Ben-Shachar, M.S.; Patil, I.; Waggoner, P.; Makowski, D. Performance: An R Package for Assessment, Comparison and Testing of Statistical Models. J. Open Source Softw. 2021, 6, 3139. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. 2022. Available online: https://www.r-project.org/ (accessed on 15 November 2022).
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall: London, UK; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate Change and Forest Disturbances: Climate Change Can Affect Forests by Altering the Frequency, Intensity, Duration, and Timing of Fire, Drought, Introduced Species, Insect and Pathogen Outbreaks, Hurricanes, Windstorms, Ice Storms, or Landslides. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Vercelli, M.; Novelli, S.; Ferrazzi, P.; Lentini, G.; Ferracini, C. A qualitative analysis of beekeepers’ perceptions and farm management adaptations to the impact of climate change on honey bees. Insects 2021, 12, 228. [Google Scholar] [CrossRef]
- Mirchev, P.; Georgiev, G.; Tsankov, G. Long-term studies on egg parasitoids of pine processionary moth (Thaumetopoea pityocampa) in a new locality in Bulgaria. J. Entomol. Res. Soc. 2017, 19, 15–25. [Google Scholar]
- Sinclair, B.; Addo-Bediako, A.; Chown, S. Climatic variability and the evolution of insect freeze tolerance. Biol. Rev. 2003, 78, 181–195. [Google Scholar] [CrossRef]
- Bonsignore, C.P.; Manti, F. Influence of habitat and climate on the capture of male pine processionary moths. Bull. Insectol. 2013, 66, 27–34. [Google Scholar]
- Ishiguri, Y.; Shirai, Y. Flight activity of the peach fruit moth, Carposina sasakii (Lepidoptera: Carposinidae), measured by a flight mill. Appl. Entomol. Zool. 2004, 39, 127–131. [Google Scholar] [CrossRef][Green Version]
- Tamburini, G.; Marini, L.; Hellrigl, K.; Salvadori, C.; Battisti, A. Effects of climate and density-dependent factors on population dynamics of the pine processionary moth in the Southern Alps. Clim. Chang. 2013, 121, 701–712. [Google Scholar] [CrossRef]
- Rocha, S.; Kerdelhué, C.; Jamaa, M.B.; Dhahri, S.; Burban, C.; Branco, M. Effect of heat waves on embryo mortality in the pine processionary moth. Bull. Entomol. Res. 2017, 107, 583–591. [Google Scholar] [CrossRef]
- Démolin, G. Comportement des adultes de Thaumetopoea pityocampa Schiff. Dispersion spatiale, importance écologique. Ann. Sci. For. 1969, 26, 81–102. [Google Scholar] [CrossRef]
- Battisti, A.; Bernardi, M.; Ghiraldo, C. Predation by the hoopoe (Upupa epops) on pupae of Thaumetopoea pityocampa and the likely influence on other natural enemies. BioControl 2000, 45, 311–323. [Google Scholar] [CrossRef]
- Torres-Muros, L.; Hódar, J.; Zamora, R. Effect of habitat type and soil moisture on pupal stage of a Mediterranean forest pest (Thaumetopoea pityocampa). Agric. For. Entomol. 2017, 19, 130–138. [Google Scholar] [CrossRef]
- Weseloh, R.; Andreadis, T.; Onstad, D. Modeling the influence of rainfall and temperature on the phenology of infection of gypsy moth, Lymantria dispar, larvae by the fungus Entomophaga maimaiga. Biol. Control 1993, 3, 311–318. [Google Scholar] [CrossRef]
- Baronio, P.; Rocchetta, G.; Baldassarri, N. Una stima delle popolazioni di Thaumetopoea pityocampa (Den. & Schiff.) (Lepidoptera: Thaumetopoeidae) in alcune vallate dell’Appennino forlivese. Boll. Ist. Entomol. Guid. Grandi Univ. Bologna 1994, 48, 11–17. [Google Scholar]
- Devkota, B.; Breuer, M.; Schmidt, G.H. Observations on the flight activity of the pine processionary moth Thaumetopoea pityocampa (Den. and Schiff.) in Greece, using synthetic sex-pheromone and light traps (Insecta: Lepidoptera: Thaumetopoeidae). Boll. Zool. Agrar. Bachic. 1992, 24, 147–157. [Google Scholar]
- Roversi, P. Observations on the use of traps with the sex pheromone of Thaumetopoea pityocampa (Den. & Schiff.) on the Gargano promontory (Lepidoptera, Thaumetopoeidae). Redia 1985, 68, 1–17. [Google Scholar]
- Auger-Rozenberg, M.A.; Barbaro, L.; Battisti, A.; Blache, S.; Charbonnier, Y.; Denux, O.; Garcia, J.; Goussard, F.; Imbert, C.E.; Kerdelhué, C.; et al. Ecological responses of parasitoids, predators and associated insect communities to the climate–driven expansion of the pine processionary moth. In Processionary Moths and Climate Change: An Update; Roques, A., Ed.; Springer: Dordrecht, The Netherlands, 2015; pp. 311–357. ISBN 9789401793407. [Google Scholar]
- Masutti, L.; Battisti, A. Thaumetopoea pityocampa (Den. & Schiff.) in Italy Bionomics and perspectives of integrated control. J. Appl. Entomol. 1990, 110, 229–234. [Google Scholar] [CrossRef]
- Gazol, A.; Hernández-Alonso, R.; Camarero, J.J. Patterns and drivers of pine processionary moth defoliation in Mediterranean mountain forests. Front. Ecol. Evol. 2019, 7, 458. [Google Scholar] [CrossRef]
- Leidemer, T.; Gonroudobou, O.B.H.; Nguyen, H.T.; Ferracini, C.; Burkhard, B.; Diez, Y.; Lopez Caceres, M.L. Classifying the Degree of Bark Beetle-Induced Damage on Fir (Abies mariesii) Forests, from UAV-Acquired RGB Images. Computation 2022, 10, 63. [Google Scholar] [CrossRef]
- Otsu, K.; Pla, M.; Vayreda, J.; Brotons, L. Calibrating the severity of forest defoliation by pine processionary moth with Landsat and UAV imagery. Sensors 2018, 18, 3278. [Google Scholar] [CrossRef] [PubMed]
- Cardil, A.; Otsu, K.; Pla, M.; Silva, C.A.; Brotons, L. Quantifying pine processionary moth defoliation in a pine-oak mixed forest using unmanned aerial systems and multispectral imagery. PLoS ONE 2019, 14, e0213027. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Daudin, J.J.; Piou, D.; Robinet, C.; Jactel, H. Periodicity and synchrony of pine processionary moth outbreaks in France. For. Ecol. Manag. 2015, 354, 309–317. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Original Variables | Correlation | Contribution | ||
|---|---|---|---|---|
| PC 1 | PC 2 | PC 1 | PC 2 | |
| Altitude | −0.48 | −0.27 | 20.2 | 12.1 |
| Intensity of attack | −0.27 | 0.13 | 11.4 | 5.9 |
| Rain at DTmax | −0.30 | 0.61 | 12.6 | 27.9 |
| Rain at May | −0.32 | 0. 61 | 13.6 | 27.8 |
| Temp at DTmax | 0.50 | 0.30 | 21.0 | 13.7 |
| Temp at May | 0.50 | 0.28 | 21.1 | 12.6 |
| (Percentage of variance explained) | 40.47% | 31.76% | ||
| Maximum capture | Parametric Coefficients | Estimate | SE | t-Value | p-Value |
| (Intercept) | 6.32 | 0.156 | 40.47 | <0.0001 | |
| PC1 | −0.05 | 0.067 | −0.73 | 0.46 | |
| PC2 | 0.12 | 0.059 | 2.12 | 0.035 | |
| Smooth term | Estimated d.f. | Reference d.f. | F-value | p-value | |
| s(year) | 2.02 | 4 | 6.69 | <0.0001 | |
| AIC = 906.34, ΔAIC (vs. model with no random eff.) = 17.26, LRT = 19.26, p < 0.0001 | |||||
| Total capture | Parametric coefficients | Estimate | SE | t-value | p-value |
| (Intercept) | 8.08 | 0.153 | 52.71 | <0.0001 | |
| PC1 | −0.05 | 0.069 | −0.76 | 0.45 | |
| PC2 | 0.10 | 0.062 | 1.66 | 0.099 | |
| Smooth term | Estimated d.f. | Reference d.f. | F-value | p-value | |
| s(year) | 1.77 | 4 | 9.16 | <0.0001 | |
| AIC = 955.92, ΔAIC (vs. model with no random eff.) = 8.69, LRT = 10.69, p = 0.0011 | |||||
| Day at maximum capture | Parametric coefficients | Estimate | SE | t-value | p-value |
| (Intercept) | 7.66 | 0.046 | 166.43 | <0.0001 | |
| PC1 | 0.09 | 0.007 | 13.75 | <0.0001 | |
| PC2 | 0.07 | 0.005 | 13.20 | <0.0001 | |
| Smooth term | Estimated d.f. | Reference d.f. | F-value | p-value | |
| s(year) | 3.89 | 4 | 73.96 | <0.0001 | |
| AIC = −741.62, ΔAIC (vs. model with no random eff.) = 92.53, LRT = 94.53, p < 0.0001 | |||||
| Day at first capture | Parametric coefficients | Estimate | SE | t-value | p-value |
| (Intercept) | 7.48 | 0.008 | 959.95 | <0.0001 | |
| PC1 | 0.003 | 0.004 | 0.78 | 0.43 | |
| PC2 | 0.001 | 0.005 | 0.43 | 0.67 | |
| Smooth term | Estimated d.f. | Reference d.f. | F-value | p-value | |
| s(year) | 3.86 | 4 | 24.04 | <0.0001 | |
| AIC = −779.21, ΔAIC (vs. model with no random eff.) = 16.27, LRT = 18.27, p < 0.0001 | |||||
| Day at last capture | Parametric coefficients | Estimate | SE | t-value | p-value |
| (Intercept) | 7.89 | 0.007 | 1154.34 | <0.0001 | |
| PC1 | −0.003 | 0.003 | −0.98 | 0.33 | |
| PC2 | 0.01 | 0.003 | 2.01 | 0.046 | |
| Smooth term | Estimated d.f. | Reference d.f. | F-value | p-value | |
| s(year) | 2.88 | 4 | 28.44 | <0.0001 | |
| AIC = −875.93, ΔAIC (vs. model with no random eff.) = 22.81, LRT = 24.81, p < 0.0001 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferracini, C.; Saitta, V.; Rondoni, G.; Rollet, I. Variables Affecting the Pine Processionary Moth Flight: A Survey in the North-Western Italian Alps. Forests 2023, 14, 31. https://doi.org/10.3390/f14010031
Ferracini C, Saitta V, Rondoni G, Rollet I. Variables Affecting the Pine Processionary Moth Flight: A Survey in the North-Western Italian Alps. Forests. 2023; 14(1):31. https://doi.org/10.3390/f14010031
Chicago/Turabian StyleFerracini, Chiara, Valerio Saitta, Gabriele Rondoni, and Ivan Rollet. 2023. "Variables Affecting the Pine Processionary Moth Flight: A Survey in the North-Western Italian Alps" Forests 14, no. 1: 31. https://doi.org/10.3390/f14010031
APA StyleFerracini, C., Saitta, V., Rondoni, G., & Rollet, I. (2023). Variables Affecting the Pine Processionary Moth Flight: A Survey in the North-Western Italian Alps. Forests, 14(1), 31. https://doi.org/10.3390/f14010031