
Variation of Cones Production in a Silver Fir (Abies alba Mill.) Clonal Seed Orchard


Abstract
1. Introduction
2. Materials and Methods
2.1. Silver Fir Clonal Seed Orchard
2.2. Data Analysis
2.2.1. Genetic Variation
2.2.2. Female Fertility Variation
3. Results
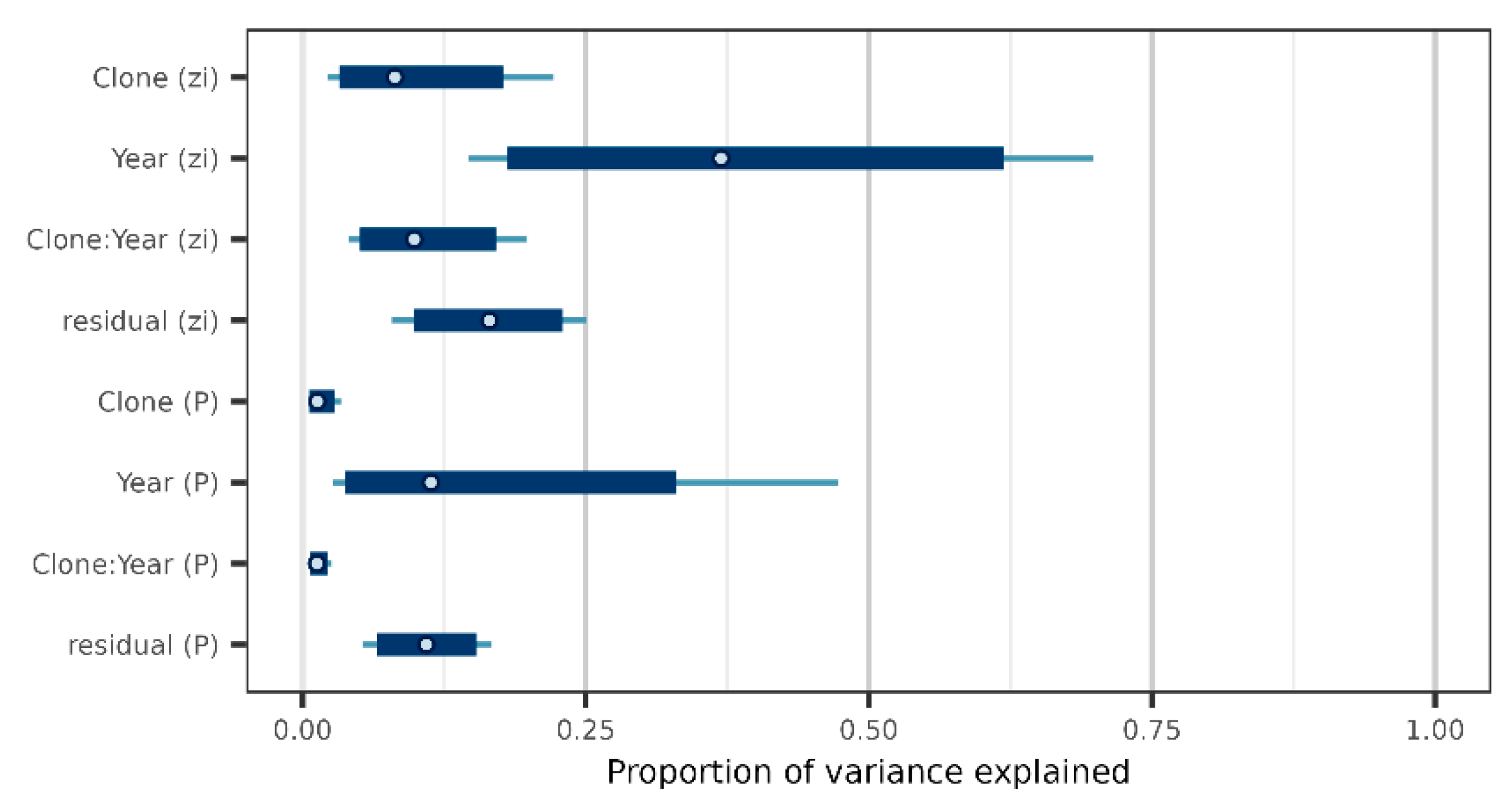
3.1. Genetic Variation
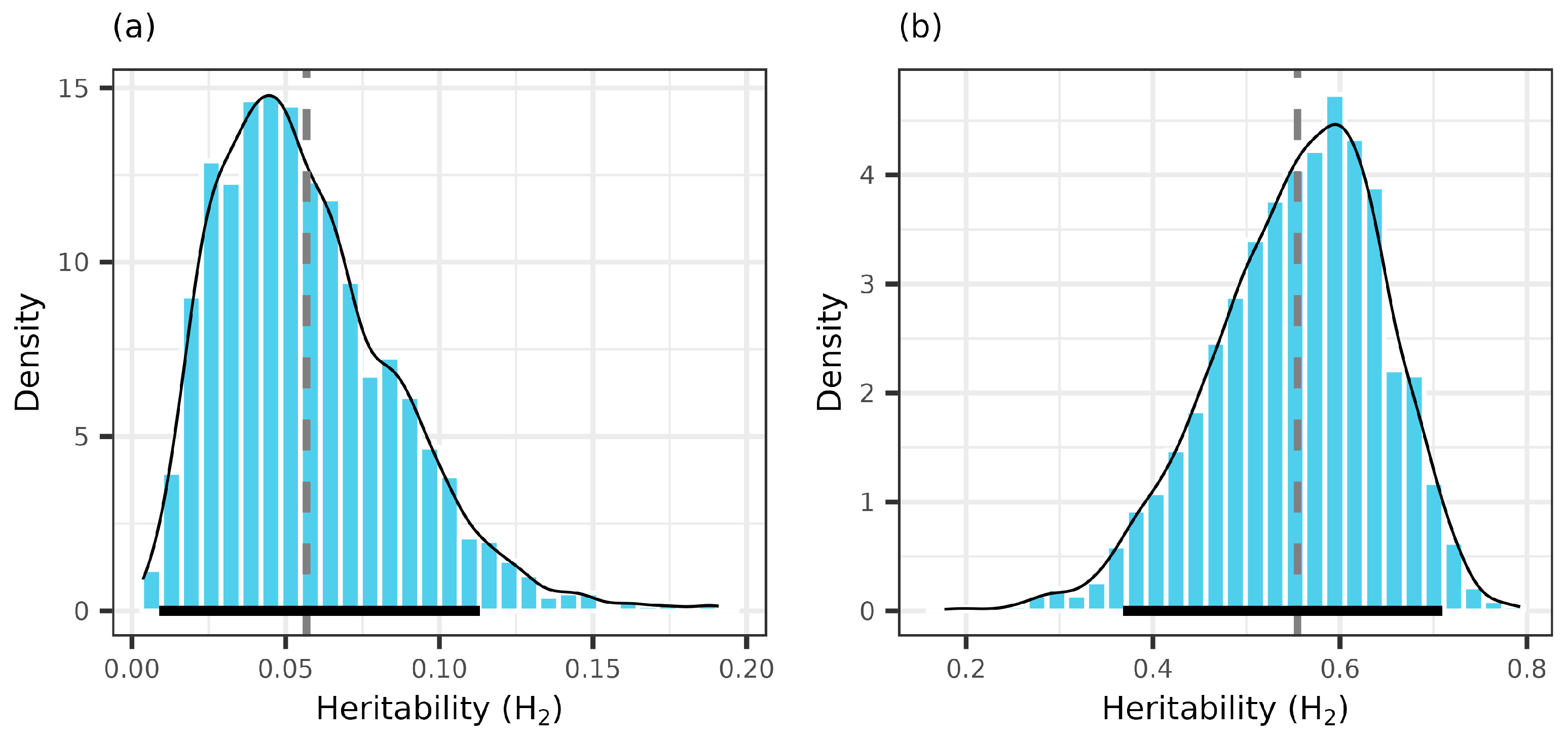
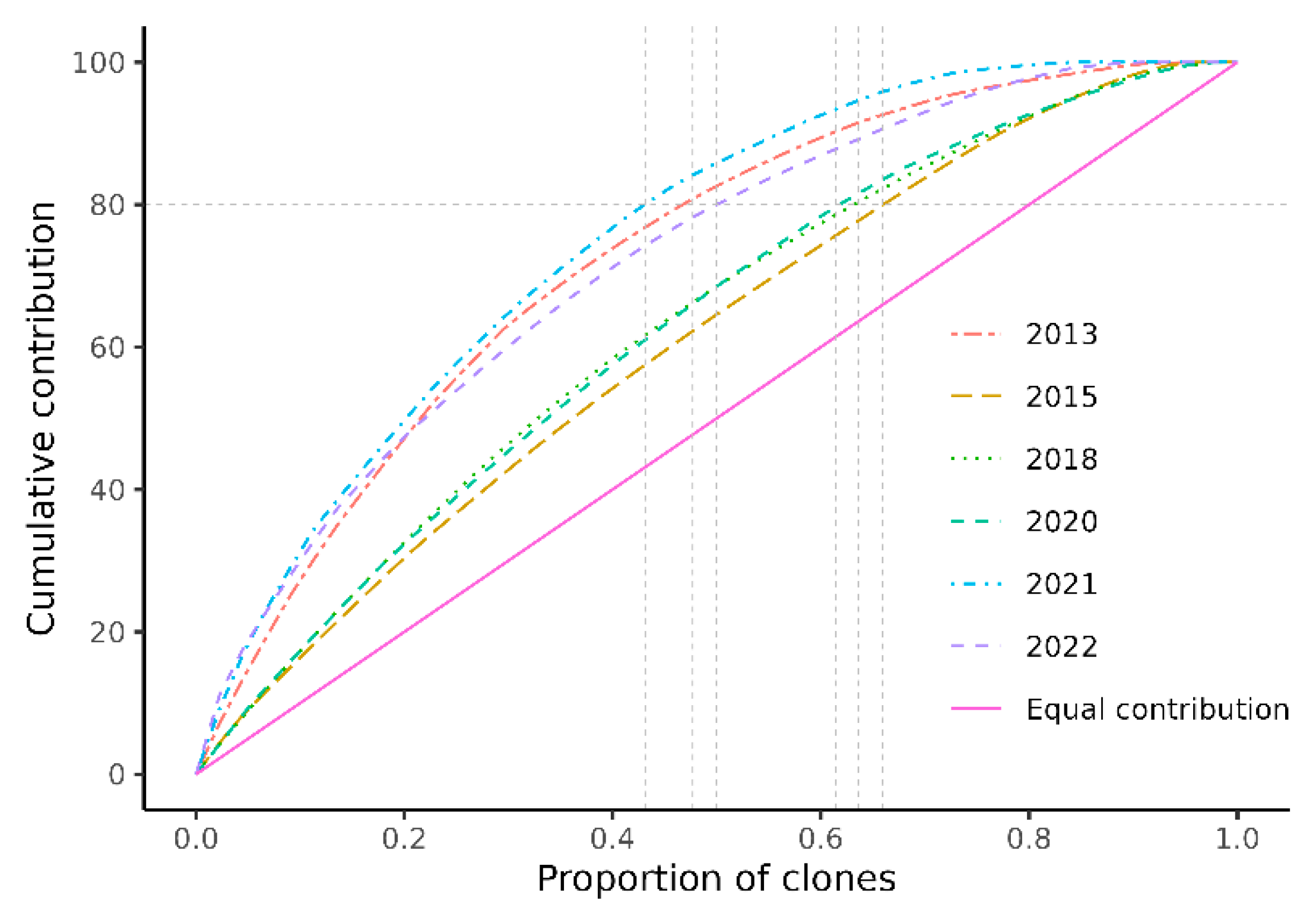
3.2. Female Fertility Variation and Genetic Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Kassaby, Y.; Fashler, A.K.; Sziklai, O. Reproductive Phenology and Its Impact on Genetically Improved Seed Production in a Douglas-fir Seed Orchard. Silvae Genet. 1984, 33, 120–125. [Google Scholar]
- Codesido, V.; Fernández-López, J. Juvenile Radiata Pine Clonal Seed Orchard Management in Galicia (NW Spain). Eur. J. For. Res. 2014, 133, 177–190. [Google Scholar] [CrossRef]
- Funda, T.; El-Kassaby, Y.A. Seed Orchard Genetics. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2012, 7, 1–23. [Google Scholar] [CrossRef]
- Kang, K.-S.; Kim, C.-S. Clonal Fertility Variation and Its Effects on the Effective Population Size in the Seed Orchard of Dioecious Species, Fraxinus Rhynchophylla. Silvae Genet. 2012, 61, 79–84. [Google Scholar] [CrossRef][Green Version]
- Bilir, N.; Kang, K.-S. Fertility Variation, Seed Collection and Gene Diversity in Natural Stands of Taurus Cedar (Cedrus libani). Eur. J. For. Res. 2021, 140, 199–208. [Google Scholar] [CrossRef]
- Kang, K.; Harju, A.; Lindgren, D.; Nikkanen, T.; Almqvist, C.; Suh, G. Variation in Effective Number of Clones in Seed Orchards. New For. 2001, 21, 17–33. [Google Scholar] [CrossRef]
- Prescher, F.; Lindgren, D.; Almqvist, C.; Kroon, J.; Lestander, T.A.; Mullin, T.J. Female Fertility Variation in Mature Pinus sylvestris Clonal Seed Orchards. Scand. J. For. Res. 2007, 22, 280–289. [Google Scholar] [CrossRef]
- Kang, K.-S. Estimation of Fertility Variation in Forest Tree Populations. Forestry 2003, 76, 329–344. [Google Scholar] [CrossRef]
- Chen, X.; Guo, Q.; Brockway, D.G. Analyzing the Complexity of Cone Production in Longleaf Pine by Multiscale Entropy. J. Sustain. For. 2016, 35, 172–182. [Google Scholar] [CrossRef]
- Matziris, D. Variation in Cone Production in a Clonal Seed Orchard of Black Pine. Silvae Genet. 1993, 42, 136–141. [Google Scholar]
- Griffin, A. Clonal Variation in Radiata Pine Seed Orchards. I: Some Flowering, Cone and Seed Production Traits. Aust. For. Res. 1983, 12, 295–302. [Google Scholar]
- Askew, G. Estimation of Gamete Pool Compositions in Clonal Seed Orchards. Silvae Genet. 1988, 37, 5–6. [Google Scholar]
- Any, Y.E.-K.; FAsıER, A.; Crown, M. Variation in Fruitfulness in a Douglas-fir Seed Orchard and Its Effect on Crop-Management Decisions. Silvae Genet. 1989, 38, 3–4. [Google Scholar]
- Kang, K.S.; Mullin, T.J. Variation in Clone Fertility and Its Effect on the Gene Diversity of Seeds From a Seed Orchard of Chamaecyparis obtusa in Korea. Silvae Genet. 2007, 56, 134–137. [Google Scholar] [CrossRef][Green Version]
- Muñoz-Gutiérrez, L.; Vargas-Hernández, J.J.; López-Upton, J.; Ramírez-Herrera, C.; Jiménez-Casas, M. Clonal Variation in Phenological Synchronization and Cone Production in a Pinus patula Seed Orchard. Silvae Genet. 2020, 69, 130–138. [Google Scholar] [CrossRef]
- Anonymous. Twentieth Annual Report. N.C. State Cooperative Tree Improvement and Hardwood Research Program; School of Forest Resources, North Carolina State University: Raleigh, NC, USA, 1976. [Google Scholar]
- Park, J.-M.; Kang, H.-I.; Yeom, D.-B.; Kang, K.-S.; El-Kassaby, Y.A.; Lee, K.-M. Gender, Reproductive Output Covariation and Their Role on Gene Diversity of Pinus Koraiensis Seed Orchard Crops. BMC Plant Biol. 2020, 20, 418. [Google Scholar] [CrossRef]
- Pick, J.L.; Lemon, H.E.; Thomson, C.E.; Hadfield, J.D. Decomposing Phenotypic Skew and Its Effects on the Predicted Response to Strong Selection. Nat. Ecol. Evol. 2022, 6, 774–785. [Google Scholar] [CrossRef]
- Martin, T.G.; Wintle, B.A.; Rhodes, J.R.; Kuhnert, P.M.; Field, S.A.; Low-Choy, S.J.; Tyre, A.J.; Possingham, H.P. Zero Tolerance Ecology: Improving Ecological Inference by Modelling the Source of Zero Observations: Modelling Excess Zeros in Ecology. Ecol. Lett. 2005, 8, 1235–1246. [Google Scholar] [CrossRef]
- Calama, R.; Mutke, S.; Tomé, J.; Gordo, J.; Montero, G.; Tomé, M. Modelling Spatial and Temporal Variability in a Zero-Inflated Variable: The Case of Stone Pine (Pinus pinea L.) Cone Production. Ecol. Model. 2011, 222, 606–618. [Google Scholar] [CrossRef]
- Andrus, R.A.; Harvey, B.J.; Hoffman, A.; Veblen, T.T. Reproductive Maturity and Cone Abundance Vary with Tree Size and Stand Basal Area for Two Widely Distributed Conifers. Ecosphere 2020, 11, e03092. [Google Scholar] [CrossRef]
- Kang, K.-S. Clonal and Annual Variation of Flower Production and Composition of Gamete Gene Pool in a Clonal Seed Orchard of Pinus Densiflora. Can. J. For. Res. 2000, 30, 1275–1280. [Google Scholar] [CrossRef]
- O’Hara, R.B.; Kotze, D.J. Do Not Log-Transform Count Data. Methods Ecol. Evol. 2010, 1, 118–122. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. Repeatability for Gaussian and Non-Gaussian Data: A Practical Guide for Biologists. Biol. Rev. 2010, 85, 935–956. [Google Scholar] [CrossRef] [PubMed]
- de Villemereuil, P.; Rutschmann, A.; Lee, K.D.; Ewen, J.G.; Brekke, P.; Santure, A.W. Little Adaptive Potential in a Threatened Passerine Bird. Curr. Biol. 2019, 29, 889–894.e3. [Google Scholar] [CrossRef] [PubMed]
- Moiron, M.; Charmantier, A.; Bouwhuis, S. The Quantitative Genetics of Fitness in a Wild Seabird. Evolution 2022, 76, 1443–1452. [Google Scholar] [CrossRef]
- Ferreira, F.M.; Evangelista, J.S.P.C.; Chaves, S.F.d.S.; Alves, R.S.; Silva, D.B.; Malikouski, R.G.; Resende, M.D.V.; Bhering, L.L.; Santos, G.A. Multivariate Bayesian Analysis for Genetic Evaluation and Selection of Eucalyptus in Multiple Environment Trials. Bragantia 2022, 81, e2922. [Google Scholar] [CrossRef]
- Peixoto, M.A.; Evangelista, J.S.P.C.; Coelho, I.F.; Alves, R.S.; Laviola, B.G.; Fonseca e Silva, F.; Resende, M.D.V.d.; Bhering, L.L. Multiple-Trait Model Through Bayesian Inference Applied to Jatropha curcas Breeding for Bioenergy. PLoS ONE 2021, 16, e0247775. [Google Scholar] [CrossRef]
- Tinner, W.; Colombaroli, D.; Heiri, O.; Henne, P.D.; Steinacher, M.; Untenecker, J.; Vescovi, E.; Allen, J.R.M.; Carraro, G.; Conedera, M.; et al. The Past Ecology of Abies alba Provides New Perspectives on Future Responses of Silver Fir Forests to Global Warming. Ecol. Monogr. 2013, 83, 419–439. [Google Scholar] [CrossRef]
- Bucher, H.U. Abies alba. In Enzyklopädie der Holzgewächse: Handbuch und Atlas der Dendrologie; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; pp. 1–18. ISBN 978-3-527-67851-8. [Google Scholar]
- San-Miguel-Ayanz, J.; De Rigo, D.; Caudullo, G.; Durrant, T.H.; Mauri, A. European Atlas of Forest Tree Species. Abies alba in Europe: Distribution, Habitat, Usage and Threats; Publications Office of the European Union: Luxemburg, 2016. [Google Scholar]
- Dobrowolska, D.; Bončina, A.; Klumpp, R. Ecology and Silviculture of Silver Fir (Abies alba Mill.): A Review. J. For. Res. 2017, 22, 326–335. [Google Scholar] [CrossRef]
- Fady, B.; Aravanopoulos, F.A.; Alizoti, P.; Mátyás, C.; von Wühlisch, G.; Westergren, M.; Belletti, P.; Cvjetkovic, B.; Ducci, F.; Huber, G.; et al. Evolution-Based Approach Needed for the Conservation and Silviculture of Peripheral Forest Tree Populations. For. Ecol. Manag. 2016, 375, 66–75. [Google Scholar] [CrossRef]
- Teodosiu, M.; Mihai, G.; Fussi, B.; Ciocîrlan, E. Genetic Diversity and Structure of Silver Fir (Abies alba Mill.) At the South-Eastern Limit of Its Distribution Range. Ann. For. Res. 2019, 62, 139. [Google Scholar] [CrossRef]
- Mihai, G.; Bîrsan, M.-V.; Dumitrescu, A.; Alexandru, A.; Mirancea, I.; Ivanov, P.; Stuparu, E.; Teodosiu, M.; Daia, M. Adaptive Genetic Potential of European Silver Fir in Romania in the Context of Climate Change. Ann. For. Res. 2018, 61, 95–108. [Google Scholar] [CrossRef]
- NIS National Institute of Statistics. INSEE-Statistics Databases. TEMPO-Online Timeseries; National Institute of Statistics: Bucharest, Romania, 2019. [Google Scholar]
- Mihai, G. O generație avansată de plantaje de brad. An. ICAS 2007, 50, 45–55. [Google Scholar]
- Mohytych, V.; Sułkowska, M.; Klisz, M. Reproduction of Silver Fir (Abies alba Mill) Forests in the Ukrainian Carpathians. Folia For. Pol. 2019, 61, 156–158. [Google Scholar] [CrossRef]
- Woods, J.H. Methods for Estimating Gamete Contributions to Orchard Seed Crops and Vegetative Lots in British Columbia; British Columbia Forest Science Program: Victoria, Canada, 2005. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R Package Version 0.4.6: 2022. Available online: http://florianhartig.github.io/DHARMa/ (accessed on 22 July 2022).
- Lüdecke, D.; Ben-Shachar, M.S.; Patil, I.; Waggoner, P.; Makowski, D. performance: An R Package for Assessment, Comparison and Testing of Statistical Models. J. Open Source Softw. 2021, 6, 3139. [Google Scholar] [CrossRef]
- Campbell, H. The Consequences of Checking for Zero-inflation and Overdispersion in the Analysis of Count Data. Methods Ecol. Evol. 2021, 12, 665–680. [Google Scholar] [CrossRef]
- Blasco-Moreno, A.; Pérez-Casany, M.; Puig, P.; Morante, M.; Castells, E. What does a zero mean? Understanding false, random and structural zeros in ecology. Methods Ecol. Evol. 2019, 10, 949–959. [Google Scholar] [CrossRef]
- Hadfield, J.D. MCMC Methods for Multi-Response Generalized Linear Mixed Models: The MCMCglmm R Package. J. Stat. Soft. 2010, 33, 1–22. [Google Scholar] [CrossRef]
- Banner, K.M.; Irvine, K.M.; Rodhouse, T.J. The Use of Bayesian Priors in Ecology: The Good, the Bad and the Not Great. Methods Ecol. Evol. 2020, 11, 882–889. [Google Scholar] [CrossRef]
- de Villemereuil, P.; Gimenez, O.; Doligez, B. Comparing Parent-Offspring Regression with Frequentist and Bayesian Animal Models to Estimate Heritability in Wild Populations: A Simulation Study for Gaussian and Binary Traits. Methods Ecol. Evol. 2013, 4, 260–275. [Google Scholar] [CrossRef]
- Nikbin, S.; Almasi, F.; Alenizi, D.; Jenvey, C.; Sloan, S.; Preston, S.; Piedrafita, D.; Jonsson, N.; Stear, M. The Heritability of Nematodirus Battus Fecal Egg Counts. Parasitology 2022, 149, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, J. MCMCglmm Course Notes. 2022. Available online: https://mran.microsoft.com/snapshot/2018-08-24/web/packages/MCMCglmm/Vignettes/CourseNotes.Pdf (accessed on 22 July 2022).
- Plummer, M.; Best, N.; Cowles, K.; Vines, K. CODA: Convergence Diagnosis and Output Analysis for MCMC. R News 2006, 6, 7–11. [Google Scholar]
- de Villemereuil, P.; Schielzeth, H.; Nakagawa, S.; Morrissey, M. General Methods for Evolutionary Quantitative Genetic Inference from Generalized Mixed Models. Genetics 2016, 204, 1281–1294. [Google Scholar] [CrossRef]
- Park, J.-M.; Kwon, S.-H.; Lee, H.-J.; Na, S.-J.; El-Kassaby, Y.A.; Kang, K.-S. Integrating Fecundity Variation and Genetic Relatedness in Estimating the Gene Diversity of Seed Crops: Pinus koraiensis Seed Orchard as an Example. Can. J. For. Res. 2017, 47, 366–370. [Google Scholar] [CrossRef]
- Chaisurisri, K.; El-Kassaby, Y.A. Estimation of Clonal Contribution to Cone and Seed Crops in a Sitka Spruce Seed Orchard. Ann. For. Sci. 1993, 50, 461–467. [Google Scholar] [CrossRef][Green Version]
- Kang, K.; Lindgren, D. Fertility Variation and Its Effect on the Relatedness of Seeds in Pinus densiflora, Pinus thunbergii and Pinus koraiensis Clonal Seed Orchards. Silvae Genet. 1998, 47, 196–201. [Google Scholar]
- Varghese, M.; Nicodemus, A.; Nagarajan, B.; Lindgren, D. Impact of Fertility Variation on Gene Diversity and Drift in Two Clonal Seed Orchards of Teak (Tectona grandis Linn. f.). New For. 2006, 31, 497–512. [Google Scholar] [CrossRef]
- Jiao, S.-Q.; Li, M.; Zhu, Y.-J.; Zhou, S.-S.; Zhao, S.-W.; Li, Z.-C.; Bao, Y.-T.; Shi, T.-L.; Zhang, H.-J.; Yang, X.-L.; et al. Variation in Platycladus orientalis (Cupressaceae) Reproductive Output and Its Effect on Seed Orchard Crops’ Genetic Diversity. Forests 2021, 12, 1429. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Kroon, J.; Wennström, U.; Prescher, F.; Lindgren, D.; Mullin, T.J. Estimation of Clonal Variation in Seed Cone Production Over Time in a Scots Pine (Pinus sylvestris L.) Seed Orchard. Silvae Genet. 2009, 58, 53–62. [Google Scholar] [CrossRef][Green Version]
- Li, W.; Wang, X.; Li, Y. Stability in and Correlation Between Factors Influencing Genetic Quality of Seed Lots in Seed Orchard of Pinus tabuliformis Carr. Over a 12-Year Span. PLoS ONE 2011, 6, e23544. [Google Scholar] [CrossRef] [PubMed]
- Gömöry, D.; Bruchanik, R.; Paule, L. Effective Population Number Estimation of Three Scots Pine (Pinus sylvestris L.) Seed Orchards Based on an Integrated Assessment of Flowering, Floral Phenology, and Seed Orchard Design. For. Genet. 2000, 7, 65–75. [Google Scholar]
- Enescu, V. Climate and the Choice of Seed Orchard Sites. For. Ecol. Manag. 1987, 19, 257–265. [Google Scholar] [CrossRef]
- Fernández-Martínez, M.; Vicca, S.; Janssens, I.A.; Espelta, J.M.; Peñuelas, J. The North Atlantic Oscillation Synchronises Fruit Production in Western European Forests. Ecography 2017, 40, 864–874. [Google Scholar] [CrossRef]
- Garcia-Barreda, S.; Sangüesa-Barreda, G.; Madrigal-González, J.; Seijo, F.; González de Andrés, E.; Camarero, J.J. Reproductive Phenology Determines the Linkages Between Radial Growth, Fruit Production and Climate in Four Mediterranean Tree Species. Agric. For. Meteorol. 2021, 307, 108493. [Google Scholar] [CrossRef]
- Byram, T.; Lowe, W.; McGriff, J. Clonal and Annual Variation in Cone Production in Loblolly Pine Seed Orchards. For. Sci. 1986, 32, 1067–1073. [Google Scholar]
- Jonsson, A.; Ekberg, I.; Eriksson, G. Flowering in a Seed Orchard of Pinus Sylvestris L. Studia For. Suec. 1976, 135, 38. [Google Scholar]
- Matziris, D. Variation in Growth, Flowering and Cone Production in a Clonal Seed Orchard of Aleppo Pine Grown in Greece. Silvae Genet. 1997, 46, 224–228. [Google Scholar]
- Kang, K.-S. Genetic Gain and Gene Diversity of Seed Orchard Crops; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2001. [Google Scholar]
- Royauté, R.; Dochtermann, N.A. Comparing Ecological and Evolutionary Variability Within Datasets. Behav. Ecol. Sociobiol. 2021, 75, 127. [Google Scholar] [CrossRef]
- Jayawickrama, K.J.S.; Jett, J.B.; McKeand, S.E. Rootstock Effects in Grafted Conifers: A Review. New For. 1991, 5, 157–173. [Google Scholar] [CrossRef]
- Kamalakannan, R.; Varghese, M.; Park, J.-M.; Kwon, S.-H.; Song, J.-H.; Kang, K.-S. Fertility Variation and Its Impact on Effective Population Size in Seed Stands of Tamarindus indica and Azadirachta indica. Silvae Genet. 2015, 64, 91–99. [Google Scholar] [CrossRef]
- Picarda, N.; Marchi, M.; Serra-Varela, M.J.; Westergren, M.; Cavers, S.; Notivol, E.; Piotti, A.; Alizoti, P.; Bozzano, M.; Santiago, C.; et al. Marginality indices for biodiversity conservation in forest trees. Ecol. Indic. 2022, 143, 109367. [Google Scholar] [CrossRef]
- Mihai, G.; Alexandru, A.M.; Nita, I.A.; Birsan, M.V. Climate Change in the Provenance Regions of Romania over the Last 70 Years: Implications for Forest Management. Forests 2022, 13, 1203. [Google Scholar] [CrossRef]
- Mihai, G.; Alexandru, A.M. Silver Fir Seeds Conservation; Forestry Publishing: Bucharest, Romania, 2021; 65p. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Provenance (Provenance Region) | Clone IDs | Number of Clones | Latitude N | Longitude E | Altitude (m) | Mean Annual Temperature (°C) | Mean Annual Precipitations (mm) |
|---|---|---|---|---|---|---|---|
| Avrig (C1) | 3.1–3.18 | 7 | 45°39′ | 24°29′ | 750 | 7.5 | 694 |
| Rusca Montană (D2) | 3.55–3.72 | 4 | 45°39′ | 24°29′ | 1000 | 6.6 | 878 |
| Sinaia (B2) | 4.17–4.36 | 7 | 45°19′ | 25°32′ | 1150 | 4.8 | 783 |
| Văratec (A2) | 158–181 | 26 | 47°08′ | 26°15′ | 600 | 6.9 | 606 |
| Parameter/Year | 2013 | 2015 | 2018 | 2020 | 2021 | 2022 | Pooled |
|---|---|---|---|---|---|---|---|
| Alive ramets (n) | 636 | 636 | 636 | 636 | 636 | 631 | - |
| Fertile ramets (%) | 75.3 | 95.12 | 91.5 | 94.3 | 42.6 | 67.5 | 77.7 |
| Mean | 28.0 | 67.92 | 79.6 | 47.5 | 7.1 | 16.2 | 41.1 |
| Std. deviation | 35.9 | 41.18 | 61.4 | 36.3 | 15.1 | 21.3 | 46.4 |
| Median | 14.5 | 67.5 | 70.0 | 42.0 | 0.0 | 7.0 | 25.0 |
| Range | 0–206 | 0–200 | 0–350 | 0–215 | 0–115 | 0–110 | 0–350 |
| Parameter/Year | 2013 | 2015 | 2018 | 2020 | 2021 | 2022 |
|---|---|---|---|---|---|---|
| Clone(zi) | 33.2 (17.9–50.4) | 18.9 (0.1–55.6) | 56.8 (36.9–74.6) | 20.2 (0.1–52.6) | 26.4 (13.1–42.7) | 24.6 (10.1–42.8) |
| residual(zi) | 27.3 (20.1–34.2) | 55.4 (30.3–69.9) | 25.9 (15.5–38) | 46 (27.3–58.7) | 31.3 (23.8–38.3) | 37 (27.9–45) |
| Clone(P) | 10.2 (5–18.3) | 5.5 (2.2–10.6) | 2.6 (1–5.4) | 6.2 (2.6–11.7) | 5.7 (1.1–13.4) | 6 (2.3–12) |
| residual(P) | 29.3 (21.2–37.4) | 20.2 (10.9–26.3) | 14.6 (8.5–21.7) | 27.6 (16.3–36) | 36.7 (27.4–46.2) | 32.4 (24–40.2) |
| H2(zi) | 0.127 (0.07, 0.237) | 0 (0, 0.049) | 0.088 (0.033, 0.188) | 0 (0, 0.065) | 0.147 (0.07, 0.229) | 0.106 (0.037, 0.191) |
| H2l(P) | 0.83 (0.723, 0.914) | 0.797 (0.663, 0.891) | 0.733 (0.548, 0.846) | 0.783 (0.616, 0.866) | 0.698 (0.323, 0.868) | 0.724 (0.541, 0.877) |
| H2(P) | 0.607 (0.543, 0.633) | 0.662 (0.536, 0.751) | 0.616 (0.46, 0.715) | 0.614 (0.486, 0.698) | 0.443 (0.188, 0.594) | 0.549 (0.353, 0.628) |
| Year | 2013 | 2015 | 2018 | 2020 | 2021 | 2022 |
|---|---|---|---|---|---|---|
| 2013 | 1 | 0.473 *** | 0.462 * | 0.297 * | 0.276 | 0.001 |
| 2015 | 0.535 *** | 1 | 0.797 *** | 0.560 *** | 0.343 * | −0.003 |
| 2018 | 0.492 ** | 0.811 *** | 1 | 0.588 *** | 0.162 | 0.077 |
| 2020 | 0.346 * | 0.599 *** | 0.596 *** | 1 | −0.069 | 0.163 |
| 2021 | 0.280 | 0.329 * | 0.159 | −0.049 | 1 | 0.136 |
| 2022 | 0.151 | 0.28 | 0.302 | 0.262 | 0.295 | 1.000 |
| Parameter/Year | 2013 | 2015 | 2018 | 2020 | 2021 | 2022 | Pooled |
|---|---|---|---|---|---|---|---|
| CV | 128.24 | 60.63 | 77.16 | 76.44 | 211.96 | 131.06 | 113.00 |
| Ψ(f) | 2.64 | 1.37 | 1.60 | 1.58 | 5.49 | 2.72 | 2.28 |
| Np | 17.02 | 32.90 | 28.21 | 28.40 | 8.19 | 16.56 | 19.76 |
| Nr% | 37.82 | 73.12 | 62.68 | 63.12 | 18.20 | 36.80 | 43.92 |
| GD | 0.971 | 0.985 | 0.982 | 0.982 | 0.939 | 0.970 | 0.975 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teodosiu, M.; Botezatu, A.; Ciocîrlan, E.; Mihai, G. Variation of Cones Production in a Silver Fir (Abies alba Mill.) Clonal Seed Orchard. Forests 2023, 14, 17. https://doi.org/10.3390/f14010017
Teodosiu M, Botezatu A, Ciocîrlan E, Mihai G. Variation of Cones Production in a Silver Fir (Abies alba Mill.) Clonal Seed Orchard. Forests. 2023; 14(1):17. https://doi.org/10.3390/f14010017
Chicago/Turabian StyleTeodosiu, Maria, Anca Botezatu, Elena Ciocîrlan, and Georgeta Mihai. 2023. "Variation of Cones Production in a Silver Fir (Abies alba Mill.) Clonal Seed Orchard" Forests 14, no. 1: 17. https://doi.org/10.3390/f14010017
APA StyleTeodosiu, M., Botezatu, A., Ciocîrlan, E., & Mihai, G. (2023). Variation of Cones Production in a Silver Fir (Abies alba Mill.) Clonal Seed Orchard. Forests, 14(1), 17. https://doi.org/10.3390/f14010017