
Spatial Variations in Fine Root Turnover, Biomass, and Necromass of Two Vegetation Types in a Karst Ecosystem, Southwestern China




Abstract
:1. Introduction
2. Material and Methods
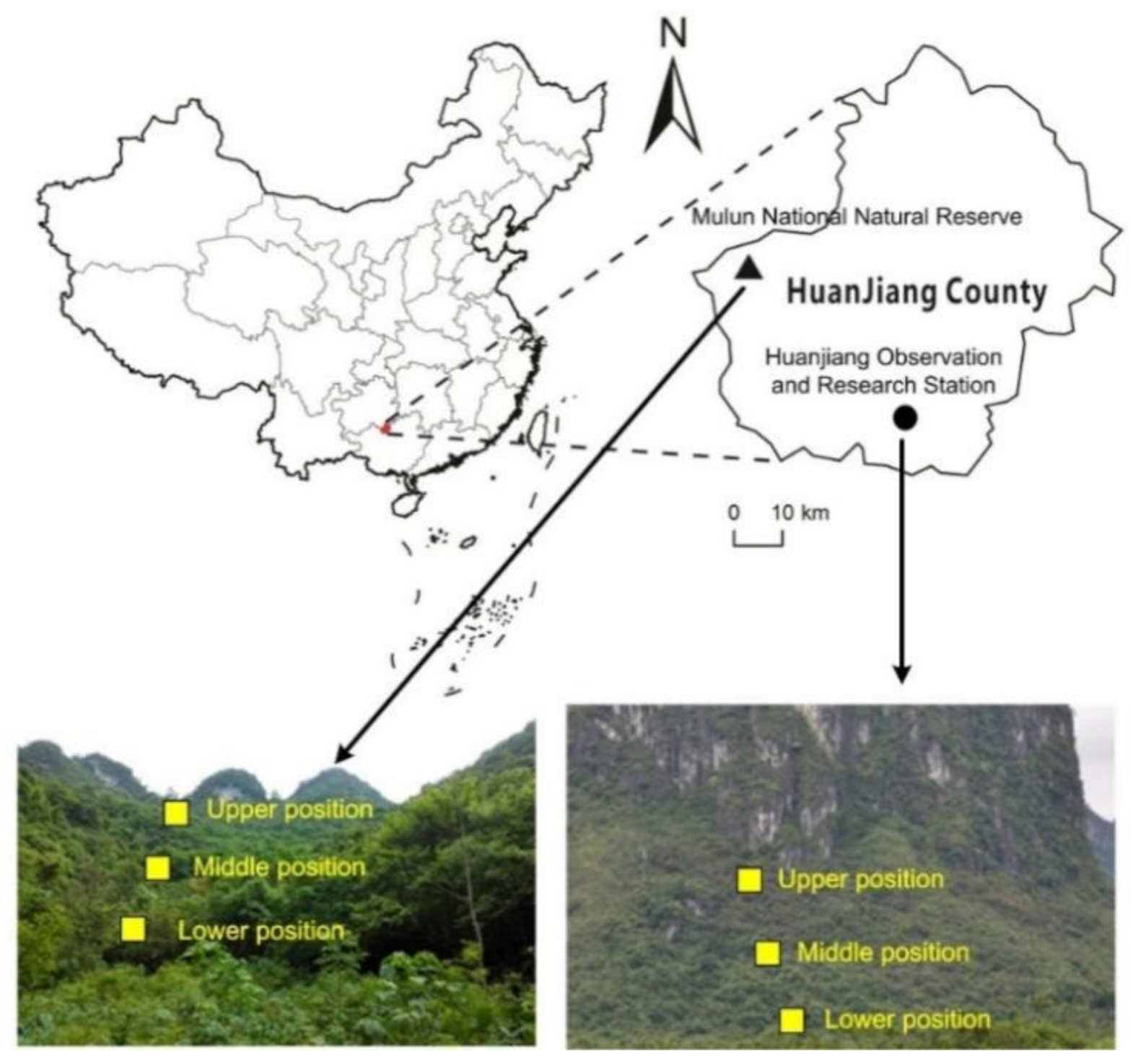
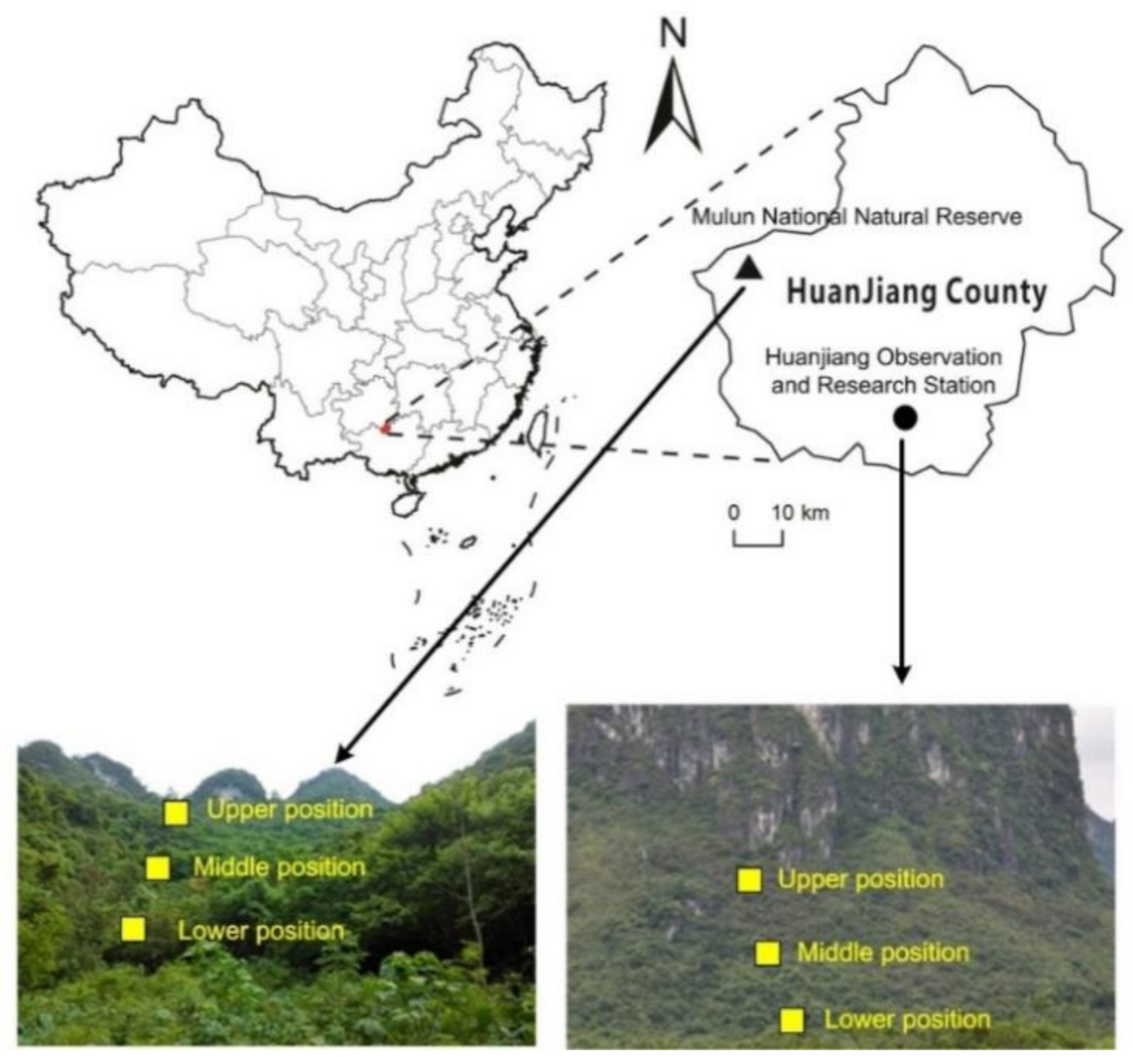
2.1. Study Site
2.2. Sampling
- (1)
- The sequential coring
- (2)
- The ingrowth cores
2.3. Laboratory Analyses
2.4. Calculation of Turnover Rates
2.5. Statistical Analyses
3. Results
3.1. Characteristics of Soil Nutrients
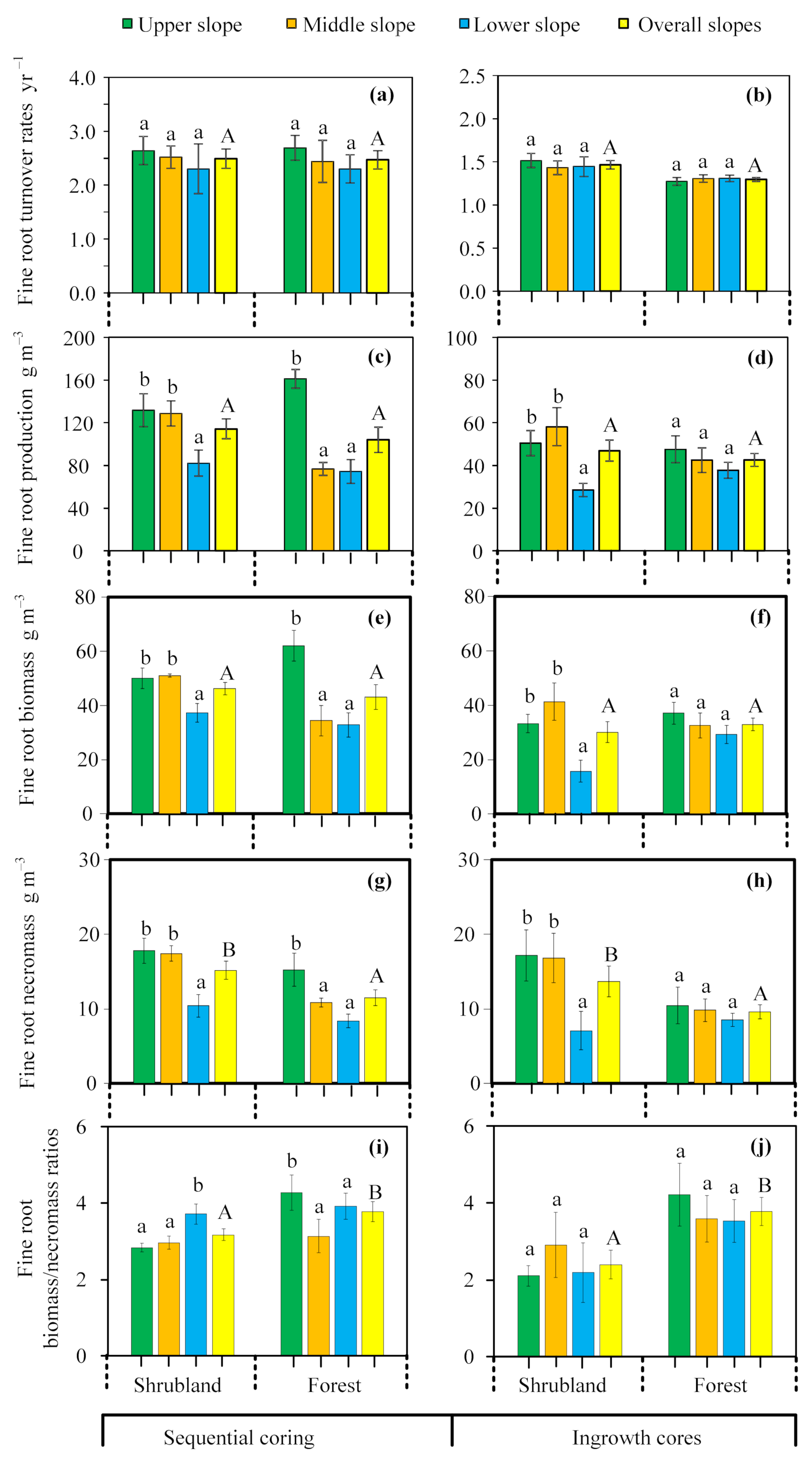
3.2. Spatial Patterns in Fine Root Turnover Rate and Related Parameters
3.3. Spatial Patterns of Living Fine Root Nutrients
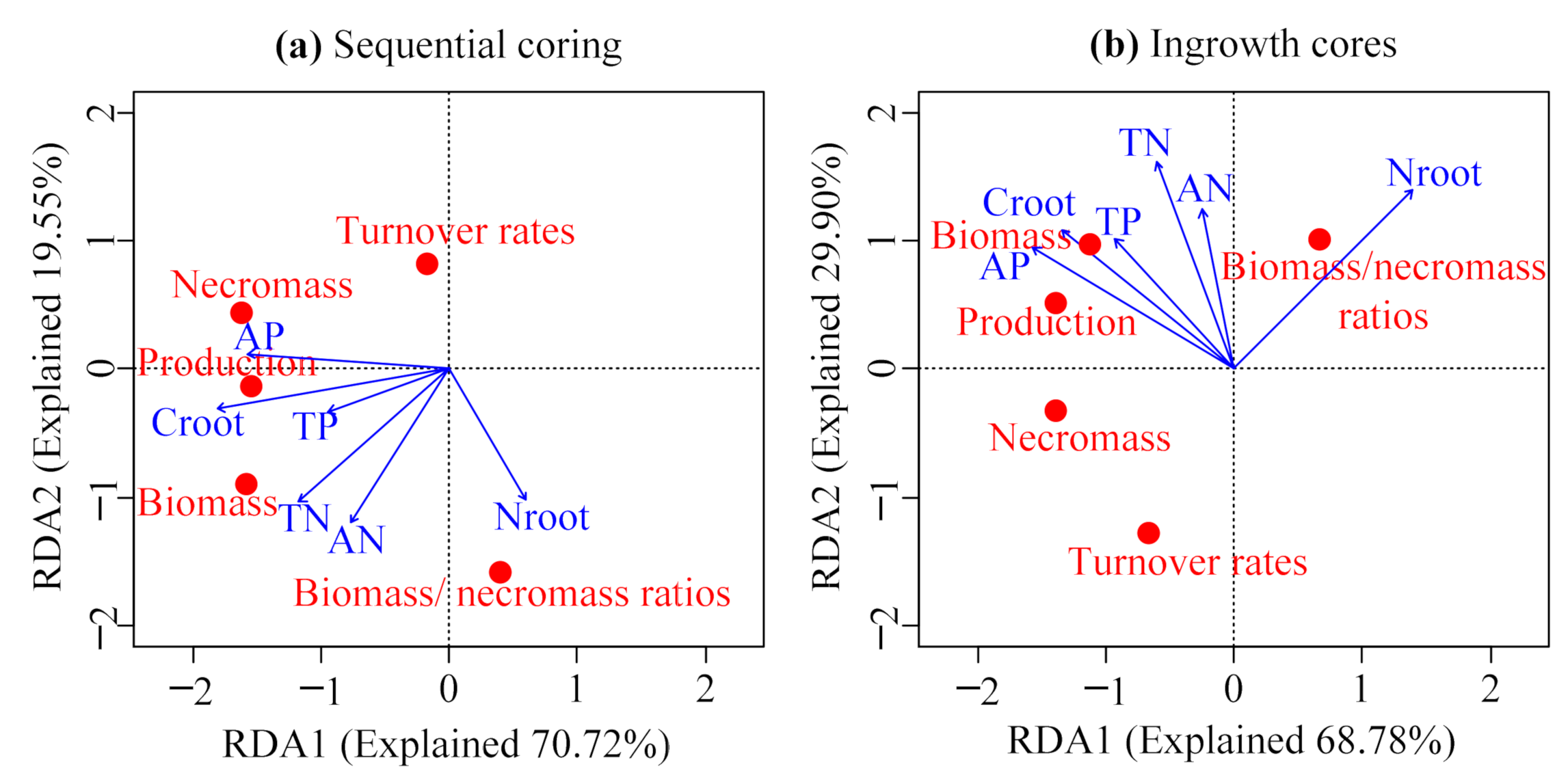
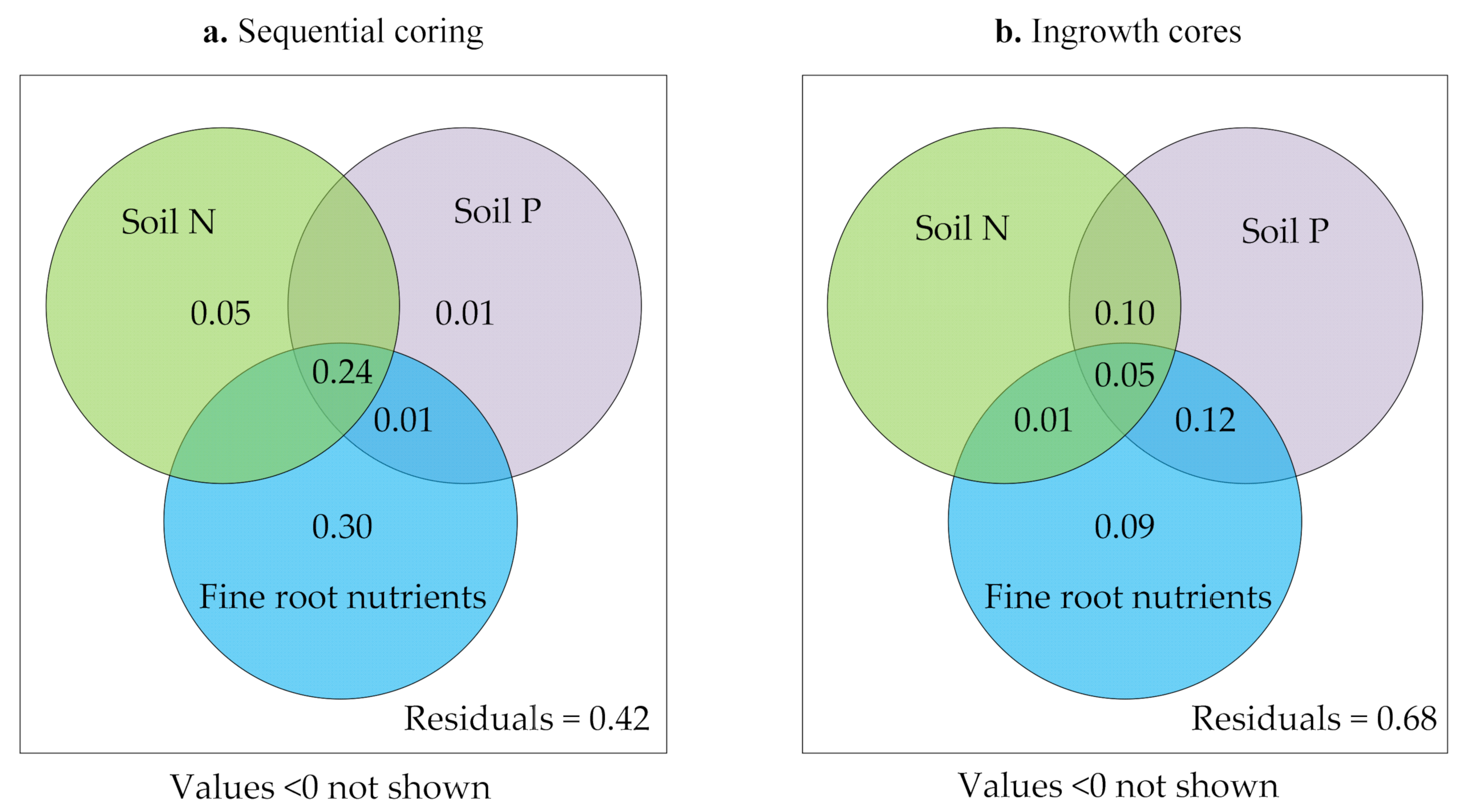
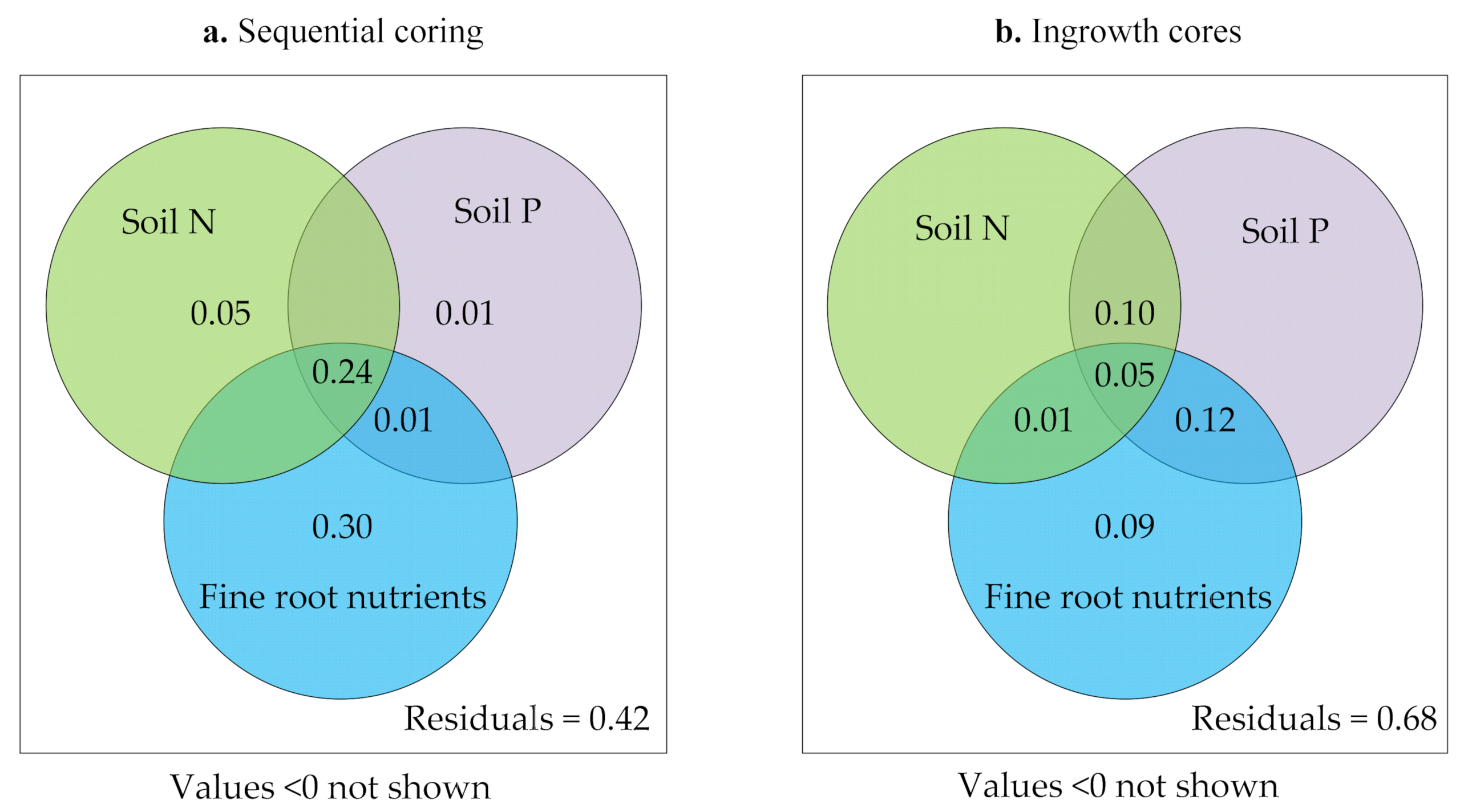
3.4. Relationships between Fine Roots and Soil Characteristics
4. Discussion
4.1. Fine Root Turnover Rates in Karst Ecosystems
4.2. Variations in Fine Root Turnover with Changes in Slope Positions and Vegetation Types
4.3. Fine Root Turnover and Dynamics Related to Soil and Root Nutrients
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuan, D.X. On the karst ecosystem. Acta Geol. Sin.-Engl. Ed. 2001, 75, 336–338. [Google Scholar]
- Pan, F.J.; Zhang, W.; Liang, Y.M.; Liu, S.J.; Wang, K.L. Increased associated effects of topography and litter and soil nutrients on soil enzyme activities and microbial biomass along vegetation successions in karst ecosystem, southwestern China. Environ. Sci. Pollut. Res. 2018, 25, 16979–16990. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.J.; Liu, Q.M.; Zhang, D.F. Karst rocky desertification in southwestern China: Geomorphology, landuse, impact and rehabilitation. Land Degrad. Dev. 2004, 15, 115–121. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, W.; Wang, K.; Hou, Y. Soil organic carbon and total nitrogen as affected by land use types in karst and non-karst areas of northwest Guangxi, China. J. Sci. Food Agric. 2012, 92, 1086–1093. [Google Scholar] [CrossRef]
- Qi, X.; Wang, K.; Zhang, C. Effectiveness of ecological restoration projects in a karst region of southwest China assessed using vegetation succession mapping. Ecol. Eng. 2013, 54, 245–253. [Google Scholar] [CrossRef]
- Jiang, Z.C.; Luo, W.Q.; Tong, L.Q.; Cheng, Y.; Yang, L.Y.; Wu, Z.Y. Evolution features of rocky desertification and influence factors in karst areas of southwest China in the 21st century. Carsologica Sin. 2016, 35, 461–468. (In Chinese) [Google Scholar]
- Wang, K.L.; Yue, Y.M.; Chen, H.S.; Wu, X.B.; Xiao, J.; Qi, X.K.; Zhang, W.; Du, H. The comprehensive treatment of karst rocky desertification and its regional restoration effects. Acta Ecol. Sin. 2019, 39, 7432–7440. (In Chinese) [Google Scholar]
- Liang, Y.; He, X.; Chen, C.; Feng, S.; Liu, L.; Chen, X.; Zhao, Z.; Su, Y. Influence of plant communities and soil properties during natural vegetation restoration on arbuscular mycorrhizal fungal communities in a karst region. Ecol. Eng. 2015, 82, 57–65. [Google Scholar] [CrossRef]
- Pan, F.; Zhang, W.; Liu, S.; Li, D.; Wang, K. Leaf N:P stoichiometry across plant functional groups in the karst region of southwestern China. Trees 2015, 29, 883–892. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, J.; Pan, F.; Li, D.; Chen, H.; Wang, K. Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in southwest China. Plant Soil 2015, 391, 77–91. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, W.; Wang, K.; Pan, F.; Yang, S.; Shu, S. Factors controlling accumulation of soil organic carbon along vegetation succession in a typical karst region in Southwest China. Sci. Total Environ. 2015, 521–522, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; He, T.; Chen, H.; Peng, W.; Song, T.; Wang, K.; Li, D. Impacts of vegetation restoration strategies on soil organic carbon and nitrogen dynamics in a karst area, southwest China. Ecol. Eng. 2017, 101, 247–254. [Google Scholar] [CrossRef]
- Thorley, R.M.S.; Taylor, L.; Banwart, S.; Leake, J.; Beerling, D. The role of forest trees and their mycorrhizal fungi in carbonate rock weathering and its significance for global carbon cycling. Plant Cell Environ. 2014, 38, 1947–1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; He, X.; Wang, K.; Xie, Y.; Xie, Q.; O’Donnell, A.G.; Chen, C. The Bradyrhizobium-legume symbiosis is dominant in the shrubby ecosystem of the Karst region, Southwest China. Eur. J. Soil Biol. 2015, 68, 1–8. [Google Scholar] [CrossRef]
- Clarholm, M.; Skyllberg, U.; Rosling, A. Organic acid induced release of nutrients from metal-stabilized soil organic matter—The unbutton model. Soil Biol. Biochem. 2015, 84, 168–176. [Google Scholar] [CrossRef]
- Pan, F.; Liang, Y.; Zhang, W.; Zhao, J.; Wang, K. Enhanced Nitrogen Availability in Karst Ecosystems by Oxalic Acid Release in the Rhizosphere. Front. Plant Sci. 2016, 7, 687. [Google Scholar] [CrossRef] [Green Version]
- Pan, F.; Liang, Y.; Wang, K.; Zhang, W. Responses of Fine Root Functional Traits to Soil Nutrient Limitations in a Karst Ecosystem of Southwest China. Forests 2018, 9, 743. [Google Scholar] [CrossRef] [Green Version]
- Wurzburger, N.; Wright, S.J. Fine-root responses to fertilization reveal multiple nutrient limitation in a lowland tropical forest. Ecology 2015, 96, 2137–2146. [Google Scholar] [CrossRef] [Green Version]
- Finér, L.; Helmisaari, H.S.; Lõhmus, K.; Majdi, H.; Brunner, I.; Børja, I.; Eldhuset, T.; Godbold, D.; Grebenc, T.; Konôpka, B.; et al. Variation in fine root biomass of three European tree species: Beech (Fagus sylvatica L.), Norway spruce (Picea abies L. Karst.), and Scots pine (Pinus sylvestris L.). Plant Biosyst. 2007, 141, 394–405. [Google Scholar] [CrossRef]
- Matamala, R.; Stover, D. Introduction to a Virtual Special Issue: Modeling the hidden half—the root of our problem. New Phytol. 2013, 200, 939–942. [Google Scholar] [CrossRef]
- Artacho, P.; Bonomelli, C. Changes in fine-root production, phenology and spatial distribution in response to N application in irrigated sweet cherry trees. Tree Physiol. 2016, 36, 601–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vormstein, S.; Kaiser, M.; Piepho, H.P.; Joergensen, R.G.; Ludwig, B. Effects of fine root characteristics of beech on carbon turnover in the topsoil and subsoil of a sandy Cambisol. Eur. J. Soil Sci. 2017, 68, 177–188. [Google Scholar] [CrossRef]
- Persson, H.Å.; Stadenberg, I. Spatial distribution of fine-roots in boreal forests in eastern Sweden. Plant Soil 2009, 318, 1–14. [Google Scholar] [CrossRef]
- Bowsher, A.W.; Mason, C.M.; Goolsby, E.W.; Donovan, L.A. Fine root tradeoffs between nitrogen concentration and xylem vessel traits preclude unified whole-plant resource strategies in Helianthus. Ecol. Evol. 2016, 6, 1016–1031. [Google Scholar] [CrossRef] [PubMed]
- Majdi, H.; Pregitzer, K.; Morén, A.S.; Nylund, J.E.; Ågren, G.I. Measuring Fine Root Turnover in Forest Ecosystems. Plant Soil 2005, 276, 1–8. [Google Scholar] [CrossRef]
- Ostonen, I.; Lõhmus, K.; Pajuste, K. Fine root biomass, production and its proportion of NPP in a fertile middle-aged Norway spruce forest: Comparison of soil core and ingrowth core methods. For. Ecol. Manag. 2005, 212, 264–277. [Google Scholar] [CrossRef]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and Associated Fungi Drive Long-Term Carbon Sequestration in Boreal Forest. Science 2013, 339, 1615–1618. [Google Scholar] [CrossRef]
- Brunner, I.; Bakker, M.; Björk, R.; Hirano, Y.; Lukac, M.; Aranda, X.; Børja, I.; Eldhuset, T.D.; Helmisaari, H.S.; Jourdan, C.; et al. Fine-root turnover rates of European forests revisited: An analysis of data from sequential coring and ingrowth cores. Plant Soil 2013, 362, 357–372. [Google Scholar] [CrossRef]
- Matamala, R.; Gonzaàlez-Meler, M.A.; Jastrow, J.D.; Norby, R.J.; Schlesinger, W.H. Impacts of Fine Root Turnover on Forest NPP and Soil C Sequestration Potential. Science 2003, 302, 1385–1387. [Google Scholar] [CrossRef]
- Quinto, H.; Caicedo, H.; Perez, M.T.; Moreno, F. Fine root dynamics and its relationship with soil fertility in tropical rainforests of Chocó. Rev. Biol. Trop. 2016, 64, 1709–1719. [Google Scholar]
- Huang, G.; Zhao, X.Y.; Su, Y.G.; Zhao, H.L.; Zhang, T.H. Vertical distribution, biomass, production and turnover of fine roots along a topographical gradient in a sandy shrubland. Plant Soil 2008, 308, 201–212. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, K.; Chen, H.; He, X.; Zhang, J. Ancillary information improves kriging on soil organic carbon data for a typical karst peak cluster depression landscape. J. Sci. Food Agric. 2012, 92, 1094–1102. [Google Scholar] [CrossRef]
- Liu, J.; Qiu, L.; Ao, G.; Zheng, B.; Wei, X.; Zhang, Y.; Gao, H.; Cheng, J. Effect of clipping on aboveground biomass and nutrients varies with slope position but not with slope aspect in a hilly semiarid restored grassland. Ecol. Eng. 2019, 134, 47–55. [Google Scholar] [CrossRef]
- Wei, X.; Shao, M.; Fu, X.; Horton, R. Changes in soil organic carbon and total nitrogen after 28 years grassland afforestation: Effects of tree species, slope position, and soil order. Plant Soil 2010, 331, 165–179. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, P.; Hu, Y.; Zhao, Y. Vegetation succession and soil infiltration characteristics under different aged refuse dumps at the Heidaigou opencast coal mine. Glob. Ecol. Conserv. 2015, 4, 255–263. [Google Scholar] [CrossRef] [Green Version]
- van Hall, R.L.; Cammeraat, E.; Keesstra, S.D.; Zorn, M. Impact of secondary vegetation succession on soil quality in a humid Mediterranean landscape. Catena 2017, 149, 836–843. [Google Scholar] [CrossRef]
- Freschet, G.T.; Valverde-Barrantes, O.J.; Tucker, C.M.; Craine, J.M.; McCormack, M.L.; Violle, C.; Fort, F.; Blackwood, C.; Urban-Mead, K.R.; Iversen, C.M.; et al. Climate, soil and plant functional types as drivers of global fine-root trait variation. J. Ecol. 2017, 105, 1182–1196. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Chen, H.Y. Positive species mixture effects on fine root turnover and mortality in natural boreal forests. Soil Biol. Biochem. 2018, 121, 130–137. [Google Scholar] [CrossRef]
- Grassein, F.; Lemauviel-Lavenant, S.; Lavorel, S.; Bahn, M.; Bardgett, R.D.; Desclos-Theveniau, M.; Laîné, P. Relationships between functional traits and inorganic nitrogen acquisition among eight contrasting European grass species. Ann. Bot. 2015, 115, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Valverde-Barrantes, O.J.; Smemo, K.A.; Blackwood, C. Fine root morphology is phylogenetically structured, but nitrogen is related to the plant economics spectrum in temperate trees. Funct. Ecol. 2015, 29, 796–807. [Google Scholar] [CrossRef]
- Cornejo, N.S.; Hertel, D.; Becker, J.N.; Hemp, A.; Leuschner, C. Biomass, Morphology, and Dynamics of the Fine Root System Across a 3000-M Elevation Gradient on Mt. Kilimanjaro. Front. Plant Sci. 2020, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulmatiski, A.; Beard, K.H.; Norton, J.M.; Heavilin, J.E.; Forero, L.E.; Grenzer, J. Live long and prosper: Plant-soil feedback, lifespan, and landscape abundance covary. Ecology 2017, 98, 3063–3073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Zhang, W.; Wang, K.; Fu, W. Soil moisture dynamics under different land uses on karst hillslope in northwest Guangxi, China. Environ. Earth Sci. 2010, 61, 1105–1111. [Google Scholar] [CrossRef]
- Nie, Y.P.; Chen, H.S.; Wang, K.L.; Yang, J. Water source utilization by woody plants growing on dolomite outcrops and nearby soils during dry seasons in karst region of Southwest China. J. Hydrol. 2012, 420–421, 264–274. [Google Scholar] [CrossRef]
- Zongming, S.; Xiankun, L. The types of natural vegetation in karst region of Guangxi and its classified system. Guangxi Zhiwu 2003, 23, 289–293. (In Chinese) [Google Scholar]
- Fairley, R.I.; Alexander, I.J. Methods of calculating fine root production in forests. In Ecological Interactions in Soil: Plants, Microbes and Animals; Fitter, A.H., Atkinson, D., Read, D.J., Eds.; Blackwell: Oxford, UK, 1985; pp. 37–42. [Google Scholar]
- Burke, M.K.; Raynal, D.J. Fine root growth phenology, production, and turnover in a northern hardwood forest ecosystem. Plant Soil 1994, 162, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Montagnoli, A.; Terzaghi, M.; Di Iorio, A.; Scippa, G.; Chiatante, D. Fine-root seasonal pattern, production and turnover rate of European beech (Fagus sylvatica L.) stands in Italy Prealps: Possible implications of coppice conversion to high forest. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2012, 146, 1012–1022. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Lin, S.; Reinsch, T.; Loges, R.; Hasler, M.; Taube, F. Comparison of ingrowth core and sequential soil core methods for estimating belowground net primary production in grass-clover swards. Grass Forage Sci. 2015, 71, 515–528. [Google Scholar] [CrossRef]
- Fu, Z.; Chen, H.; Xu, Q.; Jia, J.; Wang, S.; Wang, K. Role of epikarst in near-surface hydrological processes in a soil mantled subtropical dolomite karst slope: Implications of field rainfall simulation experiments. Hydrol. Process. 2016, 30, 795–811. [Google Scholar] [CrossRef]
- Song, M.; Peng, W.; Zeng, F.; Du, H.; Peng, Q.; Xu, Q.; Chen, L.; Zhang, F. Spatial Patterns and Drivers of Microbial Taxa in a Karst Broadleaf Forest. Front. Microbiol. 2018, 9, 1691. [Google Scholar] [CrossRef]
- Peng, W.; Zhu, Y.; Song, M.; Du, H.; Song, T.; Zeng, F.; Zhang, F.; Wang, K.; Luo, Y.; Lan, X.; et al. The spatial distribution and drivers of soil microbial richness and diversity in a karst broadleaf forest. For. Ecol. Manag. 2019, 449, 117241. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: https://www.R-project.org/ (accessed on 20 January 2022).
- Abbas, S.; Nichol, J.E.; Zhang, J.; Fischer, G.A. The accumulation of species and recovery of species composition along a 70 year succession in a tropical secondary forest. Ecol. Indic. 2019, 106, 105524. [Google Scholar] [CrossRef]
- Gill, R.A.; Jackson, R.B. Global patterns of root turnover for terrestrial ecosystems. New Phytol. 2000, 147, 13–31. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Z.; Gu, J. Variation patterns of fine root biomass, production and turnover in Chinese forests. J. For. Res. 2017, 28, 1185–1194. [Google Scholar] [CrossRef]
- Hendricks, J.J.; Hendrick, R.L.; Wilson, C.A.; Mitchell, R.J.; Pecot, S.D.; Guo, D. Assessing the patterns and controls of fine root dynamics: An empirical test and methodological review. J. Ecol. 2006, 94, 40–57. [Google Scholar] [CrossRef]
- Steele, S.J.; Gower, S.T.; Vogel, J.G.; Norman, J.M. Root mass, net primary production and turnover in aspen, jack pine and black spruce forests in Saskatchewan and Manitoba, Canada. Tree Physiol. 1997, 17, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Hansson, K.; Helmisaari, H.S.; Sah, S.P.; Lange, H. Fine root production and turnover of tree and understorey vegetation in Scots pine, silver birch and Norway spruce stands in SW Sweden. For. Ecol. Manag. 2013, 309, 58–65. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Fine root dynamics with stand development in the boreal forest. Funct. Ecol. 2012, 26, 991–998. [Google Scholar] [CrossRef]
- Sun, T.; Dong, L.; Mao, Z.; Li, Y. Fine root dynamics of trees and understorey vegetation in a chronosequence of Betula platyphylla stands. For. Ecol. Manag. 2015, 346, 1–9. [Google Scholar] [CrossRef]
- Du, H.; Liu, L.; Su, L.; Zeng, F.; Wang, K.; Peng, W.; Zhang, H.; Song, T. Seasonal Changes and Vertical Distribution of Fine Root Biomass during Vegetation Restoration in a Karst Area, Southwest China. Front. Plant Sci. 2019, 9, 2001. [Google Scholar] [CrossRef] [Green Version]
- Kochsiek, A.; Tan, S.; Russo, S.E. Fine root dynamics in relation to nutrients in oligotrophic Bornean rain forest soils. Plant Ecol. 2013, 214, 869–882. [Google Scholar] [CrossRef]
- Pinno, B.D.; Wilson, S.D. Fine root response to soil resource heterogeneity differs between grassland and forest. Plant Ecol. 2013, 214, 821–829. [Google Scholar] [CrossRef]
- Göransson, H.; Edwards, P.J.; Perreijn, K.; Smittenberg, R.; Venterink, H.O. Rocks create nitrogen hotspots and N:P heterogeneity by funnelling rain. Biogeochemistry 2014, 121, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.B.; Bai, X.; He, X.B. Soil creeping in the weathering crust of carbonate rocks and underground soil losses in the karst mountain areas of southwest china. Carbonates Evaporites 2011, 26, 149–153. [Google Scholar] [CrossRef]
- Espeleta, J.F.; Donovan, L.A. Fine root demography and morphology in response to soil resources availability among xeric and mesicsandhill tree species. Funct. Ecol. 2002, 16, 113–121. [Google Scholar] [CrossRef]
- Wang, G.; Xue, S.; Liu, F.; Liu, G. Nitrogen addition increases the production and turnover of the lower-order roots but not of the higher-order roots of Bothriochloa ischaemum. Plant Soil 2017, 415, 423–434. [Google Scholar] [CrossRef]
- Hu, L.; Su, Y.; He, X.; Wu, J.; Zheng, H.; Li, Y.; Wang, A. Response of soil organic carbon mineralization in typical Karst soils following the addition of 14C-labeled rice straw and CaCO3. J. Sci. Food Agric. 2012, 92, 1112–1118. [Google Scholar] [CrossRef]
- Li, X.F.; Han, S.J.; Zhang, Y. Decomposition of leaf litter of quercus mongolica from four precipitation gradients in mongolian oak forest. Chin. J. Appl. Ecol. 2007, 18, 261–266. (In Chinese) [Google Scholar]
- Pan, F.J.; Zhang, W.; Wang, K.L.; He, X.Y.; Liang, S.C.; Wei, G.F. Litter C:N:P ecological stoichiometry character of plant communities in typical karst peak-cluster depression. Acta Ecol. Sin. 2011, 31, 335–343. (In Chinese) [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetations | Number | Altitude (m) | Slope (°) | Community Height (m) | Dominant Species |
|---|---|---|---|---|---|
| Shrubland | 15 | 280 ~ 420 | 23.2 ± 5.8 | 2.5 ± 0.2 | Vitex negundo, Croton lachnocarpus, Ligustrum sinense, Cipadessa cinerascens, Platycarya longipes |
| Forest | 15 | 320 ~ 480 | 35.3 ± 6.1 | 7.4 ± 0.5 | Pteroceltis tatarinowii, Rapanea kwangsiensis, Eurycorymbus cavaleriei, Murraya exotica, Cipadessa cinerascens |
| Slope Positions | Total Nitrogen (TN g kg−1) | Available Nitrogen (AN mg kg−1) | Total Phosphorus (TP g kg−1) | Available Phosphorus (AP mg kg−1) |
|---|---|---|---|---|
| Shrubland | ||||
| Upper | 7.86 ± 0.22 b | 424.92 ± 4.69 b | 1.48 ± 0.20 b | 12.18 ± 0.69 b |
| Middle | 6.75 ± 0.46 b | 388.43 ± 16.25 b | 1.46 ± 0.10 b | 11.30 ± 0.46 b |
| Lower | 4.34 ± 0.24 a | 358.33 ± 23.93 a | 0.96 ± 0.08 a | 4.63 ± 1.14 a |
| Overall | 6.32 ± 0.43 A | 390.56 ± 11.61 A | 1.30 ± 0.10 A | 9.37 ± 1.00 A |
| Forest | ||||
| Upper | 13.54 ± 1.39 b | 584.23 ± 63.21 b | 1.64 ± 0.07 b | 10.50 ± 1.42 b |
| Middle | 9.63 ± 2.43 b | 508.15 ± 90.28 b | 1.66 ± 0.21 b | 9.52 ± 1.67 b |
| Lower | 4.70 ± 0.17 a | 314.68 ± 9.14 a | 1.14 ± 0.10 a | 4.32 ± 0.32 a |
| Overall | 9.29 ± 1.30 B | 469.02 ± 45.65 B | 1.48 ± 0.10 A | 8.11 ± 1.00 A |
| Response Variable | Slope Position | Vegetation | Slope Position × Vegetation | |||
|---|---|---|---|---|---|---|
| F Value | p | F Value | p | F Value | p | |
| Sequential coring | ||||||
| Fine root turnover rates | 1.452 | 0.239 | 0.003 | 0.957 | 0.007 | 0.935 |
| Fine root production | 26.948 | <0.001 | 0.888 | 0.355 | 2.009 | 0.168 |
| Fine root biomass | 19.675 | <0.001 | 0.720 | 0.404 | 2.911 | 0.099 |
| Fine root necromass | 22.874 | <0.001 | 10.248 | 0.004 | 0.076 | 0.786 |
| Biomass/necromass ratios | 0.473 | 0.498 | 4.540 | 0.042 | 3.182 | 0.086 |
| Ingrowth cores | ||||||
| Fine root turnover rates | 0.129 | 0.722 | 9.811 | 0.004 | 0.633 | 0.434 |
| Fine root production | 5.094 | 0.033 | 0.496 | 0.488 | 0.637 | 0.432 |
| Fine root biomass | 4.147 | 0.052 | 0.074 | 0.788 | 0.215 | 0.647 |
| Fine root necromass | 4.038 | 0.055 | 5.733 | 0.025 | 1.465 | 0.237 |
| Biomass/necromass ratios | 0.001 | 0.977 | 5.280 | 0.030 | 1.087 | 0.307 |
| Response Variable | Slope Position | Vegetation | Slope Position × Vegetation | |||
|---|---|---|---|---|---|---|
| F Value | p | F Value | p | F Value | p | |
| Sequential coring | ||||||
| Fine root C concentration | 28.194 | <0.001 | 0.238 | 0.626 | 2.923 | 0.056 |
| Fine root N concentration | 2.009 | 0.137 | 217.471 | <0.001 | 11.936 | <0.001 |
| Fine root C:N ratios | 1.639 | 0.197 | 175.577 | <0.001 | 9.883 | <0.001 |
| Ingrowth cores | ||||||
| Fine root C concentration | 11.446 | <0.001 | 1.666 | 0.210 | 2.525 | 0.102 |
| Fine root N concentration | 1.494 | 0.245 | 20.277 | <0.001 | 11.764 | <0.001 |
| Fine root C:N ratios | 0.190 | 0.828 | 22.35 | <0.001 | 14.35 | <0.001 |
| Production | Biomass | Necromass | Biomass/Necromass Ratios | |
|---|---|---|---|---|
| Sequential coring | ||||
| Turnover rates | 0.492 ** | −0.145 | 0.014 | −0.233 |
| Production | – | 0.768 ** | 0.658 ** | 0.051 |
| Biomass | – | – | 0.739 ** | 0.230 |
| Necromass | – | – | – | −0.454 * |
| Ingrowth cores | ||||
| Turnover rates | 0.191 | −0.141 | 0.716 ** | −0.862 ** |
| Production | – | 0.941 ** | 0.806 ** | −0.223 |
| Biomass | – | – | 0.558 ** | 0.063 |
| Necromass | – | – | – | −0.659 ** |
| Soil TN | Soil AN | Soil TP | Soil AP | Croot | Nroot | C/N Ratios | |
|---|---|---|---|---|---|---|---|
| Sequential coring | |||||||
| Turnover rates | −0.110 | −0.230 | 0.290 | 0.109 | 0.182 | 0.024 | −0.011 |
| Production | 0.439 * | 0.263 | 0.344 | 0.530 ** | 0.712 ** | −0.041 | 0.188 |
| Biomass | 0.612 ** | 0.512 ** | 0.393 * | 0.542 ** | 0.684 ** | −0.040 | 0.197 |
| Necromass | 0.318 | 0.173 | 0.315 | 0.590 ** | 0.570 ** | −0.432 * | 0.572 ** |
| Biomass: Necromass ratios | 0.283 | 0.336 | 0.068 | −0.165 | 0.033 | 0.511 ** | −0.504 ** |
| Ingrowth cores | |||||||
| Turnover rates | −0.028 | −0.002 | 0.033 | 0.044 | −0.095 | −0.51 ** | 0.479 ** |
| Production | 0.381 * | 0.252 | 0.376 * | 0.506 ** | 0.503 ** | −0.226 | 0.405 * |
| Biomass | 0.468 ** | 0.324 | 0.392 * | 0.503 ** | 0.540 ** | −0.049 | 0.237 |
| Necromass | 0.120 | 0.052 | 0.250 | 0.381 * | 0.290 | −0.468 * | 0.580 ** |
| Biomass: Necromass ratios | 0.340 | 0.336 | 0.186 | 0.045 | 0.020 | 0.514 ** | −0.482 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, F.; Qian, Q.; Liang, Y.; Wang, K.; Zhang, W. Spatial Variations in Fine Root Turnover, Biomass, and Necromass of Two Vegetation Types in a Karst Ecosystem, Southwestern China. Forests 2022, 13, 611. https://doi.org/10.3390/f13040611
Pan F, Qian Q, Liang Y, Wang K, Zhang W. Spatial Variations in Fine Root Turnover, Biomass, and Necromass of Two Vegetation Types in a Karst Ecosystem, Southwestern China. Forests. 2022; 13(4):611. https://doi.org/10.3390/f13040611
Chicago/Turabian StylePan, Fujing, Qian Qian, Yueming Liang, Kelin Wang, and Wei Zhang. 2022. "Spatial Variations in Fine Root Turnover, Biomass, and Necromass of Two Vegetation Types in a Karst Ecosystem, Southwestern China" Forests 13, no. 4: 611. https://doi.org/10.3390/f13040611
APA StylePan, F., Qian, Q., Liang, Y., Wang, K., & Zhang, W. (2022). Spatial Variations in Fine Root Turnover, Biomass, and Necromass of Two Vegetation Types in a Karst Ecosystem, Southwestern China. Forests, 13(4), 611. https://doi.org/10.3390/f13040611