Post-Fire Habitat Heterogeneity Leads to Black Spruce–Kalmia L. Shrub Savannah Alternate State


Abstract
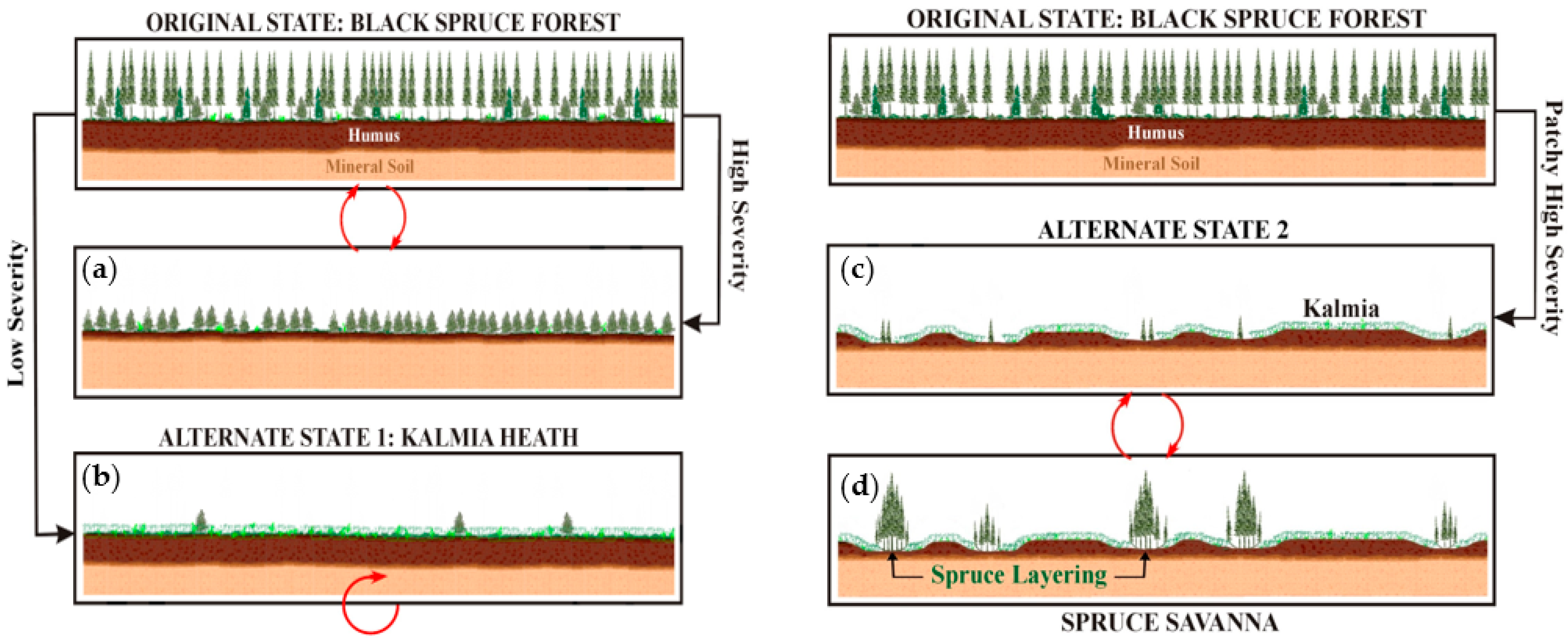
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Site
2.2.1. Black Spruce Stem Density and Growth
2.2.2. Foliar Nutrient Analysis
2.2.3. Soil Characteristics
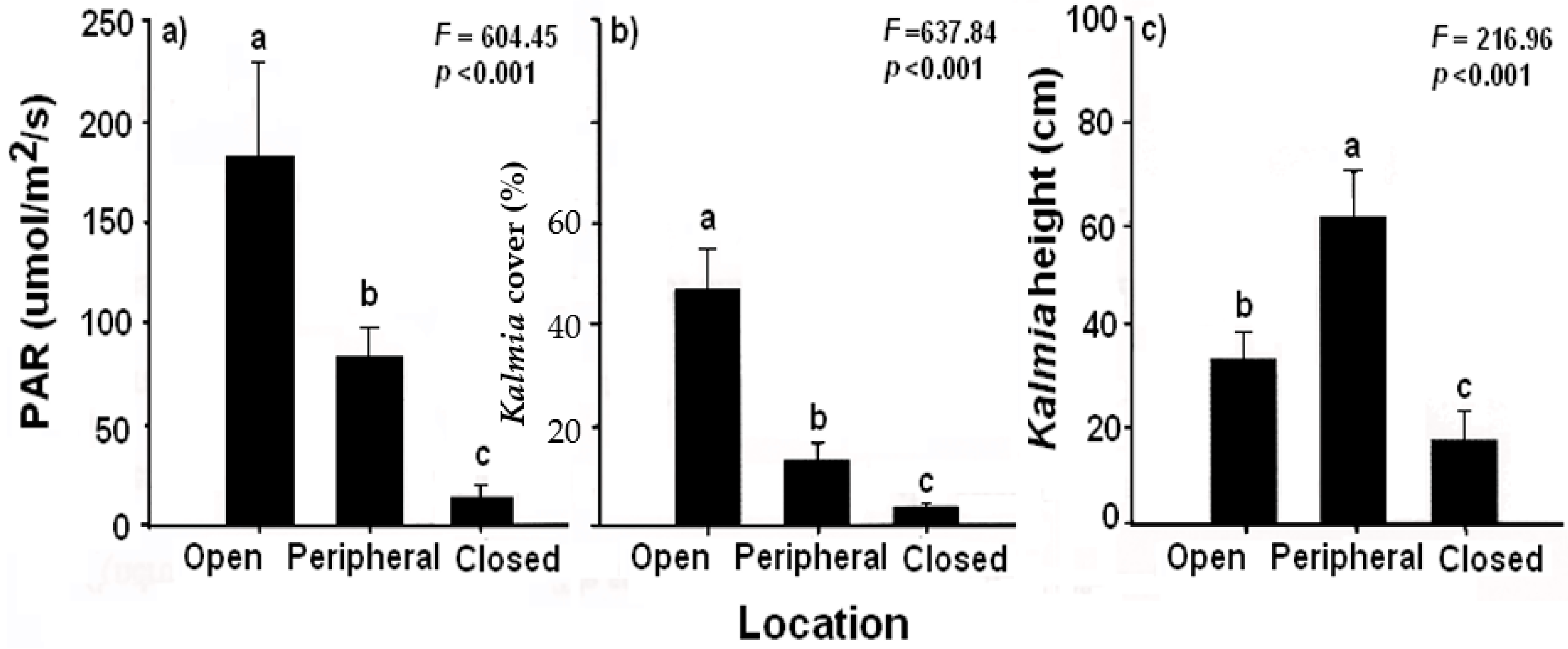
2.2.4. Black Spruce Shade Effects on Kalmia
2.2.5. Community Composition
2.3. Statistical Analysis
3. Results
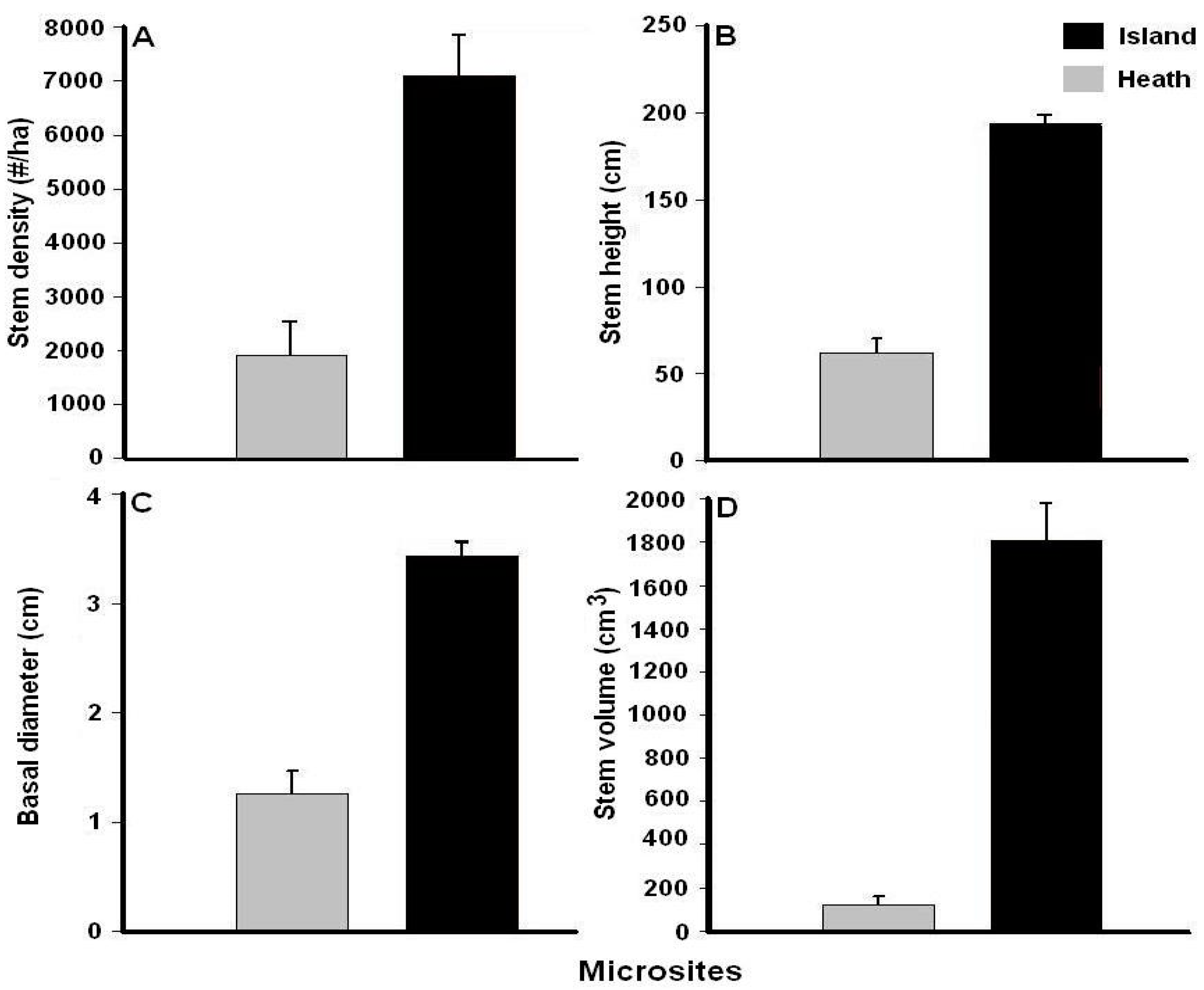
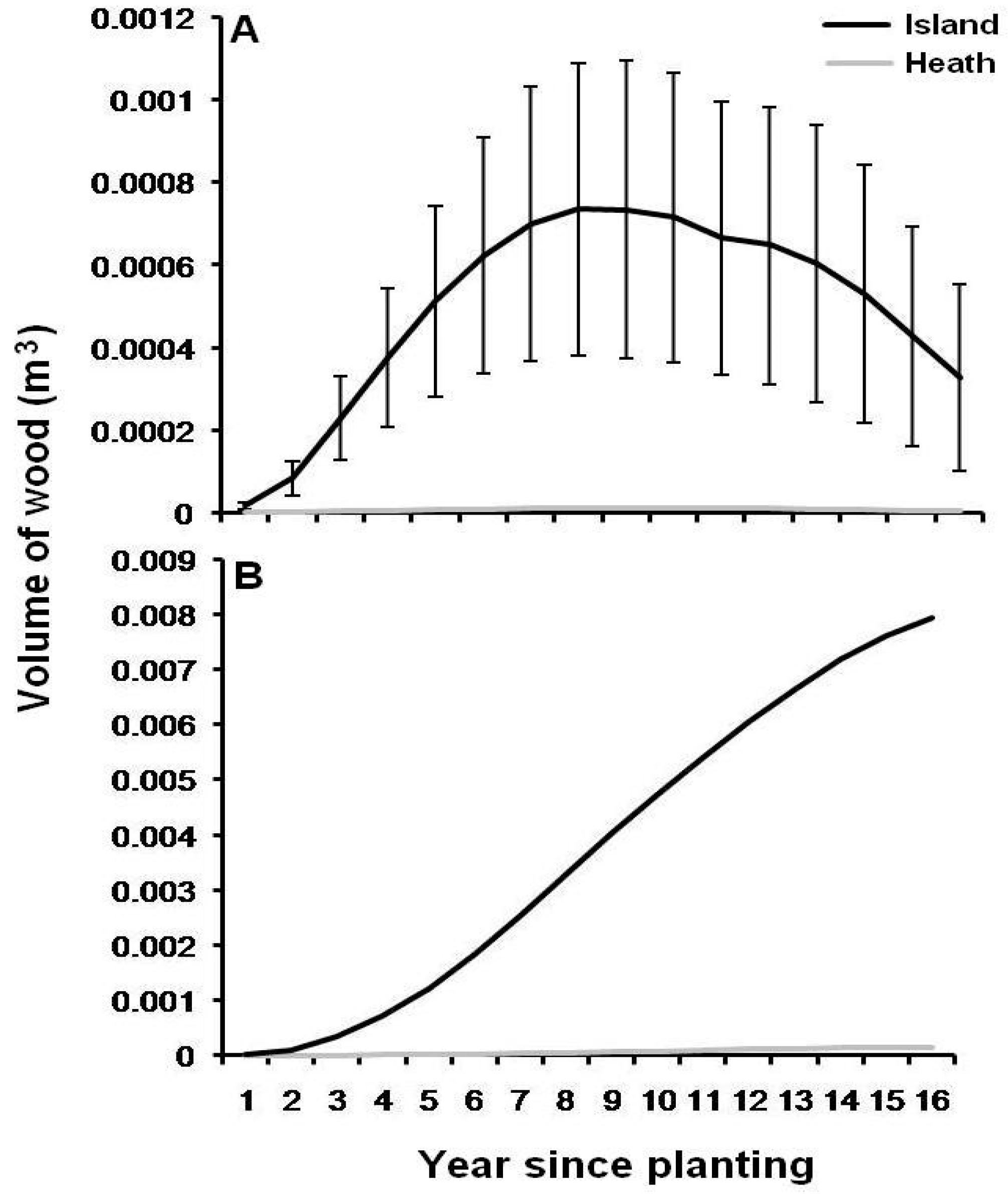
3.1. Black Spruce Stem Density and Growth
3.2. Foliar Nutrients
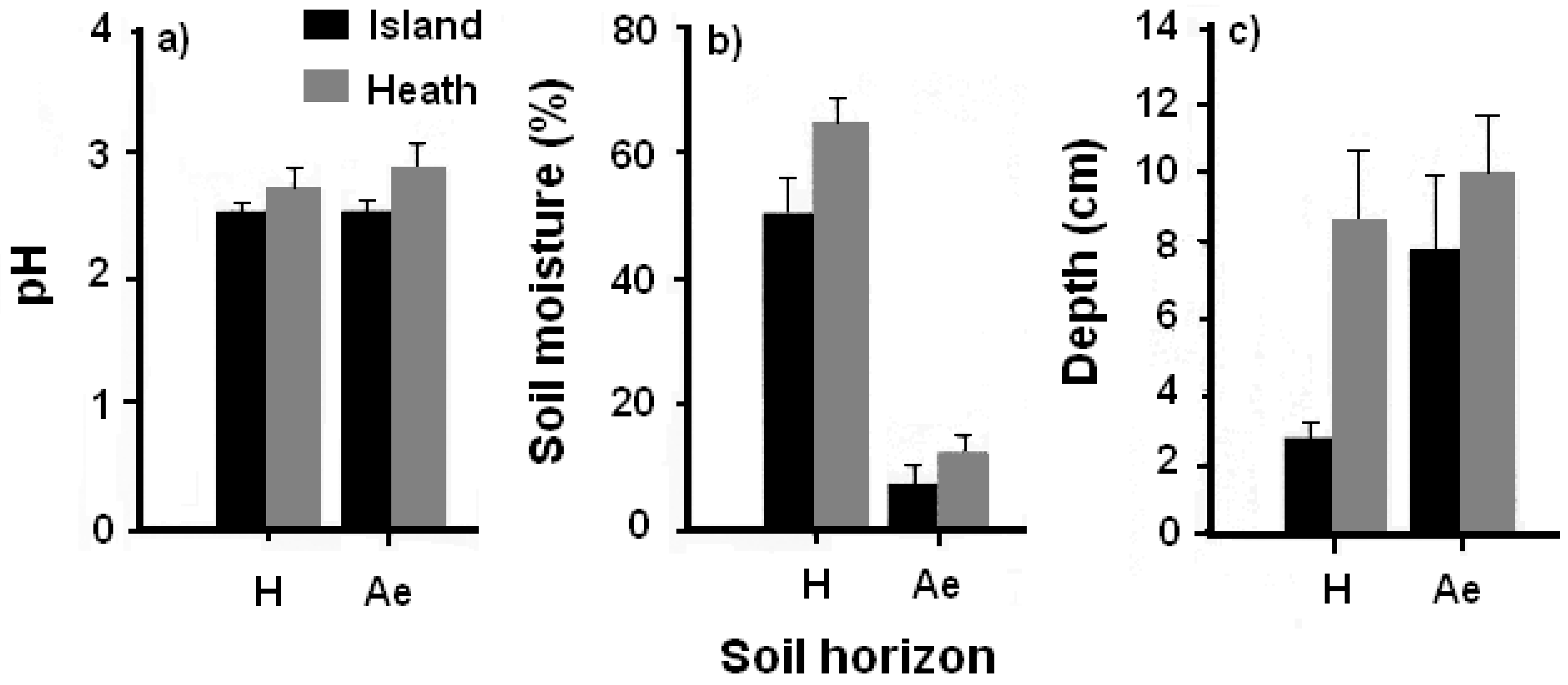
3.2.1. Soil pH, Moisture, and Humus and Ae Horizon Depth
3.2.2. Soil Nutrients
3.3. Black Spruce Shade Effects on Kalmia
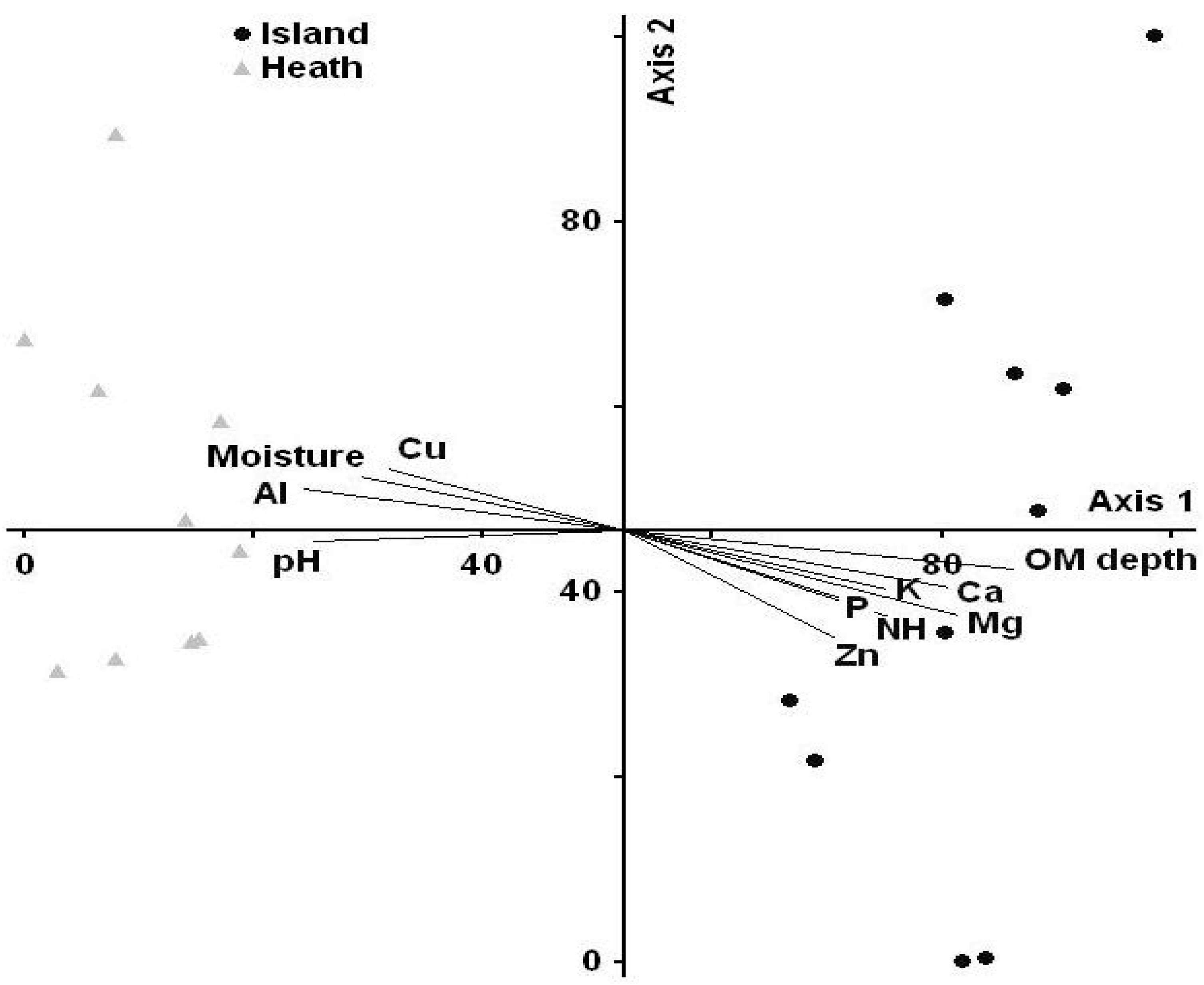
3.4. Community Composition
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Candy, R.H. Reproduction on cutover and burned over land in Canada. Canada Department of Research and Development, Forest Research Division. Silvic. Res. Note 1951, 92, 224. [Google Scholar]
- Page, G. The Development of Kalmia Angustifolia on Black Spruce Cutover in Central Newfoundland; Internal Report N-27; Forest Research Laboratory: St John’s, NL, Canada, 1970; 7p. [Google Scholar]
- Richardson, J.; Hall, J.P. Natural Regeneration after Disturbance in the Forest of Central Newfoundland; Information Report N-X-86; Canada Department of Environment, Canadian Forestry Service: Ottawa, QC, Canada, 1973; p. 63. [Google Scholar]
- Damman, A.W. H Permanent changes in the chronosequence of a boreal forest habitat. In Sukessionsforschung; Schmidt, W., Ed.; Cramer: Vanduz, Germany, 1975; pp. 499–515. [Google Scholar]
- Yamasaki, S.H.; Fyles, J.W.; Egger, N.E.; Titus, B.D. The effect of Kalmia angustifolia on growth, nutrition and ectomycorrhizal symbiont community of black spruce. For. Ecol. Manag. 1998, 105, 197–207. [Google Scholar] [CrossRef]
- Thiffault, N.; Titus, B.D.; Munson, A.D. Black spruce seedlings in a Kalmia–Vaccinium association: Microsite manipulation to explore interactions in the field. Can. J. For. Res. 2004, 34, 1657–1668. [Google Scholar] [CrossRef]
- Mallik, A.U. Growth and physiological responses of Black Spruce (Picea mariana) to sites dominated by Ledum groenlandicum. J. Chem. Ecol. 1996, 22, 575–585. [Google Scholar] [CrossRef]
- Jäderlund, A.; Zackrisson, O.; Dahlberg, X.; Nilsson, M.-C. Interference of Vaccinium myrtillus on establishment, growth and nutrition of Picea abies in a northern boreal site. Can. J. For. Res. 1997, 27, 2017–2025. [Google Scholar] [CrossRef]
- Mallik, A.U. Allelopathy and competition in coniferous forests. In Environmental Forest Science; Sassa, K., Ed.; Kluwer Academic Publishers: London, UK, 1998; pp. 309–315. [Google Scholar]
- Mallik, A.U. Black spruce growth, and understory species diversity in contiguous plots with and without sheep laurel (Kalmia angustifolia). Agron. J. 2001, 93, 92–98. [Google Scholar] [CrossRef]
- Mallik, A.U.; Prescott, C.E. Growth inhibitory effects of salal (Gaultheria shallon) on western hemlock (Thuia heterophylla) and western red cedar (Thuia plicata). Agron J. 2001, 93, 85–92. [Google Scholar] [CrossRef]
- English, B.; Hackett, R. The Impact of Kalmia on Plantation Performance in Central Newfoundland. Silvic. Noteb. 1994, 2, 12. [Google Scholar]
- Krause, H.H. Ericaceous vegetation as a site factor in jack pine growth of a New Brunswick plantation. In Proceedings of the IUFRO Workshop, Fredericton, NB, Canada, 20–24 July 1980; Fredericton, N.B. Canadian Forestry Service: Fredericton, NB, Canada. [Google Scholar]
- Damman, A.W.H. Effect of Vegetation Changes on the Fertility of a Newfoundland Forest Site. Ecol. Monogr. 1971, 41, 253–270. [Google Scholar] [CrossRef]
- Bradley, R.; Titus, B.; Fyles, J. Nitrogen acquisition and competitive ability of Kalmia angustifolia L., paper birch (Betula papyrifera Marsh.) and black spruce (Picea mariana (Mill.) B.S.P.) seedlings grown on different humus forms. Plant Soil 1997, 195, 209–220. [Google Scholar] [CrossRef]
- Meades, W.J.; Moores, L. Forest Site Classification Manual: A Field Guide to the Damman Forest Types of Newfoundland; Forest Resource Development Agreement Rep. 003; Western Newfoundland Model Forest Inc.: Corner Brook, NL, Canada, 1989; 226p. [Google Scholar]
- Mallik, A.U. Competitive ability and allelopathy of ericaceous plants as potential causes of conifer regeneration failures. J. Kor. For. Soc. 1996, 84, 394–406. [Google Scholar]
- Mallik, A.U. Conversion of temperate forests into heaths: Role of ecosystem disturbance and ericaceous plants. Environ. Manag. 1995, 19, 675–684. [Google Scholar] [CrossRef]
- Mallik, A.U.; Biswas, S.R.; Collier, L.C.S. Belowground negative interactions between ericaceous plants and conifers –the example of sheep laurel and black spruce. Plant Soil 2016, 403, 471–483. [Google Scholar] [CrossRef]
- Mallik, A. Allelopathic potential of Kalmia angustifolia to black spruce (Picea mariana). For. Ecol. Manag. 1987, 20, 43–51. [Google Scholar] [CrossRef]
- Zhu, H.; Mallik, A.U. Interactions betweenKalmia and black spruce: Isolation and identification of allelopathic compounds. J. Chem. Ecol. 1994, 20, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Mallik, A. Effect of phenolic compounds on selected soil properties. For. Ecol. Manag. 1997, 92, 11–18. [Google Scholar] [CrossRef]
- Bloom, R.G.; Mallik, A.U. Relationships between ericaceous vegetation and soil nutrient status in a post-fire Kalmia angustifolia-black spruce chronosequence. Plant Soil 2006, 289, 211–226. [Google Scholar] [CrossRef]
- Joanisse, G.D.; Bradley, R.L.; Preston, C.M.; Bending, G.D. Sequestration of soil nitrogen as tannin–protein complexes may improve the competitive ability of sheep laurel (Kalmia angustifolia) relative to black spruce (Picea mariana). New Phytol. 2008, 181, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Joanisse, G.D.; Bradley, R.L.; Preston, C.M.; Munson, A.D. Soil enzyme inhibition by condensed litter tannins may drive ecosystem structure and processes: The case of Kalmia angustifolia. New Phytol. 2007, 175, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Read, D.J.; Leake, J.; Perez-Moreno, J. Mycorrhizal fungi as drivers of ecosystem processes in heathland and boreal forest biomes. Can. J. Bot. 2004, 82, 1243–1263. [Google Scholar] [CrossRef]
- Martin, P.S.; Mallik, A.U. Soil chemistry drives below ground traits in an alternate successional pathway from forest to heath. Oecologia 2021, 195, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Mallik, A.U. Conifer regeneration problems in boreal and temperate forests with ericaceous understory: Role of disturbance, seedbed limitation and keystone species change (invited paper). Crit. Rev. Plant Sci. 2003, 22, 341–366. [Google Scholar] [CrossRef]
- Newton, P. An integrated approach to deriving site-specific black spruce regeneration standards by management objective. For. Ecol. Manag. 1998, 102, 143–156. [Google Scholar] [CrossRef]
- Wardle, D.A.; Nilsson, M.-C.; Gallet, C.; Zackrisson, O. An ecosystem level perspective of allelopathy. Biol. Rev. 1998, 73, 305–319. [Google Scholar] [CrossRef]
- Collier, L.C.S.; Mallik, A. Does post-fire abiotic habitat filtering create divergent plant communities in black spruce forests of eastern Canada? Oecologia 2010, 164, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK, 1977. [Google Scholar]
- Nyland, R.D. Patterns of lodgepole pine regeneration following the 1988 Yellowstone fires. For. Ecol. Manag. 1998, 111, 23–33. [Google Scholar] [CrossRef]
- Mallik, A.U. Ecology of a forest weed of Newfoundland: Vegetative regeneration strategy of Kalmia angustifolia. Can. J. Bot. 1993, 71, 161–166. [Google Scholar] [CrossRef]
- Bloom, R.G.; Mallik, A.U. Indirect effects of black spruce (Picea mariana) cover on community structure and function in sheep laurel (Kalmia angustifolia) dominated heath of eastern Canada. Plant Soil 2004, 265, 279–293. [Google Scholar] [CrossRef]
- Mallik, A.U.; Bloom, R.G.; Whisenant, S.G. Seedbed filter controls post-fire succession. Basic Appl. Ecol. 2010, 11, 170–181. [Google Scholar] [CrossRef]
- Mallik, A. Autecological response of Kalmia angustifolia to forest types and disturbance regimes. For. Ecol. Manag. 1994, 65, 231–249. [Google Scholar] [CrossRef]
- Mallik, A.; Newton, P. Inhibition of black spruce seedling growth by forest-floor substrates of central Newfoundland. For. Ecol. Manag. 1988, 23, 273–283. [Google Scholar] [CrossRef]
- Mallik, A.; Kayes, I. Lichen matted seedbeds inhibit while moss dominated seedbeds facilitate black spruce (Picea mariana) seedling regeneration in post-fire boreal forest. For. Ecol. Manag. 2018, 427, 260–274. [Google Scholar] [CrossRef]
- Jasinski, J.P.P.; Payette, S. THE Creation of Alternative Stable States in the Southern Boreal Forest, Quebec, Canada. Ecol. Monogr. 2005, 75, 561–583. [Google Scholar] [CrossRef]
- Enright, N.J.; Fontaine, J.B.; Bowman, D.M.J.S.; Bradstock, R.A.; Williams, R.J. Interval squeeze: Altered fire regimes and demographic responses interact to threaten woody species persistence as climate changes. Front. Ecol. Environ. 2015, 13, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Girard, F.; Payette, S.; Gagnon, R. Rapid expansion of lichen woodlands within the closed-crown boreal forest zone over the last 50 years caused by stand disturbances in eastern Canada. J. Biogeogr. 2007, 35, 529–537. [Google Scholar] [CrossRef]
- South, R. Biogeography and Ecology of the Island of Newfoundland; Springer: Berlin/Heidelberg, Germany, 1983; p. 723. [Google Scholar]
- Soil Classification Working Group. Canadian Soil Classification System, 3rd ed.; Agriculture and Agri-Food Canada Publication 1646, NRC Press: Ottawa, QC, Canada, 1998. [Google Scholar]
- Allen, S.E. Chemical Analysis of Ecological Materials; Blackwell Scientific Publications: London, UK, 1989; p. 368. [Google Scholar]
- van den Driessche, R. Prediction of mineral nutrient status of trees by foliar analysis. Bot. Rev. 1974, 40, 347–394. [Google Scholar] [CrossRef]
- Garten, C.T., Jr. Correlations between concentrations of elements in plants. Nature 1976, 261, 686–688. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Tyron, P.R. Habitat, and leaf habit as determinants of growth, nutrient absorption and nu-trient use by Alaskan taiga forest species. Can. J. For. Res. 1983, 13, 818–826. [Google Scholar] [CrossRef]
- Inderjit, M.A.U. Nutrient status of black spruce forest soils dominated by Kalmia angustifolia L. Acta Oecol. 1999, 20, 87–92. [Google Scholar] [CrossRef]
- Ludwig, J.A.; Reynolds, J.F. Statistical Ecology; John Wiley and Sons: New York, NY, USA, 1988. [Google Scholar]
- Zimmerman, G.M.; Goetz, H.; Mielke, P.W., Jr. Use of improved statistical method for group comparisons to study the effects of prairie fire. Ecology 1985, 66, 606–611. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data, Version 4.0; MjM Software Design: Gleneden Beach, OR, USA, 1999. [Google Scholar]
- Bloom, R.G. Direct and Indirect Effects of Post-Fire Successional Pathways and Ecological Processes in Black Spruce-Kalmia Forests. Ph.D. Thesis, Lakehead University, Thunder Bay, ON, Canada, 2001. [Google Scholar]
- Zenk, M.H.; Muller, D. In vivo destruction of exogenously applied indol-3-acetic acid as influenced by naturally occurring phenolic acids. Nature 1963, 200, 761–763. [Google Scholar] [CrossRef]
- Tomaszewski, M.; Thimann, K. V. Interactions of phenolic acids, metalic ions and chelating agents on auxin induced growth. Plant Physiol. 1966, 41, 1443–1454. [Google Scholar] [CrossRef] [PubMed]
- Inderjit, M.A.U. The nature of interference potential of Kalmia angustifolia. Can. J. For. Res. 1996, 26, 1899–1904. [Google Scholar] [CrossRef]
- Appel, H.M. Phenolics in ecological interactions: The importance of oxidation. J. Chem. Ecol. 1993, 19, 1521–1552. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Martin, P.S.; Mallik, A.U. Alternate successional pathway yields alternate pattern of functional diversity. J. Veg. Sci. 2019, 30, 461–470. [Google Scholar] [CrossRef]
- Mallik, A.; Kravchenko, D. Black spruce (Picea mariana) restoration in Kalmia heath by scarification and microsite mulching. For. Ecol. Manag. 2016, 362, 10–19. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Kalmia Heath | Black Spruce Island | t-Statistic | p-Value | Power | Effect Size |
|---|---|---|---|---|---|---|
| N (%) | 0.748 ± 0.049 | 1.051 ± 0.033 | 2.234 | 0.040 | 1.0 | 7.25 |
| P (ppm) | 12.507 ± 0.981 | 11.860 ± 0.226 | −0.560 | 0.588 | 0.60 | 0.91 |
| K (ppm) | 4998 ± 297.92 | 5127.778 ± 208.944 | 0.412 | 0.686 | 0.28 | 0.50 |
| Al (ppm) | 1.021 ± 0.164 | 0.643 ± 0.059 | −2.00 | 0.071 | 1.0 | 3.07 |
| Ca (ppm) | 25.290 ± 11.85 | 49.094 ± 1.575 | −1.394 | 0.196 | 1.0 | 2.82 |
| Cu (ppm) | 0.024 ± 0.00 | 0.023 ± 0.004 | −0.339 | 0.739 | 0.13 | 0.25 |
| Fe (ppm) | 0.313 ± 0.01 | 0.281 ± 0.023 | −0.838 | 0.417 | 0.97 | 1.71 |
| Mg (ppm) | 4.881 ± 0.42 | 8.011 ± 0.216 | 6.761 | <0.001 | 1.0 | 9.32 |
| Mn (ppm) | 28.956 ± 5.53 | 15.652 ± 1.529 | 10.528 | 0.049 | 1.0 | 3.28 |
| Zn (ppm) | 0.607 ± 0.11 | 0.409 ± 0.026 | −2.317 | 0.042 | 1.0 | 2.54 |
| Nutrient | Soil Horizon | Kalmia Heath | Black Spruce Island | t-Statistic | p-Value | Power | Effect Size |
|---|---|---|---|---|---|---|---|
| NH+ (%) | Oh | 3.62 ± 1.02 | 3.01 ± 1.18 | −0.908 | 0.388 | 0.31 | 0.553 |
| Ae | 2.26 ± 0.60 | 3.51 ± 0.92 | 8.427 | <0.001 | 0.96 | 1.610 | |
| Ca (ppm) | Oh | 4.14 ± 2.23 | 4.45 ± 2.10 | 0.229 | 0.824 | 0.09 | 0.143 |
| Ae | 2.42 ± 0.65 | 4.86 ± 1.39 | 4.130 | 0.003 | 1.0 | 2.249 | |
| K (ppm) | Oh | 4.30 ± 1.73 | 4.35 ± 1.24 | 0.054 | 0.958 | 0.06 | 0.033 |
| Ae | 3.07 ± 0.37 | 4.60 ± 1.34 | 3.958 | 0.003 | 0.95 | 1.557 | |
| Mg (ppm) | Oh | 3.10 ± 2.03 | 3.79 ± 1.55 | 0.617 | 0.553 | 0.20 | 0.382 |
| Ae | 1.92 ± 0.45 | 4.28± 1.27 | 4.818 | 0.001 | 1.0 | 2.477 | |
| Na (ppm) | Oh | 3.01 ± 1.07 | 2.72 ± 0.56 | −0.658 | 0.527 | 0.17 | 0.340 |
| Ae | 2.24 ± 0.62 | 2.78 ± 0.82 | 1.981 | 0.079 | 0.46 | 0.743 | |
| Al (ppm) | Oh | 5.92 ± 11.10 | 5.10 ± 0.82 | −1.373 | 0.203 | 0.55 | 0.845 |
| Ae | 6.29 ±0.80 | 4.70 ± 0.55 | −3.871 | 0.004 | 1.0 | 2.316 | |
| Cu (ppm) | Oh | −0.84 ± 11.53 | −0.32 ± 1.32 | 0.583 | 0.574 | 0.19 | 0.364 |
| Ae | 0.35 ± 0.34 | −0.37 ± 0.80 | −3.389 | 0.008 | 0.79 | 1.171 | |
| Fe (ppm) | Oh | 3.91 ± 0.79 | 3.54 ± 0.62 | −0.981 | 0.352 | 0.29 | 0.521 |
| Ae | 3.68 ± 0.93 | 3.05± 0.26 | −2.163 | 0.059 | 0.61 | 0.922 | |
| Mn (ppm) | Oh | 3.24 ± 1.65 | 2.94 ± 2.71 | −0.228 | 0.825 | 0.09 | 0.134 |
| Ae | 0.54 ± 2.30 | 2.36 ± 2.43 | 2.586 | 0.029 | 0.48 | 0.769 | |
| Zn (ppm) | Oh | 1.36 ± 1.36 | 1.75 ± 1.13 | 0.501 | 0.628 | 0.16 | 0.312 |
| Ae | −0.33 ± 0.32 | 1.46± 1.57 | 2.624 | 0.028 | 0.95 | 1.580 | |
| P (ppm) | Oh | 4.01 ± 0.96 | 3.22 ± 0.47 | −1.861 | 0.096 | 0.70 | 1.045 |
| Ae | 2.80 ± 0.51 | 3.52 ± 0.47 | 4.716 | 0.001 | 0.92 | 1.468 |
| Site | Average Distance | N | MRPP Statistics |
|---|---|---|---|
| Black spruce island Kalmia heath | 0.4741 0.3167 | 10 10 | Observed Delta = 0.3959 Expected Delta = 0.5836 T = −12.25 A = 0.321 P = < 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mallik, A.U. Post-Fire Habitat Heterogeneity Leads to Black Spruce–Kalmia L. Shrub Savannah Alternate State. Forests 2022, 13, 570. https://doi.org/10.3390/f13040570
Mallik AU. Post-Fire Habitat Heterogeneity Leads to Black Spruce–Kalmia L. Shrub Savannah Alternate State. Forests. 2022; 13(4):570. https://doi.org/10.3390/f13040570
Chicago/Turabian StyleMallik, Azim U. 2022. "Post-Fire Habitat Heterogeneity Leads to Black Spruce–Kalmia L. Shrub Savannah Alternate State" Forests 13, no. 4: 570. https://doi.org/10.3390/f13040570
APA StyleMallik, A. U. (2022). Post-Fire Habitat Heterogeneity Leads to Black Spruce–Kalmia L. Shrub Savannah Alternate State. Forests, 13(4), 570. https://doi.org/10.3390/f13040570