
Gross Primary Production of Dwarf Bamboo, Sasa senanensis, in Cool-Temperate Secondary Forests with Different Canopy Structures

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
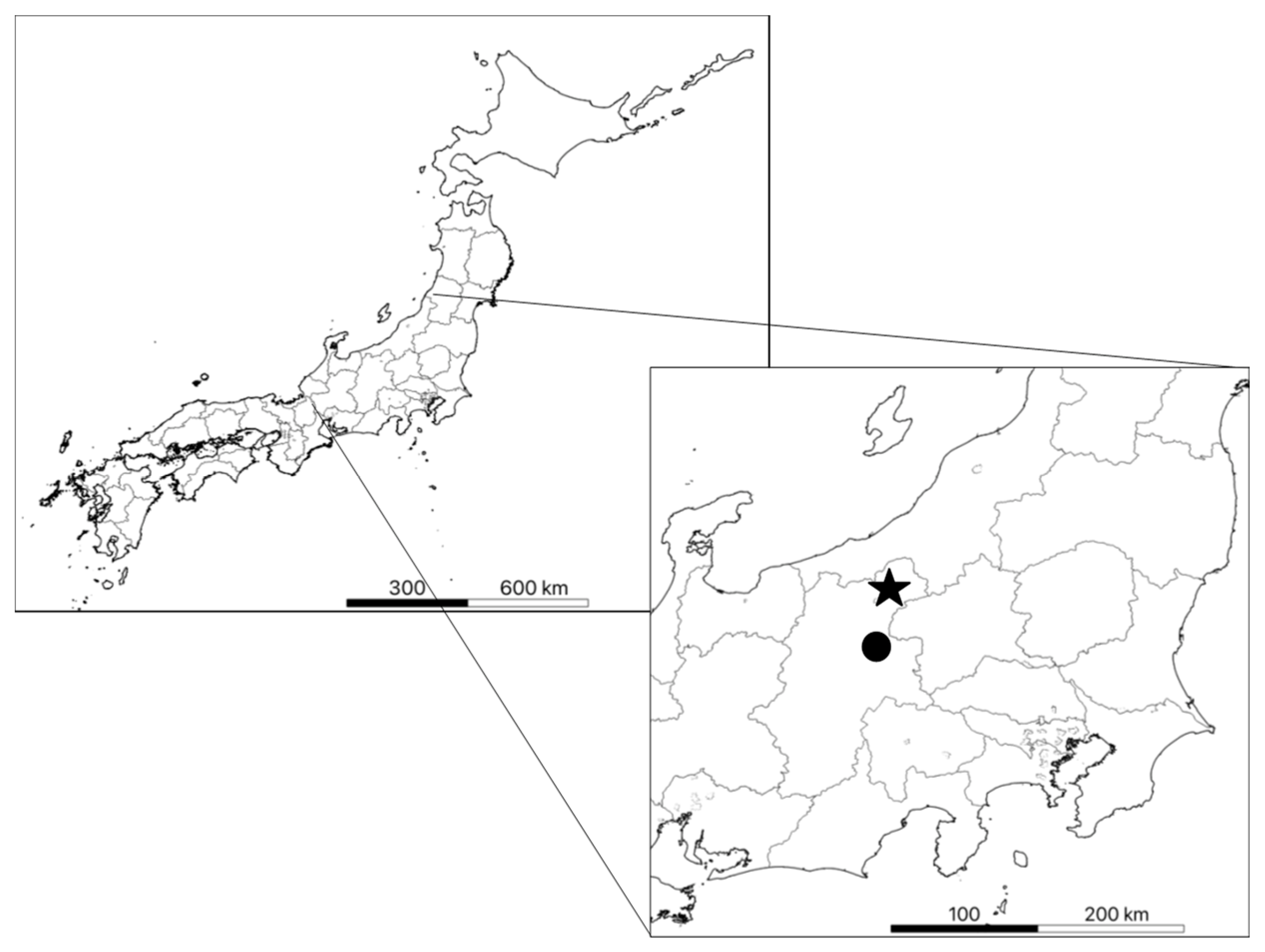
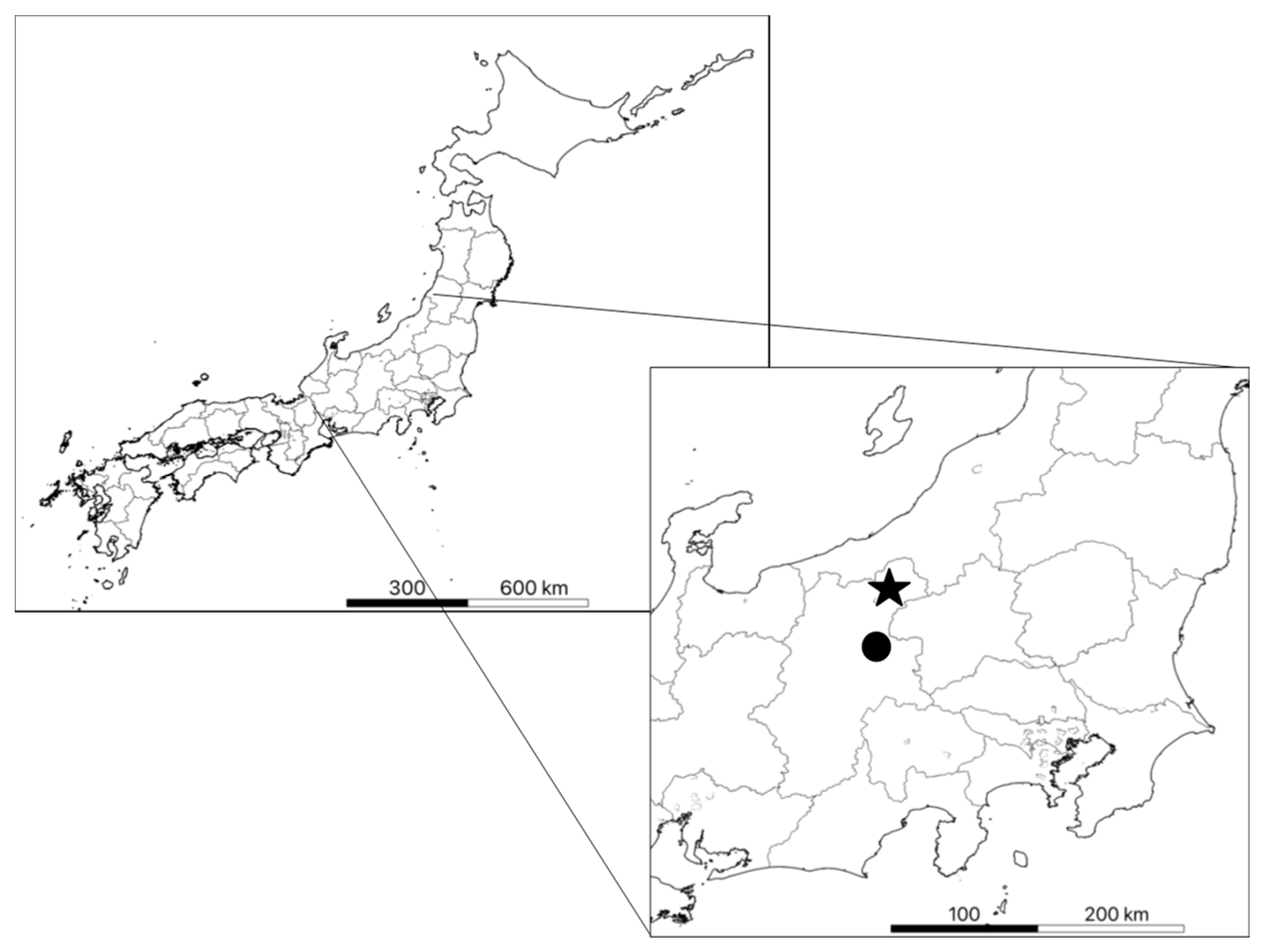
2.1. Study Area
2.2. Measurement of Metrological Variables
2.3. CO2 Flux Measurement and Calculation
2.4. Sasa Senanensis GPP at the Community Scale and its Relationship with PPFD
2.5. Biomass and Leaf Area of S. senanensis
2.6. Sasa GPP at the Forest Ecosystem Scale and Its Contribution to Forest GPP
2.7. Statistical Analysis
3. Results
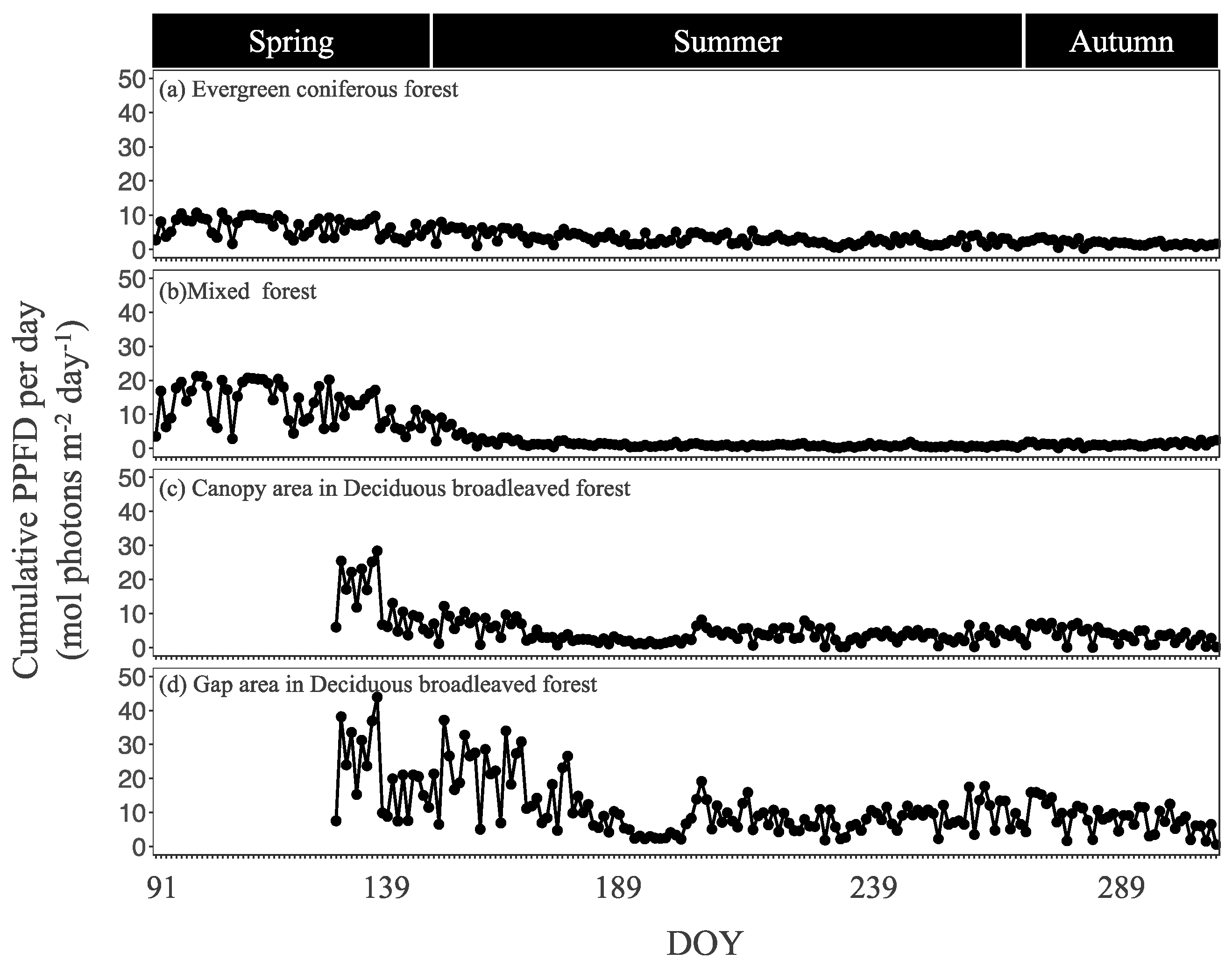
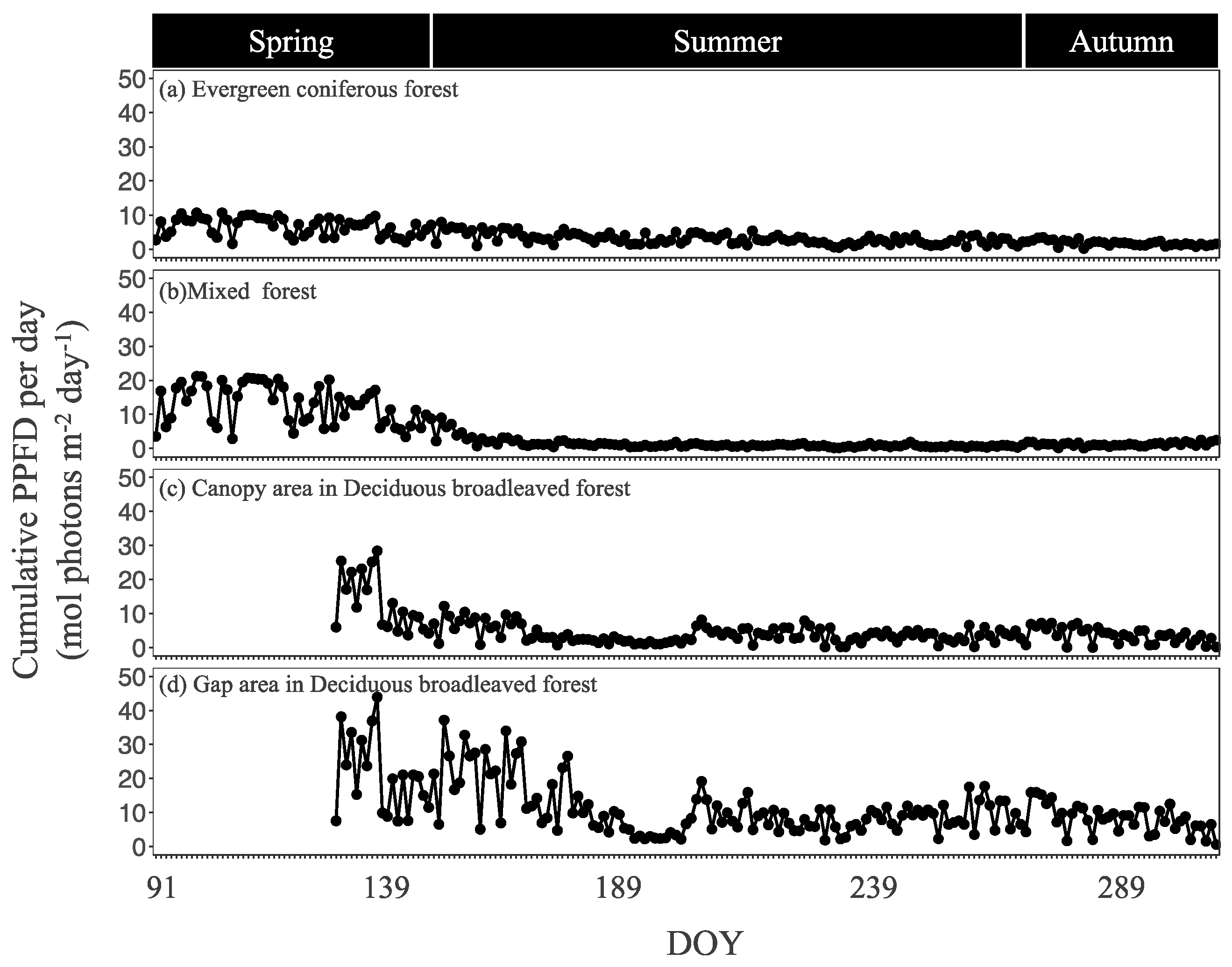
3.1. Seasonal Dynamics in Light Intensity and Other Environmental Factors
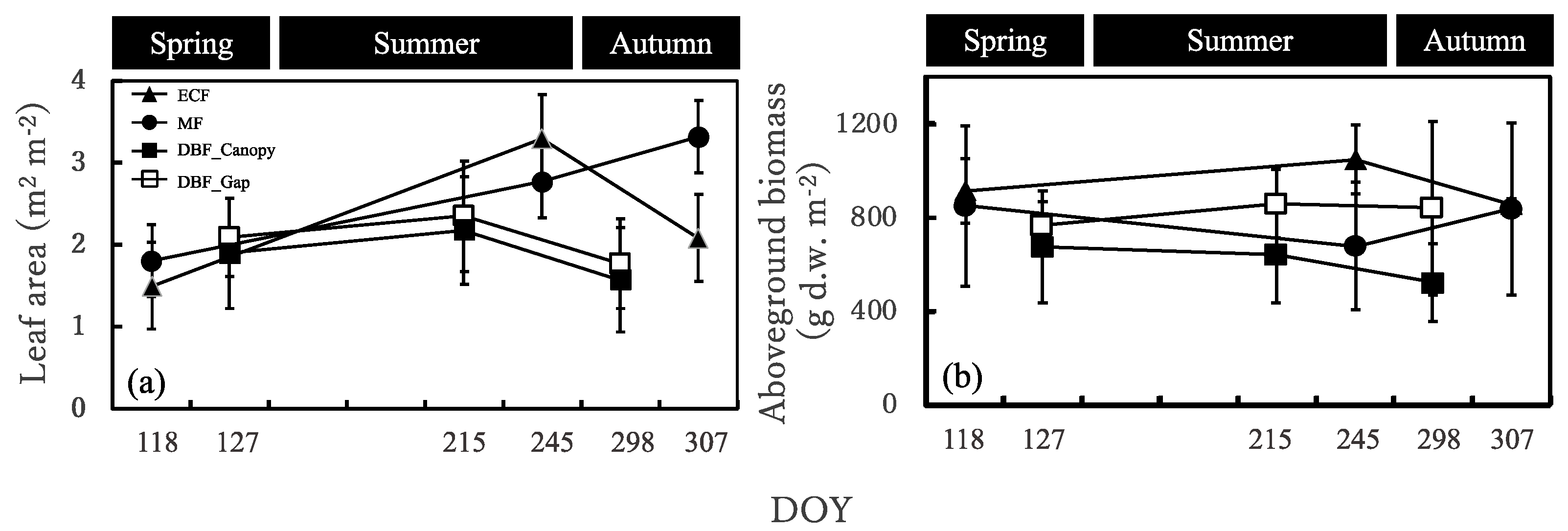
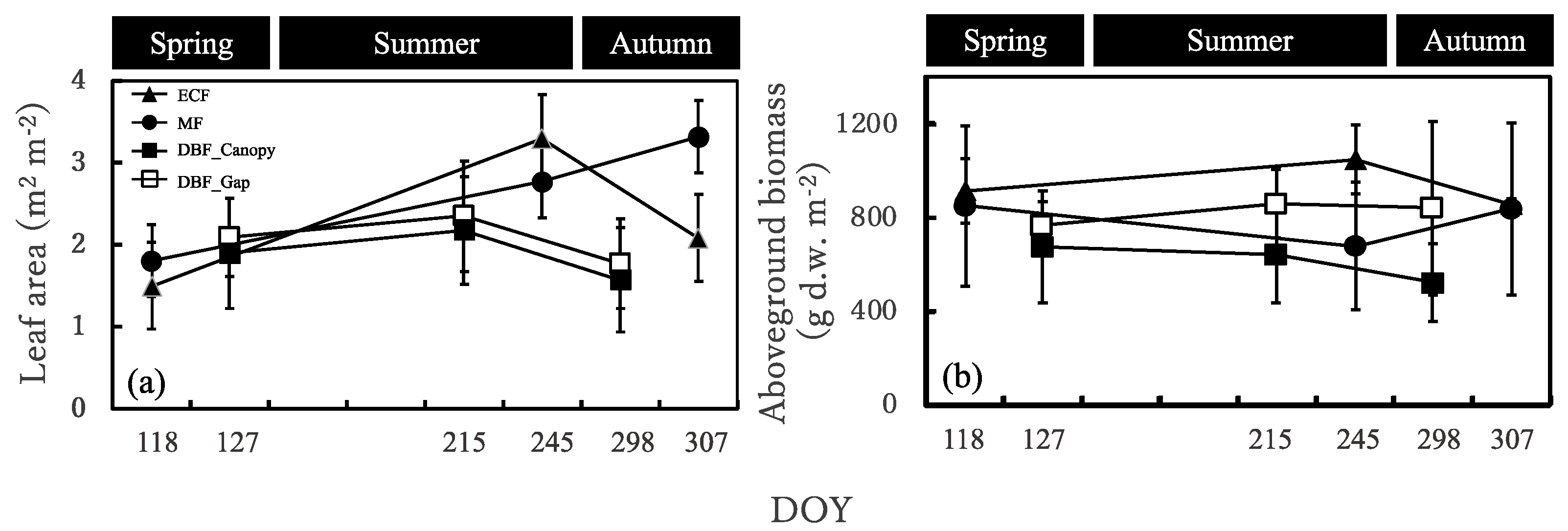
3.2. Seasonal Dynamics in the Sasa Community Leaf Area and Aboveground Biomass
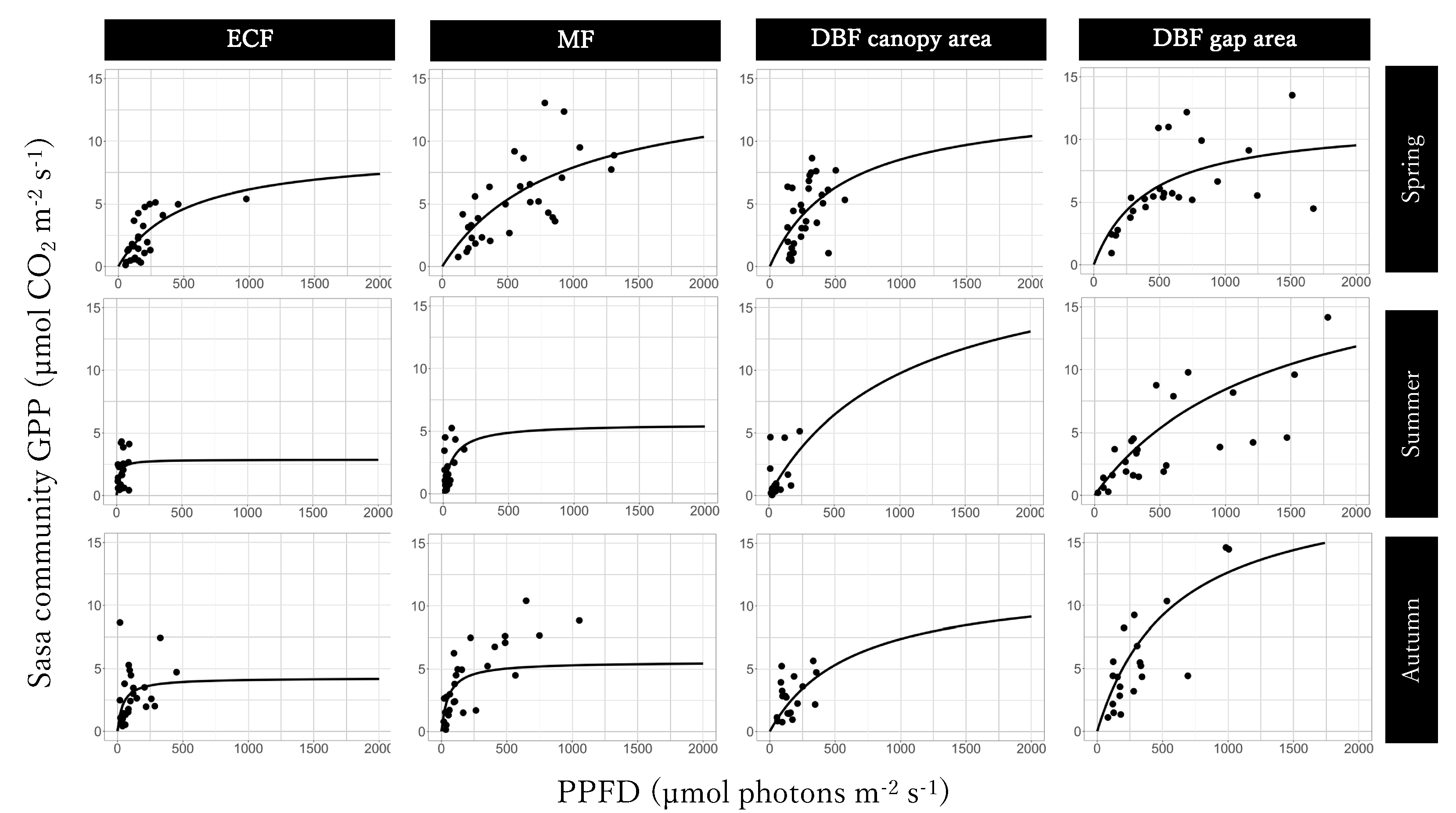
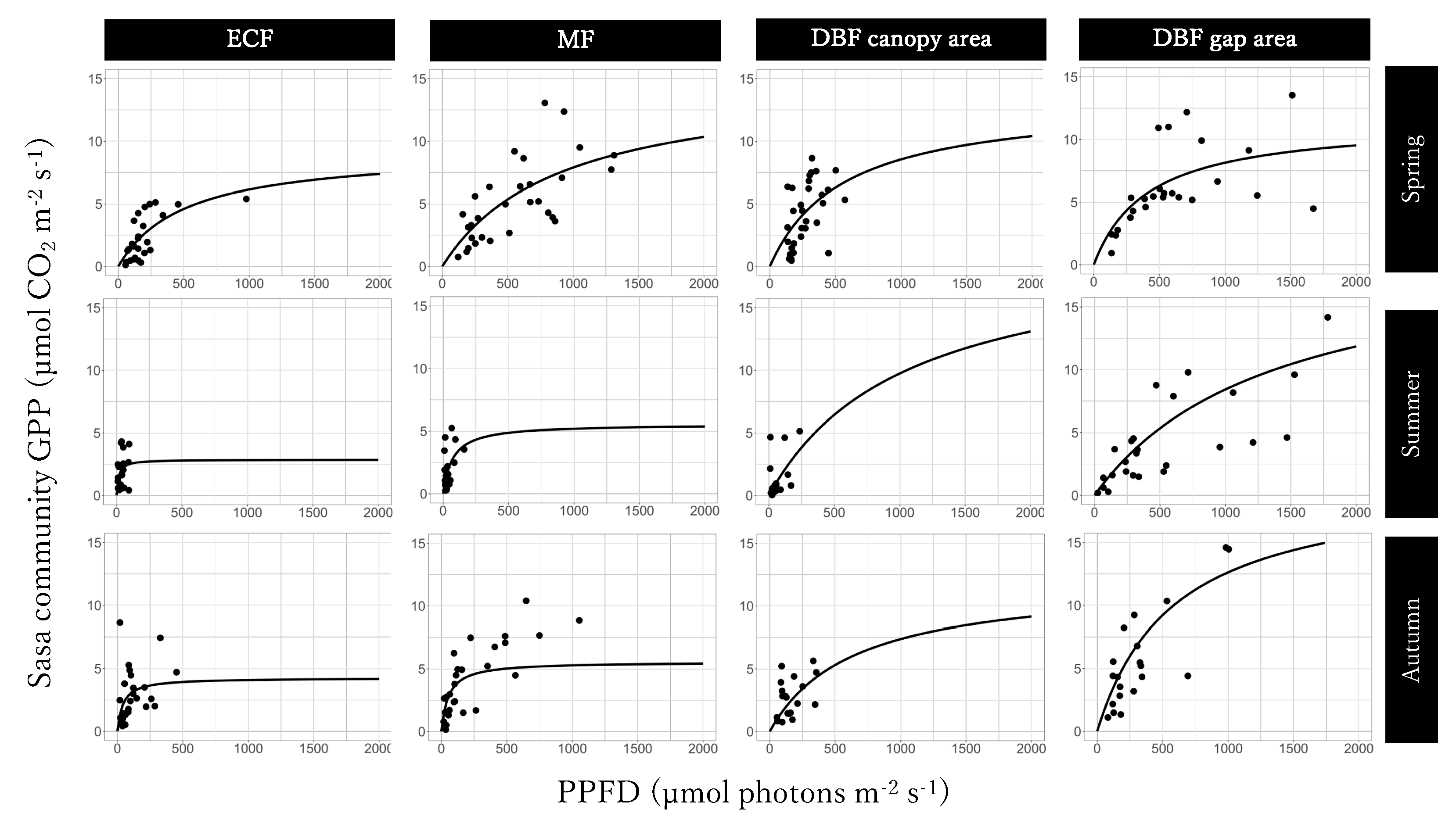
3.3. Light-GPPSasa-community Curves
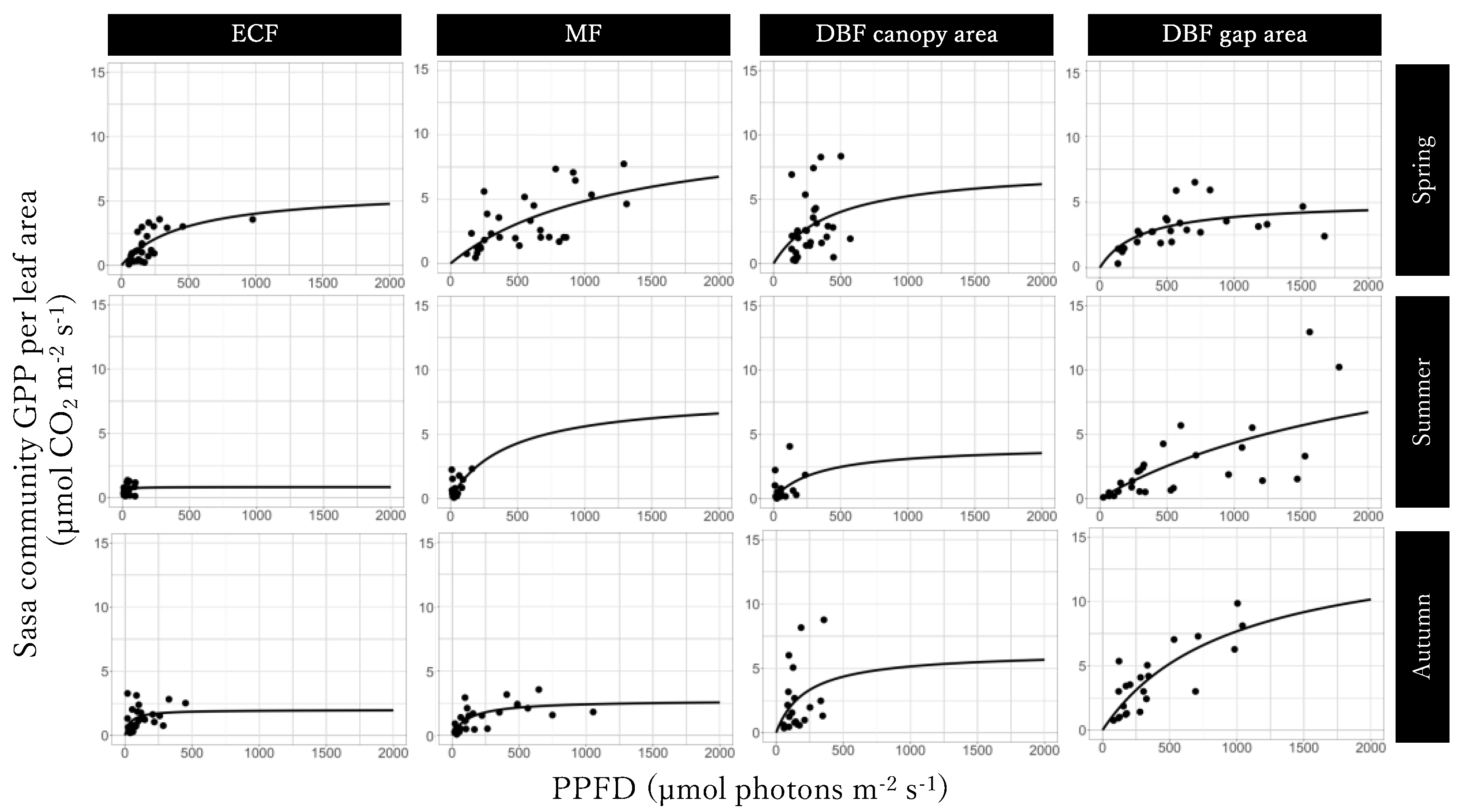
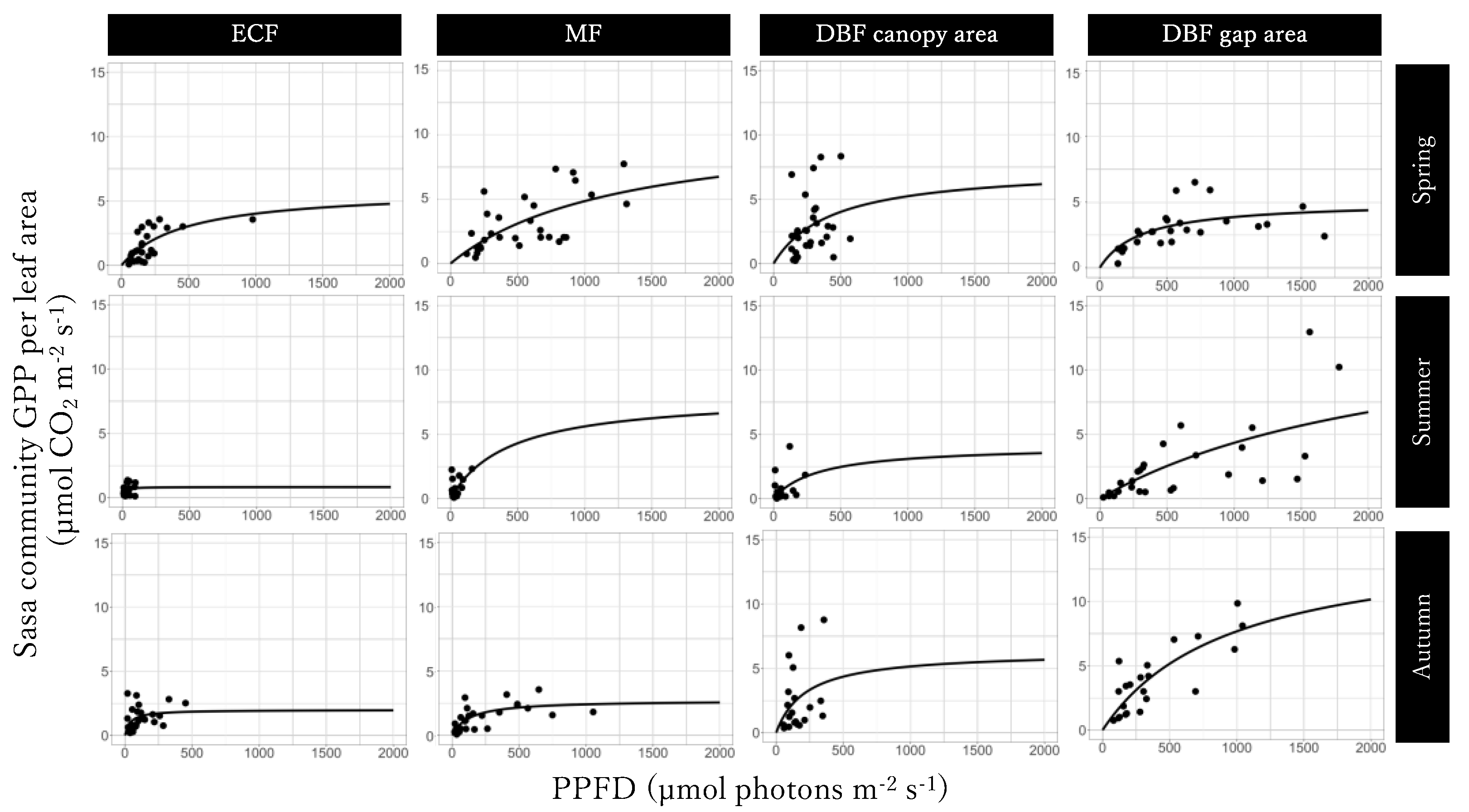
3.4. Light-GPPSasa-community Curves per Leaf Area
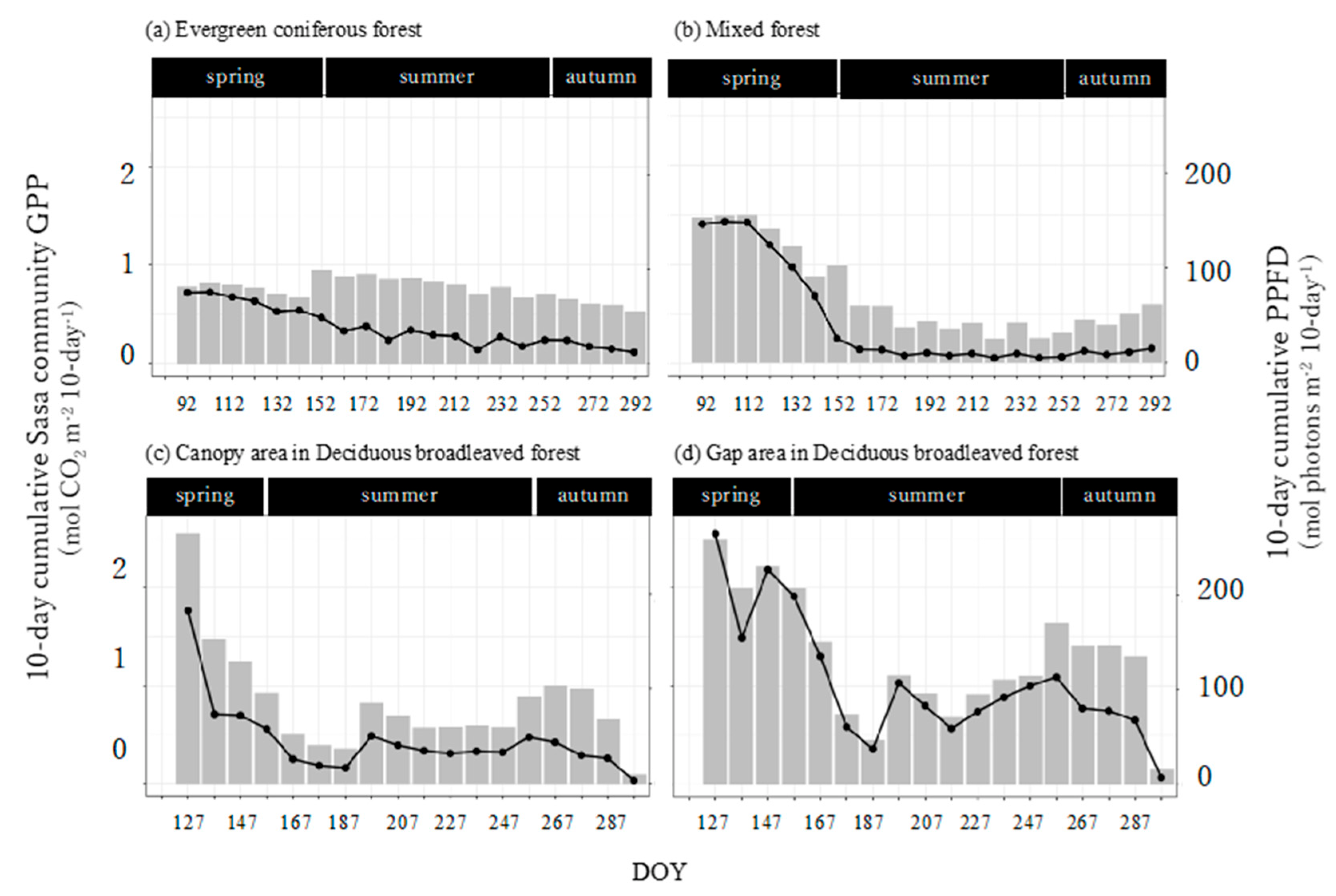
3.5. Seasonal Dynamics in the Sasa Community GPP and Its Controlling Factors
3.6. Annual Sasa GPP and Community GPP at the Forest Scale for Each Forest Type
4. Discussion
4.1. Photosynthetic Ability of the Sasa Community
4.2. Seasonal Dynamics in Sasa Community GPP and Annual Sasa GPP
4.3. Annual Changes in Sasa GPP
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chokkalingam, U.; De Jong, W. Secondary forests: A working definition and typology. Int. For. Rev. 2001, 3, 19–26. [Google Scholar]
- Sugiura, K.; Sonohara, W. Application of an Adaptive Forest Management Simulation Model Based on Zoning in a Man-Made Forest. Forests 2019, 10, 482. [Google Scholar] [CrossRef] [Green Version]
- Luyssaert, S.; Schulze, E.-D.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2013: The Physical Science Basis; Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Chapin, F.S.; Matson, P.A.; Mooney, H.A. Principles of Terrestrial Ecosystem Ecology, 2nd ed.; Springer: New York, NY, USA, 2011. [Google Scholar]
- Desai, A.R.; Richardson, A.D.; Moffat, A.M.; Kattge, J.; Hollinger, D.Y.; Barr, A.; Falge, E.; Noormets, A.; Papale, D.; Reichstein, M.; et al. Cross-site evaluation of eddy covariance GPP and RE decomposition techniques. Agric. For. Meteorol. 2008, 148, 821–838. [Google Scholar] [CrossRef]
- Mäkelä, A.; Kolari, P.; Karimäki, J.; Nikinmaa, E.; Perämäki, M.; Hari, P. Modelling five years of weather-driven variation of GPP in a boreal forest. Agric. For. Meteorol. 2006, 139, 382–398. [Google Scholar] [CrossRef]
- Croft, H.; Chen, J.M.; Froelich, N.J.; Chen, B.; Staebler, R.M. Seasonal controls of canopy chlorophyll content on forest carbon uptake: Implications for GPP modeling. J. Geophys. Res. Biogeosci. 2015, 120, 1576–1586. [Google Scholar] [CrossRef] [Green Version]
- Misson, L.; Baldocchi, D.; Black, T.; Blanken, P.; Brunet, Y.; Yuste, J.C.; Dorsey, J.; Falk, M.; Granier, A.; Irvine, M.; et al. Partitioning forest carbon fluxes with overstory and understory eddy-covariance measurements: A synthesis based on FLUXNET data. Agric. For. Meteorol. 2007, 144, 14–31. [Google Scholar] [CrossRef] [Green Version]
- Kolari, P.; Pumpanen, J.; Kulmala, L.; Ilvesniemi, H.; Nikinmaa, E.; Grönholm, T.; Hari, P. Forest floor vegetation plays an important role in photosynthetic production of boreal forests. For. Ecol. Manag. 2006, 221, 241–248. [Google Scholar] [CrossRef]
- Sakai, T.; Akiyama, T.; Saigusa, N.; Yamamoto, S.; Yasuoka, Y. The contribution of gross primary production of understory dwarf bamboo, Sasa senanensis, in a cool-temperate deciduous broadleaved forest in central Japan. For. Ecol. Manag. 2006, 236, 259–267. [Google Scholar] [CrossRef]
- Cai, Y.; Tanioka, Y.; Kitawaga, T.; Ida, H.; Hirota, M. Gross primary production of dwarf bamboo, Sasa senanensis, in a mature beech forest with a substantial gap-mosaic structure. J. Plant Res. 2021, 134, 209–221. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Law, B.E.; Anthoni, P.M. On measuring and modeling energy fluxes above the floor of a homogeneous and heterogeneous conifer forest. Agric. For. Meteorol. 2000, 102, 187–206. [Google Scholar] [CrossRef]
- Launiainen, S.; Rinne, J.; Pumpanen, J.; Kulmala, L.; Kolari, P.; Keronen, P.; Siivola, E.; Pohja, T.; Hari, P.; Vesala, T. Eddy covariance measurements of CO2 and sensible and latent heat fluxes during a full year in a boreal pine forest trunk-space. Boreal Environ. Res. 2005, 10, 569–588. [Google Scholar]
- Oshima, Y. Ecological Studies of Sasa Communities I. Productive structure of some of the Sasa communities in Japan. Bot. Mag. Tokyo 1961, 74, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Narukawa, Y.; Yamamoto, S. Effects of dwarf bamboo (Sasa sp.) and forest floor microsites on conifer seedling recruitment in a subalpine forest, Japan. For. Ecol. Manag. 2002, 163, 61–70. [Google Scholar] [CrossRef]
- Doležal, J.; Matsuki, S.; Hara, T. Effects of dwarf-bamboo understory on tree seedling emergence and survival in a mixed-oak forest in northern Japan: A multi-site experimental study. Community Ecol. 2009, 10, 225–235. [Google Scholar] [CrossRef]
- Abe, M.; Izaki, J.; Miguchi, H.; Masaki, T.; Makita, A.; Nakashizuka, T. The effects of Sasa and canopy gap formation on tree regeneration in an old beech forest. J. Veg. Sci. 2002, 13, 565–574. [Google Scholar] [CrossRef]
- Cho, S.; Lee, K.; Choung, Y. Distribution, abundance, and effect on plant species diversity of Sasa borealis in Korean forests. J. Ecol. Environ. 2018, 42, 9. [Google Scholar] [CrossRef] [Green Version]
- Tsuyama, I.; Matsui, T.; Horikawa, M.; Kominami, Y.; Tanaka, N. Habitat prediction and impact assessment of climate change on dwarf bamboo of the Section Sasa in Japan. Theory Appl. GIS 2008, 16, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Suyama, Y.; Obayashi, K.; Hayashi, I. Clonal structure in a dwarf bamboo (Sasa senanensis) population inferred from amplified fragment length polymorphism (AFLP) fingerprints. Mol. Ecol. 2000, 9, 901–906. [Google Scholar] [CrossRef]
- Ida, H. Forest structure in a beech (Fagus crenata Blume) stand on a 1-ha permanent plot for the Monitoring Sites 1000 Project in Kayanodaira, central Japanese snowbelt. Bull. Inst. Nat. Educ. Shiga Height. Shinshu Univ. 2013, 50, 33–40. [Google Scholar]
- Ishihara, M.; Center, Y.F.E.C.; Suzuki, S.; Nakamura, M.; Enoki, T.; Fujiwara, A.; Hiura, T.; Homma, K.; Hoshino, D.; Hoshizaki, K.; et al. Forest stand structure, composition, and dynamics in 34 sites over Japan. Ecol. Res. 2011, 26, 1007–1008. [Google Scholar] [CrossRef]
- Thornley, M.N.; Johnson, I.R. Plant and Crop Modeling: A Mathematical Approach to Plant and Crop Physiology; Clarendon Press: Oxford, UK, 1990. [Google Scholar]
- Waring, R.H.; Landsberg, J.J.; Williams, M. Net primary production of forests: A constant fraction of gross primary production? Tree Physiol. 1998, 18, 129–134. [Google Scholar] [CrossRef]
- DeLucia, E.H.; Drake, J.; Thomas, R.B.; Gonzalez-Meler, M. Forest carbon use efficiency: Is respiration a constant fraction of gross primary production? Glob. Chang. Biol. 2007, 13, 1157–1167. [Google Scholar] [CrossRef] [Green Version]
- Otsuka, T. Carbon cycling at Takayama Forest: Results from intensive studies in the last decade, and further studies for a next decade. Jpn. J. Ecol. 2012, 62, 31–44. [Google Scholar]
- Komiyama, A.; Nakagawa, M.; Kato, S. The Static Model of Plant Form, the Pipe Model. Ter. Mikaelian Korzukhin 2011, 31, 220225. [Google Scholar]
- Kato, T.; Tang, Y. Spatial variability and major controlling factors of CO2 sink strength in Asian terrestrial ecosystems: Evidence from eddy covariance data. Glob. Chang. Biol. 2008, 14, 2333–2348. [Google Scholar] [CrossRef]
- Saigusa, N.; Li, S.-G.; Kwon, H.; Takagi, K.; Zhang, L.-M.; Ide, R.; Ueyama, M.; Asanuma, J.; Choi, Y.-J.; Chun, J.H.; et al. Dataset of CarboEastAsia and uncertainties in the CO2 budget evaluation caused by different data processing. J. For. Res. 2013, 18, 41–48. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Yang, S.-J.; Sun, M.; Zhang, Y.-J.; Cochard, H.; Cao, K.-F. Strong leaf morphological, anatomical, and physiological responses of a subtropical woody bamboo (Sinarundinaria nitida) to contrasting light environments. Plant Ecol. 2014, 215, 97–109. [Google Scholar] [CrossRef]
- Tsunoda, Y.; Furukawa, S.; Mizunaga, H. How does the longevity of Sasa kurilensis ramets respond to a light gradient? An analysis of ontogenetic changes to hydraulic resistance and carbon budget within a ramet. Ecol. Res. 2017, 32, 117–128. [Google Scholar] [CrossRef]
- Messier, C.; Parent, S.; Bergeron, Y. Effects of overstory and understory vegetation on the understory light environment in mixed boreal forests. J. Veg. Sci. 1998, 9, 511–520. [Google Scholar] [CrossRef]
- Vierling, L.A.; Wessman, C.A. Photosynthetically active radiation heterogeneity within a monodominant Congolese rain forest canopy. Agric. For. Meteorol. 2000, 103, 265–278. [Google Scholar] [CrossRef]
- Yamori, W.; Hikosaka, K.; Way, D. Temperature response of photosynthesis in C3, C4, and CAM plants: Temperature acclimation and temperature adaptation. Photosynth. Res. 2014, 119, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Kudo, G.; Aoshima, Y.; Miyata, R.; Winkler, D.E. Altered morphologies and physiological compensation in a rapidly expanding dwarf bamboo in alpine ecosystems. Arctic Antarct. Alp. Res. 2018, 50, e1463733. [Google Scholar] [CrossRef] [Green Version]
- Muscolo, A.; Bagnato, S.; Sidari, M.; Mercurio, R. A review of the roles of forest canopy gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) Evergreen coniferous e forest | |||||
| season | GPPmax | P | α | P | Residual SE |
| µmol m−2 s−1 | µmol m−2 s−1 | ||||
| spring | 9.26 | * | 0.018 | *** | 1.26 |
| summer | 2.87 | * | 0.20 | 1.36 | |
| autumn | 4.25 | ** | 0.10 | 2.06 | |
| (b) Mixed forest | |||||
| season | GPPmax | P | α | P | Residual SE |
| µmol m−2 s−1 | µmol m−2 s−1 | ||||
| spring | 15.57 | * | 0.016 | ** | 2.35 |
| summer | 5.57 | 0.077 | 1.44 | ||
| autumn | 9.93 | *** | 0.048 | *** | 1.67 |
| (c) Canopy area in Deciduous broadleaved forest | |||||
| season | GPPmax | P | α | P | Residual SE |
| µmol m−2 s−1 | µmol m−2 s−1 | ||||
| spring | 13.31 | * | 0.024 | * | 2.16 |
| summer | 8.00 | 0.023 | 1.33 | ||
| autumn | 4.90 | * | 0.041 | 1.45 | |
| (d) Gap area in Deciduous broadleaved forest | |||||
| season | GPPmax | P | α | P | Residual SE |
| µmol m−2 s−1 | µmol m−2 s−1 | ||||
| spring | 11.50 | *** | 0.028 | * | 2.50 |
| summer | 20.00 | * | 0.015 | * | 3.27 |
| autumn | 20.00 | ** | 0.034 | ** | 3.12 |
| (a) Evergreen coniferous forest | |||||
| season | GPPmax | P | α | P | Residual SE |
| µmol m−2 s−1 | µmol m−2 s−1 | ||||
| spring | 5.90 | * | 0.013 | *** | 1.26 |
| summer | 0.80 | 0.071 | * | 1.36 | |
| autumn | 1.99 | *** | 0.051 | 2.06 | |
| (b) Mixed forest | |||||
| season | GPPmax | P | α | P | Residual SE |
| µmol m−2 s−1 | µmol m−2 s−1 | ||||
| spring | 11.06 | 0.0086 | * | 2.35 | |
| summer | 8.07 | 0.018 | * | 1.44 | |
| autumn | 2.72 | *** | 0.021 | * | 1.67 |
| (c) Canopy area in Deciduous broadleaved forest | |||||
| season | GPPmax | P | α | P | Residual SE |
| µmol m−2 s−1 | µmol m−2 s−1 | ||||
| spring | 7.51 | 0.017 | 2.16 | ||
| summer | 4.13 | 0.012 | 1.33 | ||
| autumn | 6.30 | 0.028 | 1.45 | ||
| (d) Gap area in Deciduous broadleaved forest | |||||
| season | GPPmax | P | α | P | Residual SE |
| µmol m−2 s−1 | µmol m−2 s−1 | ||||
| spring | 5.06 | *** | 0.016 | * | 2.50 |
| summer | 20.00 | 0.0055 | * | 3.27 | |
| autumn | 15.36 | * | 0.016 | *** | 3.12 |
| (a) Evergreen coniferous forest | |||
| Estimate | p Value | ||
| intercept | −0.49 | <0.001 | |
| PPFD | 1.15 | <0.001 | |
| SWC | 0.077 | 0.38 | |
| Leaf area | 0.56 | <0.001 | |
| TA | −0.11 | 0.090 | |
| RH | 0.048 | 0.55 | |
| Adjusted R2 | 0.85 | p value | <0.001 |
| (b) Mixed forest | |||
| Estimate | p Value | ||
| intercept | −0.75 | <0.001 | |
| PPFD | 1.31 | <0.001 | |
| Leaf area | 0.56 | <0.001 | |
| TA | 0.34 | <0.001 | |
| RH | −0.071 | 0.32 | |
| Adjusted R2 | 0.75 | p value | <0.001 |
| (c) Canopy area in Deciduous broadleaved forest | |||
| Estimate | p Value | ||
| intercept | 0.18 | <0.01 | |
| PPFD | 0.92 | <0.001 | |
| Leaf area | −0.056 | 0.097 | |
| TA | −0.081 | 0.14 | |
| RH | −0.47 | <0.01 | |
| Adjusted R2 | 0.75 | p value | <0.001 |
| (d) Gap area in Deciduous broadleaved forest | |||
| Estimate | p Value | ||
| intercept | 0.023 | <0.01 | |
| PPFD | 0.96 | <0.001 | |
| Leaf area | −0.048 | 0.11 | |
| TA | −0.21 | <0.001 | |
| RH | −0.058 | 0.095 | |
| Adjusted R2 | 0.92 | p value | <0.001 |
| Sasa Community GPP per Leaf | Sasa Community GPP | |
|---|---|---|
| g C m−2 yr−1 | g C m−2 yr−1 | |
| Evergreen coniferous forest | 83.2 | 190.8 |
| Mixed forest | 80.7 | 176.8 |
| Canopy area in Deciduous broadleaved forest | 93.2 | 178.6 |
| Gap area in Deciduous broadleaved forest | 130.8 | 276.7 |
| Annual Sasa GPP | Tree GPP | Contribution | |
|---|---|---|---|
| t C ha−1 yr−1 | t C ha−1 yr−1 | % | |
| Evergreen coniferous forest | 1.9 | 11.2 ± 1.3 | 13.1 ± 1.3 |
| Mixed forest | 1.8 | 8.4 ± 4.0 | 19.1 ± 6.9 |
| Deciduous broadleaved forest | 2.1 | 16.8 ± 1.7 | 11.2 ± 0.9 |
| Canopy area | 1.8 | - | - |
| Gap area | 2.8 | - | - |
| Annual Sasa GPP | Cumulative PPFD | Leaf Biomass | ||
|---|---|---|---|---|
| t C ha−1 yr−1 | mol photons m−2 yr−1 | g DW m−2 | ||
| Cai et al. 2021 | DBF | 3.7 | - | - |
| Canopy area | 1.9 | 432 | 152 | |
| Gap area | 7.0 | 2309 | 286 | |
| This study | DBF | 2.1 | - | - |
| Canopy area | 1.8 | 819 | 128 | |
| Gap area | 2.8 | 1943 | 154 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, Y.; Koido, R.; Umino, T.; Sakamoto, H.; Hasebe, Y.; Sarmah, R.; Yoneda, M.; Ida, H.; Hirota, M. Gross Primary Production of Dwarf Bamboo, Sasa senanensis, in Cool-Temperate Secondary Forests with Different Canopy Structures. Forests 2022, 13, 564. https://doi.org/10.3390/f13040564
Cai Y, Koido R, Umino T, Sakamoto H, Hasebe Y, Sarmah R, Yoneda M, Ida H, Hirota M. Gross Primary Production of Dwarf Bamboo, Sasa senanensis, in Cool-Temperate Secondary Forests with Different Canopy Structures. Forests. 2022; 13(4):564. https://doi.org/10.3390/f13040564
Chicago/Turabian StyleCai, Yihan, Ririko Koido, Taichi Umino, Hiroki Sakamoto, Yuki Hasebe, Ragini Sarmah, Mio Yoneda, Hideyuki Ida, and Mitsuru Hirota. 2022. "Gross Primary Production of Dwarf Bamboo, Sasa senanensis, in Cool-Temperate Secondary Forests with Different Canopy Structures" Forests 13, no. 4: 564. https://doi.org/10.3390/f13040564
APA StyleCai, Y., Koido, R., Umino, T., Sakamoto, H., Hasebe, Y., Sarmah, R., Yoneda, M., Ida, H., & Hirota, M. (2022). Gross Primary Production of Dwarf Bamboo, Sasa senanensis, in Cool-Temperate Secondary Forests with Different Canopy Structures. Forests, 13(4), 564. https://doi.org/10.3390/f13040564