

Assessment of Past Decadal Dynamics of Tree Stands in Forest–Tundra Transition Zone on the Polar Ural Mountains Calibrated Using Historical and Modern Field Measurements


Abstract
1. Introduction
2. Materials and Methods
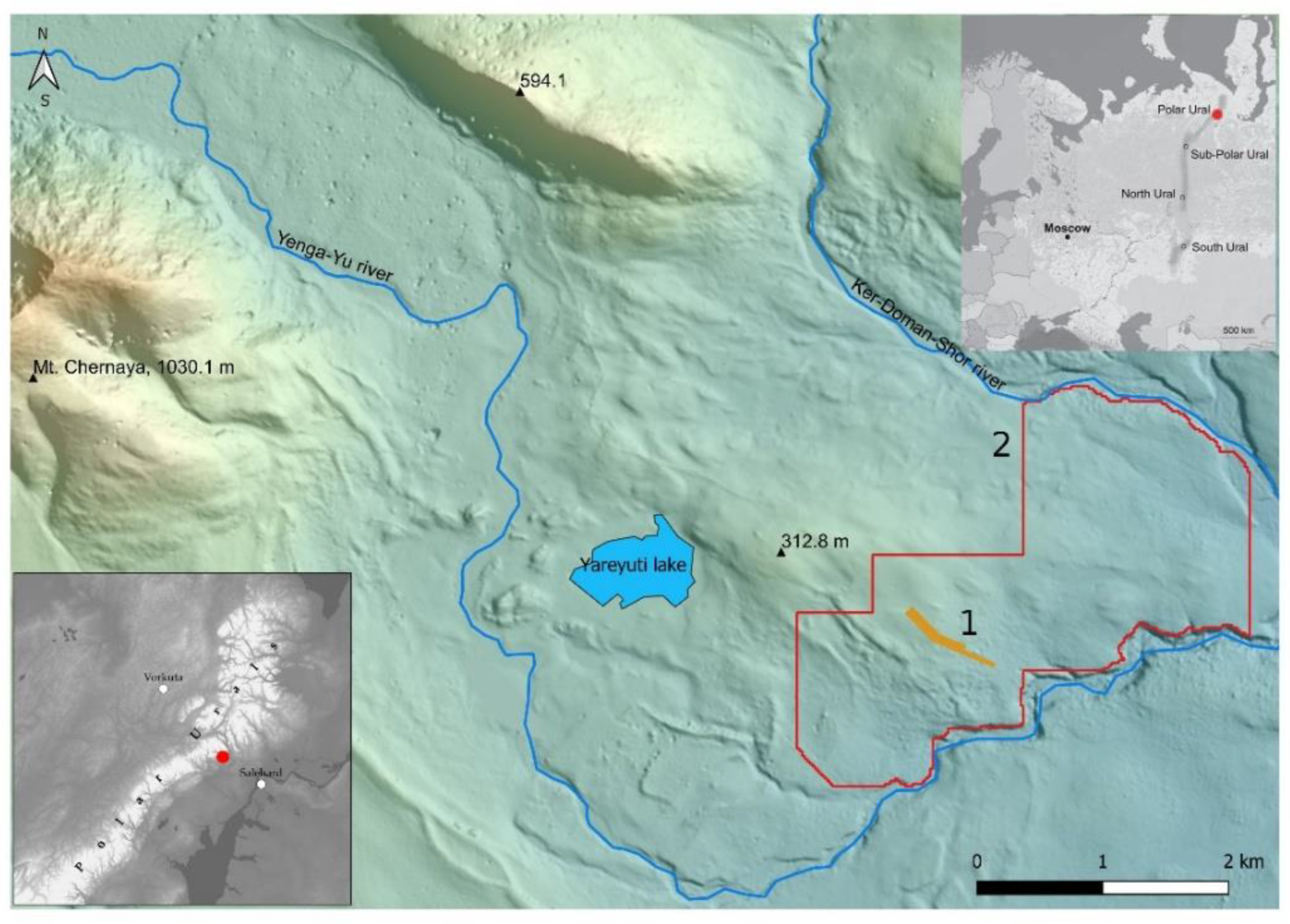
2.1. Study Site
2.2. Trees and Stands Data Sampling and Calculation
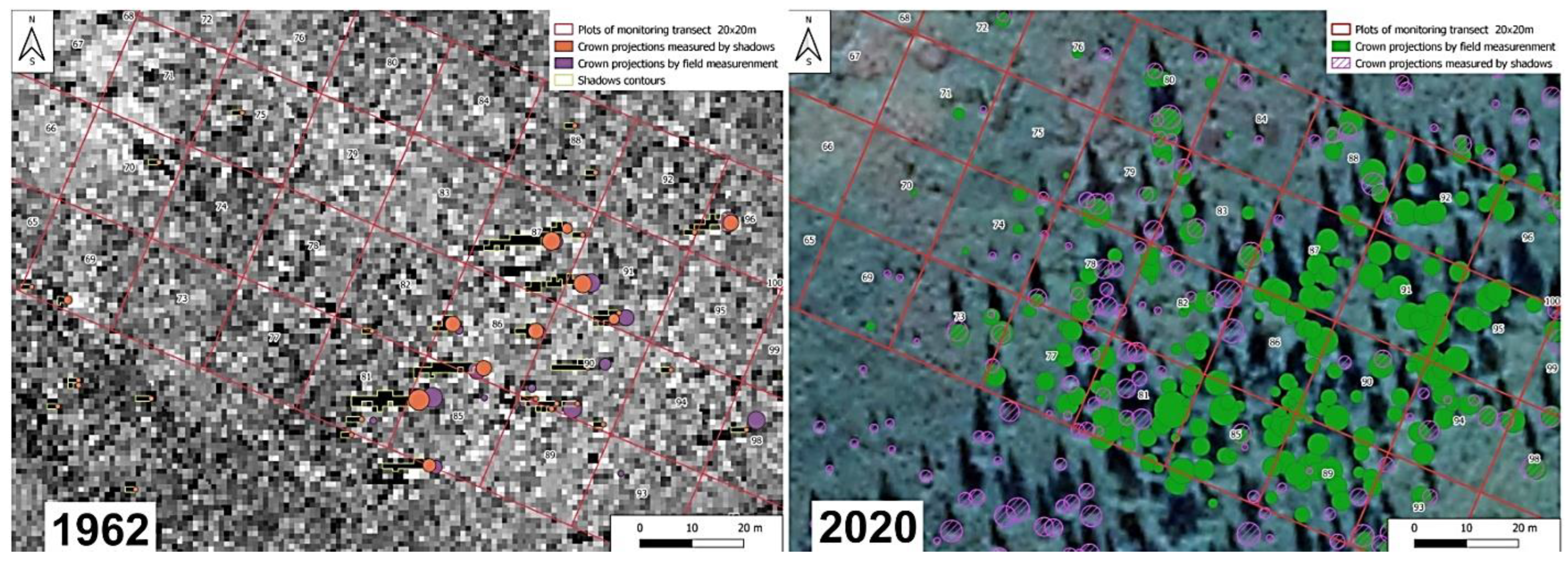
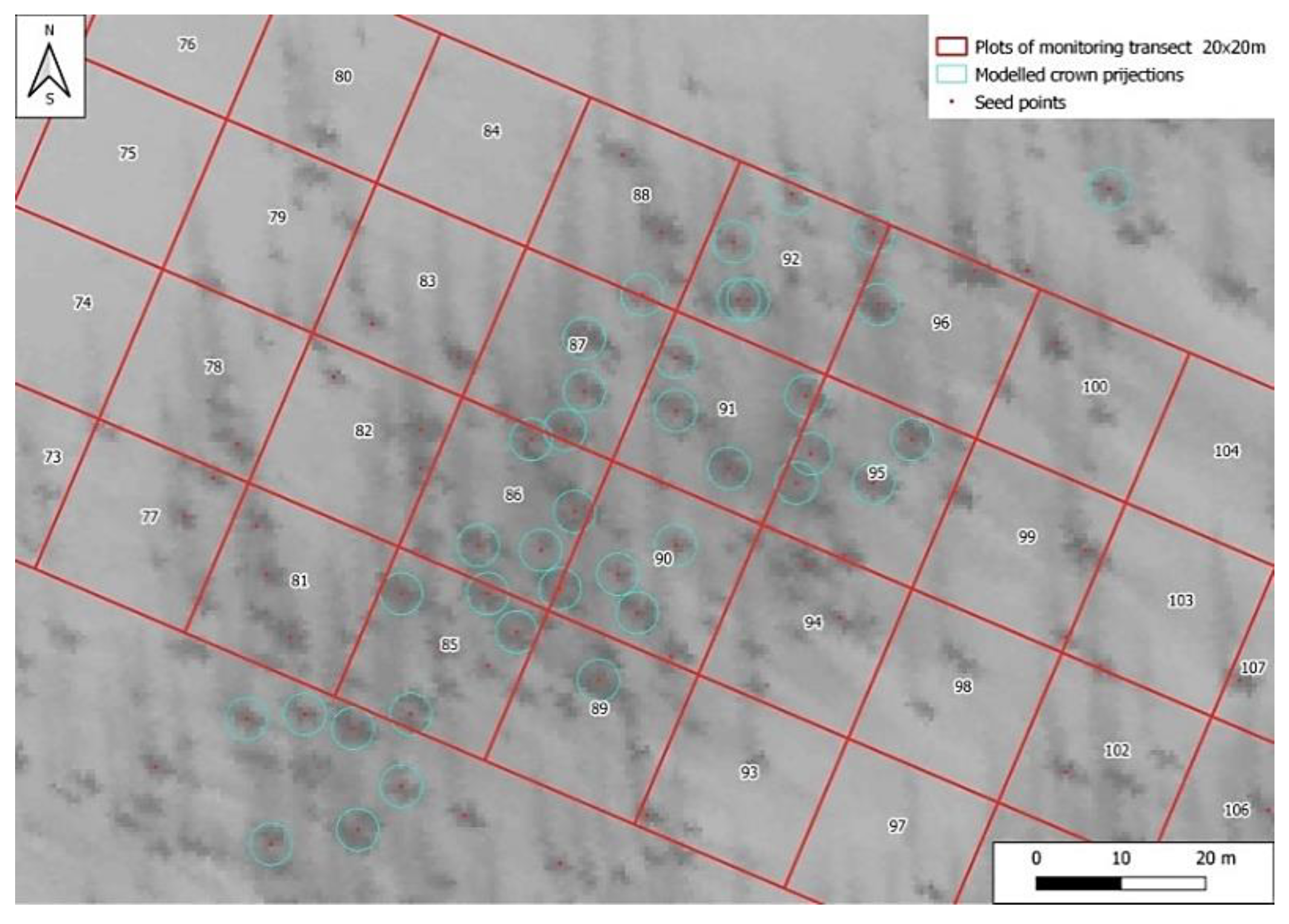
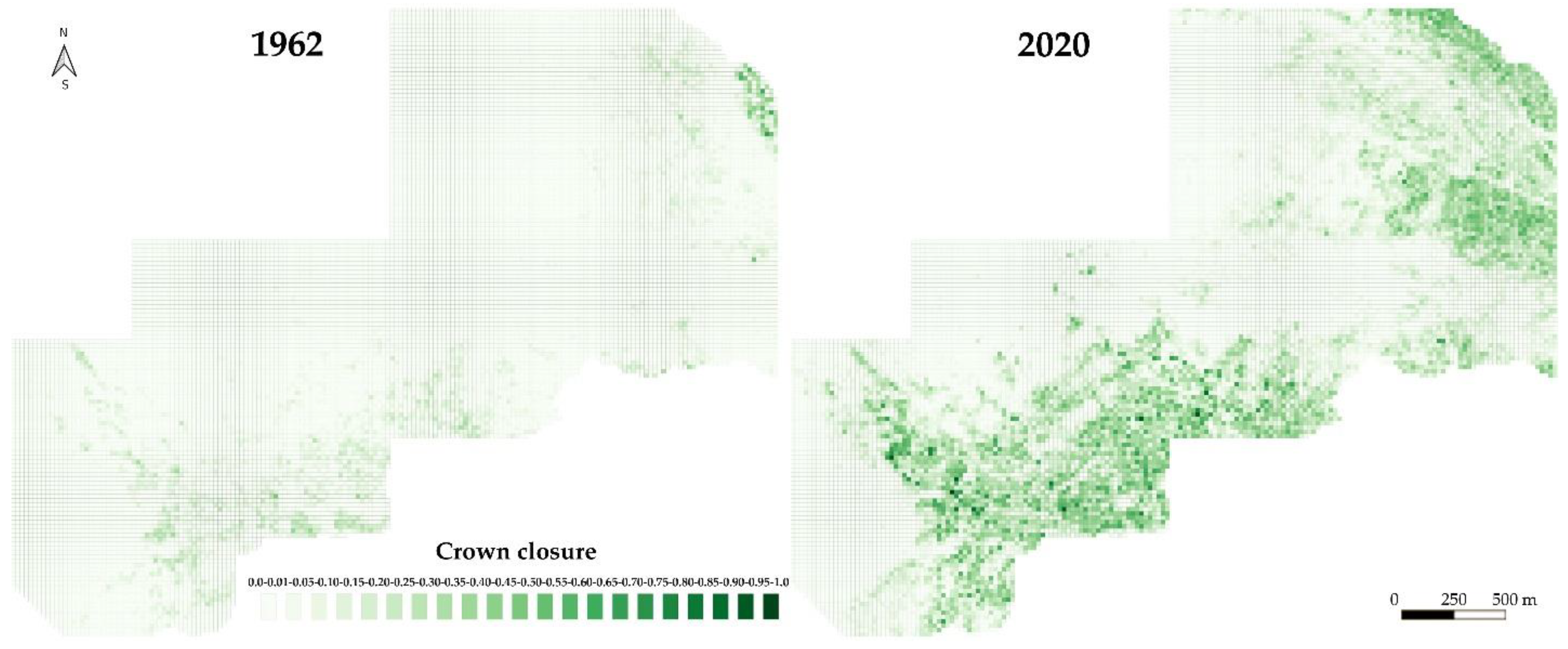
2.3. Estimation of Crown Closure by Interpreting Aerial Photos and Satellite Images
2.4. Calibration of Remote Sensing Data Using Field Observations
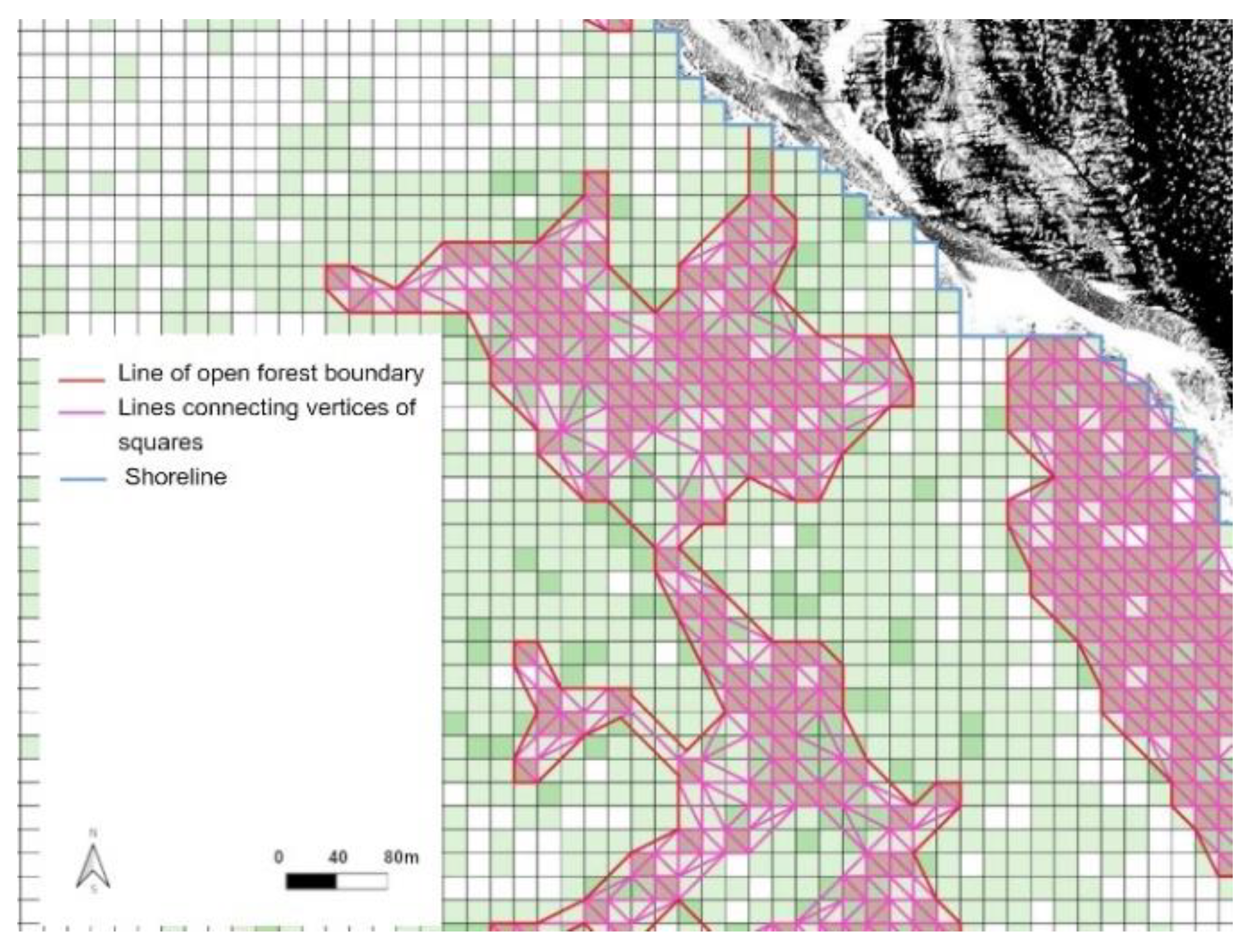
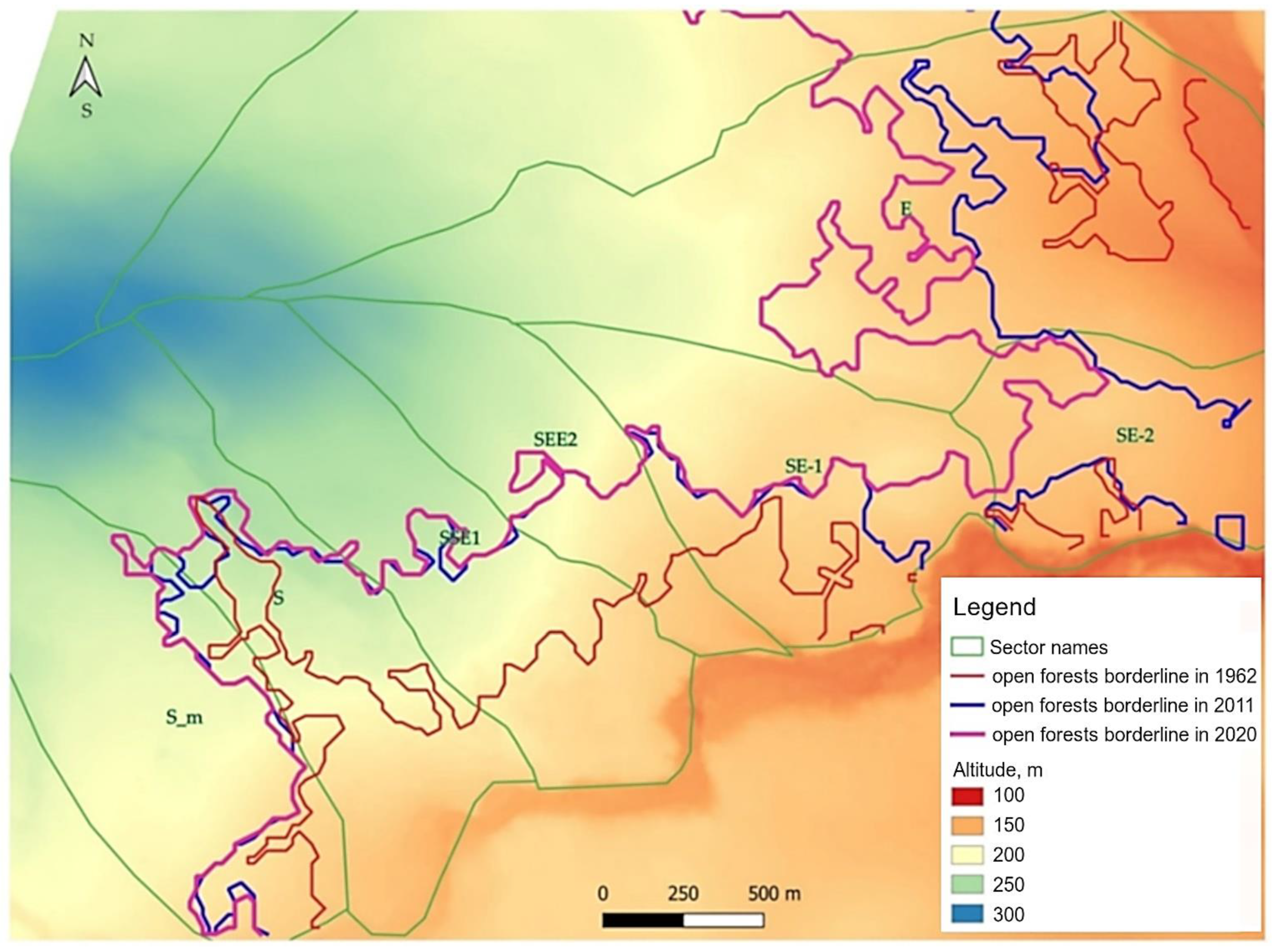
2.5. Detection of Position of Upper Boundaries of Open Forests
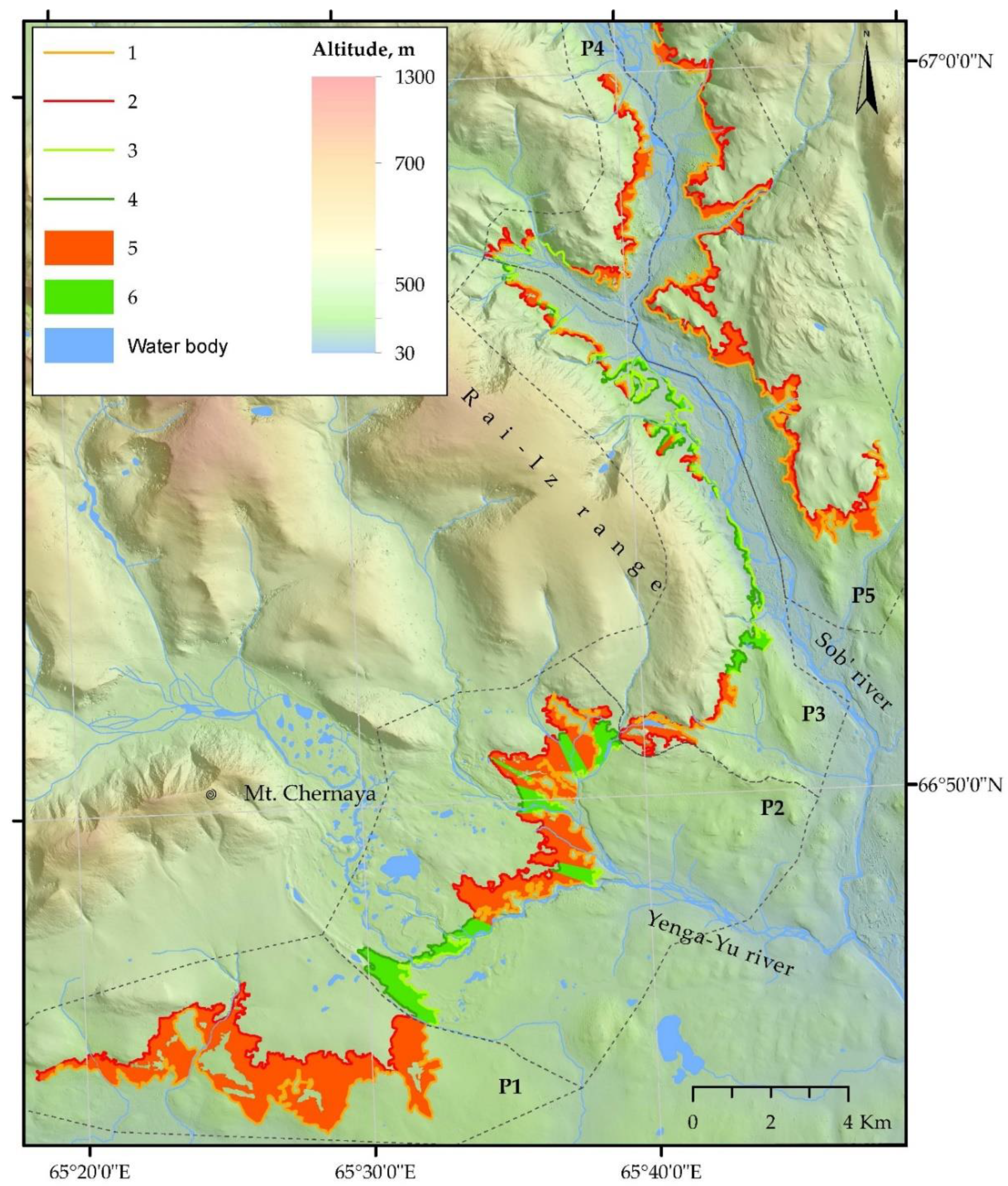
2.6. Creation of Landscape Map and Detection of Upper Open Forest Boundary
2.7. Estimation of Upper Open Forest Boundary Shift
3. Results
3.1. Tree Stand Dynamics within the Test Polygon Close to Mt. Chernaya
3.2. Characteristics of Open Forest Boundary and Its Dynamics at Studied Subregion of Polar Urals
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. 2021: Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; ISBN 978-92-9169-158-6. [Google Scholar]
- Lloyd, A.H.; Fastie, C.L. Spatial and temporal variability in the growth and climate response of treeline trees in Alaska. Clim. Chang. 2002, 52, 481–509. [Google Scholar] [CrossRef]
- Ackerly, D.D.; Cornwell, W.K.; Weiss, S.B.; Flint, L.E.; Flint, A.L. A geographic mosaic of climate change impacts on terrestrial vegetation: Which areas are most at risk? PLoS ONE 2015, 10, e130629. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. A re-assessment of high elevation treeline positions and their explanation. Oecologia 1998, 115, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Penuelas, J.; Boada, M. A global change-induced biome shift in the Montseny mountains (NE Spain). Glob. Chang. Biol. 2003, 9, 131–140. [Google Scholar] [CrossRef]
- Gehrig-Fasel, J. Treeline and Climate Change: Analyzing and Modeling Patterns and Shifts in the Swiss Alps. Ph.D. Thesis, Ecole Polytechnique, Palaiseau, France, 2007. [Google Scholar]
- Payette, S. Contrasted dynamics of Northern Labrador tree lines caused by climate change and migrational lag. Ecology 2007, 88, 770–780. [Google Scholar] [CrossRef]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A Significant Upward Shift in Plant Species Optimum Elevation During the 20th Century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- Walsh, S.J.; Butler, D.R.; Malanson, G.P.; Crews-Meyer, K.A.; Messina, J.P.; Xiao, N. Mapping, modeling, and visualization of the influences of geomorphic processes on the alpine treeline ecotone, Glacier National Park, MT, USA. Geomorphology 2003, 53, 129–145. [Google Scholar] [CrossRef]
- Danby, R.K.; Hik, D.S. Variability, contingency and rapid change in recent subarctic alpine tree line dynamics. J. Ecol. 2007, 95, 352–363. [Google Scholar] [CrossRef]
- Macias-Fauria, M.; Johnson, E.A. Warming-induced upslope advance of subalpine forest is severely limited by geomorphic processes. Proc. Natl. Acad. Sci. USA 2013, 110, 8117–8122. [Google Scholar] [CrossRef]
- Treml, V.; Chuman, T. Ecotonal Dynamics of the Altitudinal Forest Limit are Affected by Terrain and Vegetation Structure Variables: An Example from the Sudetes Mountains in Central Europe. Arct. Antarct. Alp. Res. 2015, 47, 133–146. [Google Scholar] [CrossRef]
- Holtmeier, F.K.; Broll, G. Wind as an ecological agent at treelines in North America, the Alps, and the European Subarctic. Phys. Geogr. 2010, 31, 203–233. [Google Scholar] [CrossRef]
- MacDonald, G.M.; Szeicz, J.M.; Claricoates, J.; Dale, K.A. Response of the central Canadian treeline to recent climatic changes. Ann. Assoc. Am. Geogr. 1998, 88, 183–208. [Google Scholar] [CrossRef]
- Holtmeier, F.K.; Broll, G.; Müterthies, A.; Anschlag, K. Regeneration of trees in the treeline ecotone: Northern Finnish Lapland. Fennia 2003, 181, 103–128. [Google Scholar]
- Holtmeier, F.-K.; Broll, G. Soils at the Altitudinal and Northern Treeline: European Alps, Northern Europe, Rocky Mountains—A Review. Insights For. Res. 2018, 2, 67–83. [Google Scholar] [CrossRef]
- Mckenzie, D.; Peterson, D.W.; Peterson, D.L.; Seattle, P.E.T. Climatic and biophysical controls on conifer species distributions in mountain forests of Washington State, USA. For. Sci. 2003, 30, 1093–1108. [Google Scholar] [CrossRef]
- Lloyd, A.H. Ecological Histories From Alaskan Tree Lines Provide Insight Into Future Change. Ecology 2005, 86, 1687–1695. [Google Scholar] [CrossRef]
- Kullman, L.; Öberg, L. Post-Little Ice Age tree line rise and climate warming in the Swedish Scandes: A landscape ecological perspective. J. Ecol. 2009, 97, 415–429. [Google Scholar] [CrossRef]
- Holtmeier, F.-K. Mountain Timberlines: Ecology, Patchiness, and Dynamics; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 1402097042. [Google Scholar]
- Grafius, D.; Malanson, G.; Weiss, D. Secondary controls of alpine treeline elevations in the Western USA. Phys. Geogr. 2012, 33, 146–164. [Google Scholar] [CrossRef]
- Weiss, D.J.; Malanson, G.P.; Walsh, S.J. Multiscale Relationships Between Alpine Treeline Elevation and Hypothesized Environmental Controls in the Western United States. Ann. Assoc. Am. Geogr. 2015, 105, 437–453. [Google Scholar] [CrossRef]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, R.; Luoto, M.; RäMä, T.; Mikkola, K.; Hjort, J.; Grytnes, J.A.; Birks, H.J.B. Recent vegetation changes at the high-latitude tree line ecotone are controlled by geomorphological disturbance, productivity and diversity. Glob. Ecol. Biogeogr. 2010, 19, 810–821. [Google Scholar] [CrossRef]
- Vitasse, Y.; Hoch, G.; Randin, C.F.; Lenz, A.; Kollas, C.; Körner, C. Tree recruitment of European tree species at their current upper elevational limits in the Swiss Alps. J. Biogeogr. 2012, 39, 1439–1449. [Google Scholar] [CrossRef]
- Austin, M.P.; Van Niel, K.P. Impact of landscape predictors on climate change modelling of species distributions: A case study with Eucalyptus fastigata in southern New South Wales, Australia. J. Biogeogr. 2011, 38, 9–19. [Google Scholar] [CrossRef]
- Randin, C.F.; Engler, R.; Normand, S.; Zappa, M.; Zimmermann, N.E.; Pearman, P.B.; Vittoz, P.; Thuiller, W.; Guisan, A. Climate change and plant distribution: Local models predict high-elevation persistence. Glob. Chang. Biol. 2009, 15, 1557–1569. [Google Scholar] [CrossRef]
- Trivedi, M.R.; Berry, P.M.; Morecroft, M.D.; Dawson, T.P. Spatial scale affects bioclimate model projections of climate change impacts on mountain plants. Glob. Chang. Biol. 2008, 14, 1089–1103. [Google Scholar] [CrossRef]
- Hagedorn, F.; Shiyatov, S.G.; Mazepa, V.S.; Devi, N.M.; Grigor’ev, A.A.; Bartysh, A.A.; Fomin, V.V.; Kapralov, D.S.; Terent’ev, M.; Bugman, H.; et al. Treeline advances along the Urals mountain range—Driven by improved winter conditions? Glob. Chang. Biol. 2014, 20, 3530–3543. [Google Scholar] [CrossRef]
- Grigoriev, A.A.; Shalaumova, Y.V.; Vyukhin, S.O.; Balakin, D.S.; Kukarskikh, V.V.; Vyukhina, A.A.; Camarero, J.J.; Moiseev, P.A. Upward treeline shifts in two regions of subarctic Russia are governed by summer thermal and winter snow conditions. Forests 2022, 14, 174. [Google Scholar] [CrossRef]
- Shugart, H.H.; French, N.H.F.; Kasischke, E.S.; Slawski, J.J.; Dull, C.W.; Shuchman, R.A.; Mwangi, J. Detection of vegetation change using reconnaissance imagery. Glob. Chang. Biol. 2001, 7, 247–252. [Google Scholar] [CrossRef]
- Hofgaard, A.; Tømmervik, H.; Rees, G.; Hanssen, F. Latitudinal forest advance in northernmost Norway since the early 20th century. J. Biogeogr. 2013, 40, 938–949. [Google Scholar] [CrossRef]
- Rannow, S. Do shifting forest limits in south-west Norway keep up with climate change? Scand. J. For. Res. 2013, 28, 574–580. [Google Scholar] [CrossRef]
- Greenwood, S.; Chen, J.C.; Chen, C.T.; Jump, A.S. Strong topographic sheltering effects lead to spatially complex treeline advance and increased forest density in a subtropical mountain region. Glob. Chang. Biol. 2014, 20, 3756–3766. [Google Scholar] [CrossRef] [PubMed]
- Mathisen, I.E.; Mikheeva, A.; Tutubalina, O.V.; Aune, S.; Hofgaard, A. Fifty years of tree line change in the Khibiny Mountains, Russia: Advantages of combined remote sensing and dendroecological approaches. Appl. Veg. Sci. 2014, 17, 6–16. [Google Scholar] [CrossRef]
- Beckage, B.; Osborne, B.; Gavin, D.G.; Pucko, C.; Siccama, T.; Perkins, T. A rapid upward shift of a forest ecotone during 40 years of warming in the Green Mountains of Vermont. Proc. Natl. Acad. Sci. USA 2008, 105, 4197–4202. [Google Scholar] [CrossRef] [PubMed]
- Shiyatov, S.G. Age Structure and Formation of Larch Open Forests in the Upper Timberline in the Sob’ River Basin (the Polar Urals). In Geografiya i Dinamika Rastitel’nogo Pokrova: Trudy Instituta Biologii Ural’skogo Filiala Akademii Nauk SSSR; Institut Biologii Ural’skogo Filiala Akademii Nauk SSSR: Sverdlovsk, Russia, 1965; Volume 42, pp. 81–96. [Google Scholar]
- Shiyatov, S.G.; Mazepa, V.S. Climate-driven dynamics of the forest-tundra vegetation in the Polar Ural Mountains. Contemp. Probl. Ecol. 2011, 4, 758–768. [Google Scholar] [CrossRef]
- Shiyatov, S.G. Dynamics of Upper forest Boundary on the Eastern Slope of Polar Urals; Institute of Biology of Ural Department of Academy of Sciense of SSSR: Sverdlovsk, Russia, 1964. [Google Scholar]
- Shyatov, S.G. Climate fluctuation and stand age structure of larch open forests in mounatains of the Polar Urals. In Vegetation fo Forest-Tundra and Ways of Its Development; Nauka: Lenengrad, Russia, 1967; pp. 271–278. [Google Scholar]
- Shiyatov, S.G.; Terent’ev, M.M.; Fomin, V.V.; Zimmermann, N.E. Altitudinal and horizontal shifts of the upper boundaries of open and closed forests in the Polar Urals in the 20th century. Russ. J. Ecol. 2007, 38, 223–227. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Grabherr, G. Effects of climate change on mountain ecosystems—Upward shifting of alpine plants. World Resour. Rev. 1996, 8, 382–390. [Google Scholar]
- Lamprecht, A.; Semenchuk, P.R.; Steinbauer, K.; Winkler, M.; Pauli, H. Climate change leads to accelerated transformation of high-elevation vegetation in the central Alps. New Phytol. 2018, 220, 447–459. [Google Scholar] [CrossRef]
- Holtmeier, F.K.; Broll, G. Altitudinal and polar treelines in the northern hemisphere causes and response to climate change. Polarforschung 2009, 79, 139–153. [Google Scholar] [CrossRef]
- Bourgeron, P.S.; Humphries, H.C.; Liptzin, D.; Seastedt, T.R. The forest–alpine ecotone: A multi-scale approach to spatial and temporal dynamics of treeline change at Niwot Ridge. Plant Ecol. Divers. 2015, 8, 763–779. [Google Scholar] [CrossRef]
- Dullinger, S.; Dirnböck, T.; Grabherr, G. Modelling climate change-driven treeline shifts: Relative effects of temperature increase, dispersal and invasibility. J. Ecol. 2004, 92, 241–252. [Google Scholar] [CrossRef]
- Mencuccini, M.; Piussi, P.; Sulli, A.Z. Thirty years of seed production in a subalpine Norway spruce forest: Patterns of temporal and special variation. For. Ecol. Manag. 1995, 76, 109–125. [Google Scholar] [CrossRef]
- Kroiss, S.J.; Hillerislambers, J.; D’Amato, A.W. Recruitment limitation of long-lived conifers: Implications for climate change responses. Ecology 2015, 96, 1286–1297. [Google Scholar] [CrossRef]
- Kambo, D.; Danby, R.K. Constraints on treeline advance in a warming climate: A test of the reproduction limitation hypothesis. J. Plant Ecol. 2018, 11, 411–422. [Google Scholar] [CrossRef]
- Gamache, I.; Payette, S. Latitudinal response of subarctic tree lines to recent climate change in eastern Canada. J. Biogeogr. 2005, 32, 849–862. [Google Scholar] [CrossRef]
- Trant, A.J.; Jameson, R.G.; Hermanutz, L. Variation in reproductive potential across a multi-species treeline. Arct. Antarct. Alp. Res. 2018, 50, e1524191. [Google Scholar] [CrossRef]
- Koshkina, N.B.; Moiseev, P.A.; Goryaeva, A.V. Reproduction of the Siberian spruce in the timberline ecotone of the Iremel’ Massif. Russ. J. Ecol. 2008, 39, 83–91. [Google Scholar] [CrossRef]
- Grigorieva, A.V.; Moiseev, P.A. Peculiarities and Determinants of Regeneration of Siberian Larch on the Upper Limit of Its Growth in the Urals. Contemp. Probl. Ecol. 2018, 11, 13–25. [Google Scholar] [CrossRef]
- Davis, E.L.; Hager, H.A.; Gedalof, Z. Soil properties as constraints to seedling regeneration beyond alpine treelines in the Canadian Rocky Mountains. Arct. Antarct. Alp. Res. 2018, 50, e1415625. [Google Scholar] [CrossRef]
- Lloyd, A.H.; Rupp, T.S.; Fastie, C.L.; Starfield, A.M. Patterns and dynamics of treeline advance on the Seward Peninsula, Alaska. J. Geophys. Res. 2002, 108, 8161. [Google Scholar] [CrossRef]
- Hagedorn, F.; Gavazov, K.; Alexander, J.M. Above- and belowground linkages shape responses ofmountain vegetation to climate change. Science 2019, 365, 1119–1123. [Google Scholar] [CrossRef]
- Brodersen, C.R.; Germino, M.J.; Johnson, D.M.; Reinhardt, K. Seedling Survival at Timberline Is Critical to Conifer Mountain Forest Elevation and Extent. Front. For. Glob. Chang. 2019, 2, 9. [Google Scholar] [CrossRef]
- Johnson, A.C.; Yeakley, J.A. Microsites and climate zones: Seedling regeneration in the alpine treeline ecotone worldwide. Forests 2019, 10, 864. [Google Scholar] [CrossRef]
- Lett, S.; Dorrepaal, E. Global drivers of tree seedling establishment at alpine treelines in a changing climate. Funct. Ecol. 2018, 32, 1666–1680. [Google Scholar] [CrossRef]
- Holtmeier, F.K.; Broll, G. Treeline advance—driving processes and adverse factors. Landsc. Online 2007, 1, 1–33. [Google Scholar] [CrossRef]
- Marsman, F.; Nystuen, K.O.; Opedal, Ø.H.; Foest, J.J.; Sørensen, M.V.; De Frenne, P.; Graae, B.J.; Limpens, J. Determinants of tree seedling establishment in alpine tundra. J. Veg. Sci. 2020, 31, e12948. [Google Scholar] [CrossRef]
- Kambo, D.; Danby, R.K. Factors influencing the establishment and growth of tree seedlings at Subarctic alpine treelines. Ecosphere 2018, 9, e02176. [Google Scholar] [CrossRef]
- Moiseev, P.A.; Galimova, A.A.; Bubnov, M.O.; Devi, N.M.; Fomin, V.V. Tree Stands and Their Productivity Dynamics at the Upper Growing Limit in Khibiny on the Background of Modern Climate Changes. Russ. J. Ecol. 2019, 50, 431–444. [Google Scholar] [CrossRef]
- Grigor’ev, A.A.; Devi, N.M.; Kukarskikh, V.V.; V’yukhin, S.O.; Galimova, A.A.; Moiseev, P.A.; Fomin, V.V. Structure and Dynamics of Tree Stands at the Upper Timberline in the Western Part of the Putorana Plateau. Russ. J. Ecol. 2019, 50, 311–322. [Google Scholar] [CrossRef]
- Kuyek, N.J.; Thomas, S.C. Trees are larger on south slopes in late-seral conifer stands in northwestern British Columbia. Can. J. For. Res. 2019, 49, 1349–1356. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carraro, V. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes. Oecologia 2007, 152, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springe: Basel, Switzerland, 2012; ISBN 978-3034-80395-3. [Google Scholar]
- Reger, B.; Kölling, C.; Ewald, J. Modelling effective thermal climate for mountain forests in the Bavarian Alps: Which is the best model? J. Veg. Sci. 2011, 22, 677–687. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crown closure | 0.00 | 0.01–0.05 | 0.05–0.1 | 0.11–0.15 | 0.16–0.2 | 0.21–0.25 | 0.26–0.3 | 0.31–0.35 | 0.36–0.4 | 0.41–0.45 | 0.46–0.5 | 0.51–0.55 | 0.56–0.6 | 0.61–0.65 | 0.66–0.7 | 0.71–0.75 | 0.76–0.8 | 0.81–0.85 | 0.86–0.9 | 0.91–0.95 | 0.96–1.0 |
| 1962 | 65.5 | 18.6 | 7.9 | 4.4 | 1.9 | 1.0 | 0.5 | 0.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2020 | 34.5 | 16.4 | 9.4 | 7.1 | 4.7 | 5.7 | 5.7 | 5.1 | 3.6 | 2.9 | 1.6 | 1.4 | 1.0 | 0.5 | 0.2 | 0.1 | 0.02 | 0.04 | 0.02 | 0.01 | 0.01 |
| 2020 | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Crown closure | 0.00 | 0.01–0.05 | 0.05–0.1 | 0.11–0.15 | 0.16–0.2 | 0.21–0.25 | 0.26–0.3 | 0.31–0.35 | 0.36–0.4 | 0.41–0.45 | 0.46–0.5 | 0.51–0.55 | 0.56–0.6 | 0.61–0.65 | 0.66–0.7 | 0.71–0.75 | 0.76–0.8 | 0.81–0.85 | 0.86–0.9 | 0.91–0.95 | 0.96–1.0 | |
| 1962 | 0.00 | 52.1 | 22.2 | 8.9 | 5.1 | 3.1 | 2.2 | 2.1 | 1.4 | 1.0 | 0.7 | 0.4 | 0.3 | 0.2 | 0.2 | 0.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| 0.01–0.05 | 0.0 | 10.1 | 14.4 | 13.5 | 9.0 | 11.3 | 11.1 | 9.5 | 6.9 | 5.2 | 3.2 | 2.0 | 1.9 | 1.1 | 0.5 | 0.1 | 0.1 | 0.0 | 0.0 | 0.1 | 0.0 | |
| 0.05–0.1 | 0.0 | 0.0 | 11.5 | 8.4 | 7.6 | 12.1 | 15.0 | 15.3 | 9.6 | 7.6 | 4.1 | 4.6 | 1.9 | 0.6 | 0.3 | 0.5 | 0.0 | 0.2 | 0.5 | 0.2 | 0.0 | |
| 0.11–0.15 | 0.0 | 0.0 | 0.0 | 12.5 | 5.7 | 16.7 | 13.1 | 14.5 | 11.4 | 9.6 | 3.7 | 4.5 | 4.3 | 1.1 | 1.1 | 1.1 | 0.0 | 0.3 | 0.0 | 0.0 | 0.3 | |
| 0.16–0.2 | 0.0 | 0.0 | 0.0 | 0.0 | 6.4 | 16.1 | 14.1 | 14.7 | 15.4 | 11.5 | 6.4 | 7.1 | 3.2 | 3.2 | 1.3 | 0.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 0.21–0.25 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 17.1 | 11.0 | 23.2 | 8.6 | 12.2 | 8.5 | 6.1 | 8.5 | 2.4 | 1.2 | 0.0 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | |
| 0.26–0.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 27.0 | 13.5 | 10.9 | 18.9 | 8.1 | 8.1 | 8.1 | 5.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 0.31–0.35 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 25.0 | 25.0 | 25.0 | 0.0 | 16.7 | 8.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Sector Name | Altitudinal Position (m a.s.l.) | Altitudinal Difference (m a.s.l.) | Altitudinal Shift (m 10 y−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 1962 | 2011 | 2020 | 1962 and 2011 | 2011 and 2020 | 1962 and 2020 | in 1962–2011 | in 2011–2020 | in 1962–2020 | |
| S | 220 ± 17.8 | 243 ± 12.8 | 245 ± 10.9 | 23 | 3 | 25 | 4.6 | 3.0 | 4.4 |
| SSE1 | 195 ± 6.5 | 234 ± 7.7 | 236 ± 7.3 | 40 | 1 | 41 | 8.1 | 1.4 | 7.1 |
| SEE2 | 182 ± 2.2 | 208 ± 9.0 | 214 ± 10.3 | 26 | 5 | 32 | 5.4 | 6.0 | 5.5 |
| E | 158 ± 9.2 | 172 ± 6.9 | 186 ± 7.9 | 13 | 14 | 28 | 2.7 | 15.8 | 4.7 |
| Mean for group | 189 ± 9 | 214 ± 9 | 220 ± 9 | 26 | 6 | 32 | 5.2 | 6.6 | 5.4 |
| S_m | 198 ± 16.6 | 210 ± 19.6 | 213 ± 21.7 | 12 | 3 | 14 | 2.4 | 3.2 | 2.5 |
| SE-1 | 176 ± 5.4 | 185 ± 11.2 | 189 ± 7.8 | 9 | 4 | 13 | 1.9 | 4.3 | 2.3 |
| SE-2 | 167 ± 4.7 | 168 ± 6.8 | 177 ± 2.2 | 2 | 9 | 10 | 0.3 | 9.6 | 1.7 |
| Mean for group | 180 ± 9 | 188 ± 13 | 193 ± 11 | 8 | 5 | 12 | 1.5 | 5.7 | 2.2 |
| Mean for all polygons | 185 ± 9 | 203 ± 11 | 209 ± 10 | 18 | 6 | 23 | 3.6 | 6.2 | 4.0 |
| Sector Name | Absolute Horizontal Shift (m) | Horizontal Shift per 10 Years (m) | ||||
|---|---|---|---|---|---|---|
| 1962–2011 | 2011–2020 | 1962–2020 | 1962–2011 | 2011–2020 | 1962–2020 | |
| S | 152 ± 135 | 21 ± 30 | 167 ± 132 | 30.9 | 23.8 | 28.8 |
| SSE1 | 344 ± 43 | 6 ± 11 | 356 ± 47 | 70.1 | 6.4 | 61.4 |
| SEE2 | 260 ± 56 | 1 ± 4 | 268 ± 53 | 53.0 | 1.3 | 46.2 |
| E | 131 ± 112 | 194 ± 160 | 478 ± 144 | 26.6 | 215.9 | 82.3 |
| S_m | 62 ± 57 | 9 ± 19 | 67 ± 59 | 12.6 | 10.3 | 11.6 |
| SE-1 | 126 ± 86 | 45 ± 74 | 193 ± 117 | 25.7 | 50.2 | 33.3 |
| SE-2 | 36 ± 47 | 229 ± 188 | 254 ± 145 | 7.4 | 254.2 | 43.8 |
| Part of Study Area | Portion of Total Length | Altitude in 2020 (m asl) | Distribution of Slope Inclination (Degrees) | Distribution of Slope Exposition | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Min | Max | 0–10 | 10–20 | 20–30 | N | NE | E | SE | S | SW | W | NW | ||
| Slopes without wide spread of moraine deposits and boulder fields | ||||||||||||||
| 1 | 1 | 146 | 279 | 0.97 | 0.03 | 0.00 | 0.01 | 0.05 | 0.10 | 0.20 | 0.34 | 0.23 | 0.05 | 0.02 |
| 2 | 0.66 | 136 | 330 | 0.92 | 0.07 | 0.01 | 0.04 | 0.13 | 0.18 | 0.24 | 0.27 | 0.11 | 0.02 | 0.01 |
| 3 | 0.36 | 85 | 311 | 0.66 | 0.29 | 0.05 | 0.16 | 0.23 | 0.24 | 0.12 | 0.07 | 0.09 | 0.05 | 0.04 |
| 4 | 0.82 | 92 | 243 | 0.37 | 0.48 | 0.12 | 0.04 | 0.22 | 0.43 | 0.15 | 0.07 | 0.06 | 0.02 | 0.01 |
| 5 | 1 | 97 | 288 | 0.42 | 0.35 | 0.16 | 0.03 | 0.04 | 0.08 | 0.10 | 0.17 | 0.24 | 0.24 | 0.10 |
| Slopes with a prevalence of moraine deposits and boulder fields | ||||||||||||||
| 1 | 0 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| 2 | 0.34 | 145 | 253 | 0.91 | 0.08 | 0.01 | 0.06 | 0.23 | 0.18 | 0.18 | 0.20 | 0.11 | 0.02 | 0.02 |
| 3 | 0.64 | 73 | 235 | 0.76 | 0.20 | 0.03 | 0.15 | 0.30 | 0.25 | 0.10 | 0.05 | 0.05 | 0.04 | 0.06 |
| 4 | 0.18 | 140 | 228 | 0.53 | 0.36 | 0.08 | 0.08 | 0.17 | 0.11 | 0.13 | 0.21 | 0.12 | 0.10 | 0.08 |
| 5 | 0 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Part of Subregion | Altitudinal Position (m a.s.l.) in | Altitudinal Differences for Period 1960–2020 (m a.s.l.) | Shift in 1960–2020 (m 10 year−1) | Area Occupied by Open and Closed Forests (km2) | ||
|---|---|---|---|---|---|---|
| 1960 | 2020 | Vertical | Horizontal | |||
| Slopes with slight influences of edaphic constrains | ||||||
| 1 | 202 ± 28 | 244 ± 23 | 42 | 7.0 | 48.2 | 9.81 |
| 2 | 199 ± 39 | 231 ± 38 | 32 | 5.3 | 33.7 | 5.39 |
| 3 | 213 ± 61 | 238 ± 62 | 25 | 4.2 | 25.5 | 1.54 |
| 4 | 162 ± 39 | 200 ± 35 | 38 | 6.3 | 14.7 | 1.37 |
| 5 | 207 ± 39 | 238 ± 42 | 31 | 5.2 | 19.8 | 4.33 |
| All | 200 ± 42 | 233 ± 43 | 33 | 5.5 | 35.5 | 22.4 |
| Slopes with strong influences of edaphic constrains | ||||||
| 1 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| 2 | 194 ± 25 | 226 ± 29 | 32 | 5.3 | 41.3 | 3.59 |
| 3 | 119 ± 36 | 130 ± 42 | 11 | 1.8 | 13.8 | 1.71 |
| 4 | 169 ± 18 | 169 ± 20 | 0 | 0.0 | 3.2 | 0.03 |
| 5 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| All | 149 ± 47 | 158 ± 57 | 9 | 1.5 | 32.8 | 5.33 |
| Part of Subregion | 1 | 2 | 3 | 4 | 5 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Year | 1960 | 2020 | 1960 | 2020 | 1960 | 2020 | 1960 | 2020 | 1960 | 2020 |
| Slopes slightly influence by edaphic constrains | ||||||||||
| Prevail slope exposure | S, SW | SE, S | NE, E | NE, E | SW, W | |||||
| Mean | 211 | 238 | 204 | 234 | 179 | 205 | 160 | 197 | 204 | 234 |
| SD | 43 | 24 | 36 | 35 | 42 | 60 | 36 | 36 | 42 | 46 |
| Slopes strongly influence with edaphic constrains | ||||||||||
| Prevail slope exposure | n.d. | S, SW | NE, E | NE, S | n.d. | |||||
| Mean | n.d. | n.d. | 197 | 240 | 120 | 131 | 167 | 167 | n.d. | n.d. |
| SD | n.d. | n.d. | 32 | 34 | 36 | 42 | 14 | 16 | n.d. | n.d. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nizametdinov, N.F.; Shalaumova, Y.V.; Mazepa, V.S.; Moiseev, P.A. Assessment of Past Decadal Dynamics of Tree Stands in Forest–Tundra Transition Zone on the Polar Ural Mountains Calibrated Using Historical and Modern Field Measurements. Forests 2022, 13, 2107. https://doi.org/10.3390/f13122107
Nizametdinov NF, Shalaumova YV, Mazepa VS, Moiseev PA. Assessment of Past Decadal Dynamics of Tree Stands in Forest–Tundra Transition Zone on the Polar Ural Mountains Calibrated Using Historical and Modern Field Measurements. Forests. 2022; 13(12):2107. https://doi.org/10.3390/f13122107
Chicago/Turabian StyleNizametdinov, Nail’ F., Yulia V. Shalaumova, Valery S. Mazepa, and Pavel A. Moiseev. 2022. "Assessment of Past Decadal Dynamics of Tree Stands in Forest–Tundra Transition Zone on the Polar Ural Mountains Calibrated Using Historical and Modern Field Measurements" Forests 13, no. 12: 2107. https://doi.org/10.3390/f13122107
APA StyleNizametdinov, N. F., Shalaumova, Y. V., Mazepa, V. S., & Moiseev, P. A. (2022). Assessment of Past Decadal Dynamics of Tree Stands in Forest–Tundra Transition Zone on the Polar Ural Mountains Calibrated Using Historical and Modern Field Measurements. Forests, 13(12), 2107. https://doi.org/10.3390/f13122107