
Influence of Leaf Litter and Humus Composition on the Development of Black Spruce Seedlings: A Greenhouse Experimentation



Abstract
1. Introduction
2. Materials and Methods
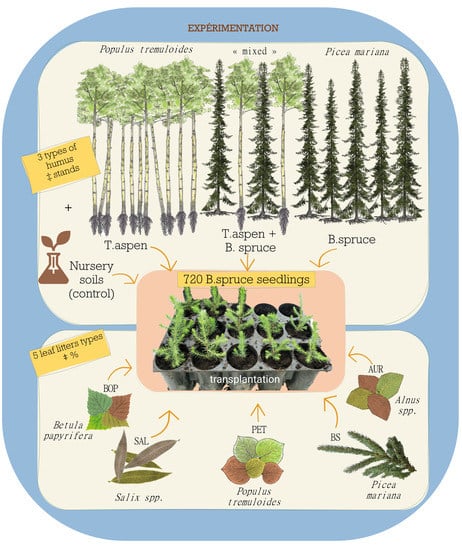
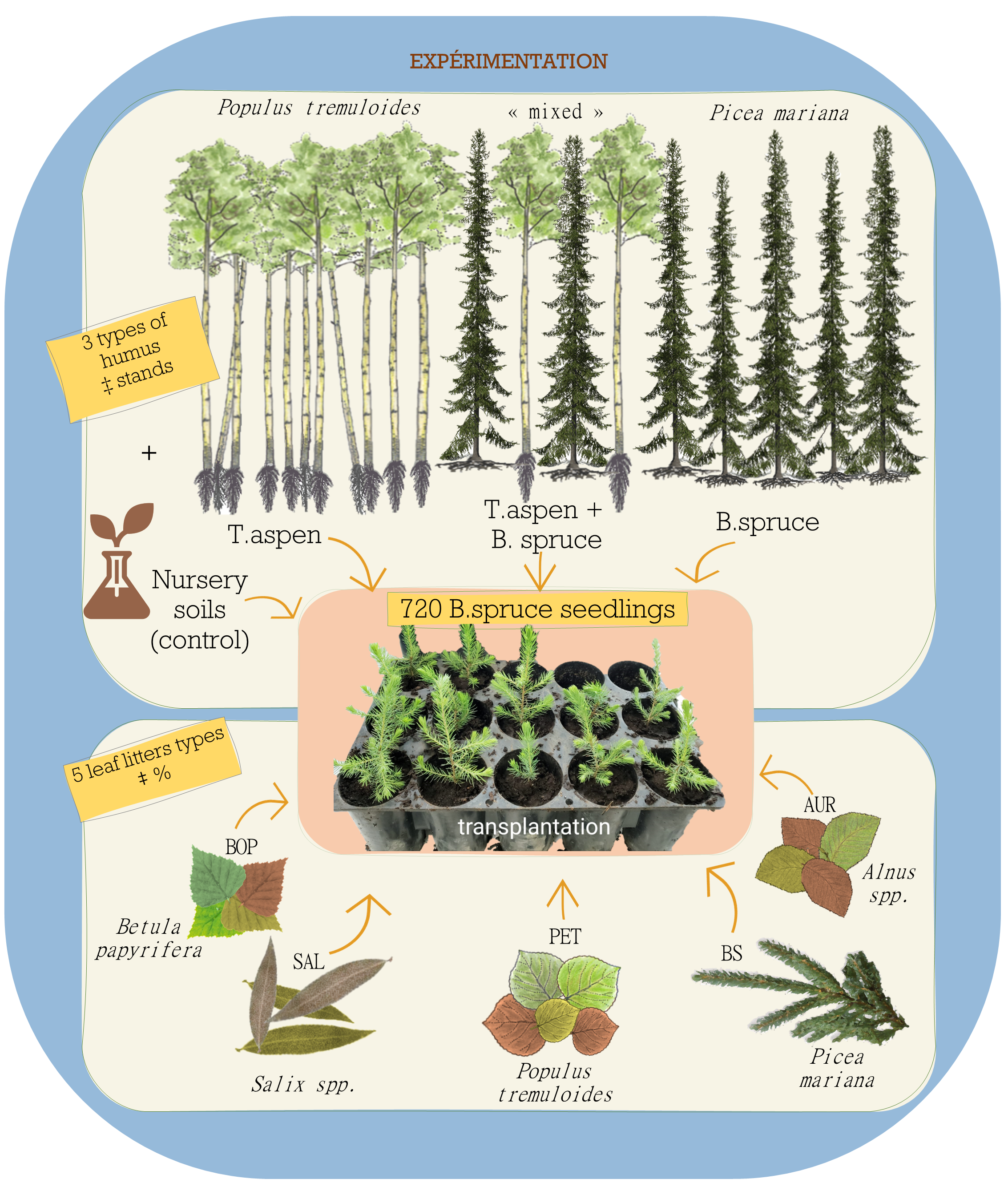
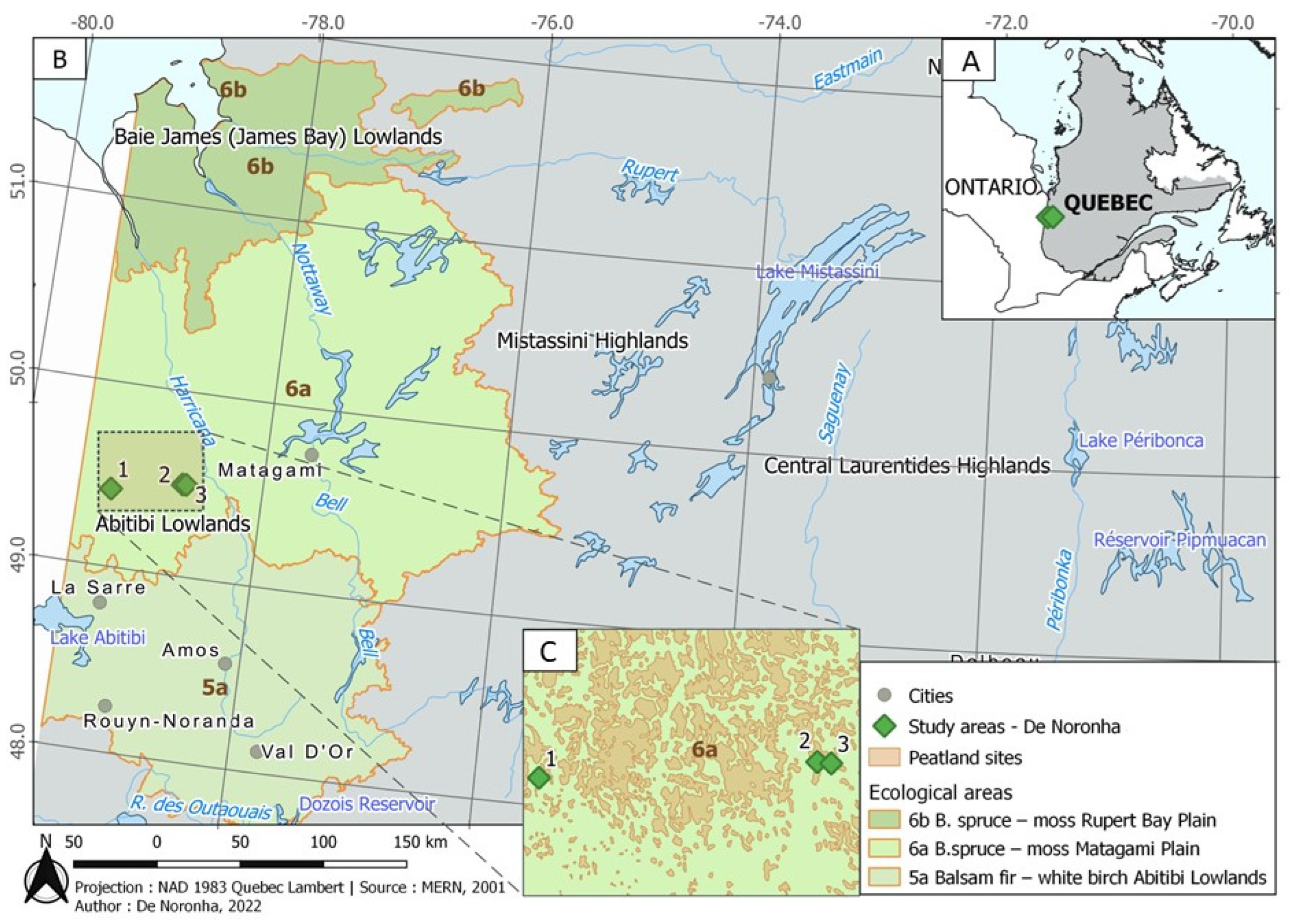
2.1. Growing Substrate Source
2.2. Greenhouse Experiment and Sampling Design
2.3. Statistical Analysis
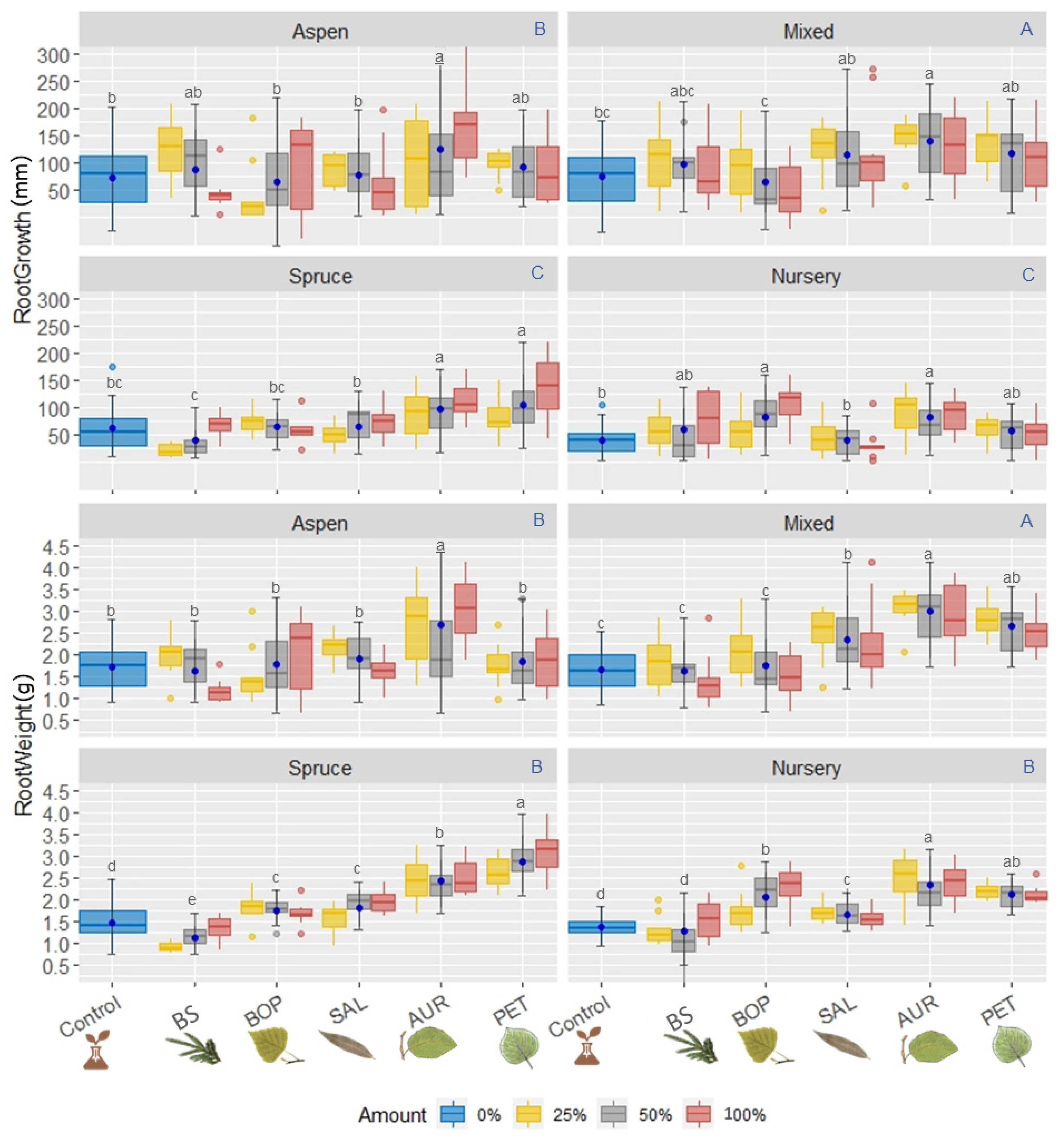
3. Results
3.1. Leaf Litter and Humus Nutrients before Treatment
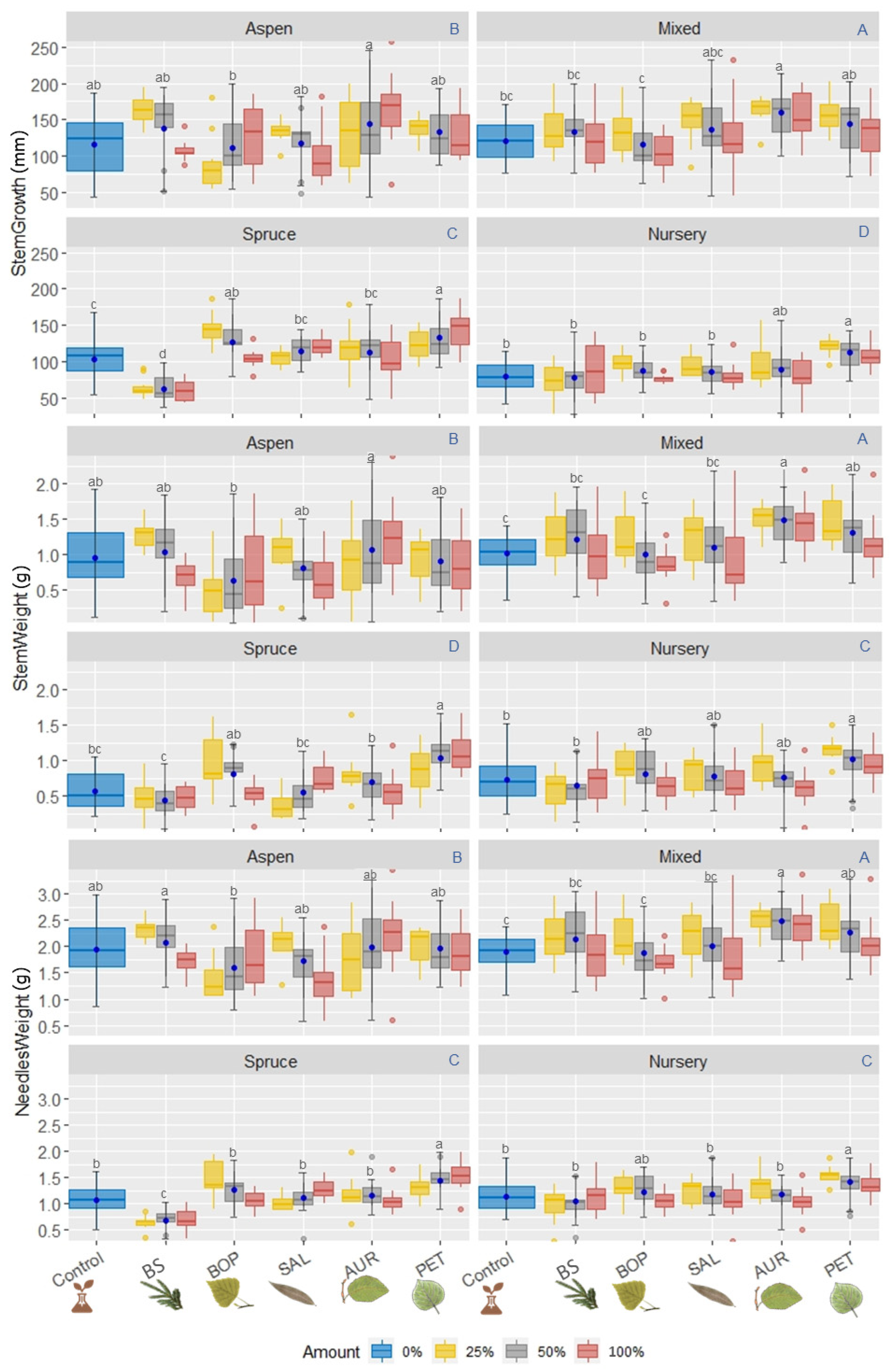
3.2. Seedling Development and Humus Characteristics
3.2.1. Aerial Parts of B. spruce Seedlings
3.2.2. Root Development
Root Growth
Root Weight
3.3. Humus pH
3.4. Humus Moisture Content
3.5. Foliage Nutrient Concentrations after Experiment
3.6. Humus Substrate Composition at the End of Experiment
4. Discussion
4.1. Humus Type Effect
4.2. Litter Type and Litter Addition Rate Effect
4.3. Management Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brassard, B.W.; Chen, H.Y.; Bergeron, Y.; Paré, D. Differences in Fine Root Productivity between Mixed-and Single-Species Stands. Funct. Ecol. 2011, 25, 238–246. [Google Scholar] [CrossRef]
- Levia, D.F.; Herwitz, S.R. Interspecific Variation of Bark Water Storage Capacity of Three Deciduous Tree Species in Relation to Stemflow Yield and Solute Flux to Forest Soils. Catena 2005, 64, 117–137. [Google Scholar] [CrossRef]
- Crispo, M.; Jean, M.; Fenton, N.J.; Leduc, A.; Bergeron, Y. Factors Explaining the Composition and Diversity of Vascular Plant Understories along a Transcontinental Gradient in the Canadian Boreal Forest. J. Veg. Sci. 2021, 32, e13058. [Google Scholar] [CrossRef]
- Chávez, V.; Macdonald, S.E. Partitioning Vascular Understory Diversity in Mixedwood Boreal Forests: The Importance of Mixed Canopies for Diversity Conservation. For. Ecol. Manag. 2012, 271, 19–26. [Google Scholar] [CrossRef]
- Messier, C.; Parent, S.; Bergeron, Y. Effects of Overstory and Understory Vegetation on the Understory Light Environment in Mixed Boreal Forests. J. Veg. Sci. 1998, 9, 511–520. [Google Scholar] [CrossRef]
- Mestre, L.; Toro-Manríquez, M.; Soler, R.; Huertas-Herrera, A.; Martínez-Pastur, G.; Lencinas, M.V. The Influence of Canopy-Layer Composition on Understory Plant Diversity in Southern Temperate Forests. For. Ecosyst. 2017, 4, 6. [Google Scholar] [CrossRef]
- Ghotsa Mekontchou, C.; Houle, D.; Bergeron, Y.; Drobyshev, I. Contrasting Root System Structure and Belowground Interactions between Black Spruce (Picea Mariana (Mill.) BSP) and Trembling Aspen (Populus Tremuloides Michx) in Boreal Mixedwoods of Eastern Canada. Forests 2020, 11, 127. [Google Scholar] [CrossRef]
- Adamczyk, B.; Sietiö, O.-M.; Straková, P.; Prommer, J.; Wild, B.; Hagner, M.; Pihlatie, M.; Fritze, H.; Richter, A.; Heinonsalo, J. Plant Roots Increase Both Decomposition and Stable Organic Matter Formation in Boreal Forest Soil. Nat. Commun. 2019, 10, 3982. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, B.; Liu, Z. Impacts of Plant Secondary Metabolites from Conifer Litter on the Decomposition of Populus Purdomii Litter. J. For. Res. 2019, 30, 2237–2245. [Google Scholar] [CrossRef]
- Initial Litter Chemical Composition. In Plant Litter: Decomposition, Humus Formation, Carbon Sequestration; Berg, B., McClaugherty, C., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 67–100. ISBN 978-3-030-59631-6. [Google Scholar]
- Berg, B.; Laskowski, R.; Santo, A.V. Estimated Nitrogen Concentrations in Humus Based on Initial Nitrogen Concentrations in Foliar Litter: A Synthesis. XII. Long-Term Decomposition in a Scots Pine Forest. Can. J. Bot. 2000, 77, 1712–1722. [Google Scholar] [CrossRef]
- Staaf, H. Plant Nutrient Changes in Beech Leaves during Senescence as Influenced by Site Characteristics. Acta Oecol. Oecol. Plant 1982, 3, 161–170. [Google Scholar]
- Priputina, I.V.; Frolova, G.G.; Shanin, V.N.; Myakshina, T.N.; Grabarnik, P.Y. Spatial Distribution of Organic Matter and Nitrogen in the Entic Podzols of the Prioksko-Terrasnyi Reserve and Its Relationship with the Structure of Forest Phytocenoses. Eurasian Soil Sci. 2020, 53, 1021–1032. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, J.C.; Bergeron, Y.; Kembel, S.W.; Fenton, N.J. Dominance of Coniferous and Broadleaved Trees Drives Bacterial Associations with Boreal Feather Mosses. bioRxiv, 2022; bioRxiv preprint. [Google Scholar] [CrossRef]
- Saetre, P. Spatial Patterns of Ground Vegetation, Soil Microbial Biomass and Activity in a Mixed Spruce-Birch Stand. Ecography 1999, 22, 183–192. [Google Scholar] [CrossRef]
- Jean, M.; Holland-Moritz, H.; Melvin, A.M.; Johnstone, J.F.; Mack, M.C. Experimental Assessment of Tree Canopy and Leaf Litter Controls on the Microbiome and Nitrogen Fixation Rates of Two Boreal Mosses. New Phytol. 2020, 227, 1335–1349. [Google Scholar] [CrossRef] [PubMed]
- Rinnan, R.; Michelsen, A.; Jonasson, S. Effects of Litter Addition and Warming on Soil Carbon, Nutrient Pools and Microbial Communities in a Subarctic Heath Ecosystem. Appl. Soil Ecol. 2008, 39, 271–281. [Google Scholar] [CrossRef]
- Vesterdal, L.; Schmidt, I.K.; Callesen, I.; Nilsson, L.O.; Gundersen, P. Carbon and Nitrogen in Forest Floor and Mineral Soil under Six Common European Tree Species. For. Ecol. Manag. 2008, 255, 35–48. [Google Scholar] [CrossRef]
- Forest Regions. The Canadian Encyclopedia. Available online: https://www.thecanadianencyclopedia.ca/en/article/forest-regions (accessed on 17 January 2022).
- Canada, N.R. State-Canadas-Forests-Report. Available online: https://www.nrcan.gc.ca/our-natural-resources/forests/state-canadas-forests-report/16496 (accessed on 30 April 2022).
- Comeau, P.G.; Thomas, K.D. Silviculture of Temperate and Boreal Broadleaf-Conifer Mixtures. In Land Management Handbook; No. 36; Technical Report; Government of British Columbia: Victoria, BC, Canada, 1996. [Google Scholar]
- Légaré, S.; Paré, D.; Bergeron, Y. The Responses of Black Spruce Growth to an Increased Proportion of Aspen in Mixed Stands. Can. J. For. Res. 2004, 34, 405–416. [Google Scholar] [CrossRef]
- Fenton, N.; Bergeron, Y. Dynamic Old-Growth Forests? A Case Study of Boreal Black Spruce Forest Bryophytes. Silva Fenn. 2011, 45, 983–994. [Google Scholar] [CrossRef]
- Qian, H.; Klinka, K.; Økland, R.H.; Krestov, P.; Kayahara, G.J. Understorey Vegetation in Boreal Picea Mariana and Populus Tremuloides Stands in British Columbia. J. Veg. Sci. 2003, 14, 173–184. [Google Scholar] [CrossRef]
- Pacé, M.; Fenton, N.J.; Paré, D.; Bergeron, Y. Ground-Layer Composition Affects Tree Fine Root Biomass and Soil Nutrient Availability in Jack Pine and Black Spruce Forests under Extreme Drainage Conditions. Can. J. For. Res. 2017, 47, 433–444. [Google Scholar] [CrossRef]
- Comeau, P.G. Relationships between Stand Parameters and Understorey Light in Boreal Aspen Stands. J. Ecosyst. Manag. 2002, 1, 2. [Google Scholar]
- Kumar, P.; Chen, H.Y.; Searle, E.B.; Shahi, C. Dynamics of Understorey Biomass, Production and Turnover Associated with Long-Term Overstorey Succession in Boreal Forest of Canada. For. Ecol. Manag. 2018, 427, 152–161. [Google Scholar] [CrossRef]
- Hart, S.A.; Chen, H.Y. Understory Vegetation Dynamics of North American Boreal Forests. Crit. Rev. Plant Sci. 2006, 25, 381–397. [Google Scholar] [CrossRef]
- Canada, L. A Search—Theses Canada. Available online: https://www.bac-lac.gc.ca/eng/services/theses/Pages/item.aspx?idNumber=1033011249 (accessed on 15 March 2022).
- Liu, T.-Y.; Lin, K.-C.; Vadeboncoeur, M.A.; Chen, M.-Z.; Huang, M.-Y.; Lin, T.-C. Understorey Plant Community and Light Availability in Conifer Plantations and Natural Hardwood Forests in Taiwan. Appl. Veg. Sci. 2015, 18, 591–602. [Google Scholar] [CrossRef]
- Kristensen, H.L.; Gundersen, P.; Callesen, I.; Reinds, G.J. Throughfall Nitrogen Deposition Has Different Impacts on Soil Solution Nitrate Concentration in European Coniferous and Deciduous Forests. Ecosystems 2004, 7, 180–192. [Google Scholar] [CrossRef]
- Gundersen, P.; Sevel, L.; Christiansen, J.R.; Vesterdal, L.; Hansen, K.; Bastrup-Birk, A. Do Indicators of Nitrogen Retention and Leaching Differ between Coniferous and Broadleaved Forests in Denmark? For. Ecol. Manag. 2009, 258, 1137–1146. [Google Scholar] [CrossRef]
- Viereck, L.A.; Dyrness, C.T.; Cleve, K.V.; Foote, M.J. Vegetation, Soils, and Forest Productivity in Selected Forest Types in Interior Alaska. Can. J. For. Res. 1983, 13, 703–720. [Google Scholar] [CrossRef]
- Tamm, C.O. Nitrogen-Limited and Nitrogen-Depleted Terrestrial Ecosystems: Ecological Characteristics. In Nitrogen in Terrestrial Ecosystems; Springer: Berlin/Heidelberg, Germany, 1991; pp. 34–49. [Google Scholar]
- Farooq, T.H.; Chen, X.; Shakoor, A.; Li, Y.; Wang, J.; Rashid, M.H.U.; Kumar, U.; Yan, W. Unraveling the Influence of Land-Use Change on Δ13C, Δ15N, and Soil Nutritional Status in Coniferous, Broadleaved, and Mixed Forests in Southern China: A Field Investigation. Plants 2021, 10, 1499. [Google Scholar] [CrossRef]
- Cavard, X.; Bergeron, Y.; Chen, H.Y.; Paré, D.; Laganière, J.; Brassard, B. Competition and Facilitation between Tree Species Change with Stand Development. Oikos 2011, 120, 1683–1695. [Google Scholar] [CrossRef]
- Simard, M.; Lecomte, N.; Bergeron, Y.; Bernier, P.Y.; Paré, D. Forest Productivity Decline Caused by Successional Paludification of Boreal Soils. Ecol. Appl. 2007, 17, 1619–1637. [Google Scholar] [CrossRef]
- Payette, S.; Rochefort, L. Écologie Des Tourbières Du Québec-Labrador; Presses Université Laval: Québec, QC, Canada, 2001. [Google Scholar]
- Paré, D.; Banville, J.L.; Garneau, M.; Bergeron, Y. Soil Carbon Stocks and Soil Carbon Quality in the Upland Portion of a Boreal Landscape, James Bay, Quebec. Ecosystems 2011, 14, 533–546. [Google Scholar] [CrossRef]
- Magnan, G.; Le Stum-Boivin, É.; Garneau, M.; Grondin, P.; Fenton, N.; Bergeron, Y. Holocene Vegetation Dynamics and Hydrological Variability in Forested Peatlands of the Clay Belt, Eastern Canada, Reconstructed Using a Palaeoecological Approach. Boreas 2019, 48, 131–146. [Google Scholar] [CrossRef]
- Simard, M.; Bernier, P.Y.; Bergeron, Y.; Pare, D.; Guérine, L. Paludification Dynamics in the Boreal Forest of the James Bay Lowlands: Effect of Time since Fire and Topography. Can. J. For. Res. 2009, 39, 546–552. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Messier, C.; Stadt, K.J.; Gendron, F.; Comeau, P.G. Predicting and Managing Light in the Understory of Boreal Forests. Can. J. For. Res. 1999, 29, 796–811. [Google Scholar] [CrossRef]
- Ouimet, R.; Duchesne, L.; Tremblay, S. Long-Term Soil Fertility and Site Productivity in Stem-Only and Whole-Tree Harvested Stands in Boreal Forest of Quebec (Canada). Forests 2021, 12, 583. [Google Scholar] [CrossRef]
- Trettin, C.C.; Jurgensen, M.F.; Gale, M.R.; McLaughlin, J.W. Soil Carbon in Northern Forested Wetlands: Impacts of Silvicultural Practices. Carbon Forms Funct. For. Soils 1995, 53711, 437–461. [Google Scholar]
- Prescott, C.E. Effects of Clearcutting and Alternative Silvicultural Systems on Rates of Decomposition and Nitrogen Mineralization in a Coastal Montane Coniferous Forest. For. Ecol. Manag. 1997, 95, 253–260. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; Huang, Y. Comparisons of Litterfall, Litter Decomposition and Nutrient Return in a Monoculture Cunninghamia Lanceolata and a Mixed Stand in Southern China. For. Ecol. Manag. 2008, 255, 1210–1218. [Google Scholar] [CrossRef]
- Wu, W.; Zhou, X.; Wen, Y.; Zhu, H.; You, Y.; Qin, Z.; Li, Y.; Huang, X.; Yan, L.; Li, H. Coniferous-Broadleaf Mixture Increases Soil Microbial Biomass and Functions Accompanied by Improved Stand Biomass and Litter Production in Subtropical China. Forests 2019, 10, 879. [Google Scholar] [CrossRef]
- Prescott, C.E.; Zabek, L.M.; Staley, C.L.; Kabzems, R. Decomposition of Broadleaf and Needle Litter in Forests of British Columbia: Influences of Litter Type, Forest Type, and Litter Mixtures. Can. J. For. Res. 2000, 30, 1742–1750. [Google Scholar] [CrossRef]
- Vanguelova, E.I.; Pitman, R.M. Nutrient and Carbon Cycling along Nitrogen Deposition Gradients in Broadleaf and Conifer Forest Stands in the East of England. For. Ecol. Manag. 2019, 447, 180–194. [Google Scholar] [CrossRef]
- Taylor, B.R.; Parsons, W.F.; Parkinson, D. Decomposition of Populus Tremuloides Leaf Litter Accelerated by Addition of Alnus Crispa Litter. Can. J. For. Res. 1989, 19, 674–679. [Google Scholar] [CrossRef]
- Augusto, L.; De Schrijver, A.; Vesterdal, L.; Smolander, A.; Prescott, C.; Ranger, J. Influences of Evergreen Gymnosperm and Deciduous Angiosperm Tree Species on the Functioning of Temperate and Boreal Forests. Biol. Rev. 2015, 90, 444–466. [Google Scholar] [CrossRef] [PubMed]
- Légaré, S.; Paré, D.; Bergeron, Y. Influence of Aspen on Forest Floor Properties in Black Spruce-Dominated Stands. Plant Soil 2005, 275, 207–220. [Google Scholar] [CrossRef]
- Fradette, J. Effet de La Proximité Des Feuillus Sur La Croissance de l’épinette Noire et de l’épinette Blanche Dans Un Contexte de Plantation: Le Dégagement Est-Il Toujours Souhaitable? Ph.D. Thesis, Université du Québec en Abitibi-Témiscamingue, Rouyn Noranda, QC, Canada, 2014. [Google Scholar]
- Felton, A.; Nilsson, U.; Sonesson, J.; Felton, A.M.; Roberge, J.-M.; Ranius, T.; Ahlström, M.; Bergh, J.; Björkman, C.; Boberg, J. Replacing Monocultures with Mixed-Species Stands: Ecosystem Service Implications of Two Production Forest Alternatives in Sweden. Ambio 2016, 45, 124–139. [Google Scholar] [CrossRef]
- Asplund, J.; Hustoft, E.; Nybakken, L.; Ohlson, M.; Lie, M.H. Litter Impair Spruce Seedling Emergence in Beech Forests: A Litter Manipulation Experiment. Scand. J. For. Res. 2018, 33, 332–337. [Google Scholar] [CrossRef]
- Peterson, C.J.; Facelli, J.M. Contrasting Germination and Seedling Growth of Betula Alleghaniensis and Rhus Typhina Subjected to Various Amounts and Types of Plant Litter. Am. J. Bot. 1992, 79, 1209–1216. [Google Scholar] [CrossRef]
- Morsing, J.; Kepfer-Rojas, S.; Baastrup-Spohr, L.; Rodriguez, A.L.; Raulund-Rasmussen, K. Litter Legacy after Spruce Plantation Removal Hampers Initial Vegetation Establishment. Basic Appl. Ecol. 2020, 42, 4–14. [Google Scholar] [CrossRef]
- Uselman, S.M.; Qualls, R.G.; Lilienfein, J. Quality of Soluble Organic C, N, and P Produced by Different Types and Species of Litter: Root Litter versus Leaf Litter. Soil Biol. Biochem. 2012, 54, 57–67. [Google Scholar] [CrossRef]
- Wieder, W.R.; Cleveland, C.C.; Townsend, A.R. Tropical Tree Species Composition Affects the Oxidation of Dissolved Organic Matter from Litter. Biogeochemistry 2008, 88, 127–138. [Google Scholar] [CrossRef]
- Natural Regions. The Canadian Encyclopedia. Available online: https://www.thecanadianencyclopedia.ca/en/article/natural-regions (accessed on 14 January 2022).
- Blouin, J.; Berger, J.-P.; Gosselin, J.; Québec (Province); Direction des Inventaires Forestiers; Québec (Province); Ministère des Ressources Naturelles; de la Faune et des Parcs. Direction des Communications Guide de Reconnaissance des Types Écologiques: Région Écologique 6a, Plaine du Lac Matagami: Région Écologique 6b, Plaine de la Baie de Rupert; Direction des Communications; Ministére des Ressources Naturelles et de la Faune, Forêt Québec, Direction des Inventaires Forestiers: Québec, QC, Canada, 2005; ISBN 978-2-551-22842-3. [Google Scholar]
- Description of Natural Provinces—Province F—Abitibi and James Bay Lowlands. Available online: https://www.environnement.gouv.qc.ca/biodiversite/aires_protegees/provinces/partie4f.htm (accessed on 14 January 2022).
- Sommaire Des Normales Climatiques 1981–2010—Ministère Du Développement Durable, de l’Environnement, de La Faune et Des Parcs. Available online: https://www.environnement.gouv.qc.ca/climat/normales/sommaire.asp?cle=7093376 (accessed on 14 January 2022).
- Tran, T.S.; Simard, R.R. Mehlich III-Extractable Elements. Soil Sampl. Methods Anal. 1993, 43, 49. [Google Scholar]
- Hossner, L.R. Dissolution for Total Elemental Analysis. Methods Soil Anal. Part 3 Chem. Methods 1996, 5, 49–64. [Google Scholar]
- Jones, J.B., Jr.; Case, V.W. Sampling, Handling, and Analyzing Plant Tissue Samples. Soil Test. Plant Anal. 1990, 3, 389–427. [Google Scholar]
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.L.; Takahashi, K.; Wilke, C.; Woo, K.; Yutani, H.; Dunnington, D. RStudio Ggplot2: Create Elegant Data Visualisations Using the Grammar of Graphics. 2021. Available online: https://ggplot2.tidyverse.org/reference/ggplot2-package.html (accessed on 14 January 2022).
- Wickham, H.; François, R.; Henry, L.; Müller, K. RStudio Dplyr: A Grammar of Data Manipulation. 2022. Available online: http://stat599.cwick.co.nz/lectures/02-dplyr.pdf (accessed on 14 January 2022).
- R-core Nlme: Linear and Nonlinear Mixed Effects Models 2022. Available online: https://cran.r-project.org/web/packages/nlme/index.html (accessed on 2 November 2022).
- Hothorn, T.; Bretz, F.; Westfall, P.; Heiberger, R.M.; Schuetzenmeister, A.; Scheibe, S. Multcomp: Simultaneous Inference in General Parametric Models. 2022. Available online: https://cran.r-project.org/web/packages/multcomp/vignettes/generalsiminf.pdf (accessed on 20 January 2022).
- Chomel, M.; Guittonny-Larchevêque, M.; DesRochers, A.; Baldy, V. Effect of Mixing Herbaceous Litter with Tree Litters on Decomposition and N Release in Boreal Plantations. Plant Soil 2016, 398, 229–241. [Google Scholar] [CrossRef]
- Becker, H.; Aosaar, J.; Varik, M.; Morozov, G.; Aun, K.; Mander, Ü.; Soosaar, K.; Uri, V. Annual Net Nitrogen Mineralization and Litter Flux in Well-Drained Downy Birch, Norway Spruce and Scots Pine Forest Ecosystems. Silva Fenn. 2018, 52, 10013. [Google Scholar] [CrossRef]
- Hansson, K.; Olsson, B.A.; Olsson, M.; Johansson, U.; Kleja, D.B. Differences in Soil Properties in Adjacent Stands of Scots Pine, Norway Spruce and Silver Birch in SW Sweden. For. Ecol. Manag. 2011, 262, 522–530. [Google Scholar] [CrossRef]
- Hojjati, S.M.; Hagen-Thorn, A.; Lamersdorf, N.P. Canopy Composition as a Measure to Identify Patterns of Nutrient Input in a Mixed European Beech and Norway Spruce Forest in Central Europe. Eur. J. For. Res. 2009, 128, 13–25. [Google Scholar] [CrossRef]
- Thelin, G.; Rosengren, U.; Callesen, I.; Ingerslev, M. The Nutrient Status of Norway Spruce in Pure and in Mixed-Species Stands. For. Ecol. Manag. 2002, 160, 115–125. [Google Scholar] [CrossRef]
- Daněk, P.; Šamonil, P.; Hort, L. Forest Floor Alteration by Canopy Trees and Soil Wetness Drive Regeneration of a Spruce-Beech Forest. For. Ecol. Manag. 2022, 504, 119802. [Google Scholar] [CrossRef]
- Likulunga, L.E.; Pérez, C.A.R.; Schneider, D.; Daniel, R.; Polle, A. Forest Tree Species Composition and Abiotic Site Conditions Drive Soil Fungal Communities and Functional Groups. bioRxiv, 2021. Available online: https://www.biorxiv.org/content/10.1101/2021.07.21.453256v1.full.pdf (accessed on 20 April 2022).
- Zeller, B.; Legout, A.; Bienaimé, S.; Gratia, B.; Santenoise, P.; Bonnaud, P.; Ranger, J. Douglas Fir Stimulates Nitrification in French Forest Soils. Sci. Rep. 2019, 9, 10687. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Climate Gradients. Substrate Quality versus Climate and Their Interactions. In Plant Litter; Springer: Berlin/Heidelberg, Germany, 2020; pp. 165–187. [Google Scholar]
- Zhou, S.; Butenschoen, O.; Barantal, S.; Handa, I.T.; Makkonen, M.; Vos, V.; Aerts, R.; Berg, M.P.; McKie, B.; Van Ruijven, J. Decomposition of Leaf Litter Mixtures across Biomes: The Role of Litter Identity, Diversity and Soil Fauna. J. Ecol. 2020, 108, 2283–2297. [Google Scholar] [CrossRef]
- Rode, M.W. The Interaction between Organic Layer and Forest Growth and Forest Development on Former Heathland. For. Ecol. Manag. 1999, 114, 117–127. [Google Scholar] [CrossRef]
- Nilsson, M.-C.; Wardle, D.A.; Dahlberg, A. Effects of Plant Litter Species Composition and Diversity on the Boreal Forest Plant-Soil System. Oikos 1999, 86, 16–26. [Google Scholar] [CrossRef]
- Simard, M.-J.; Bergeron, Y.; Sirois, L. Conifer Seedling Recruitment in a Southeastern Canadian Boreal Forest: The Importance of Substrate. J. Veg. Sci. 1998, 9, 575–582. [Google Scholar] [CrossRef]
- Wardle, D.A.; Zackrisson, O.; Nilsson, M.-C. The Charcoal Effect in Boreal Forests: Mechanisms and Ecological Consequences. Oecologia 1998, 115, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Wahl, N.A.; Bens, O.; Schäfer, B.; Hüttl, R.F. Impact of Changes in Land-Use Management on Soil Hydraulic Properties: Hydraulic Conductivity, Water Repellency and Water Retention. Phys. Chem. Earth Parts ABC 2003, 28, 1377–1387. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Hokkanen, T.J.; Järvinen, E.; Pukkala, T. Factors Related to Seedling Growth in a Boreal Scots Pine Stand: A Spatial Analysis of a Vegetation—Soil System. Can. J. For. Res. 1993, 23, 2101–2109. [Google Scholar] [CrossRef]
- Oberhuber, W.; Hammerle, A.; Kofler, W. Tree Water Status and Growth of Saplings and Mature Norway Spruce (Picea Abies) at a Dry Distribution Limit. Front. Plant Sci. 2015, 6, 703. [Google Scholar] [CrossRef]
- Sousa, T.R.; Schietti, J.; Ribeiro, I.O.; Emílio, T.; Fernández, R.H.; ter Steege, H.; Castilho, C.V.; Esquivel-Muelbert, A.; Baker, T.; Pontes-Lopes, A. Water Table Depth Modulates Productivity and Biomass across Amazonian Forests. Glob. Ecol. Biogeogr. 2022, 31, 1571–1588. [Google Scholar] [CrossRef]
- Berg, B. Litter Decomposition and Organic Matter Turnover in Northern Forest Soils. For. Ecol. Manag. 2000, 133, 13–22. [Google Scholar] [CrossRef]
- Henneb, M.; Thiffault, N.; Valeria, O. Regional Climate, Edaphic Conditions and Establishment Substrates Interact to Influence Initial Growth of Black Spruce and Jack Pine Planted in the Boreal Forest. Forests 2020, 11, 139. [Google Scholar] [CrossRef]
- Mason, W.L.; Edwards, C.; Hale, S.E. Survival and Early Seedling Growth of Conifers with Different Shade Tolerance in a Sitka Spruce Spacing Trial and Relationship to Understorey Light Climate. Silva Fenn. 2004, 38, 357–370. [Google Scholar] [CrossRef]
- Boucher, D.; Gauthier, S.; Thiffault, N.; Marchand, W.; Girardin, M.; Urli, M. How Climate Change Might Affect Tree Regeneration Following Fire at Northern Latitudes: A Review. New For. 2020, 51, 543–571. [Google Scholar] [CrossRef]
- Liepe, K.J.; Hamann, A.; Smets, P.; Fitzpatrick, C.R.; Aitken, S.N. Adaptation of Lodgepole Pine and Interior Spruce to Climate: Implications for Reforestation in a Warming World. Evol. Appl. 2016, 9, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.A.; Gonçalves, D.S.; Souza, P.A.; de Lucena, F.R.; Silva, R.R. da Brondani, G.E. Luminosity Levels Affect the Initial Seedlings Growth and Nutrient Accumulation in Khaya Senegalensis A. Juss. Cerne 2018, 24, 344–351. [Google Scholar] [CrossRef]
- De Vries, W.; Van der Salm, C.; Reinds, G.J.; Erisman, J.W. Element Fluxes through European Forest Ecosystems and Their Relationships with Stand and Site Characteristics. Environ. Pollut. 2007, 148, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Houle, D.; Marty, C.; Duchesne, L.; Gagnon, C. Humus Layer Is the Main Locus of Secondary SO4 Production in Boreal Forests. Geochim. Cosmochim. Acta 2014, 126, 18–29. [Google Scholar] [CrossRef]
- Lindroos, A.-J.; Derome, J.; Derome, K.; Smolander, A. The Effect of Scots Pine, Norway Spruce and Silver Birch on the Chemical Composition of Stand Throughfall and Upper Soil Percolation Water in Northern Finland. 2011. Available online: https://jukuri.luke.fi/bitstream/handle/10024/516371/Lindroos.pdf?sequence=1&isAllowed=y (accessed on 12 April 2022).
- Driscoll, C.T.; Lawrence, G.B.; Bulger, A.J.; Butler, T.J.; Cronan, C.S.; Eagar, C.; Lambert, K.F.; Likens, G.E.; Stoddard, J.L.; Weathers, K.C. Acidic Deposition in the Northeastern United States: Sources and Inputs, Ecosystem Effects, and Management Strategies: The Effects of Acidic Deposition in the Northeastern United States Include the Acidification of Soil and Water, Which Stresses Terrestrial and Aquatic Biota. BioScience 2001, 51, 180–198. [Google Scholar]
- Jones, D.L.; Kielland, K. Soil Amino Acid Turnover Dominates the Nitrogen Flux in Permafrost-Dominated Taiga Forest Soils. Soil Biol. Biochem. 2002, 34, 209–219. [Google Scholar] [CrossRef]
- Adamczyk, B.; Ahvenainen, A.; Sietiö, O.-M.; Kanerva, S.; Kieloaho, A.-J.; Smolander, A.; Kitunen, V.; Saranpää, P.; Laakso, T.; Straková, P. The Contribution of Ericoid Plants to Soil Nitrogen Chemistry and Organic Matter Decomposition in Boreal Forest Soil. Soil Biol. Biochem. 2016, 103, 394–404. [Google Scholar] [CrossRef]
- Killham, K. Nitrification in Coniferous Forest Soils. Plant Soil 1990, 128, 31–44. [Google Scholar] [CrossRef]
- Kendrick, W.B. The Time Factor in the Decomposition of Coniferous Leaf Litter. Can. J. Bot. 1959, 37, 907–912. [Google Scholar] [CrossRef]
- Urli, M.; Thiffault, N.; Houle, D.; Gauthier, S.; Bergeron, Y. Role of Green Alder in Boreal Conifer Growth: Competitor or Facilitator? Facets 2020, 5, 166–181. [Google Scholar] [CrossRef]
- Nikonov, V.V.; Lukina, N.V.; Polyanskaya, L.M.; Panikova, A.N. Distribution of Microorganisms in the Al–Fe–Humus Podzols of Natural and Anthropogenically Impacted Boreal Spruce Forests. Microbiology 2001, 70, 319–328. [Google Scholar] [CrossRef]
- Giesler, R.; Satoh, F.; Ilstedt, U.; Nordgren, A. Microbially Available Phosphorus in Boreal Forests: Effects of Aluminum and Iron Accumulation in the Humus Layer. Ecosystems 2004, 7, 208–217. [Google Scholar] [CrossRef]
- Caccia, F.D.; Ballaré, C.L. Effects of Tree Cover, Understory Vegetation, and Litter on Regeneration of Douglas-Fir (Pseudotsuga Menziesii) in Southwestern Argentina. Can. J. For. Res. 1998, 28, 683–692. [Google Scholar] [CrossRef]
- Simard, M.-J.; Bergeron, Y.; Sirois, L. Substrate and Litterfall Effects on Conifer Seedling Survivorship in Southern Boreal Stands of Canada. Can. J. For. Res. 2003, 33, 672–681. [Google Scholar] [CrossRef]
- Simola, H. Persistent Carbon Loss from the Humus Layer of Tilled Boreal Forest Soil. Eur. J. Soil Sci. 2018, 69, 303–314. [Google Scholar] [CrossRef]
- Munson, A.D.; Timmer, V.R. Soil Nitrogen Dynamics and Nutrition of Pine Following Silvicultural Treatments in Boreal and Great Lakes-St. Lawrence Plantations. For. Ecol. Manag. 1995, 76, 169–179. [Google Scholar] [CrossRef]
- Prescott, C.E.; Maynard, D.G.; Laiho, R. Humus in Northern Forests: Friend or Foe? For. Ecol. Manag. 2000, 133, 23–36. [Google Scholar] [CrossRef]
- Natalia, S.; Lieffers, V.J.; Landhäusser, S.M. Effects of Leaf Litter on the Growth of Boreal Feather Mosses: Implication for Forest Floor Development. J. Veg. Sci. 2008, 19, 253–260. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Litter From Species | Added Amount of Litter by Cavity (g) | Field Equivalent (100%) | ||
|---|---|---|---|---|
| 25% | 50% | 100% | (t ha−1) | |
| PET | 2.4 | 4.8 | 9.6 | 24.9 |
| BOP | 4.69 | 9.38 | 18.76 | 48.8 |
| AUR | 2.3 | 4.6 | 9.2 | 23.9 |
| BS | 1.25 | 2.5 | 5.0 | 13.0 |
| SAL | 1.28 | 2.56 | 5.12 | 13.5 |
| Nutrient | Unit | Litter | Humus | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| PET | BOP | SAL | AUR | BS | T. aspen | Mixed | B. spruce | Nursery | ||
| C tot | g kg−1 | 512 | 519 | 494 | 427 | 508 | 392 | 330 | 339 | 409 |
| N tot | g kg−1 | 8.4 | 9.2 | 14 | 16 | 15 | 15 | 9 | 8 | 6 |
| C:N | 61 | 56 | 35 | 26 | 34 | 26 | 34 | 41 | 66 | |
| Concentration | Total | Available (MehlichIII) | ||||||||
| P | mg kg−1 | 800 | 850 | 740 | 1390 | 860 | 171 | 52 | 38 | 125 |
| K | mg kg−1 | 2000 | 1700 | 9100 | 6900 | 700 | 701 | 304 | 399 | 1510 |
| Ca | mg kg−1 | 20,100 | 14,200 | 20,800 | 22,000 | 8390 | 7540 | 7970 | 3530 | 2180 |
| Mg | mg kg−1 | 1500 | 2230 | 2520 | 3320 | 1130 | 1130 | 1610 | 881 | 1320 |
| Mn | mg kg−1 | 110 | 740 | 100 | 210 | 120 | 264 | 68 | 40 | 30 |
| Cu | mg kg−1 | <20 | <20 | <20 | 40 | 70 | 2 | 3 | <1 | 10 |
| Zn | mg kg−1 | 220 | 270 | 180 | 160 | 60 | 67 | 12 | 20 | 55 |
| Al | mg kg−1 | 100 | 100 | 200 | 3800 | 1200 | 1410 | 1980 | 2500 | 484 |
| Fe | mg kg−1 | 100 | 100 | 500 | 2300 | 1100 | 636 | 979 | 1090 | 628 |
| Na | mg kg−1 | 20 | 54 | 38 | 185 | |||||
| S | mg kg−1 | 125 | 64 | 24 | 71 | |||||
| CEC | cmol(+) kg−1 | 152 | 128 | 131 | 155 | |||||
| BS | % | 32 | 42 | 19 | 17 | |||||
| Litter | n | % | T. aspen | Mixed | B. spruce | Nursery |
|---|---|---|---|---|---|---|
| Litter | pH (2 cm) | pH (2 cm) | pH (2 cm) | pH (2 cm) | ||
| PET | 10 | 25 | 0.19 ± 0.02 | 0.36 ± 0.03 | 0.48 ± 0.02 | 0.36 ± 0.02 |
| 10 | 50 | 0.14 ± 0.02 | 0.18 ± 0.02 | 0.49 ± 0.03 | 0.31 ± 0.03 | |
| 10 | 100 | 0.08 ± 0.02 | 0.09 ± 0.03 | 0.50 ± 0.02 | 0.19 ± 0.03 | |
| AUR | 10 | 25 | 0.04 ± 0.02 | 0.10 ± 0.03 | 0.17 ± 0.02 | 0.32 ± 0.03 |
| 10 | 50 | 0.15 ± 0.02 | 0.08 ± 0.02 | 0.16 ± 0.2 | 0.24 ± 0.02 | |
| 10 | 100 | 0.39 ± 0.03 | 0.08 ± 0.02 | −0.41 ± 0.03 | −0.16 ± 0.03 | |
| SAL | 10 | 25 | 0.27 ± 0.02 | 0.16 ± 0.01 | 0.17 ± 0.03 | 0.28 ± 0.02 |
| 10 | 50 | 0.11 ± 0.02 | 0.18 ± 0.01 | 0.19 ± 0.01 | 0.01 ± 0.03 | |
| 10 | 100 | 0.09 ± 0.01 | 0.10 ± 0.01 | 0.26 ± 0.02 | 0.30 ± 0.04 | |
| BOP | 10 | 25 | −0.20 ± 0.03 | −0.08 ± 0.02 | 0.22 ± 0.01 | 0.15 ± 0.04 |
| 10 | 50 | −0.09 ± 0.04 | 0.13 ± 0.01 | 0.11 ± 0.02 | −0.54 ± 0.03 | |
| 10 | 100 | −0.13 ± 0.03 | 0.07 ± 0.01 | 0.07 ± 0.02 | −0.60 ± 0.02 | |
| BS | 10 | 25 | 0.35 ± 0.03 | 0.20 ± 0.02 | −0.05 ± 0.02 | −0.20 ± 0.01 |
| 10 | 50 | 0.02 ± 0.01 | −0.15 ± 0.01 | −0.22 ± 0.02 | −0.48 ± 0.01 | |
| 10 | 100 | −0.05 ± 0.02 | −0.22 ± 0.01 | −0.35 ± 0.01 | −0.51 ± 0.01 | |
| Control | 30 | 0 | −0.24 ± 0.03 | −0.55 ± 0.03 | −0.23 ± 0.02 | −0.56 ± 0.01 |
| Treatments | T. aspen | Mixed | B. spruce | Nursery | Total |
|---|---|---|---|---|---|
| PET | 5.8 ± 0.1 | 5.6 ± 0.1 | 5.3 ± 0.1 | 4.1 ± 0.2 | 5.2 ± 0.6 |
| AUR | 5.5 ± 0.0 | 50.5 ± 0.1 | 5.2 ± 0.2 | 4.0 ± 0.1 | 5.1 ± 0.5 |
| SAL | 5.5 ± 0.1 | 5.5 ± 0.1 | 4.5 ± 0.2 | 4.1 ± 0.0 | 4.9 ± 0.5 |
| BOP | 5.5 ± 0.0 | 5.5 ± 0.0 | 4.8 ± 0.2 | 4.1 ± 0.2 | 5.0 ± 0.6 |
| BS | 5.4 ± 0.0 | 5.2 ± 0.0 | 4.5 ± 0.1 | 3.9 ± 0.1 | 4.7 ± 0.6 |
| Control | 5.4 ± 0.0 | 5.2 ± 0.1 | 4.5 ± 0.1 | 3.9 ± 0.0 | 4.8 ± 0.6 |
| Average | 5.6 ± 0.1 | 5.4 ± 0.2 | 4.8 ± 0.4 | 4.0 ± 0.1 | 5.0 ± 0.6 |
| Litter Added | N | P | K | Ca | Mg | Mn | Cu | Zn |
|---|---|---|---|---|---|---|---|---|
| g kg−1 | ||||||||
| T. aspen stand humus substrate | ||||||||
| Average | 21.16 ± 3.65 | 2.24 ± 0.42 | 7.70 ± 0.85 | 11.89 ± 1.57 | 2.46 ± 0.21 | 1.40 ± 0.15 | 0.05 ± 0.01 | 0.16 ± 0.01 |
| PET | 23.27 ± 1.36 | 2.15 ± 0.33 | 7.97 ± 0.45 | 13.83 ± 0.86 | 2.36 ± 0.15 | 1.14 ± 0.05 | 0.05 ± 0.05 | 0.21 ± 0.04 |
| AUR | 23.63 ± 1.92 | 2.35 ± 0.12 | 8.40 ± 0.39 | 11.43 ± 0.78 | 2.58 ± 0.28 | 1.46 ± 0.01 | 0.05 ± 0.01 | 0.15 ± 0.04 |
| SAL | 17.43 ± 1.78 | 1.75 ± 0.09 | 6.33 ± 0.17 | 10.32 ± 0.25 | 2.55 ± 0.10 | 1.12 ± 0.03 | 0.06 ± 0.03 | 0.08 ± 0.01 |
| BOP | 16.70 ± 2.27 | 2.08 ± 0.31 | 7.67 ± 0.36 | 10.67 ± 0.65 | 2.36 ± 0.18 | 1.63 ± 0.72 | 0.05 ± 0.04 | 0.09 ± 0.02 |
| BS | 23.73 ± 2.09 | 2.64 ± 0.14 | 7.77 ± 0.74 | 12.33 ± 0.87 | 2.39 ± 0.15 | 1.62 ± 0.04 | 0.05 ± 0.01 | 0.24 ± 0.02 |
| Control | 24.30 ± 0.00 | 2.90 ± 0.00 | 8.80 ± 0.00 | 14.40 ± 0.00 | 2.68 ± 0.00 | 1.54 ± 0.00 | 0.05 ± 0.00 | 0.270 ± 0.00 |
| Mixed stand humus substrate | ||||||||
| Average | 17.93 ± 3.04 | 2.19 ± 0.24 | 8.07 ± 0.72 | 10.46 ± 0.90 | 2.20 ± 0.16 | 2.75 ± 0.44 | 0.04 ± 0.01 | 0.13 ± 0.01 |
| PET | 18.80 ± 1.40 | 2.41 ± 0.06 | 7.83 ± 0.28 | 10.90 ± 0.53 | 2.21 ± 0.12 | 2.14 ± 0.21 | 0.04 ± 0.01 | 0.15 ± 0.02 |
| AUR | 20.97 ± 1.92 | 2.44 ± 0.06 | 7.70 ± 0.51 | 11.00 ± 0.66 | 2.38 ± 0.16 | 2.78 ± 0.09 | 0.05 ± 0.01 | 0.14 ± 0.01 |
| SAL | 16.23 ± 2.03 | 2.11 ± 0.13 | 8.73 ± 0.58 | 9.61 ± 0.43 | 2.05 ± 0.07 | 2.81 ± 0.22 | 0.03 ± 0.06 | 0.11 ± 0.01 |
| BOP | 13.70 ± 0.11 | 2.20 ± 0.16 | 8.70 ± 0.64 | 9.90 ± 0.53 | 2.08 ± 0.09 | 2.71 ± 0.22 | 0.04 ± 0.00 | 0.12 ± 0.03 |
| BS | 19.83 ± 2.20 | 1.86 ± 0.07 | 7.30 ± 0.16 | 10.69 ± 1.21 | 2.23 ± 0.11 | 3.11 ± 0.48 | 0.04 ± 0.01 | 0.11 ± 0.01 |
| Control | 18.20 ± 0.00 | 2.02 ± 0.00 | 8.40 ± 0.00 | 11.10 ± 0.00 | 2.34 ± 0.00 | 3.34 ± 0.00 | 0.05 ± 0.00 | 0.13 ± 0.00 |
| B. spruce stand humus substrate | ||||||||
| Average | 14.93 ± 1.81 | 1.67 ± 0.21 | 7.91 ± 0.67 | 9.44 ± 0.96 | 2.16 ± 0.22 | 1.51 ± 0.78 | 0.06 ± 0.01 | 0.12 ± 0.01 |
| PET | 13.90 ± 0.70 | 1.65 ± 0.12 | 8.73 ± 0.40 | 9.91 ± 0.44 | 2.42 ± 0.24 | 1.17 ± 0.64 | 0.06 ± 0.01 | 0.13 ± 0.01 |
| AUR | 17.03 ± 1.52 | 1.88 ± 0.09 | 7.73 ± 0.66 | 10.39 ± 0.63 | 2.23 ± 0.08 | 2.34 ± 0.12 | 0.06 ± 0.01 | 0.14 ± 0.03 |
| SAL | 13.37 ± 0.91 | 1.47 ± 0.13 | 7.40 ± 0.37 | 8.25 ± 0.58 | 2.06 ± 0.03 | 0.67 ± 0.12 | 0.05 ± 0.01 | 0.11 ± 0.02 |
| BOP | 14.93 ± 0.54 | 1.70 ± 0.16 | 7.80 ± 0.72 | 9.89 ± 0.85 | 2.11 ± 0.21 | 2.10 ± 0.54 | 0.05 ± 0.01 | 0.13 ± 0.01 |
| BS | 15.97 ± 2.02 | 1.77 ± 0.26 | 7.83 ± 0.42 | 8.86 ± 0.38 | 1.93 ± 0.02 | 1.62 ± 0.67 | 0.06 ± 0.01 | 0.11 ± 0.01 |
| Control | 13.30 ± 0.0 | 1.53 ± 0.00 | 8.10 ± 0.00 | 9.14 ± 0.00 | 2.37 ± 0.00 | 0.52 ± 0.00 | 0.05 ± 0.00 | 0.13 ± 0.00 |
| Nursery substrate | ||||||||
| Average | 10.49 ± 1.79 | 1.47 ± 0.19 | 8.44 ± 0.39 | 6.74 ± 0.57 | 2.09 ± 0.17 | 0.67 ± 0.04 | 0.05 ± 0.05 | 0.07 ± 0.01 |
| PET | 9.50 ± 0.58 | 1.36 ± 0.08 | 8.47 ± 0.16 | 6.32 ± 0.28 | 2.013 ± 0.13 | 0.55 ± 0.02 | 0.05 ± 0.02 | 0.06 ± 0.02 |
| AUR | 13.80 ± 1.74 | 1.40 ± 0.09 | 8.60 ± 0.35 | 7.30 ± 0.65 | 2.30 ± 0.12 | 0.79 ± 0.02 | 0.05 ± 0.04 | 0.08 ± 0.02 |
| SAL | 9.73 ± 0.40 | 1.43 ± 0.15 | 8.03 ± 0.26 | 6.55 ± 0.22 | 1.95 ± 0.16 | 0.46 ± 0.19 | 0.04 ± 0.05 | 0.07 ± 0.02 |
| BOP | 9.83 ± 0.29 | 1.67 ± 0.19 | 8.53 ± 0.13 | 6.89 ± 0.68 | 2.00 ± 0.21 | 0.99 ± 0.31 | 0.04 ± 0.01 | 0.08 ± 0.01 |
| BS | 9.732 ± 0.17 | 1.61 ± 0.16 | 8.83 ± 0.21 | 6.88 ± 0.87 | 2.19 ± 0.18 | 0.62 ± 0.38 | 0.05 ± 0.01 | 0.07 ± 0.01 |
| Control | 10.00 ± 0.00 | 1.17 ± 0.00 | 7.70 ± 0.00 | 6.15 ± 0.00 | 2.11 ± 0.00 | 0.43 ± 0.00 | 0.05 ± 0.00 | 0.05 ± 0.00 |
| Average Total | 16.13 ± 4.81 | 1.90 ± 0.39 | 8.03 ± 0.67 | 9.63 ± 2.18 | 2.23 ± 0.24 | 1.58 ± 0.81 | 0.05 ± 0.14 | 0.12 ± 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Noronha, M.; Ouimet, R.; Barrette, M.; Leduc, A.; Bergeron, Y. Influence of Leaf Litter and Humus Composition on the Development of Black Spruce Seedlings: A Greenhouse Experimentation. Forests 2022, 13, 1832. https://doi.org/10.3390/f13111832
De Noronha M, Ouimet R, Barrette M, Leduc A, Bergeron Y. Influence of Leaf Litter and Humus Composition on the Development of Black Spruce Seedlings: A Greenhouse Experimentation. Forests. 2022; 13(11):1832. https://doi.org/10.3390/f13111832
Chicago/Turabian StyleDe Noronha, Maísa, Rock Ouimet, Martin Barrette, Alain Leduc, and Yves Bergeron. 2022. "Influence of Leaf Litter and Humus Composition on the Development of Black Spruce Seedlings: A Greenhouse Experimentation" Forests 13, no. 11: 1832. https://doi.org/10.3390/f13111832
APA StyleDe Noronha, M., Ouimet, R., Barrette, M., Leduc, A., & Bergeron, Y. (2022). Influence of Leaf Litter and Humus Composition on the Development of Black Spruce Seedlings: A Greenhouse Experimentation. Forests, 13(11), 1832. https://doi.org/10.3390/f13111832