Abstract
The decomposition of abnormal litter caused by extreme weather events might play an increasingly important role in carbon and nutrient cycling in forest ecosystems under climate change scenarios, which needs to be fully investigated. In August 2020, the abnormal foliar litter of the goldenrain tree (Koelreuteria bipinnata var. Integrifoliola), the camphor tree (Cinnamomum camphora), and the weeping willow (Salix babylonica) after Typhoon Hagupit disturbance were collected and incubated on the soil surface at the Plant Ecology Research Base at Taizhou University, which is located on the eastern coast of China. Simultaneously, the physiological foliar litter of these three trees collected in the spring litter peak was incubated at the same site. The abnormal litter had higher concentrations of carbon (C), nitrogen (N), and phosphorus (P) and lower concentrations of lignin and cellulose than the physiological litter. The accumulative mass loss rates of abnormal litter in the goldenrain tree, the camphor tree, and the weeping willow during the incubation period increased by 7.72%, 29.78%, and 21.76% in comparison with physiological litter, and the corresponding carbon release increased by 9.10%, 24.15% and 19.55%, respectively. The autumn litter peak period and plum-rain season had higher rates of litter mass loss and carbon release, while the winter nongrowing season had lower rates. Accumulative mass loss, accumulative carbon release, daily mass loss and the daily carbon release of foliar litter were significantly and positively correlated with temperature and initial P concentrations, and significantly and negatively correlated with the initial C/P ratio, lignin/N ratio, and lignin/P ratio (p < 0.05). Compared with the physiological litter, abnormal litter had higher initial substrate quality, which may be the most important factor contributing to their high rates of mass loss and carbon release. The results imply that increasing tropical cyclones under climate change scenarios will facilitate carbon cycling in coastal urban forest ecosystems.
1. Introduction
Tropical cyclones (typhoons and hurricanes) rank first on the list of the top 10 most dangerous natural hazards around the world [1]; these storms not only cause the loss of lives and property, but also affect the structure and function of terrestrial ecosystems [2,3], particularly the global carbon cycle [4,5]. For instance, Zeng et al. [5] indicated that, on average, 97 million trees in the United States died each year because of hurricanes from 1851 to 2000, resulting in an annual loss of 53 Tg of biomass, equivalent to an annual release of 25 Tg carbon. In particular, hurricane-induced carbon sink loss offset 9%–18% of the total forest carbon sink in the United States during 1980–1990. Chambers et al. [6] demonstrated that the effect of Hurricane Katrina on forest damage and mortality was closely related to tree species, as Hurricane Katrina in 2005 resulted in significant forest structural damage and mortality of ~320 million large trees, equivalent to a loss of 105 Tg carbon and accounting for ca. 50%–140% of the net annual forest carbon sink in US forests. According to the IPCC [7] assessment, the frequency, intensity, and disturbance region of tropical cyclones are increasing, implying that tropical cyclones are likely to have a more profound effect on the global carbon cycle. Indeed, the effects of hurricanes and typhoons on the global carbon cycle result not only from direct uprooting, branch breaking, overturning, and wind inversion, but also from abnormal litter decomposition and carbon release. However, little information is available on this topic.
Plant litter is a carrier of carbon and nutrient cycling in forest ecosystems [8,9]. Worldwide, annual forest litter production is estimated to account for approximately 20% of net ecosystem productivity (NEP) [10] and up to 34% of NEP in tropical wet evergreen forests [11]. In particular, plant litter decomposition plays crucial roles not only in site fertility maintenance and soil formation but also in the global carbon cycle [12]. Raich and Schlesinger [13] estimated that litter decomposition (including root litter) contributed 70% of the total annual carbon flux, approximately 68 PgC/a (Pg = 1015 g). Increasing evidence has indicated that the proportion of abnormal litter production to annual forest litter production is increasing due to extreme weather events, such as tropical cyclones [14,15], and is closely related to forest type and tree species [16]. As affected by Hurricane Hugo in 1989, the forests in the Luquillo Experimental Forest of Puerto Rico produced 10,800 kg abnormal litter per hectare, which was equivalent to 1.25 times the annual litterfall production in the Bisley Experimental Watersheds [17]. As affected by Hurricanes Irma and Maria in 2017, the abnormal litter production of these forests reached 32,225 kg per hectare, equivalent to 1.61 times the annual litter production [18]. Therefore, an in-depth investigation of the decomposition and carbon release from plant litter caused by tropical cyclone disturbances can provide essential data for assessing the global carbon cycle.
Abnormal litter is nonphysiologically fallen plant tissues, such as blown green leaves and broken branches and twigs, caused by the mortality or destruction of trees due to tropical cyclones (typhoons/hurricanes) [19], rainstorms, snow storms, ice disasters [20], and other extremely destructive disturbance events [21]. Litter quality, climate, and decomposer community are considered the three key factors influencing litter decomposition [22,23]. Theoretically, abnormal litter decomposition is similarly driven by these factors. Different from physiological litter, however, abnormal tropical cyclone-induced forest litter decomposition might have distinct processes for the following reasons (Figure 1). First, abnormal litter generally has higher N and P concentrations and relatively lower C/N and C/P ratios, due to the lack of nutrient resorption processes and thus may have faster rates of mass loss and carbon release [15]. Second, the concentrations of lignin and cellulose in green leaves are relatively lower due to their relatively low fibrillation and lignification during the growing peak [12], and thus abnormal foliar litter from extreme meteorological events such as typhoons may have lower lignin and cellulose concentrations and therefore relatively lower ratios of lignin/N and lignin/P, resulting in faster decomposition rates. Third, decomposer communities (soil animals and microbes) are more active in warm and humid seasons, and accelerate litter decomposition [24,25], while abnormal litter formed by tropical cyclones is directly exposed in a warm and humid season and has higher mass loss and carbon release. As a result, the increasing forest abnormal litter may accelerate the global carbon cycle under global climate change scenarios. Present abnormal litter decomposition is increasingly a concern [15,20,26]. For instance, Yang et al. [20] have found that freeze-damaged needle litter has a faster decomposition rate than physiological needle litter. Li et al. [27] compared the decomposition processes of green and senescent leaves of nine plant species and found that green leaves had relatively higher mass loss rates than senescent leaves. Moreover, the physiological and morphological responses of trees to tropical cyclone disturbances should vary greatly with tree species, which leads to large differences in the production and quality of abnormal litter from different tree species and in turn affects the processes of abnormal mass loss and carbon release at the ecosystem level. However, the mass loss and carbon release of abnormal litter from different tree species produced by tropical cyclones remain unknown.
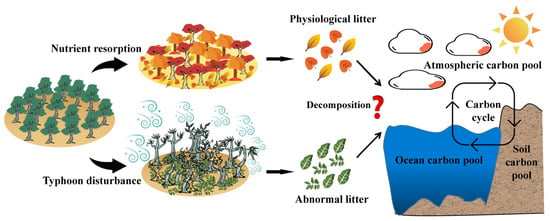
Figure 1.
The conceptual diagram depicts the lack of nutrient resorption processes in typhoon-induced abnormal litter, leading to changes in the litter’s initial substrate quality, with cascading effects on the ecosystem carbon cycle.
Typhoons are one of the most important disturbance factors affecting the structure and function of coastal forest ecosystems in the Northwest Pacific Ocean [28]. China is one of the countries most affected by typhoons [29]. Typhoons predominantly occur in the southeastern coastal region of China [30], such as in the provinces of Zhejiang, Fujian, and Guangdong. According to statistics, from 1984 to 2019, a total of 208 tropical cyclones made landfall in the southeastern coastal cities of China, of which 88 were intense tropical storms or stronger [31]. Frequent typhoons not only severely disturb coastal forests and urban garden tree species in China [32], but also have a profound impact on the homeostasis and carbon cycling processes of coastal forest ecosystems [33,34]. For instance, Huang et al. [32] investigated the effects of Typhoon Mangkhut in 2018 on urban garden tree species in Guangzhou, Hong Kong, and Macau Cities, and found that the responses of garden trees to typhoon disturbance varied greatly with tree species, and more than 60% of Radermachera hainanensis and Acacia confusa were damaged by this typhoon. Recently, we observed that Typhoon Hagupit in 2020 changed the concentrations of nitrogen, phosphorus, potassium, calcium, sodium, and magnesium in garden plant leaves. However, after a shorter period, the concentrations of these nutrients in plant leaves were restored to the level before the typhoon [35,36], implying that the initial nutrient status of abnormal litter differs from that of physiological litter, and leading to specificity in decomposition, carbon and nutrient release, which may regulate the material cycling processes in coastal forest ecosystems. However, the decomposition of abnormal litter induced by typhoons remain unknown. The mass loss and carbon release of typhoon-produced abnormal foliar litter of the goldenrain tree, the camphor tree, and the weeping willow tree, the three most prevalent garden green trees in subtropical coastal cities, were therefore investigated, and used the litterbag method in the Plant Ecology Research Base at Taizhou University, located in the southeast coastal area of Zhejiang Province, China. The main objectives of this study are to: (1) understand the effects of typhoon disturbance on litter decomposition in coastal urban forest ecosystem; (2) reveal the effects of typhoon disturbance on carbon cycling in coastal urban forest ecosystems via abnormal litter decomposition; and (3) explore the differential responses of abnormal litter decomposition from different tree species to typhoon disturbance, so as to provide scientific basis for post-typhoon recovery and reconstruction of coastal urban forests under climate change scenarios and to add important data to the global carbon cycle model.
2. Materials and Methods
2.1. Study Site
This study was conducted in the Plant Ecology Research Base at the Taizhou University, which is situated on the eastern coast of Zhejiang Province (28°63′–28°68′ N, 121°38′–121°41′ E). The area is located in the western Pacific Ocean and belongs to the mid-subtropical monsoon zone. Due to the barrier of cold currents by the high mountains in the northwest and the regulation of the high specific heat capacity of marine water, there is an abundance of sunlight and heat, with an annual average temperature of 14.8–23.8 °C, a maximum of 40 °C, and a minimum of −4 °C. The annual precipitation is 1632 mm, ranging from 1185 to 2029 mm, and rainy days are over 120 d. The plum-rain season, named the Meiyu season, occurs from early June to early July and is characterized by frequent rainfall and damp weather [30]. In particular, the ecosystems in this region are affected by frequent typhoon disturbances from July to October, and over 170 typhoons have affected all types of ecosystems in the past five decades [37,38]. In recent years, particularly in 2019, the super typhoon “Lekima” (http://news.cctv.com/special/tflqm/) (accessed on 30 September 2022) has had a dramatic impact on the security of urban property and urban forest ecosystems in Taizhou city. Taizhou city is forest urban, and urban forest covers ca. 62.2% [35], which has abundant urban garden plants. Among these, the goldenrain tree (Koelreuteria bipinnata var. Integrifoliola), the camphor tree (Cinnamomum camphora), and the weeping willow tree (Salix babylonica) are widely used to green the city. However, these garden trees are frequently attacked by typhoons [35]. Even so, there is a lack of deep investigation into the decomposition and carbon release of abnormal litter caused by typhoons.
2.2. Litter Collection and Experimental Design
To understand the role of abnormal litter in the carbon cycle, the physiological foliar litter of goldenrain trees, camphor trees and weeping willow trees, three representative urban garden tree species in Taizhou city, were collected in late spring in 2020, before the typhoon began or late autumn 2019, and were air-dried and stored in the lab. The green leaves (abnormal foliar litter) on the floor blown down by the typhoon were collected on the second day after Typhoon Hagubit (No. 4) swept through Taizhou city on 4 August 2020. The collected green leaves of the three tree species were transported to the laboratory and air-dried naturally on a flat table for two weeks. Then, 10.00 g of air-dried physiological and abnormal foliar litter were precisely weighed and put into nylon bags. The area of the nylon bag was 20 cm by 20 cm, and the mesh size was 0.04 mm on the bottom (soil contact) and 3 mm surface (atmospheric contact) aperture [39]. All the litterbags were randomly incubated on the soil surface at the Plant Ecology Research Base at Taizhou University on 9 September 2020. We considered the effects of typhoons and other factors to prevent the loss of samples. The bags were connected to each other using thick fishing lines, and the ends of the cords were fixed with iron rings, while the distance between the bags was ≥5 cm to avoid mutual influence. Finally, button thermometers (iButton DS1921G, Maxim/Dallas Semiconductor, Sunnyvale, CA, USA) were placed into the litterbags and topsoil layer to monitor the temperature dynamics of the litterbags and topsoil; meanwhile, air temperature dynamics were monitored by button thermometers hanging on the air (set to record every 2 h). The daily mean temperatures of litterbags, topsoil, and air and the actual moisture dynamics of foliar litter are presented in Supplementary Material (Figures S1 and S2).
Based on Taizhou’s historically hot and humid climate, one-year field incubation experiment periods were divided into five critical decomposition periods, i.e., the autumn litter peak period (10 September 2020–9 December 2020, ALP), winter nongrowing season (10 December 2020–9 March 2021, WNG), spring litter peak period (10 March 2021–16 May 2021, SLP), early plum-rainy season (17 May 2021–16 June 2021, EPR), and late plum-rainy season (17 June 2021–21 July 2021, LPR). A total of 90 litterbags (2 litter sources, physiological and abnormal litter × 3 species × 5 sample times × 3 replicates) were randomly collected on 9 December 2020 and on 9 March, 16 May, 16 June, and 21 July 2021.
2.3. Sampling and Chemical Analyses
Litterbags were retrieved at five critical incubation periods, i.e., ALP, WNG, SLP, EPR, and LPR, and were immediately (within 2 h) transported to the laboratory, manually separated from foreign materials oven-dried at 65 °C, and weighed to determine the dry mass loss. Additionally, three reserved litterbags of foliar litter from each tree species were also oven-dried at 65 °C and weighed to determine the moisture content of the foliar litter used for the conversion of the dry materials. The reserved, naturally air-dried foliar litter from different sources of the three tree species was also used to determine the concentrations of the initial chemical components. The oven-dried litter samples were ground in a laboratory mill to a mesh fraction of less than 0.25 mm.
The concentrations of carbon, nitrogen, phosphorus, lignin, and cellulose in the litter were then analyzed. The concentrations of carbon and nitrogen in the litter samples were determined using an element analyzer (Elementar Analysensysteme GmbH, Hanau, Germany) [40]. The instrument’s fundamental principle is the Dumas combustion equilibrium theory [41]. A prepared sample of 0.1 g litter was weighed, wrapped in a tin capsule, and fed via the autosampler into the combustion reaction tube, and a small quantity of pure oxygen was introduced into the system to help in the complete burning of the sample. The combusted sample was then subjected to a further catalytic redox process in which carbon and nitrogen were transformed into various detectable gases, including COx and NOx. The gas mixture is further separated by a separation column and finally passed through the thermal conductivity detector (TCD) to complete the detection, and the results are output by the connected computer. The total concentration of phosphorus was determined using the molybdenum-blue colorimetric method [42]. The concentrations of lignin and cellulose in litter were measured using an acid-detergent fiber-sulfuric lignin method [43] with some modification. The fractions determined by this method are now mostly known as Klason lignin or acid unhydrolysable residue (AUR). The method and procedure are given in He et al. [44]. All analyses were conducted in triplicate.
2.4. Statistical Analysis and Calculation
We calculated the rates of accumulative mass loss (Lt) and accumulative carbon release (R) from the different sources of foliar litter for three species at different critical periods, as well as the daily mass loss (Vm) and carbon release (Vc) from foliar litter at each critical period, and simulated the decomposition rate (k value) of foliar litter by the Olsen exponential equation.
Accumulative mass loss rate of foliar litter (Equation (1) [45]):
Lt(%) = (M0 − Mt)/M0 × 100
M0 = Initial dry weight of foliar litter (initial mass litter),
Mt = Litter dry weight in the corresponding moment (remaining mass litter).
Accumulative C release of foliar litter (Equation (2) [46]):
R(%) = (M0C0 − MtCt)/M0C0 × 100
C0 = Initial C concentration of foliar litter,
Ct = C concentration of foliar litter in the corresponding moment.
Daily mass loss and carbon release of foliar litter (Equations (3) and (4) [47]):
Vm(mg/d) = (Mt−1 − Mt)/ΔT
Vc(mg/d) = (Mct−1 − Mct)/ΔT
Mct = Litter carbon weight in the corresponding moment,
ΔT = The length (day number) of each critical period.
Olsen’s decomposition of the exponential equation (Equation (5) [48]):
Xt = X0e−kt
X0 = Initial dry weight of foliar litter (initial mass litter),
Xt = Litter dry weight in the corresponding moment (remaining mass litter),
t = Decomposition time (days),
k = Decomposition coefficient.
The differences in the initial element concentration of foliar litter from the three species and the daily mass loss and daily carbon release at different critical periods of foliar litter from the three species were tested using Tukey’s HSD test when the one-way ANOVA results were significant at p = 0.05. The differences in the initial element concentrations of different sources of foliar litter, and the accumulative mass loss rate, carbon concentration, accumulative carbon release rate, daily mass loss, and daily carbon release at different critical periods of different sources of foliar litter were evaluated using an independent sample t-test with an alpha level of 0.05. The effects of litter sources, tree species, critical periods, and their interactions on the mass loss rate, carbon concentration, and accumulative carbon release rate of the foliar litter were tested using a repeated-measures ANOVA with litter sources, species, and critical periods as independent categorical variables. Pearson’s correlations were performed among the investigated parameters. All statistical analyses used the SPSS 25.0 (SPSS Inc., Chicago, IL, USA), graphing with Origin 2021 (OriginLab, Northampton, MA, USA).
3. Results
3.1. Initial Litter Quality of Abnormal Foliar Litter
Abnormal litter had higher substrate quality than physiological litter (Table 1, p < 0.05). Compared to the physiological litter, the concentrations of C, N, and P in the abnormal litter were increased by 3.07%–6.73%, 17.79%–92.5%, and 28.22%–84.52%, respectively, while the concentrations of lignin and cellulose and the ratios of C/N, C/P, N/P, lignin/N, and lignin/P, were decreased by 34.82%–44.14% and 28.90%–65.78%, and 12.43%–44.58%, 18.35%–42.87%, 25.51%–27.83%, 7.97%–70.98%, and 32.75%–69.90%, respectively (Table 1). Additionally, the differences between physiological and abnormal litter varied greatly with tree species.

Table 1.
Mean initial elemental concentrations and substrate quality of foliar litter from three species of subtropical urban forest at the start of the decomposition experiment. Standard errors are in parentheses. Different capital letters indicate significant differences between different tree species (p < 0.05). Different lowercase letters indicate significant differences between different sources of foliar litter (p < 0.05).
3.2. Litter Mass Loss
The accumulative mass loss rates of foliar litter displayed characteristic decomposition patterns, which appeared to approximate an exponential function (Figure 2). The decomposition process of foliar litter was simulated by the Olsen index equation, and perfect fitting results were obtained (Table 2, R2 > 0.85, p < 0.05). Abnormal litter had higher rates of accumulative mass loss and decomposition than physiological litter (Figure 2, Table 2). After one-year decomposition, compared with the physiological litter, the rates of accumulative mass loss and decomposition of abnormal litter increased by 7.72%–29.78% and 8.00%–42.86%, respectively (Figure 2), and depended on the tree species and critical periods. The accumulative mass loss rate and decomposition rate of abnormal litter for the three species, in descending order, were weeping willow (67.98%, k = 0.0030) > goldenrain tree (63.94%, k = 0.0027) > camphor tree (49.16%, k = 0.0020). Compared with the physiological litter, the rates of mass loss and decomposition of abnormal litter increased in the following order: camphor tree > weeping willow > goldenrain tree (Figure 2, Table 2). Regardless of the tree species, the mass loss of litter was higher during the ALP, EPR, and LPR reaching 18.80–43.88 mg/d, 13.87–33.44 mg/d, and 8.28–34.00 mg/d daily mass losses, respectively, while it was lowest during the WNG, with only 4.56–12.37 mg/d daily mass losses (Figure 3). Repeated-measures ANOVA revealed that the accumulative mass loss rate was significantly influenced by the source of foliar litter, tree species, critical period, and their interactions (Table 3, p < 0.001).
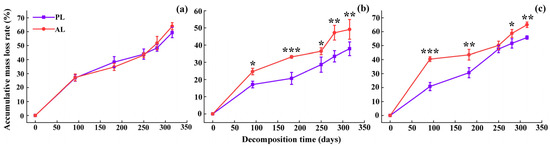
Figure 2.
The accumulative mass loss rates (%) of goldenrain tree (a), camphor tree (b), and weeping willow tree (c) foliar litter in the urban forest. PL and AL indicate physiological and abnormal litter, respectively. * p < 0.05, ** p < 0.01, *** p < 0.001.
Table 2.
The decomposition constant, correlation coefficient, and time of 50% and 95% decomposition of foliar litter from different sources for the three species in the urban forest.
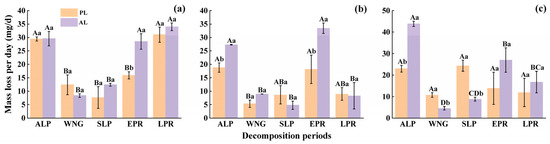
Figure 3.
The daily mass loss of goldenrain tree (a), camphor tree (b), and weeping willow tree (c) foliar litter during different critical periods in the urban forest. ALP, WNG, SLP, EPR, and LPR indicate the autumn litter peak period, winter non-growing season, spring litter peak period, early plum-rainy season, and late plum-rainy season, respectively. PL and AL indicate physiological and abnormal litter, respectively. Different capital letters indicate significant differences between different critical periods (p < 0.05). Different lowercase letters indicate significant differences between different sources of foliar litter (p < 0.05).
Table 3.
Results of repeated-measures ANOVA showing the p values for responses of accumulative mass loss rate, carbon concentration, and accumulative release rate to critical periods, species, and litter source.
3.3. Litter Carbon Release
The carbon concentrations of foliar litter fluctuated and decreased with time (Figure 4). After one-year decomposition, compared with the physiological litter, the decreased amplitude in abnormal litter carbon concentrations was elevated by 19.79%–30.11% (Figure 4) and depended on the tree species and critical periods. The carbon concentrations decreased the amplitude of abnormal litter for the three species in descending order: the weeping willow (28.83%) > the camphor tree (24.53%) > the goldenrain tree (21.02%). Compared with the physiological litter, the carbon concentrations of the abnormal litter decreased in amplitude in descending order: weeping willow trees > camphor trees > goldenrain trees (Figure 4). Regardless of the tree treatment, the litter carbon concentrations decreased mainly during the ALP and after the temperature recovery (SLP, EPR, and LPR), whereas they rose throughout the WNG (Figure 4).
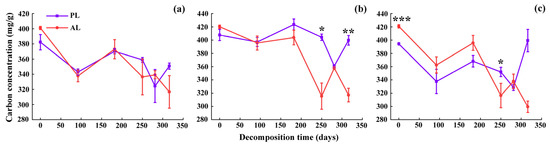
Figure 4.
The carbon concentration dynamics of goldenrain tree (a), camphor tree (b), and weeping willow tree (c) foliar litter in the urban forest. PL and AL indicate physiological and abnormal litter, respectively. * p < 0.05, ** p < 0.01, *** p < 0.001.
The accumulative carbon release rate of foliar litter assessed in the experiment increased with time, and abnormal litter had higher rates of accumulative carbon release rate than physiological litter (Figure 5). After one-year decomposition, the accumulative carbon release rate of the abnormal litter increased by 9.1%–24.15% compared with the physiological litter, depending on the tree species and critical periods (Figure 5). The accumulative carbon release rate of abnormal litter for the three species, in descending order, was the weeping willow (74.87%) > the goldenrain tree (71.64%) > the camphor tree (62.13%). Compared with the physiological litter, the accumulative carbon release rate of abnormal litter increased in the following order: the camphor tree > the weeping willow > the goldenrain tree (Figure 5). Regardless of the tree species treatment, the carbon release of litter was higher during the ALP and EPR, and the daily carbon release reached 8.61–22.25 mg/d and 3.22–16.56 mg/d, respectively, while it was lowest during the WNG, with only −0.40–2.94 mg/d daily carbon release (Figure 6). Repeated-measures ANOVA revealed that the carbon concentrations and accumulative carbon release rate were significantly influenced by the sources of foliar litter, the species, critical sampling period, and their interactions (Table 3, p < 0.001).
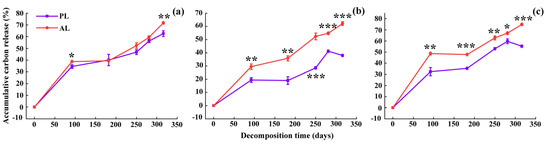
Figure 5.
The accumulative carbon release rates (%) of goldenrain tree (a), camphor tree (b), and weeping willow tree (c) foliar litter in the urban forest. PL and AL indicate physiological and abnormal litter, respectively. * p < 0.05, ** p < 0.01, *** p < 0.001.
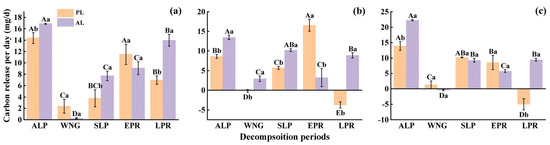
Figure 6.
The daily carbon release of goldenrain tree (a), camphor tree (b), and weeping willow tree (c) foliar litter during different critical periods. ALP, WNG, SLP, EPR, and LPR indicate the autumn litter peak period, winter non-growing season, spring litter peak period, early plum-rainy season, and late plum-rainy season, respectively. PL and AL indicate physiological and abnormal litter, respectively. Different capital letters indicate significant differences between different critical periods (p < 0.05). Different small letters indicate significant differences between different sources of foliar litter (p < 0.05).
3.4. Key Drivers of Mass Loss and Carbon Release of Abnormal Litter
Pearson’s correlation analysis results indicated that accumulative mass loss and accumulative carbon release were significantly positively correlated (p < 0.05) with the initial N and P concentrations and negatively correlated (p < 0.05) with the initial lignin concentrations, C/P, N/P, lignin/N, and lignin/P, while the opposite was true for the carbon concentration of litter (Figure 7). During different periods, daily mass loss and daily carbon release from foliar litter were strongly and positively correlated with initial P concentrations (p < 0.05) and considerably negatively correlated with initial C/P, lignin/N, and lignin/P (p < 0.05). Additionally, there were significant positive correlations (p < 0.05) between accumulative mass loss, accumulative carbon release, daily mass loss, and daily carbon release and atmospheric temperature, topsoil temperature, and foliar litter temperature, as well as a significant positive correlation (p < 0.05) between accumulative carbon release and foliar litter moisture, whereas the opposite was true for litter carbon concentration.
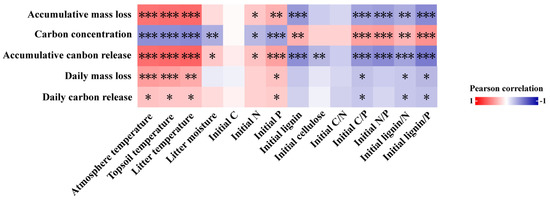
Figure 7.
Pearson’s correlation analysis between the mass loss rate, carbon concentration, and release rate and the temperature, moisture, and initial nutrient parameters of foliar litter. * p < 0.05, ** p < 0.01, *** p < 0.001.
4. Discussion
4.1. Effects of Typhoon Disturbance on Litter Quality
Typically, the initial quality of litter in a given ecosystem is the most influential element in litter decomposition [9]. Climate, plant population characteristics [49], and soil physicochemical properties [50], generally influence the quality of litter. Additionally, human actions (chemical fertilization [51], heavy metal pollution [52]) and particular meteorological occurrences such as tropical storms [28], and snowfall [20], might also alter litter quality. In subtropical urban forests affected by typhoons, abnormal litter contains higher initial concentrations of carbon, N, and P, but lower levels of lignin and cellulose. In addition, the initial C/N, C/P, lignin/N, and lignin/P ratios were lower in the abnormal litter (Table 1), indicating a higher quality substrate at the beginning of its decomposition. In the subtropical broadleaf evergreen forest on Okinawa Island, Japan [14] and the subtropical tidal wetlands of the Minjiang estuary [15], which are likewise subtropical regions, not only did the quantity of litter increase significantly after the typhoon but also the concentration of nutrients such as N and P in the litter increased significantly. An investigation of simulated tropical storm disturbances in a subtropical forest in Puerto Rico revealed that N and P concentrations in the litter layer increased considerably after a tropical storm [53]. As leaves aged, the degree of lignification and fibrillation increase, leading to an increase in lignin and cellulose concentrations. Plants have an “internal recycling” protective mechanism that recycles key nutrients prior to leaf senescence [54]. Therefore, the higher nutrient concentrations of abnormal foliar litter induced by a typhoon may be attributable to the absence of nutrient recycling prior to leaf fall. In addition, compared with the physiological litter, the extent to which the initial nutrient concentration of abnormal litter varied with tree species. On the one hand, the ability of the plant to regulate its own homeostasis varies between tree species due to the different abilities of different tree species to resist typhoon disturbances [35,55]; on the other hand, the nutrient resorption capacity of different tree species also varies. For example, for alder and acacia, which produce leaf litter with as high a concentration of N as high as that of live leaves, the difference range is c. 2%–3% [12]. Therefore, the differences between tree species result in distinct initial substrate qualities for the decomposition of abnormal litter, which then significantly affects the decomposition process.
According to the US National Weather Service’s classification scale for tropical storm damage, there are four classes: F0 through F3, with sustained wind speeds of 18–25, 26–35, 36–47, and 48–62 m/s, respectively. However, our study only focused on Typhoon Hagupit, which occurred on 4 August 2020, and affected Taizhou city with wind speeds of ca. 17.2–32.6 m/s in the wind circle and a rating of c. 8–10 level (https://e.weather.com.cn/mtqzt/3367215.shtml) (accessed on 30 September 2022). The effects of typhoons on abnormal litter production and quality are not yet fully understood [56]. To better understand the tropical cyclones on the initial quality of abnormal litter, it is necessary to further investigate the changes in abnormal litter production and quality with the intensity, disturbance frequency, and the occurrence time of tropical cyclones, as well as tree species and forest types.
4.2. Effects of Typhoon Disturbance on Abnormal Litter Mass Loss
The litter decomposition characteristics are climatically zonal [57], with litter annual mass loss rates in subtropical forests generally ranging from 40%–70% [58]. In our study, the annual mass loss of litter from different sources of the three species in the subtropical coastal urban forest ranged from 40.01%–66.55% in line with them, and the mass loss rates were considerably affected by the litter sources. Compared with the physiological litter, the rates of accumulative mass loss and decomposition of abnormal litter increased by 7.72%–29.78% and 8.00%–42.86%, respectively. This result agreed with Xu et al. [14], who have observed that tropical cyclone-generated litter has a higher decomposition rate in a subtropical forest in Okinawa. The rapid decomposition of the typhoon-produced foliar litter may be attributed to its distinct features of both physical structure and initial chemical concentrations [59]. The typhoon-produced green leaves are soft and tender in texture, not as hard as normal wilted leaves, and they have higher nutrient concentrations (e.g., N and P) and substrate quality. Increasingly investigations have shown that the initial N concentrations of litter, C/N [60], and lignin/N [61], are excellent predictors of decomposition. The higher the quality of litter is, the greater its soluble nutrition concentrations and the greater its leaching loss [27]. Our results also indicated that the initial concentrations of N, P, and lignin in physiological and typhoon-produced litter, and other substrate quality indexes, were closely correlated with the accumulative mass loss rate and daily mass loss, and well explained the predictors of litter decomposition (Figure 7). Besides, the differences of litter quality determined by tree species also affected litter mass loss and decomposition rates (Table 3). After one-year decomposition, the camphor tree foliar litter had the lowest rate of mass loss among the three tree species, owing to abnormal litter quality from different tree species. On the one hand, the foliar litter of the camphor tree has a waxy surface, which is more resistant to decomposition; on the other hand, the foliar litter of the camphor tree has lower initial N and P concentrations, and higher lignin concentration, C/N, and C/P ratios (Table 1), resulting in the lower camphor tree litter mass loss rate. In other words, the predictive indicators of physiological litter decomposition can be used to predict typhoon-induced abnormal litter decomposition.
Additionally, litter mass loss rate differed according to the critical period (Table 3, Figure 3). The litter mass loss occurred primarily during the ALP (0–92 d) and after the temperature warming (SLP, EPR, and LPR), reaching 17.04–40.34% and 15.98%–29.17%, respectively, and reaching a new peak of 8.31%–20.73% during the EPR; during the WNG, the lowest loss rate was only 3.16%–11.39%. The consensus about the decomposition process of litter is that litter has a high rate of mass loss in the earlier stages, a low rate of mass loss in the later stages, and a decomposition process with decreasing differences in the rate of mass loss between time periods over time [62]. On the one hand, the early decomposition period has the highest quality of foliar litter, and a large quantity of soluble carbohydrates and nutrients are released by leaching [63]. At the same time, it attracts the colonization and reproduction of soil organisms [64], both of which increase litter mass loss. On the other hand, the temperature and moisture of the decomposing environment are the most critical environmental factors affecting litter decomposition [65]. Our result also demonstrated that within a certain range, the foliar litter mass loss rate increased with increasing temperature and relative moisture. Under favorable temperature and moisture conditions, it is beneficial to increase soil organism activity [66] and associated decomposition enzyme activity [19], concurrently with strong leaching of foliar litter during the EPR, resulting in an increase in the foliar litter mass loss rate after the temperature rebounded and reached another peak in the EPR. In contrast, cold temperatures limit the soil organism activity significantly [67]. Consequently, the decomposition process of foliar litter is inhibited.
4.3. Effects of Typhoon Disturbance on Abnormal Litter Carbon Release
The accumulative carbon rates were considerably affected by the litter sources (Table 3). Compared with the physiological litter, the accumulative carbon release rates of abnormal litter increased by 9.1%–19.55% (Figure 5), which is consistent with the findings of Wang et al. [15]. Since carbon is the foundation of the composition of an organism, the total carbon released from litter is generally congruent with the process of mass loss [12]. As with mass loss, the rapid carbon release of the typhoon-produced foliar litter may be attributed to its distinct features of both physical structure and initial chemical concentrations. The results of correlation analyses on accumulative carbon release rates and daily carbon releases with initial nutrient concentrations and substrate quality, also indicate that the high substrate quality characteristics of abnormal litter contribute to increased carbon release (Figure 7). Tree species differences also significantly affected the carbon release rate of litter (Table 3). After one-year decomposition, the lowest rate of accumulative carbon release from abnormal litter of camphor tree was only 62.13%. Compared with the other two species, the indecomposable structure and abundant recalcitrant compounds in camphor tree litter (the highest lignin concentration in camphor tree litter, Table 1) were the primary reasons for its lower carbon release rate. The dynamics of litter carbon release varied with the critical periods. Litter carbon release rates peaked at 19.41%–48.65% and 2.29%–19.21% during ALP (0–92 d) and EPR, respectively, while the lowest was only −0.85%–6.29% during WNG, which is generally consistent with the characteristics of mass loss. The higher nutrient concentration in the early decomposing phase, along with the litter fragmentation during decomposition, the soluble sugars and small-molecule organic matter by leaching loss, and the utilization of litter nutrients by the soil biotope organisms [12], all contribute to litter carbon release. The temperature and moisture of the decomposing environment also influence carbon release rates. Within a certain range, the rate of foliar litter carbon release increased with increasing temperature and moisture (Figure 7). During WNG, cold temperatures largely limited the soil organism activity significantly [67], and the decomposition process of foliar litter is inhibited. The rate of carbonaceous organic matter decomposition is also slowed, reducing carbon release rates. It is worth noting that there is a negative carbon release during the WNG and LPR. This indicates that carbon accumulates during the litter decomposition process, possibly because the decomposition rate of material with other elemental concentrations of the litter is greater than the carbon release rate at this phase. It is accompanied by the accumulation of recalcitrant material (such as lignin and condensed phenolic compounds) that is difficult to decompose [68,69], leading to an increase in the proportion of carbon in the total weight of the litter, manifested as litter carbon concentration increase and carbon accumulation. Therefore, as mentioned above, typhoons drive material cycle processes in subtropical urban forests by altering the production and quality of litter, increasing the rate of mass loss and carbon release from litter. With global climate change, the intensity and frequency of tropical cyclones will continue to increase, as will the proportion of abnormal litter. This will then contribute to the forest ecosystem’s carbon and nutrient cycling processes [70].
5. Main Findings, Conclusions and Perspectives
(1) Compared with the physiological litter, typhoon-produced abnormal foliar litter has higher concentrations of carbon, nitrogen, and phosphorus, and lower ratios of C/N, C/P, lignin/N, and lignin/P.
(2) Compared with physiological litter, typhoon-produced abnormal litter has a higher mass loss and carbon release, implying that typhoons can accelerate the cycle of carbon in subtropical urban forest ecosystems. Under climate change scenarios, the intensity and frequency of tropical cyclones will continue to increase, as will the proportion of abnormal litter. The result suggests that tropical cyclones (hurricane/typhoon) accelerate the carbon cycle of coastal urban forest ecosystems through abnormal litter decomposition in the future.
(3) The rates of mass loss rate and the carbon release of abnormal and physiological litter varied greatly with tree species and critical periods, implying that changing the tree species composition in coastal subtropical urban forest ecosystems.
Future research should emphasize the following aspects. First of all, the production and quality of physiological and abnormal litter vary with the intensity, frequency, and the occurrence time of tropical cyclones, as well as tree species, forest types, stand characteristics, and site conditions. In addition, the relative contribution of physiological and tropical cyclone-induced abnormal litter decomposition to soil organic carbon accumulation and stability changes with tropical cyclones and forest characteristics, and in turn provides key scientific data for improving global carbon cycle prediction. Last but not least, what is the mechanism by which tropical cyclone-induced abnormal litter decomposition forms soil organic matter?
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f13111819/s1. Figure S1. The daily mean temperature dynamics of the atmosphere, topsoil, and foliar litter in the experimental area of the litter incubation experiment from 10 September 2020, to 21 July 2021. Figure S2. The moisture dynamics of goldenrain tree (a), camphor tree (b), and weeping willow tree (c) foliar litter at five key decomposition periods. ALP, WNG, SLP, EPR, and LPR indicate the autumn litter peak period, winter nongrowing season, spring litter peak period, early plum-rainy season, and late plum-rainy season, respectively. PL and AL indicate physiological and abnormal litter, respectively. Different capital letters indicate significant differences between different key periods (p < 0.05). Different lowercase letters indicate significant differences between different sources of foliar litter (p < 0.05).
Author Contributions
W.Y. and X.X. conceived the ideas and designed methodology. H.W., B.Z., S.S., Y.J. and Q.Z. conducted the experiments and collected the data. H.W., Z.W. and R.C. analyzed the data. H.W., W.Y. and X.X. led the writing of the manuscript. All authors contributed critically to the drafts and approved the final version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (grant numbers 32071554).
Data Availability Statement
The data used for this research can be found in the following repository (https://doi.org/10.6084/m9.figshare.21214493, accessed on 30 September 2022).
Acknowledgments
The authors of this study appreciate the field assistance of the Plant Ecology Research Base at Taizhou University provided by Feihai Yu.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Lu, X.; Yu, H.; Ying, M.; Zhao, B.; Zhang, S.; Lin, L.; Bai, L.; Wan, R. Western North Pacific tropical cyclone database created by the China Meteorological Administration. Adv. Atmos. Sci. 2021, 38, 690–699. [Google Scholar] [CrossRef]
- Xi, W. Synergistic effects of tropical cyclones on forest ecosystems: A global synthesis. J. For. Res. 2015, 26, 1–21. [Google Scholar] [CrossRef]
- Pruitt, J.N.; Little, A.G.; Majumdar, S.J.; Schoener, T.W.; Fisher, D.N. Call-to-action: A global consortium for tropical cyclone ecology. Trends Ecol. Evol. 2019, 34, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Vandermeer, J.; Granzow, D.L.; Cerda, I.; Boucher, D.; Perfecto, I.; Ruiz, J.E. Hurricane disturbance and tropical tree species diversity. Science 2000, 290, 788–791. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Chamber, J.Q.; Negron-Juarez, R.I.; Hurtt, G.C.; Baker, D.B.; Powell, M.D. Impacts of tropical cyclones on U.S. Forest tree mortality and carbon flux from 1851 to 2000. Proc. Natl. Acad. Sci. USA 2009, 106, 7888–7892. [Google Scholar] [CrossRef]
- Chambers, J.Q.; Fisher, J.; Zeng, H.; Chapman, E.L.; Baker, D.B.; Hurtt, G.C. Hurricane Katrina’s carbon footprint on U.S. Gulf coast forests. Science 2007, 318, 1107. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021: The Physical Science Basis; Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Vitousek, P.M. Litterfall, nutrient cycling, and nutrient limitation in tropical forests. Ecology 1984, 65, 285–298. [Google Scholar] [CrossRef]
- Krishna, M.P.; Mohan, M. Litter decomposition in forest ecosystems: A review. Energy Ecol. Environ. 2017, 2, 236–249. [Google Scholar] [CrossRef]
- Bray, J.R.; Gorham, E. Litter production in forests of the world. Adv. Ecol. Res. 1964, 2, 101–157. [Google Scholar]
- Malhi, Y.; Doughty, C.; Galbraith, D. The allocation of ecosystem net primary productivity in tropical forests. Philos. Trans. R Soc. Lond. 2011, 366, 3225–3245. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Raich, J.W.; Schlesinger, W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B 1992, 44, 81–99. [Google Scholar] [CrossRef]
- Xu, X.; Hirata, E.; Enoki, T.; Tokashiki, Y. Leaf litter decomposition and nutrient dynamics in a subtropical forest after typhoon disturbance. Plant Ecol. 2004, 173, 161–170. [Google Scholar] [CrossRef]
- Wang, W.; Sardans, J.; Tong, C.; Wang, C.; Ouyang, L.; Bartrons, M.; Penuelas, J. Typhoon enhancement of N and P release from litter and changes in the litter N:P ratio in a subtropical tidal wetland. Environ. Res. Lett. 2016, 11, 014003. [Google Scholar] [CrossRef]
- Beard, K.H.; Vogt, K.A.; Vogt, D.J.; Scatena, F.N.; Covich, A.P.; Sigurdardottir, R.; Siccama, T.G.; Crowl, T.A. Structural and functional responses of a subtropical forest to 10 years of hurricanes and droughts. Ecol. Monogr. 2005, 75, 345–361. [Google Scholar] [CrossRef]
- Lodge, D.J.; Scatena, F.N.; Asbury, C.E.; Sanchez, M.J. Fine litterfall and related nutrient inputs resulting from hurricane Hugo in subtropical wet and lower montane rain forests of Puerto Rico. Biotropica 1991, 23, 336–342. [Google Scholar] [CrossRef]
- Liu, X.; Zeng, X.; Zou, X.; González, G.; Wang, C.; Yang, S. Litterfall production prior to and during hurricanes Irma and Maria in four Puerto Rican forests. Forests 2018, 9, 367. [Google Scholar] [CrossRef]
- Wang, H.C.; Wang, S.; Lin, K.; Shaner, P.J.L.; Lin, T.C. Litterfall and element fluxes in a natural hardwood forest and a Chinese-fir plantation experiencing frequent typhoon disturbance in central Taiwan. Biotropica 2013, 45, 541–548. [Google Scholar] [CrossRef]
- Yang, Q.; Xu, M.; Chi, Y.; Zheng, Y.; Ruichang, S.; Wang, S. Effects of freeze damage on litter production, quality and decomposition in a loblolly pine forest in central China. Plant Soil 2014, 374, 449–458. [Google Scholar] [CrossRef]
- Wu, Z.; Li, Y.; Zhou, G.; Chen, B. Abnormal litterfall and its ecological significance. Sci. Silvae Sin. 2008, 44, 28–31. [Google Scholar]
- Coûteaux, M.M.; Bottner, P.; Berg, B. Litter decomposition, climate and liter quality. Trends Ecol. Evol. 1995, 10, 63–66. [Google Scholar] [CrossRef]
- Parton, W.; Silver, W. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 2007, 315, 940. [Google Scholar] [CrossRef] [PubMed]
- González, G.; Seastedt, T.R. Soil fauna and plant litter decomposition in tropical and subalpine forests. Ecology 2001, 82, 955–964. [Google Scholar] [CrossRef]
- Yan, J.; Wang, L.; Hu, Y.; Tsang, Y.F.; Zhang, Y.; Wu, J.; Fu, X.; Sun, Y. Plant litter composition selects different soil microbial structures and in turn drives different litter decomposition pattern and soil carbon sequestration capability. Geoderma 2018, 319, 194–203. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, S.; Tong, F.; Chen, B.; Wu, Z.; Luo, X. Effects of abnormal litter input on forest soil organic carbon after ice-storm: A case of Nanling. Ecol. Environ. Sci. 2013, 22, 1504–1513. [Google Scholar]
- Li, R.; Zhang, Y.; Yu, D.; Wang, Y.; Zhao, X.; Zhang, R.; Zhang, W.; Wang, Q.; Xu, M.; Chen, L.; et al. The decomposition of green leaf litter is less temperature sensitive than that of senescent leaf litter: An incubation study. Geoderma 2021, 381, 114691. [Google Scholar] [CrossRef]
- Lin, T.C.; Hamburg, S.P.; Lin, K.C.; Wang, L.J.; Chang, C.T.; Hsia, Y.; Vadeboncoeur, M.; McMullen, C.; Liu, C.P. Typhoon disturbance and forest dynamics: Lessons from a Northwest Pacific subtropical forest. Ecosystems 2011, 14, 127–143. [Google Scholar] [CrossRef]
- Liu, D.; Pang, L.; Xie, B. Typhoon disaster in China: Prediction, prevention, and mitigation. Nat. Hazards 2009, 49, 421–436. [Google Scholar] [CrossRef]
- Xuan, W.; Ma, C.; Kang, L.; Gu, H.; Pan, S.; Xu, Y. Evaluating historical simulations of CMIP5 GCMs for key climatic variables in Zhejiang province, China. Theor. Appl. Climatol. 2017, 128, 207–222. [Google Scholar] [CrossRef]
- Xie, F.; Liu, Y. China Meteorological Yearbook; Meteorological Press: Beijing, China, 2020. [Google Scholar]
- Huang, S.; Shen, H.; Chen, Z. Investigation on the influence of typhoon Mangkhut on urban trees in Guangdong-Hongkong-Macao Greater Bay Area. Guangdong Landsc. Archit. 2020, 42, 26–31. [Google Scholar]
- Gavito, M.; Sandoval-Pérez, A.; Castillo, K.; Cohen-Salgado, D.; Colarte-Avilés, M.; Mora, F.; Santibáñez-Rentería, A.; Siddique, I.; Urquijo-Ramos, C. Resilience of soil nutrient availability and organic matter decomposition to hurricane impact in a tropical dry forest ecosystem. For. Ecol. Manag. 2018, 426, 81–90. [Google Scholar] [CrossRef]
- Kominoski, J.; Weaver, C.; Armitage, A.; Pennings, S. Coastal carbon processing rates increase with mangrove cover following a hurricane in Texas, USA. Ecosphere 2022, 13, e4007. [Google Scholar] [CrossRef]
- Wang, H.; Xu, X.; Yang, W.; Cao, R.; Wang, Z.; Zheng, B.; Lv, H.; Liu, T. The ecological stoichiometry of carbon, nitrogen and phosphorus in urban garden plants with different life forms and its response to typhoon Hagupit. Acta Ecol. Sin. 2021, 41, 8931–8938. [Google Scholar]
- Wang, H.; Xu, X.; Yang, W.; Cao, R.; Wang, Z.; Li, F.; Zheng, B.; Liang, Y. Responses of metallic nutrient concentrations in urban garden plants leaves with different life forms to typhoon Hagupit. Chin. J. Appl. Environ. Biol. 2022, 28, 1–10. [Google Scholar]
- Wang, L.; Li, X.; Xu, Z. Analysis on climatic characteristics of typhoon over the past 50 years at Zhoushan. Mar. Forecast. 2011, 28, 36–43. [Google Scholar]
- Dong, J.; Huang, X. Typhoon track classification and storm fallout analysis for landfall in Zhejiang. Zhejiang Meteorol. 2019, 40, 13–19. [Google Scholar]
- Swift, M.; Heal, O.; Anderson, J. Decomposition in Terrestrial Ecosystems; University of California Press: Berkeley, CA, USA, 1979. [Google Scholar]
- Klotzbücher, T.; Kaiser, K.; Guggenberger, G.; Gatzek, C.; Kalbitz, K. A new conceptual model for the fate of lignin in decomposing plant litter. Ecology 2011, 92, 1052–1062. [Google Scholar] [CrossRef]
- Lengauer, W. On the application of the Dumas technique for the determination of nitrogen in refractory nitrides. Talanta 1991, 38, 659–663. [Google Scholar] [CrossRef]
- Lu, R.K. Soil and Agro-Chemical Analytical Methods; Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Rowland, A.P.; Roberts, J.D. Lignin and cellulose fractionation in decomposition studies using acid-detergent fibre methods. Commun. Soil Sci. Plant Anal. 1994, 25, 269–277. [Google Scholar] [CrossRef]
- He, W.; Wu, F.; Yang, W.; Tan, B.; Zhao, Y.; Wu, Q.; He, M. Lignin degradation in foliar litter of two shrub species from the gap center to the closed canopy in an alpine fir forest. Ecosystems 2016, 19, 115–128. [Google Scholar] [CrossRef]
- Peng, Y.; Yang, W.; Li, J.; Wang, B.; Zhang, C.; Yue, K.; Wu, F. Contribution of soil fauna to foliar litter-mass loss in winter in an ecotone between dry valley and montane forest in the upper reaches of the Minjiang river. PLoS ONE 2015, 10, e0124605. [Google Scholar] [CrossRef]
- Zhu, J.; Wu, F.; Yang, W.; Tan, B. Decomposition of Abies faxoniana litter varies with freeze–thaw stages and altitudes in subalpine/alpine forests of southwest China. Scand. J. For. Res. 2012, 27, 586–596. [Google Scholar] [CrossRef]
- Ma, Z.; Yang, W.; Wu, F.; Tan, B. Effects of light intensity on litter decomposition in a subtropical region. Ecosphere 2017, 8, e01770. [Google Scholar] [CrossRef]
- Berg, B. Decomposing litter; limit values; humus accumulation, locally and regionally. Appl. Soil Ecol. 2018, 123, 494–508. [Google Scholar] [CrossRef]
- Petraglia, A.; Cacciatori, C.; Chelli, S.; Fenu, G.; Calderisi, G.; Gargano, D.; Abeli, T.; Orsenigo, S.; Carbognani, M. Litter decomposition: Effects of temperature driven by soil moisture and vegetation type. Plant Soil 2019, 435, 187–200. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, L.; Li, H.; He, H.; Wei, Y.; Luo, J.; Zhang, G.; Huang, Y.; Li, Y.; Zhou, H. Soil physicochemical properties and vegetation structure along an elevation gradient and implications for the response of alpine plant development to climate change on the northern slopes of the Qilian Mountains. J. Mt. Sci. 2018, 15, 1006–1019. [Google Scholar] [CrossRef]
- Wollenweber, B. Nitrogen in terrestrial ecosystems: Questions of productivity, vegetational changes and ecosystem stability. Holocene 1992, 2, 187. [Google Scholar] [CrossRef]
- Berg, B.; Ekbohm, G.; Söderström, B.; Staaf, H. Reduction of decomposition rates of scots pine needle litter due to heavy-metal pollution. Water Air Soil Pollut. 1991, 59, 165–177. [Google Scholar] [CrossRef]
- Silver, W.L.; Hall, S.J.; González, G. Differential effects of canopy trimming and litter deposition on litterfall and nutrient dynamics in a wet subtropical forest. For. Ecol. Manag. 2014, 332, 47–55. [Google Scholar] [CrossRef]
- Gosz, J. Nitrogen cycling in coniferous ecosystems. Ecol. Bull. 1981, 33, 405–426. [Google Scholar]
- Bellingham, P.J.; Tanner, E.V.J.; Healey, J.R. Damage and responsiveness of Jamaican montane tree species after disturbance by a hurricane. Ecology 1995, 76, 2562–2580. [Google Scholar] [CrossRef]
- Hernandez, J.; Maldia, L.; Park, B. Research Trends and Methodological Approaches of the Impacts of Windstorms on Forests in Tropical, Subtropical, and Temperate Zones: Where Are We Now and How Should Research Move Forward? Plants 2020, 9, 1709. [Google Scholar] [CrossRef] [PubMed]
- Berg, B.; Berg, M.P.; Bottner, P.; Box, E.; Breymeyer, A.; de Anta, R.C.; Coûteaux, M.; Escudero, A.; Gallardo, A.; Kratz, W.; et al. Litter mass loss rates in pine forests of Europe and Eastern United States: Some relationships with climate and litter quality. Biogeochemistry 1993, 20, 127–159. [Google Scholar] [CrossRef]
- Xu, X.; Hirata, E. Decomposition patterns of leaf litter of seven common canopy species in a subtropical forest: N and P dynamics. Plant Soil 2005, 273, 279–289. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Cornelissen, J.; Vendramini, F.; Cabido, M.; Castellanos, A. Chemistry and toughness predict leaf litter decomposition rates over a wide spectrum of functional types and taxa in central Argentina. Plant Soil 2000, 218, 21–30. [Google Scholar] [CrossRef]
- Taylor, B.; Parkinson, D.; Parkinson, W. Nitrogen and lignin content as predictors of litter decay rates: A microcosm test. Ecology 1989, 70, 97–104. [Google Scholar] [CrossRef]
- Camiré, C.; Trofymow, J.A.; Duschene, L.; Moore, T.; Kozak, L.M.; Titus, B.; Kranabetter, J.; Prescott, C.; Visser, S.; Morrison, I.; et al. Rates of litter decomposition over 6 years in Canadian forests: Influence of litter quality and climate. Can. J. For. Res. 2002, 32, 789–804. [Google Scholar]
- Garcia-Pausas, J.; Casals, P.; Romanyà, J. Litter decomposition and faunal activity in Mediterranean forest soils: Effects of N content and the moss layer. Soil Biol. Biochem. 2004, 36, 989–997. [Google Scholar] [CrossRef]
- Moore, T.R.; Trofymow, J.A.; Prescott, C.E.; Titus, B.D. Nature and nurture in the dynamics of C, N and P during litter decomposition in Canadian forests. Plant Soil 2011, 339, 163–175. [Google Scholar] [CrossRef]
- Yang, X.; Chen, J. Plant litter quality influences the contribution of soil fauna to litter decomposition in humid tropical forests, southwestern China. Soil Biol. Biochem. 2009, 41, 910–918. [Google Scholar] [CrossRef]
- Salinas, N.; Malhi, Y.; Meir, P.; Silman, M.; Roman, C.R.; Huaman, J.; Salinas, D.; Huaman, V.; Gibaja, A.; Mamani, M.; et al. The sensitivity of tropical leaf litter decomposition to temperature: Results from a large-scale leaf translocation experiment along an elevation gradient in Peruvian forests. New Phytol. 2011, 189, 967–977. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. The temperature dependence of soil organic matter decomposition, and the effect of global warming on soil organic C storage. Soil Biol. Biochem. 1995, 27, 753–760. [Google Scholar] [CrossRef]
- Zhou, Y.; Clark, M.; Su, J.; Xiao, C. Litter decomposition and soil microbial community composition in three Korean pine (Pinus koraiensis) forests along an altitudinal gradient. Plant Soil 2015, 386, 171–183. [Google Scholar] [CrossRef]
- Yue, K.; Wu, F.; Yang, W.; Zhang, C.; Peng, Y.; Tan, B.; Xu, Z.; Huang, C. Cellulose Dynamics during Foliar Litter Decomposition in an Alpine Forest Meta-Ecosystem. Forests 2016, 7, 176. [Google Scholar] [CrossRef]
- He, W.; Yang, W. Loss of total phenols from leaf litter of two shrub species: Dual responses to alpine forest gap disturbance during winter and the growing season. J. Plant Ecol. 2020, 13, 369–377. [Google Scholar] [CrossRef]
- Jaramillo, V.; Martínez-Yrízar, A.; Machado, L. Hurricane-Induced Massive Nutrient Return via Tropical Dry Forest Litterfall: Has Forest Biogeochemistry Resilience Changed? Ecosystems 2022. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).