
Bioethanol Production Potential and Other Biomass Energy Properties of Invasive Reynoutria, Solidago, and Spiraea Plants



Abstract
1. Introduction
2. Materials and Methods
2.1. Invasive Plant Species and Preparation of Biomass
2.2. Analysis of the Chemical Composition of Plant Biomass
- -
- Extractive contents were determined using 96% ethanol according to Soxhlet (TAPPI-T204 cm-07) [76].
- -
- Cellulose content was determined by the Seifert method using a mixture of acetylacetone and dioxane [77].
- -
- Lignin content was determined by the Tappi (Technical Association of the Pulp and Paper Industry) method (T-222 om-06) using concentrated sulfuric acid [78].
- -
- Pentosans were determined using phloroglucinol [28].
- -
- The theoretical hemicellulose content was arithmetically calculated as the difference in holocellulose and cellulose [28].
- -
- The ash content, important for the combustion of biomass, was determined according to the DIN 51731 standards [79].
2.3. Determination of the Heat of Combustion and Calorific Value
- C is the heat capacity of the calorimeter, 12,783.69 (J·°C−1);
- Dt is the temperature rise in the main period (°C);
- k is the correction for heat exchange with the surroundings (°C);
- c is the sum of corrections for additional thermal effects (J);
- m is the mass of the fuel sample (g).
- is the average gross calorific value of solid fuel in the analytical state (J·g−1);
- The heat of vaporization of water at 25 °C is 24.42, corresponding to 1% of water in the fuel (J/g);
- Wa is the moisture content in the analytical sample of fuel (%);
- Ha is the hydrogen content in the analytical sample of fuel.
2.4. The Process of Obtaining Bioethanol
3. Results and Discussion
3.1. Analysis of Chemical Composition of Invasive Plants
3.2. Higher Heating Value and Lower Heating Value
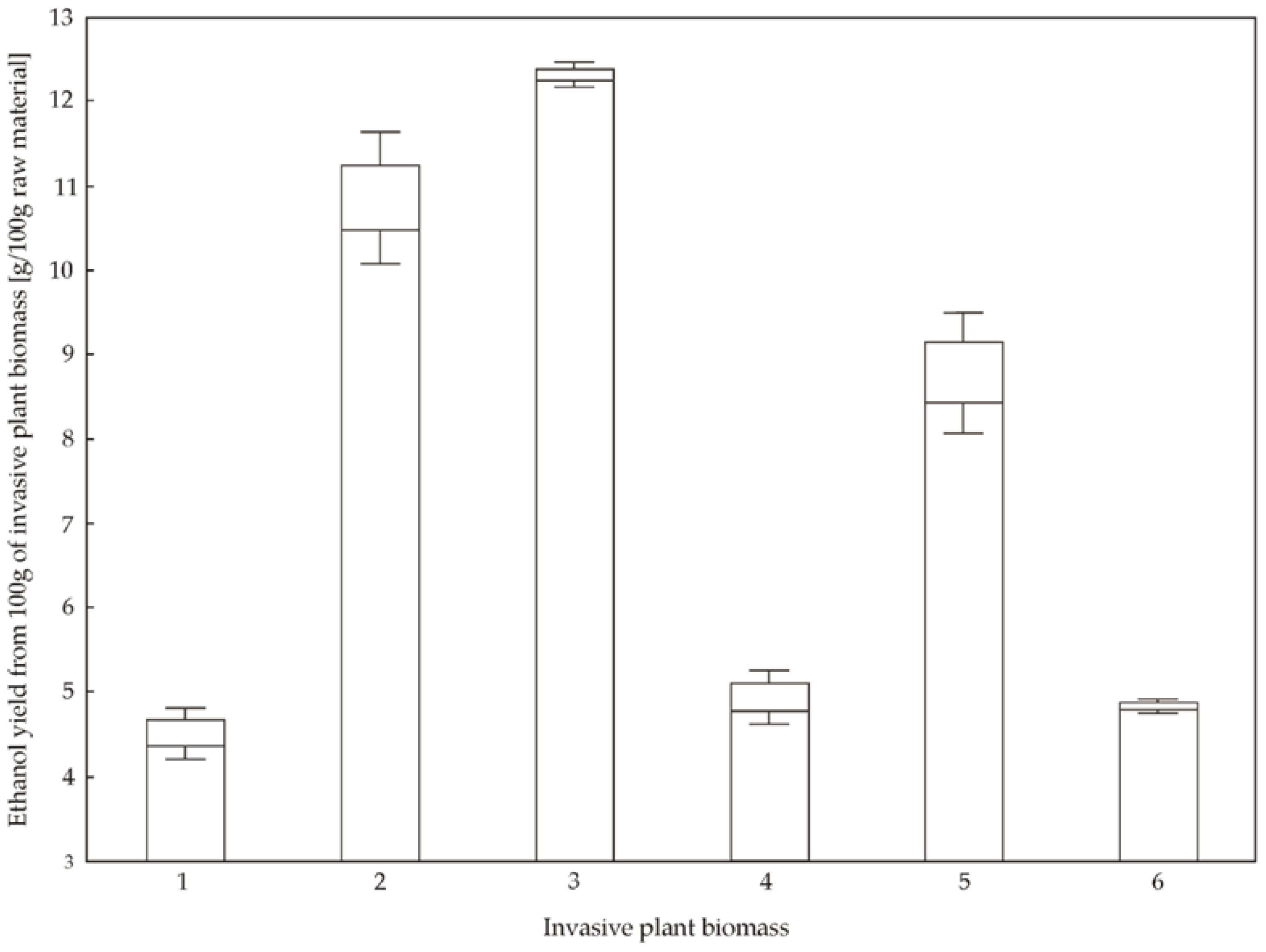
3.3. Potential of Bioethanol Production
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Taxon | Co-Ordinates | Harvest Date | |
|---|---|---|---|
| N | E | ||
| Reynoutria japonica | 52°28’49.45′′ | 17°17’00.77′′ | 13 June 2021 |
| Reynoutria sachalinensis | 52°29’12.88′′ | 17°50’31.35′′ | 13 June 2021 |
| Reynoutria × bohemica | 52°25’12.98′′ | 16°53’36.75′′ | 14 June 2021 |
| Solidago canadensis | 51°23’47.48′′ | 15°10’11.98′′ | 24 July 2021 |
| Solidago gigantea | 51°23’31.19′′ | 15°90’58.23′′ | 28 July 2021 |
| Spiraea tomentosa | 51°24’44.24′′ | 15°40’40.44′′ | 25 July 2021 |
References
- Woodford, D.J.; Richardson, D.M.; MacIsaac, H.J.; Mandrak, N.E.; Van Wilgen, B.W.; Weyl, O.L. Confronting the wicked problem of managing biological invasions. NeoBiota 2016, 31, 63–86. [Google Scholar] [CrossRef]
- Pearson, D.E.; Ortega, Y.K.; Eren, Ö.; Hierro, J.L. Community assembly theory as a framework for biological invasions. Trends Ecol. Evol. 2018, 33, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Ricciardi, A.; Iacarella, J.C.; Aldridge, D.C.; Blackburn, T.M.; Carlton, J.T.; Catford, J.A.; Dick, J.T.A.; Hulme, P.E.; Jeschke, J.M.; Liebhold, A.M. Four priority areas to advance invasion science in the face of rapid environmental change. Environ. Rev. 2021, 29, 119–141. [Google Scholar] [CrossRef]
- Soulé, M.E. Conservation: Tactics for a constant crisis. Science 1991, 253, 744–750. [Google Scholar] [CrossRef]
- Vilà, M.; Corbin, J.D.; Dukes, J.S.; Pino, J.; Smith, S.D. Linking plant invasions to global environmental change. In Terrestrial Ecosystems in a Changing World; Springer: Berlin/Heidelberg, Germany, 2007; pp. 93–102. [Google Scholar]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Butchart, S.H.; Walpole, M.; Collen, B.; Van Strien, A.; Scharlemann, J.P.; Almond, R.E.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global biodiversity: Indicators of recent declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef]
- Vitousek, P.M. Biological invasions and ecosystem processes: Towards an integration of population biology and ecosystem studies. In Ecosystem Management; Springer: New York, NY, USA, 1990; pp. 183–191. [Google Scholar]
- Charles, H.; Dukes, J.S. Impacts of invasive species on ecosystem services. In Biological Invasions; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 217–237. [Google Scholar]
- Pejchar, L.; Mooney, H.A. Invasive species, ecosystem services and human well-being. Trends Ecol. Evol. 2009, 24, 497504. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G. Ecosystem consequences of biological invasions. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 59–80. [Google Scholar] [CrossRef]
- Zhang, P.; Li, B.; Wu, J.; Hu, S. Invasive plants differentially affect soil biota through litter and rhizosphere pathways: A meta-analysis. Ecol. Lett. 2019, 22, 200–210. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Mark Lonsdale, W.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Mazza, G.; Tricarico, E. (Eds.) Invasive Species and Human Health; CABI Invasives Series: London, UK, 2018; Volume 10, p. 208. [Google Scholar]
- Kettunen, M.; Genovesi, P.; Gollasch, S.; Pagad, S.; Starfinger, U.; ten Brink, P.; Shine, C. Technical Support to EU Strategy on Invasive Alien Species (IAS); Institute for European Environmental Policy (IEEP): Brussels, Belgium, 2009; p. 124. [Google Scholar]
- Diagne, C.; Leroy, B.; Vaissière, A.-C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.M.; Corey, J.A.; Bradshaw, C.J.A.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571. [Google Scholar] [CrossRef]
- Moodley, D.; Angulo, E.; Cuthbert, R.N.; Leung, B.; Turbelin, A.; Novoa, A.; Kourantidou, M.; Heringer, G.; Haubrock, P.J.; Renault, D. Surprisingly high economic costs of biological invasions in protected areas. Biol. Invasions 2022, 24, 1995–2016. [Google Scholar] [CrossRef]
- Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions EU. Biodiversity Strategy for 2030 Bringing Nature Back into Our Lives (COM/2020/380 Final). Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:52020DC0380 (accessed on 30 March 2022).
- Wiatrowska, B.; Michalska-Hejduk, D.; Czarniecka-Wiera, M.; Derucka, M.; Derucki, M.; Mazurska, K.; Sierka, E.; Kompała-Bąba, A.; Dajdok, Z.; Krzysztofiak, L. (Eds.) General Directorate for Environmental Protection: Warsaw, Poland, 2022; p. 175. (In Polish)
- Sims, R.E. Bioenergy to mitigate for climate change and meet the needs of society, the economy and the environment. Mitig. Adapt. Strateg. Glob. Change 2003, 8, 349–370. [Google Scholar] [CrossRef]
- Raghu, S.; Anderson, R.C.; Daehler, C.C.; Davis, A.S.; Wiedenmann, R.N.; Simberloff, D.; Mack, R.N. Adding biofuels to the invasive species fire? Science 2006, 313, 1742. [Google Scholar] [CrossRef]
- Strašil, Z.; Kára, J. Study of knotweed (Reynoutria) as possible phytomass resource for energy and industrial utilization. Res. Agric. Eng. 2010, 56, 85–91. [Google Scholar] [CrossRef]
- Bajgai, R.C.; Tamang, D.T.; Kushwaha, A.; Goswami, L. Strategic consideration as feedstock resource for biofuel production as a holistic approach to control invasive plant species. In Waste-to-Energy Approaches Towards Zero Waste; Hussain, C.M., Singh, S., Goswami, L., Eds. Elsevier: Amsterdam, The Netherlands, 2022; pp. 245–268. [Google Scholar]
- Brunerova, A.; Muller, M.; Brozek, M. Potential of wild growing Japanese knotweed (Reynoutria japonica) for briquette production. In Proceedings of the 16th International Scientific Conference Engineering for Rural Development, Jelgava, Latvia, 24–26 May 2017; pp. 24–26. [Google Scholar]
- Zihare, L.; Soloha, R.; Blumberga, D. The potential use of invasive plant species as solid biofuel by using binders. EMU DSpace 2018, 16. [Google Scholar]
- Hennequin, L.M.; Polizzi, K.; Fennell, P.S.; Hallett, J.P. Rhododendron and Japanese Knotweed: Invasive species as innovative crops for second generation biofuels for the ionoSolv process. RSC Adv. 2021, 11, 18395–18403. [Google Scholar] [CrossRef]
- Lin, Y.; Zhao, Y.; Ruan, X.; Barzee, T.J.; Zhang, Z.; Kong, H.; Zhang, X. The potential of constructed wetland plants for bioethanol production. BioEnergy Res. 2020, 13, 43–49. [Google Scholar] [CrossRef]
- Prosiński, S. Wood Chemistry; PWRiL: Warsaw, Poland, 1984. (In Polish) [Google Scholar]
- Tofani, G.; Cornet, I.; Tavernier, S. Separation and recovery of lignin and hydrocarbon derivatives from cardboard. Biomass Conv. Bioref. 2020, 12, 3409–3424. [Google Scholar] [CrossRef]
- Peng, X.; Nie, S.; Li, X.; Huang, X.; Li, Q. Characteristics of the Water- and Alkali-Soluble Hemicelluloses Fractionated by Sequential Acidification and Graded-Ethanol from Sweet Maize Stems. Molecules 2019, 24, 212. [Google Scholar] [CrossRef]
- Baruah, J.; Nath, B.K.; Sharma, R.; Kumar, S.; Deka, R.C.; Baruah, D.C.; Kalita, E. Recent Trends in the Pretreatment of Lignocellulosic Biomass for Value-Added Products. Front. Energy Res. 2018, 6, 141. [Google Scholar] [CrossRef]
- Duque, A.; Álvarez, C.; Doménech, P.; Manzanares, P.; Moreno, A.D. Advanced Bioethanol Production: From Novel Raw Materials to Integrated Biorefineries. Processes 2021, 9, 206. [Google Scholar] [CrossRef]
- Vasić, K.; Knez, Ž.; Leitgeb, M. Bioethanol Production by Enzymatic Hydrolysis from Different Lignocellulosic Sources. Molecules 2021, 26, 753. [Google Scholar] [CrossRef] [PubMed]
- Rodionova, M.V.; Bozieva, A.M.; Zharmukhamedov, S.V.; Leong, Y.K.; Lan, J.C.-W.; Veziroglu, A.; Veziroglu, T.N.; Tomo, T.; Chang, J.-S.; Allakhverdiev, S.I. A comprehensive review on lignocellulosic biomass biorefinery for sustainable biofuel production. Int. J. Hydrog. Energy 2022, 47, 1481–1498. [Google Scholar] [CrossRef]
- Pascoli, D.U.; Suko, A.; Gustafson, R.; Gough, H.L.; Bura, R. Novel ethanol production using biomass preprocessing to increase ethanol yield and reduce overall costs. Biotechnol. Biofuels 2021, 14, 9. [Google Scholar] [CrossRef]
- Kumar, D.; Juneja, A.; Hohenschuh, W.; Williams, J.D.; Murthy, G.S. Chemical composition and bioethanol potential of different plant species found in Pacific Northwest conservation buffers. J. Renew. Sustain. Energy 2012, 4, 6. [Google Scholar] [CrossRef]
- Sheng, C.; Azevedo, J.L.T. Estimating the higher heating value of biomass fuels from basic analysis data. Biomass Bioenergy 2005, 28, 499–507. [Google Scholar] [CrossRef]
- Qian, X.; Lee, S.; Soto, A.M.; Chen, G. Regression model to predict the higher heating value of poultry waste from proximate analysis. Resources 2018, 7, 39. [Google Scholar] [CrossRef]
- Directive 2009/28/EC of the European Parliament and of the Council of 23 April 2009 on the Promotion of the Use of Energy from Renewable Sources and Amending and Subsequently Repealing Directives 2001/77/EC and 2003/30/EC. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX:32009L0028 (accessed on 8 March 2022).
- Flora of China. Fact Sheets: Reynoutria japonica. Available online: http://www.efloras.org/florataxon.aspx?flora_id=2&taxon_id=242343249 (accessed on 30 March 2022).
- Sukopp, H.; Starfinger, U. Reynoutria sachalinensis in Europe and in the Far East: A comparison of the species ecology in its native and adventive distribution range. In Plant Invasions—General Aspects and Special Problems; Pyšek, P., Prach, K., Rejmánek, M., Wade, M., Eds.; SPB Academic: Amsterdam, The Netherlands, 1995; pp. 151–159. [Google Scholar]
- Mandák, B.; Pyšek, P. Druhy rodu Reynoutria na území České republiky. Zprávy České botanické společnosti Materiály 1997, 32, 45–57. (In Czech) [Google Scholar]
- Bailey, J.P.; Conolly, A.P. Prize-winners to pariahs—A history of Japanese knotweed s.l. (Polygonaceae) in the British Isles. Watsonia 2000, 23, 93–110. [Google Scholar]
- Mandák, B.; Pyšek, P.; Bímová, K. History of the invasion and distribution of Reynoutria taxa in the Czech Republic: A hybrid spreading faster than its parents. Preslia 2004, 76, 15–64. [Google Scholar]
- Bailey, J.; Wisskirchen, R. The distribution and origins of Faúopia× bohemica (Polygonaceae) in Europe. Nord. J. Bot. 2000, 24, 173–199. [Google Scholar] [CrossRef]
- Drazan, D.; Smith, A.G.; Anderson, N.O.; Becker, R.; Clark, M. History of knotweed (Fallopia spp.) invasiveness. Weed Sci. 2021, 69, 617–623. [Google Scholar] [CrossRef]
- Chrtek, J.; Chrtková, A. Reynoutria × bohemica, a new hybrid from the dock family. Časopis Nàrodniho Muzea v Praze. Ser. Nat. 1983, 152, 120. [Google Scholar]
- Protopopova, V.V.; Shevera, M.V.; Orlov, O.O.; Panchenko, S.M. The transformer species of the Ukrainian Polissya. Biodiv. Res. Conserv. 2015, 39, 7. [Google Scholar] [CrossRef]
- Lavoie, C. The impact of invasive knotweed species (Reynoutria spp.) on the environment: Review and research perspectives. Biol. Invasions 2017, 19, 2319–2337. [Google Scholar] [CrossRef]
- CABI. Reynoutria japonica Datasheet. In Invasive Species Compendium; CAB International: Wallingford, UK; Available online: https://www.cabi.org/isc/datasheet/23875 (accessed on 30 March 2022).
- CABI. Reynoutria sachalinensis Datasheet. In Invasive Species Compendium; CAB International: Wallingford, UK; Available online: https://www.cabi.org/isc/datasheet/107744 (accessed on 30 March 2022).
- CABI. Reynoutria ×bohemica Datasheet. In Invasive Species Compendium; CAB International: Wallingford, UK; Available online: https://www.cabi.org/isc/datasheet/108332 (accessed on 30 March 2022).
- Papamatthaiakis, N.; Laine, A.; Haapala, A.; Ikonen, R.; Kuittinen, S.; Pappinen, A.; Kolström, M.; Mola-Yudego, B. New energy crop alternatives for Northern Europe: Yield, chemical and physical properties of Giant knotweed (Fallopia sachalinensis var.‘Igniscum’) and Virginia mallow (Sida hermaphrodita). Fuel 2021, 304, 121349. [Google Scholar] [CrossRef]
- Pude, R.; Franken, H. Reynoutria xbohemica—eine Alternative zu Miscanthus xgiganteus? Bodenkultur 2001, 52, 19–27. (In German) [Google Scholar]
- Havrland, B.; Ivanova, T.; Lapczynska-Kordon, B.; Kolarikova, M. Comparative analysis of bio-raw materials and biofuels. In Proceedings of the 12th International Scientific Conference on Engineering for Rural Development, Jelgava, Latvia, 23–24 May 2013; pp. 541–544. [Google Scholar]
- Flora of North America. Fact Sheets: Solidago canadensis. Available online: http://floranorthamerica.org/Solidago_canadensis (accessed on 30 March 2022).
- Flora of North America. Fact Sheets: Solidago gigantea. Available online: http://floranorthamerica.org/Solidago_gigantea (accessed on 30 March 2022).
- Priede, A. Invasive Non-Native Solidago species in Latvia: Expansion History and Current Distribution. Proc. Latv. Acad. Sci. B Nat. Exact Appl. Sci. 2008, 629, 78–83. [Google Scholar] [CrossRef]
- CABI. Solidago canadensis Datasheets. In Invasive Species Compendium; CAB International: Wallingford, UK; Available online: https://www.cabi.org/isc/datasheet/50599 (accessed on 30 March 2022).
- CABI. Solidago gigantea Datasheets. In Invasive Species Compendium; CAB International: Wallingford, UK; Available online: https://www.cabi.org/isc/datasheet/50575 (accessed on 30 March 2022).
- Weber, E. Current and potential ranges of three exotic goldenrods (Solidago) in Europe. Conserv. Biol. 2001, 15, 122–128. [Google Scholar] [CrossRef]
- Kabuce, N.; Priede, N. Solidago canadensis—Invasive Alien Species Fact Sheet. Online Database of the European Network on Invasive Alien Species—NOBANIS. Available online: www.nobanis.org (accessed on 29 March 2022).
- Botta-Dukát, Z. Morphological plasticity in the rhizome system of Solidago gigantea (Asteraceae): Comparison of populations in a wet and a dry habitat. Acta Bot. Hung. 2016, 58, 227–240. [Google Scholar] [CrossRef]
- Rajdus, T.; Svehlakova, H.; Plohak, P.; Stalmachova, B. Management of invasive species Solidago canadensis in Ostrava region (Czech Republic). IOP Conf. Ser. Earth Environ. Sci. 2020, 1, 012046. [Google Scholar] [CrossRef]
- Van Meerbeek, K.; Appels, L.; Dewil, R.; Calmeyn, A.; Lemmens, P.; Muys, B.; Hermy, M. Biomass of invasive plant species as a potential feedstock for bioenergy production. Biofuels Bioprod. Biorefining 2015, 9, 273–282. [Google Scholar] [CrossRef]
- Ciesielczuk, T.; Poluszyńska, J.; Rosik-Dulewska, C.; Sporek, M.; Lenkiewicz, M. Uses of weeds as an economical alternative to processed wood biomass and fossil fuels. Ecol. Eng. 2016, 95, 485–491. [Google Scholar] [CrossRef]
- Van Meerbeek, K.; Muys, B.; Hermy, M. Lignocellulosic biomass for bioenergy beyond intensive cropland and forests. Renew. Sust. Energ. Rev. 2019, 102, 139–149. [Google Scholar] [CrossRef]
- Flora of North America. Fact Sheets: Spiraea tomentosa. Available online: http://floranorthamerica.org/Spiraea_tomentosa (accessed on 30 March 2022).
- Symes, M. Charles Hamilton’s plantings at Painshill. Gard. Hist. 1983, 11, 112–124. [Google Scholar] [CrossRef]
- Dajdok, Z.; Nowak, A.; Danielewicz, W.; Kujawa-Pawlaczyk, J.; Bena, W. Spiraea tomentosa—Invasive Alien Species Fact Sheet. Online Database of the European Network on Invasive Alien Species—NOBANIS. 2011. Available online: www.nobanis.org (accessed on 19 April 2022).
- Wiatrowska, B.; Łukowski, A.; Karolewski, P.; Danielewicz, W. Invasive Spiraea tomentosa: A new host for monophagous Earias clorana? Arthropod-Plant Interact. 2018, 12, 423–434. [Google Scholar] [CrossRef]
- Wiatrowska, B.; Pietras, M.; Kolanowska, M.; Danielewicz, W. Current occurrence and potential future climatic niche distribution of the invasive shrub Spiraea tomentosa L. in its native and non-native ranges. Glob. Ecol. Conserv. 2020, 24, e01226. [Google Scholar] [CrossRef]
- Wiatrowska, B.; Danielewicz, W. Environmental conditions of steeplebush (Spiraea tomentosa L.) invasion in the Lower Silesian Forests. Sylwan 2016, 60, 696–704. (In Polish) [Google Scholar]
- Wiatrowska, B.; Czepieruk, I. Unpublished work.
- Abrahamson, W.G.; McCrea, K.D. Seasonal nutrient dynamics of Solidago altissima (Compositae). J. Torrey Bot. Soc. 1985, 112, 414–420. [Google Scholar] [CrossRef]
- TAPPI. T 204 cm-07—Solvent Extractives of Wood and Pulp; Standards Technical Association of the Pulp and Paper Industry: Atlanta, GA, USA, 2007. [Google Scholar]
- Seifert, K. Zur Frage der Cellulose-Schnellbestimmung nach der Acetylaceton-Methode. Das Pap 1960, 14, 104–106. (In German) [Google Scholar]
- TAPPI. T-222 om-06—Lignin in Wood and Pulp; Standards Technical Association of the Pulp and Paper Industry: Atlanta, GA, USA, 2006. [Google Scholar]
- DIN. DIN 51731. Testing of Solid Fuels—Compressed Untreated Wood—Requirements and Testing; Deutsches Institut für Normung, E.V. (German National Standard): Berlin, Germany, 1996. [Google Scholar]
- Kawa-Rygielska, J.; Pietrzak, W. Bread waste management for the production of bioethanol. Żywność Nauka Technologia Jakość 2011, 79, 105–118. (In Polish) [Google Scholar]
- Kim, H.Y. Analysis of variance (ANOVA) comparing means of more than two groups. Restor. Dent. Endod. 2014, 39, 74–77. [Google Scholar] [CrossRef]
- Qian, X.; Lee, S.W. The design and analysis of energy efficient building envelopes for the commercial buildings by mixed-level factorial design and statistical methods. In Proceedings of the ASEE Middle Atlantic American Society of Engineering Education, Swarthmore, PA, USA, 14–15 November 2014; pp. 14–15. [Google Scholar]
- Rowell, R.M.; Han, J.S.; Rowell, J.S. Characterization and Factors Effecting Fiber Properties. In Natural Polymers and Agrofibers Composites; Frollini, E., Leao, A.L., Mattoso, L.H.C., Eds.; Natural Polymers and Agrofibers Composites, Embrapa: Sãn Carlos, Brasil, 2000; pp. 115–134. [Google Scholar]
- Xu, P.; Cheng, S.; Han, Y.; Zhao, D.; Li, H.; Wang, Y.; Zhang, G.; Chen, C. Natural Variation of Lignocellulosic Components in Miscanthus Biomass in China. Front. Chem. 2020, 8, 595143. [Google Scholar] [CrossRef]
- Brosse, N.; Dufour, A.; Meng, X.; Sun, Q.; Ragauskas, A. Miscanthus: A fast-growing crop for biofuels and chemicals production. Biofuels Bioprod. Bioref. 2012, 6, 580–598. [Google Scholar]
- Waliszewska, B.; Grzelak, M.; Gaweł, E.; Spek-Dźwigała, A.; Sieradzka, A.; Czekała, W. Chemical Characteristics of Selected Grass Species from Polish Meadows and Their Potential Utilization for Energy Generation Purposes. Energies 2021, 14, 1669. [Google Scholar] [CrossRef]
- Gismatulina, Y.A.; Budaeva, V.V. Chemical composition of five Miscanthus sinensis harvests and nitric-acid cellulose therefrom. Ind. Crops Prod. 2017, 109, 227–232. [Google Scholar] [CrossRef]
- Qian, X.; Xue, J.; Yang, Y.; Lee, S.W. Thermal properties and combustion-related problems prediction of agricultural crop residues. Energies 2021, 14, 4619. [Google Scholar] [CrossRef]
- Krzyżaniak, M.; Stolarski, M.J.; Waliszewska, B.; Szczukowski, S.; Tworkowski, J.; Załuski, D.; Śnieg, M. Willow biomass as feedstock for an integrated multi-product biorefinery. Ind. Crops Prod. 2014, 58, 230–237. [Google Scholar] [CrossRef]
- Collura, S.; Azambre, B.; Weber, J.V. Thermal behaviour of Miscanthus grasses, an alternative biological fuel. Env. Chem. Lett. 2005, 3, 95–99. [Google Scholar] [CrossRef]
- Friedl, A.; Padouvas, E.; Rotter, H.; Varmuza, K. Prediction of heating values of biomass fuel from elemental composition. Anal. Chim. Acta 2005, 544, 191–198. [Google Scholar] [CrossRef]
- Michel, R.; Mischler, N.; Azambre, B.; Finqueneisel, G.; Machnikowski, J.; Rutkowski, P.; Zimny, T.; Weber, J.V. Miscanthus ×giganteus straw and pellets as sustainable fuels and raw material for activated carbon. Environ. Chem. Lett. 2006, 4, 185–189. [Google Scholar] [CrossRef]
- Hodgson, E.M.; Fahmi, R.; Yates, N.; Barraclough, T.; Shield, I.; Allison, G.; Bridgwater, A.V.; Donnison, I.S. Miscanthus as a feedstock for fast-pyrolysis: Does agronomic treatment affect quality? Bioresour. Technol. 2010, 101, 6185–6191. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, A.; Munazza, J.; Asma, S.; Fariha, I.; Waheed, T.; Syed, H.G. Optimizing the Effect of Chemical Pretreatment on Lignocellulosic Properties of Wheat Straw. J. Waste Biomass Manag. 2020, 2, 28–32. [Google Scholar]
- Wawro, A.; Batog, J.; Gieparda, W. Polish Varieties of Industrial Hemp and Their Utilisation in the Efficient Production of Lignocellulosic Ethanol. Molecules 2021, 26, 6467. [Google Scholar] [CrossRef]
- Suttikul, S.; Srinorakutara, T.; Butivate, E.; Orasoon, K. Comparison of SHF and SSF Processes for Ethanol Production from Alkali-Acid Pretreated Sugarcane trash. KKU. Res. J. 2016, 21, 229–235. [Google Scholar]
- Yuwa-Amornpitak, T.; Yeunyaw, P. Comparative Study of Ethanol Production from Cassava Pulp by a Mixed Culture of Amylomyces Rouxii with Zygosaccharomyces Pseudorouxii and Zymomonas Mobilis. Biomed. Pharmacol. J. 2019, 12, 199. [Google Scholar] [CrossRef]
- Suwanan, S.; Saovanee, C.; Taweesak, R.; Chayanoot, S. Bioethanol production from oil palm empty fruit bunch with SSF and SHF processes using Kluyveromyces marxianus yeast. Cellulose 2020, 27, 301–314. [Google Scholar]
- Cotana, F.; Cavalaglio, G.; Gelosia, M.; Coccia, V.; Petrozzi, A.; Ingles, D.; Pompili, E. A comparison between SHF and SSSF processes from cardoon for ethanol production. Ind. Crops Prod. 2015, 69, 424–432. [Google Scholar] [CrossRef]
- López-Sandin, I.; Zavala-García, F.; Levin, L.; Ruiz, H.A.; Hernández-Luna, C.E.; Gutiérrez-Soto, G. Evaluation of Bioethanol Production from Sweet Sorghum Variety Roger under Different Tillage and Fertilizer Treatments. Bioenerg. Res. 2021, 14, 1058–1069. [Google Scholar] [CrossRef]
- Li, H.Q.; Li, C.L.; Sang, T.; Xu, J. Pretreatment on Miscanthus lutarioriparious by liquid hot water for efficient ethanol production. Biotechnol. Biofuels 2013, 6, 76. [Google Scholar] [CrossRef] [PubMed]
- Roozeboom, K.L.; Wang, D.; McGowan, A.R.; Propheter, J.L.; Staggenborg, S.A.; Rice, C.W. Long-term biomass and potential ethanol yields of annual and perennial biofuel crops. Agron. J. 2018, 110, 74–83. [Google Scholar] [CrossRef]



| Plant Species | Extractive Substances | Ash | Pentosans | Cellulose | Hemicellulose | Lignin |
|---|---|---|---|---|---|---|
| Reynoutria japonica | 16.14 ± 0.33 | 9.49 ± 0.01 | 15.98 ± 0.15 | 31.94 ± 0.77 | 20.87 ± 0.64 | 20.18 ± 0.17 |
| Reynoutria sachalinensis | 21.16 ± 0.55 | 5.77 ± 0.01 | 20.09 ± 0.52 | 29.57 ± 0.49 | 29.80 ± 0.72 | 19.17 ± 0.11 |
| Reynoutria × bohemica | 19.72 ± 0.21 | 6.58 ± 0.09 | 20.51 ± 0.42 | 31.71 ± 0.95 | 34.48 ± 0.57 | 19.41 ± 0.40 |
| Solidago canadensis | 14.42 ± 0.52 | 2.39 ± 0.01 | 20.07 ± 0.46 | 38.95 ± 0.67 | 23.78 ± 1.17 | 28.68 ± 0.13 |
| Solidago gigantea | 13.27 ± 0.26 | 4.93 ± 0.01 | 18.54 ± 0.3 | 38.50 ± 0.18 | 29.59 ± 0.21 | 24.79 ± 0.22 |
| Spiraea tomentosa | 14.19 ± 0.32 | 6.95 ± 0.01 | 16.70 ± 0.16 | 32.32 ± 0.09 | 22.46 ± 0.82 | 23.63 ± 0.12 |
| Plant Species | Higher Heating Value [MJ∙kg−1] | Lower Heating Value [MJ∙kg−1] |
|---|---|---|
| Reynoutria japonica | 18.485 | 16.965 |
| Reynoutria sachalinensis | 19.927 | 18.405 |
| Reynoutria × bohemica | 19.210 | 17.683 |
| Solidago canadensis | 19.894 | 18.369 |
| Solidago gigantea | 19.403 | 17.868 |
| Spiraea tomentosa | 18.892 | 17.353 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiatrowska, B.M.; Wawro, A.; Gieparda, W.; Waliszewska, B. Bioethanol Production Potential and Other Biomass Energy Properties of Invasive Reynoutria, Solidago, and Spiraea Plants. Forests 2022, 13, 1582. https://doi.org/10.3390/f13101582
Wiatrowska BM, Wawro A, Gieparda W, Waliszewska B. Bioethanol Production Potential and Other Biomass Energy Properties of Invasive Reynoutria, Solidago, and Spiraea Plants. Forests. 2022; 13(10):1582. https://doi.org/10.3390/f13101582
Chicago/Turabian StyleWiatrowska, Blanka Maria, Aleksandra Wawro, Weronika Gieparda, and Bogusława Waliszewska. 2022. "Bioethanol Production Potential and Other Biomass Energy Properties of Invasive Reynoutria, Solidago, and Spiraea Plants" Forests 13, no. 10: 1582. https://doi.org/10.3390/f13101582
APA StyleWiatrowska, B. M., Wawro, A., Gieparda, W., & Waliszewska, B. (2022). Bioethanol Production Potential and Other Biomass Energy Properties of Invasive Reynoutria, Solidago, and Spiraea Plants. Forests, 13(10), 1582. https://doi.org/10.3390/f13101582