
Is the Survivability of Silver Fir under Condition of Strong Ungulate Pressure Related to Mycobiota of Bark-Stripping Wounds?


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
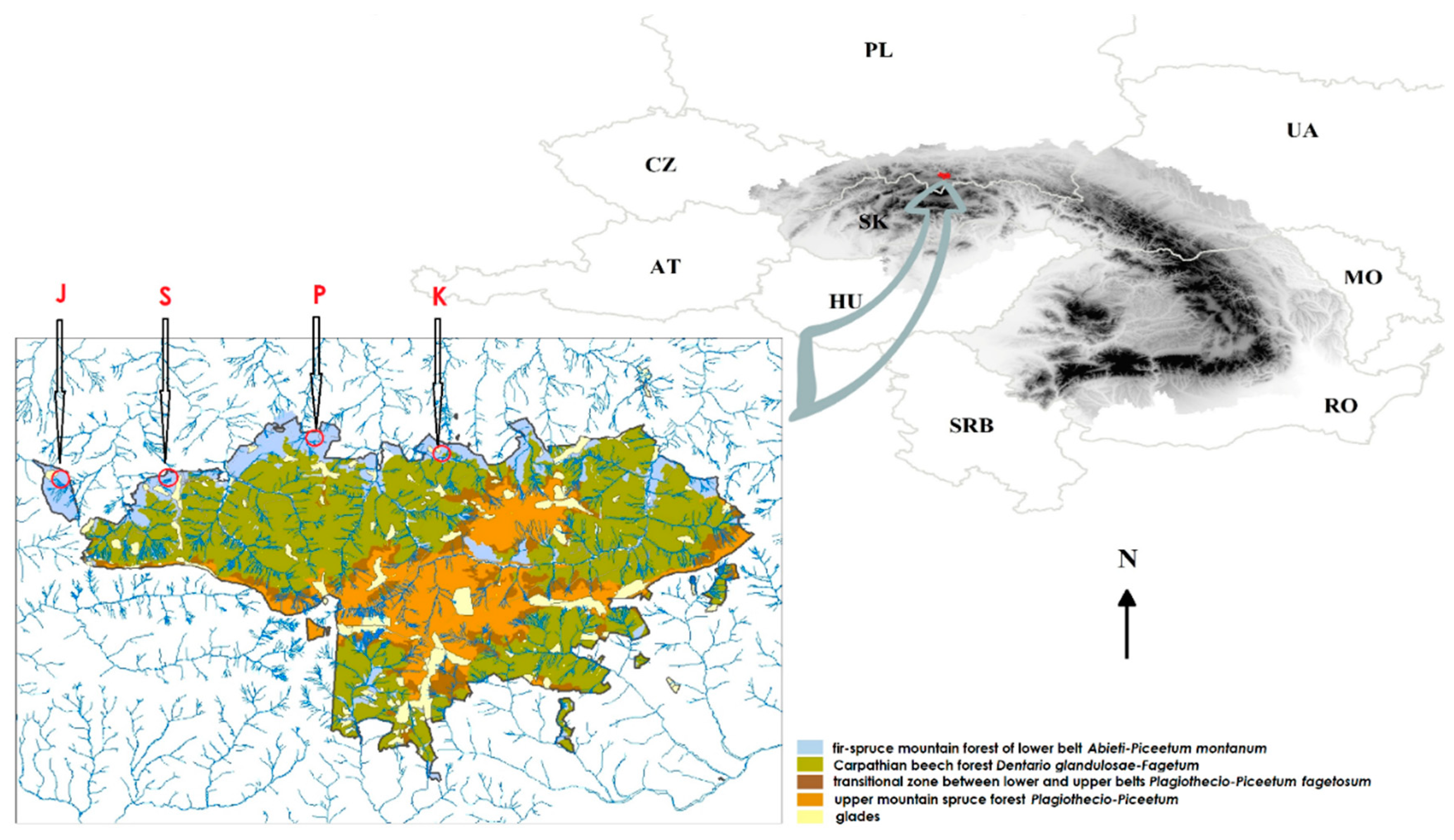
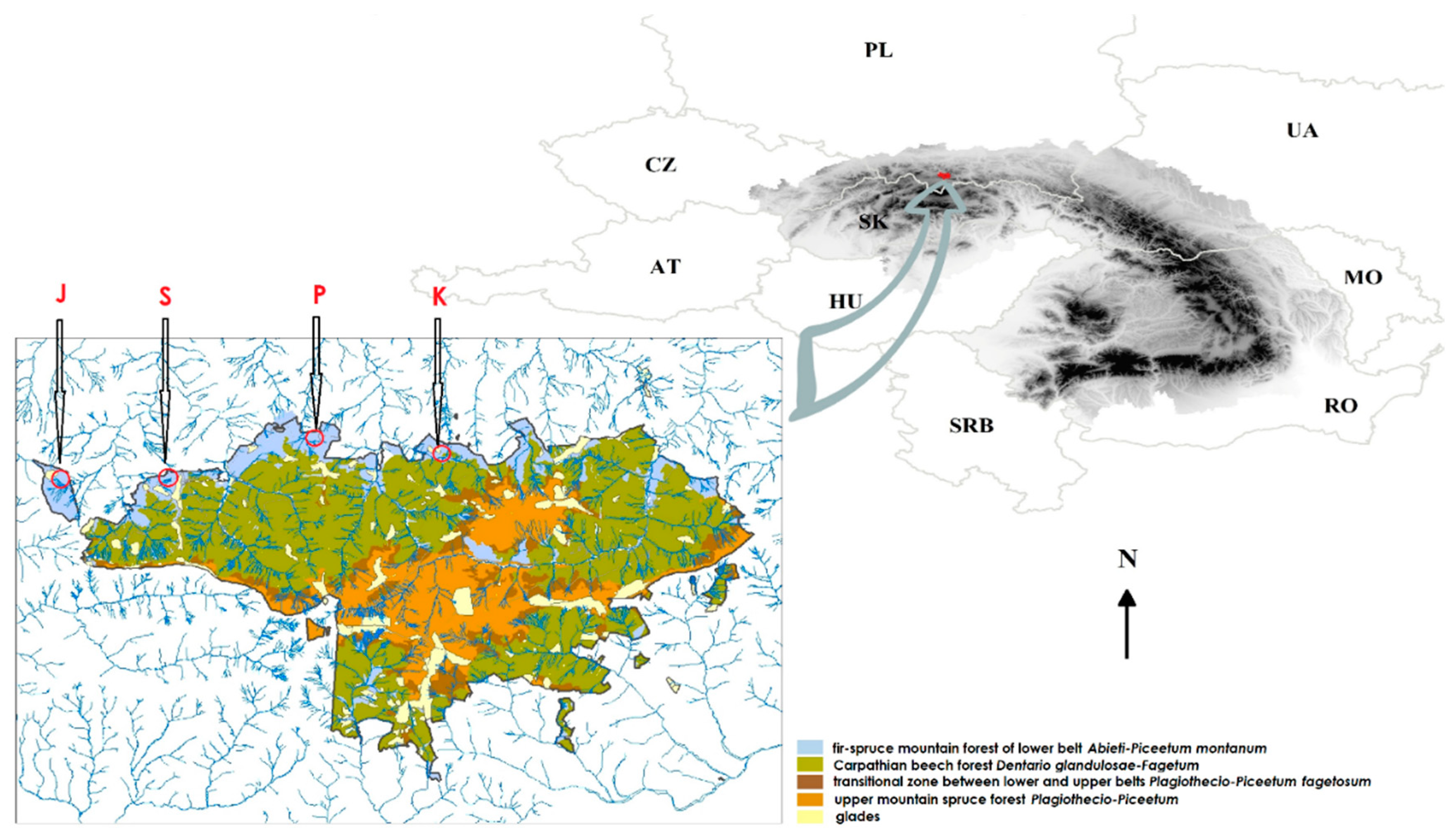
2.1. Fieldwork
2.2. Laboratory Analysis
2.3. Statistical Analysis
3. Results
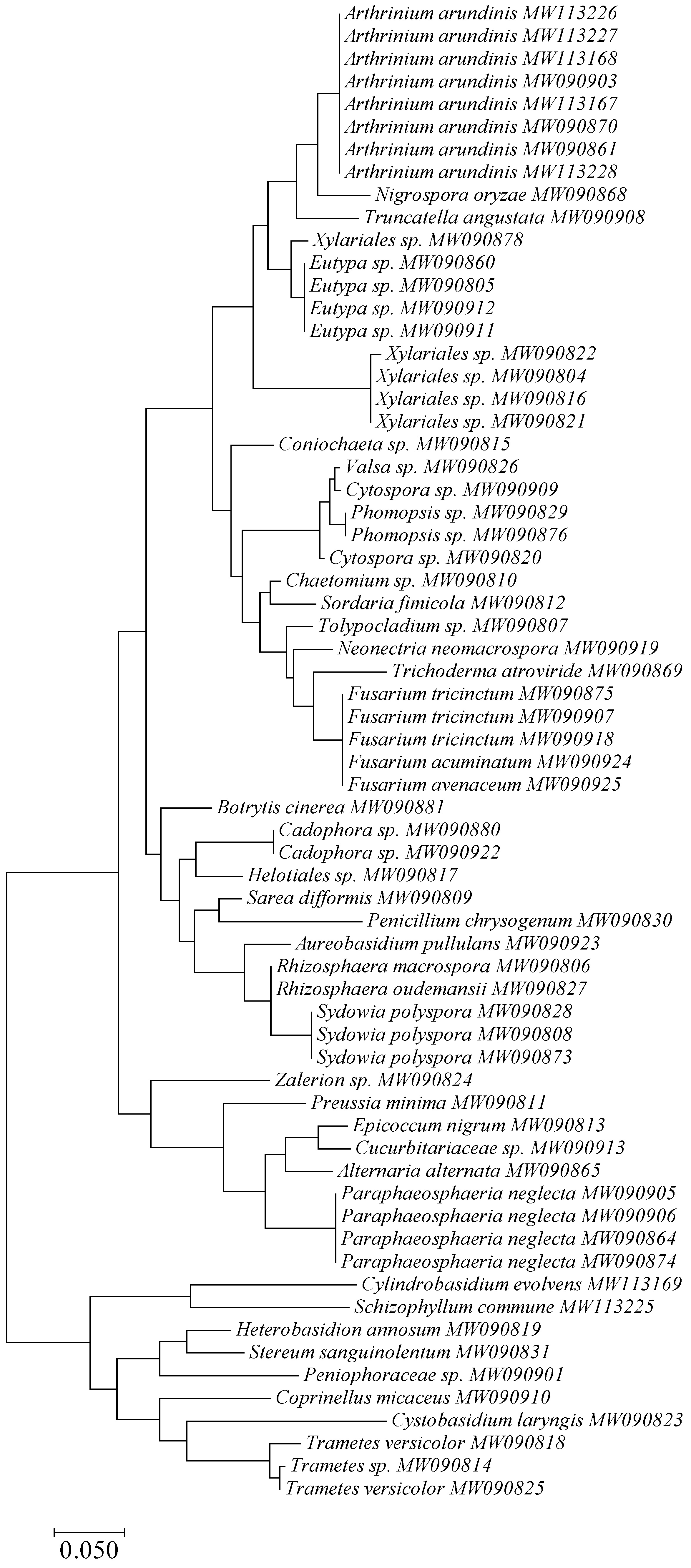
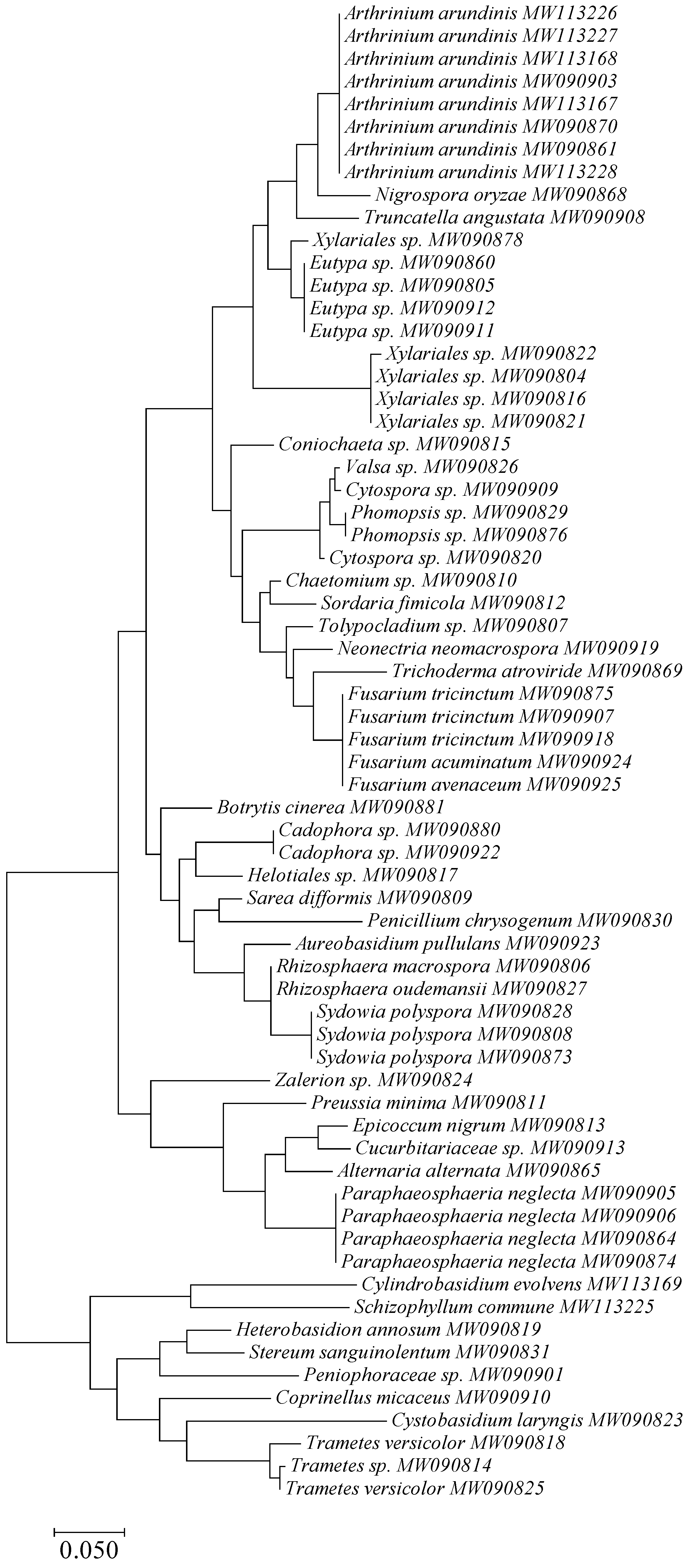
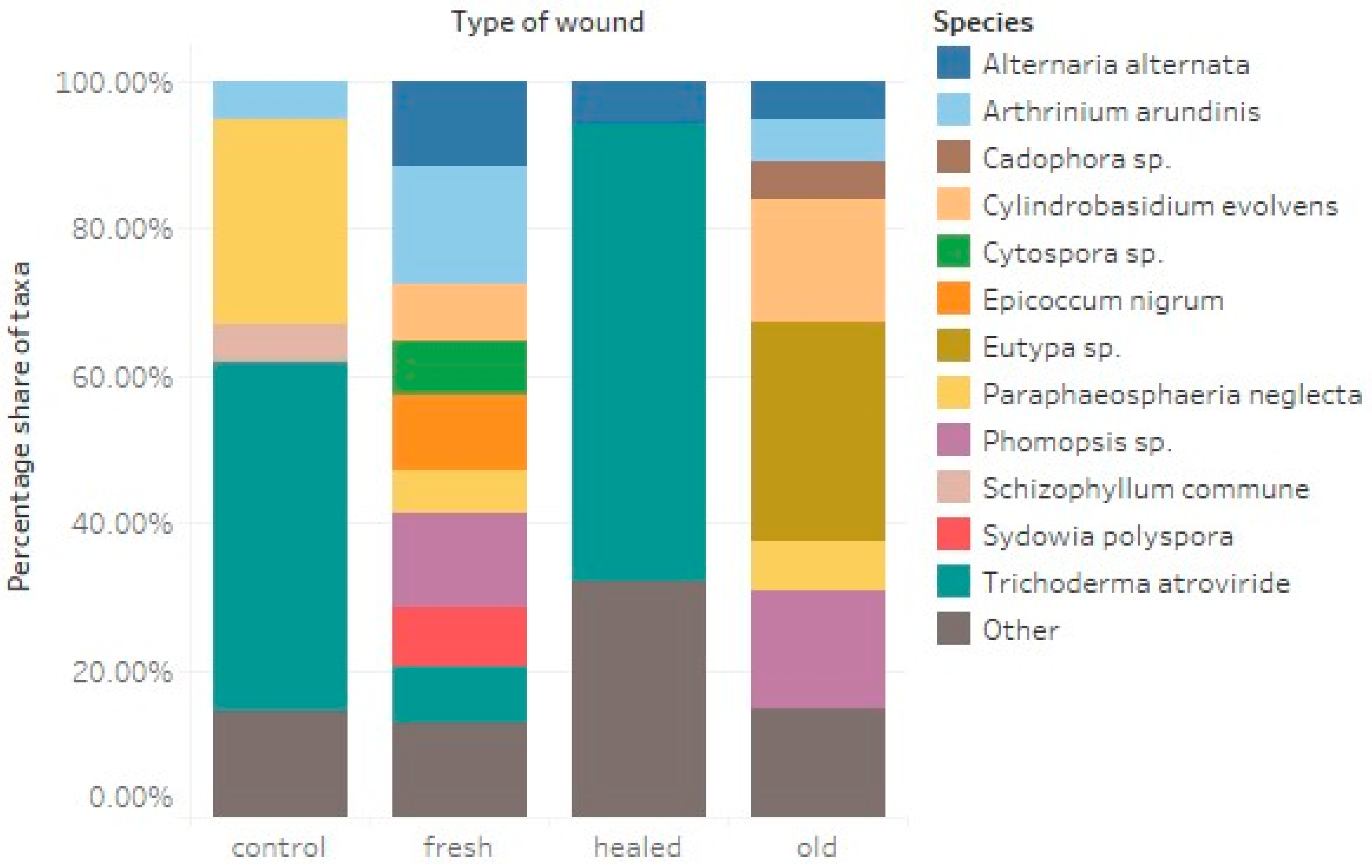
3.1. Taxonomy and Frequency of Fungal Colonies
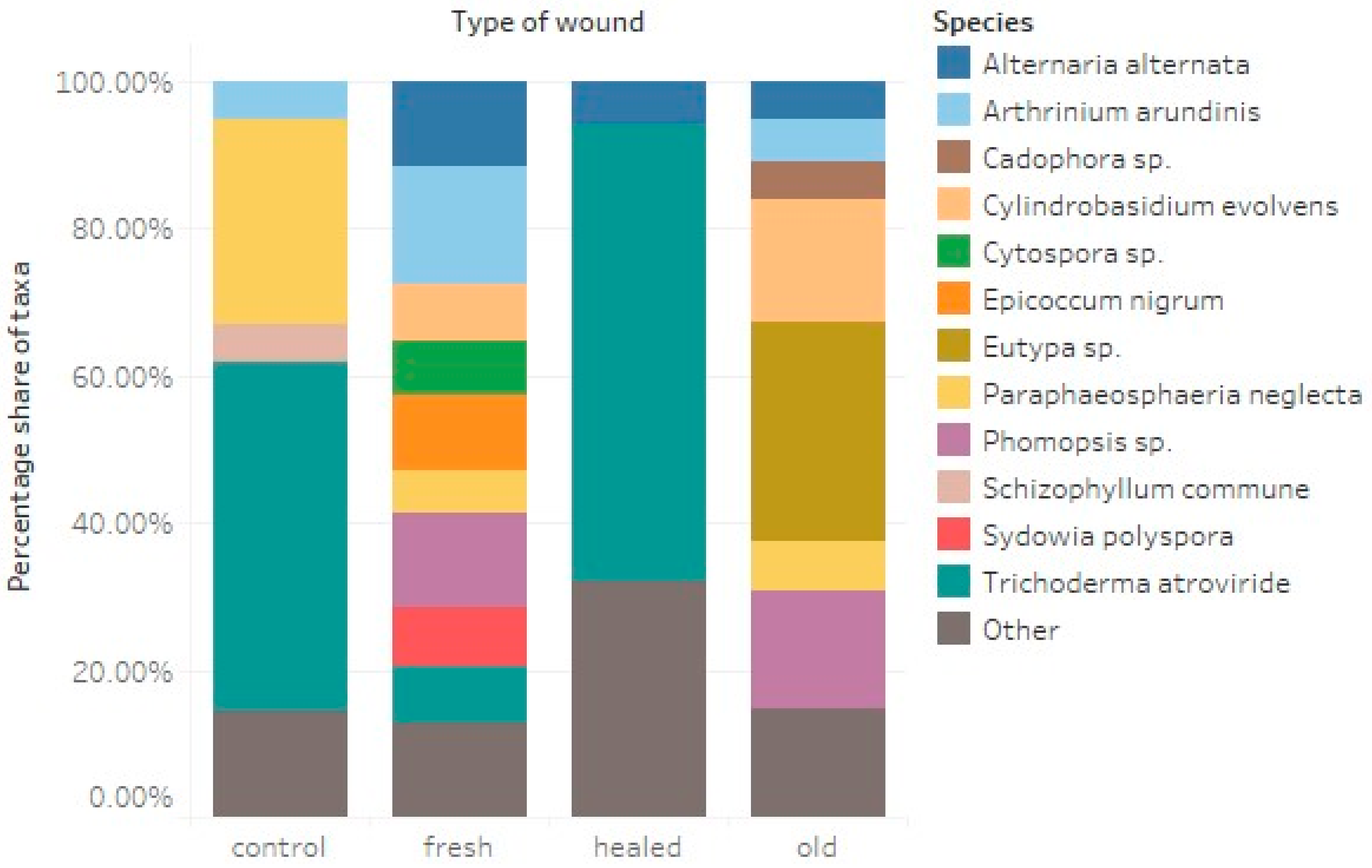
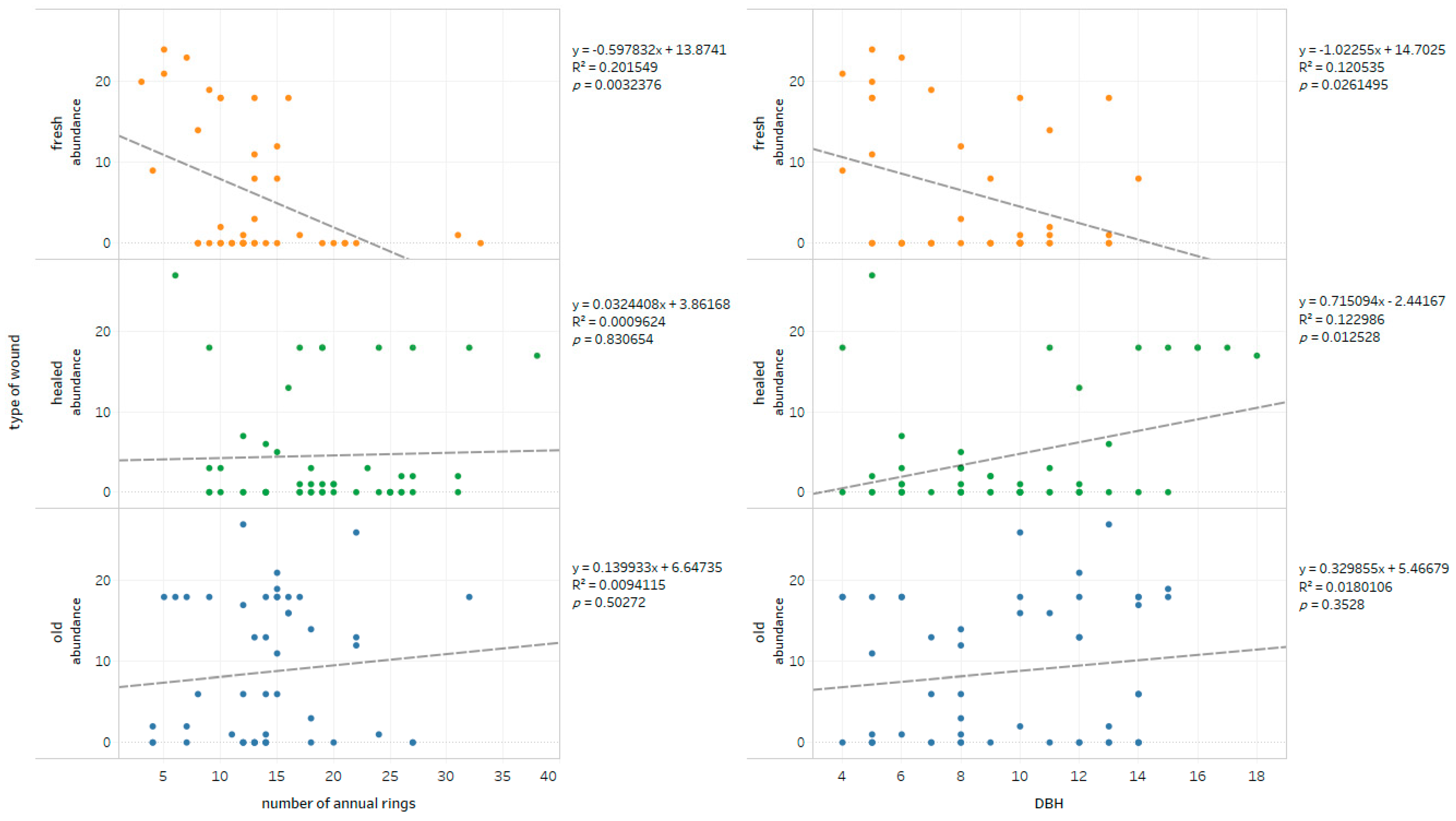
3.2. Fungi-Wounds Relationship
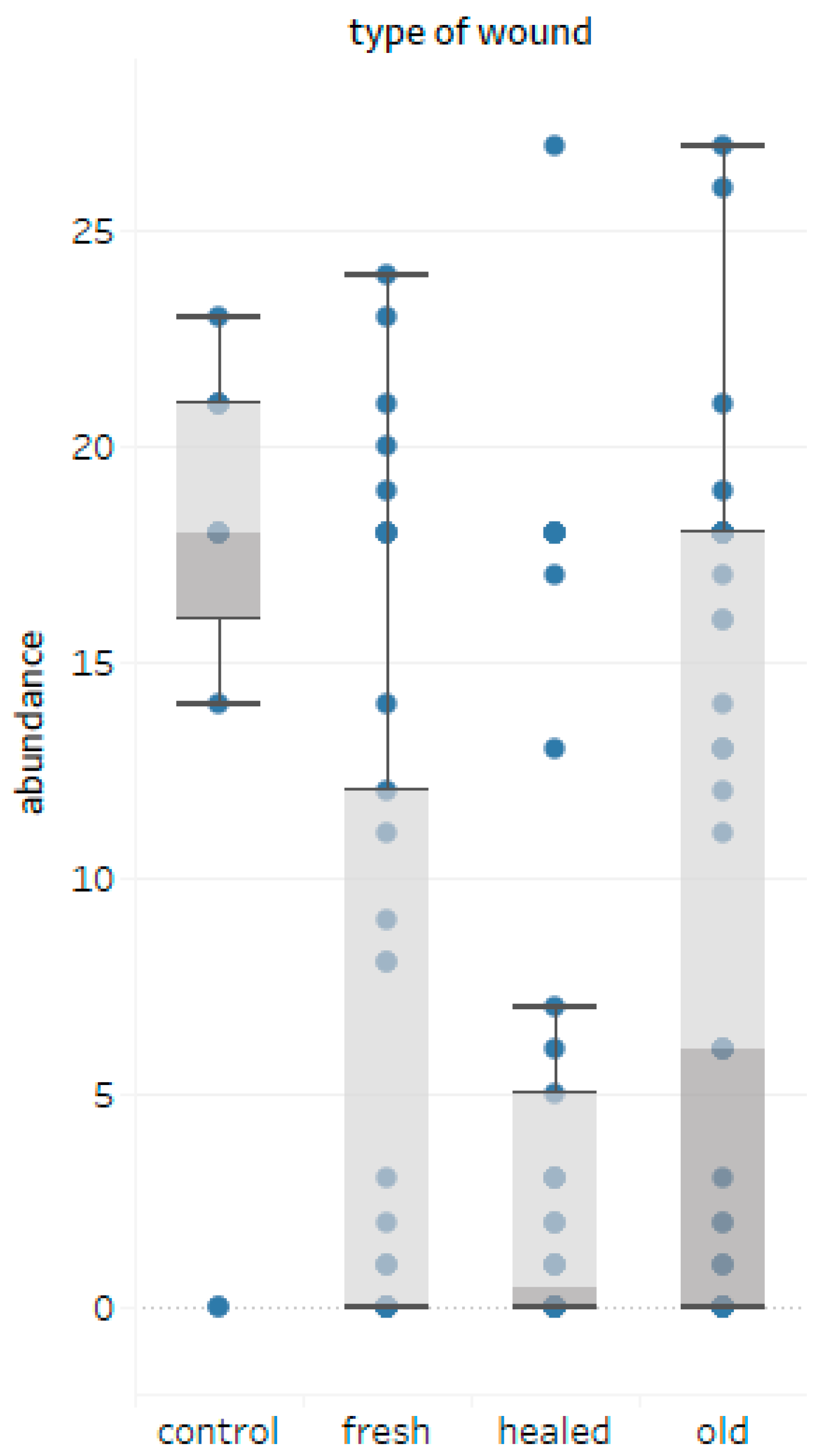
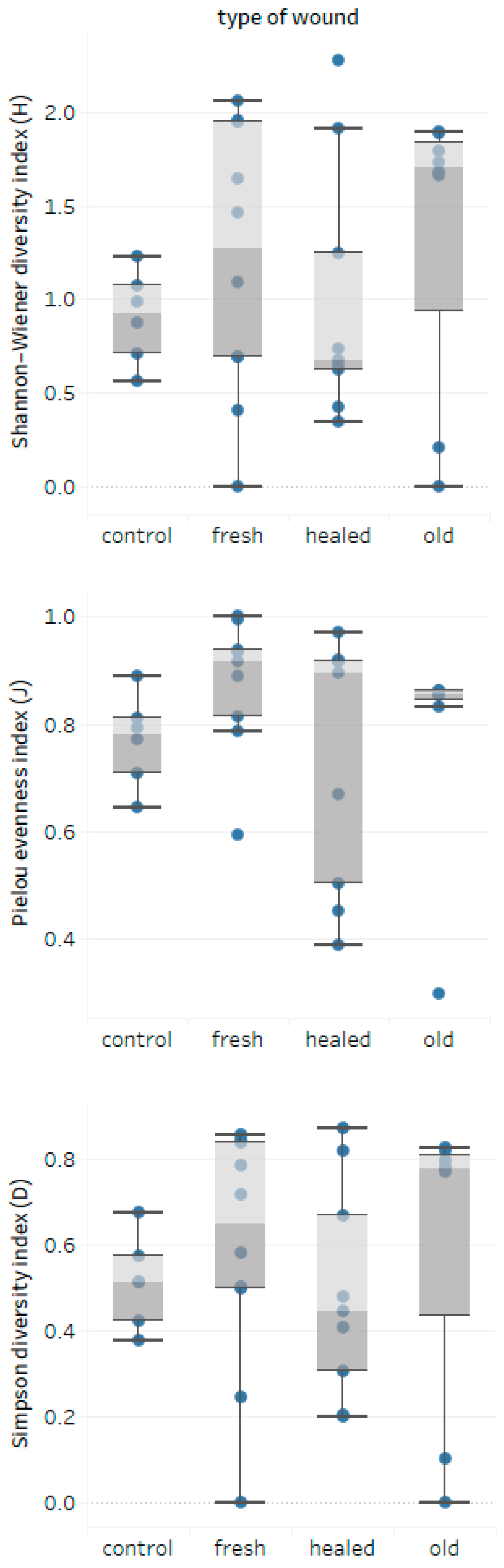
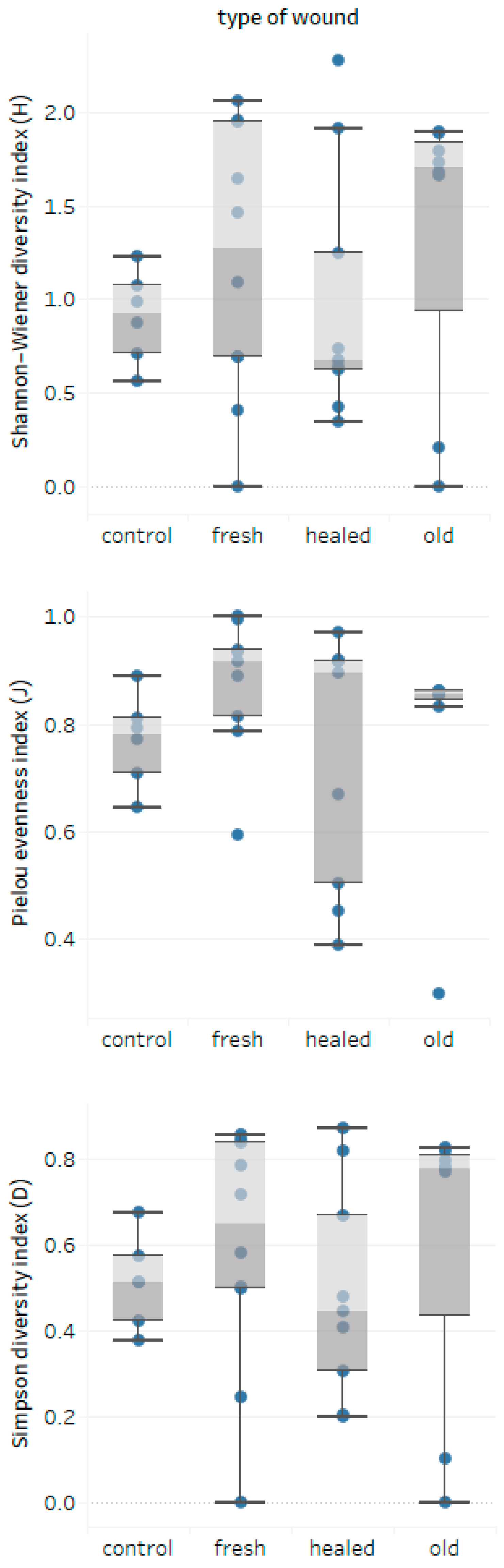
3.3. Wounds-Fungal Diversity Relationship
4. Discussion
5. Conclusions
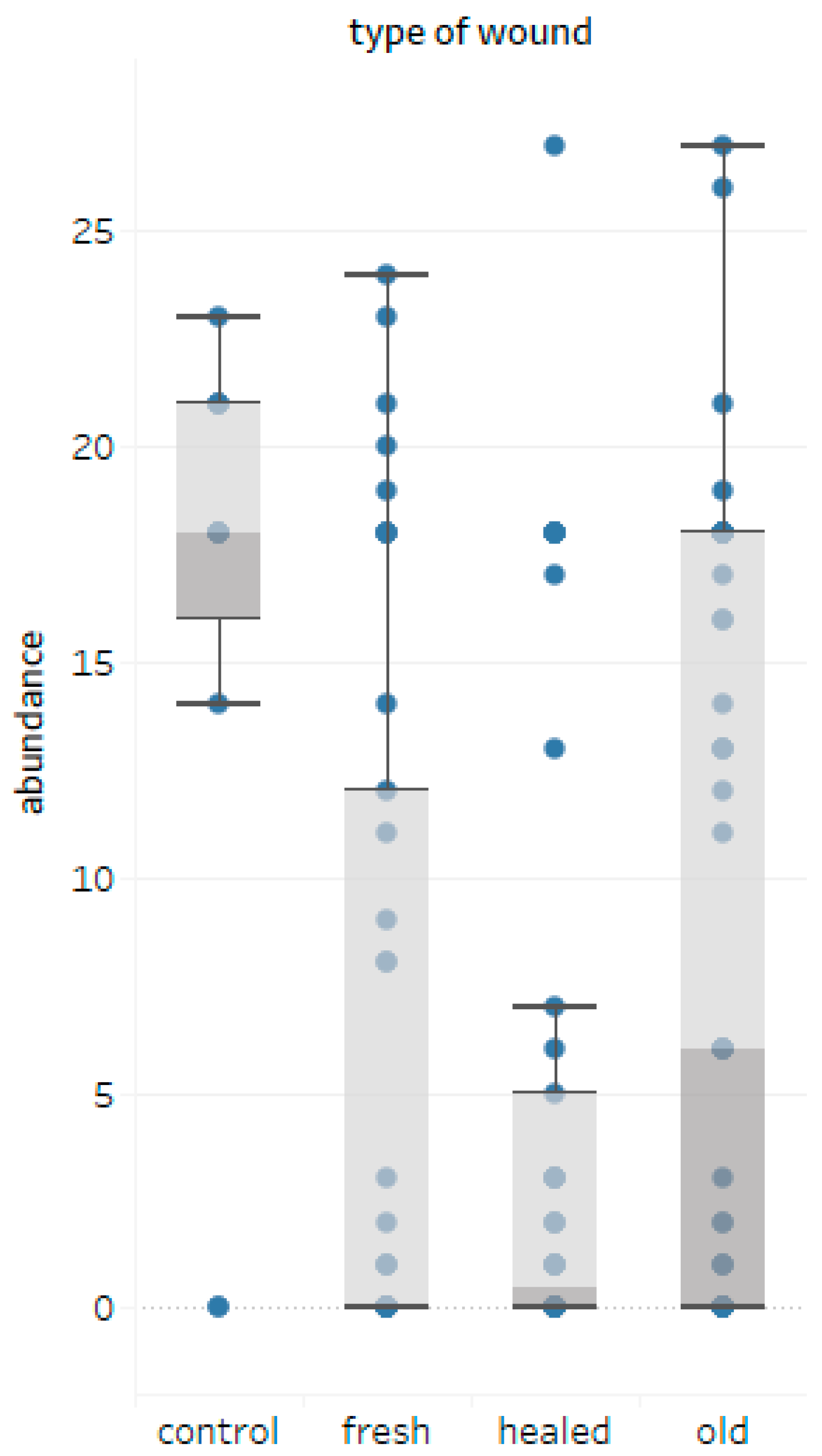
- The greatest number of fungal colonies inhabiting the silver fir wood was obtained from old open wounds, with a smaller number of taxa compared to fresh and healed wounds.
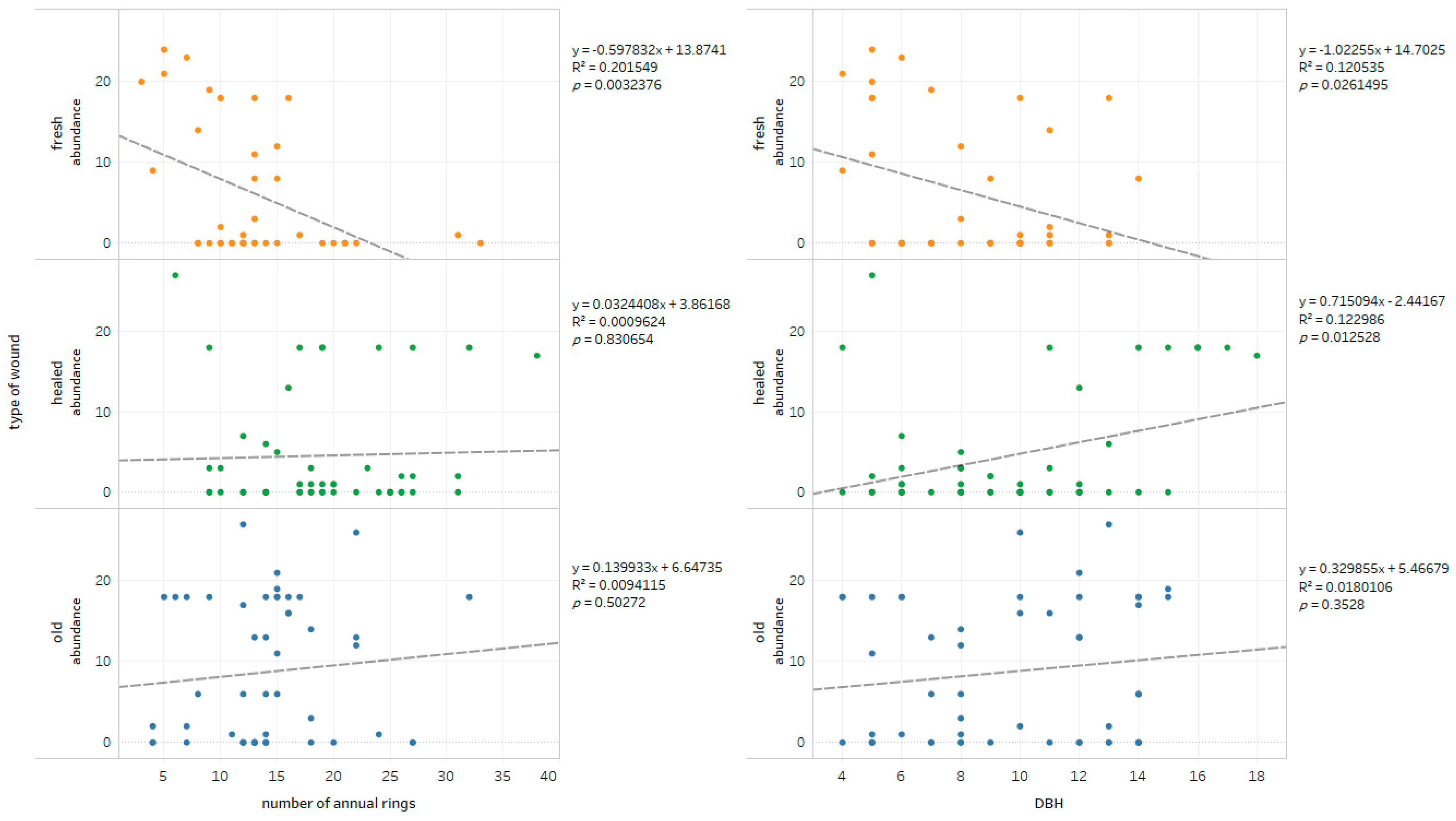
- The age and DBH of bark-stripped trees do not seem to be of importance for the degree of infection of wounds expressed by the number of colonies; nevertheless, species diversity of wood decay fungi assessed by the α-diversity Shannon-Wiener (H) and Simpson (D) indices seems to be lower in healed wounds.
- The strong presence of Arthrinium arundinis and Trichoderma atroviride in firs wounds, which are often antagonistic to pathogenic fungi, may indicate the natural defense mechanisms of fir, aimed at inhibiting disease processes and, subsequently, the decomposition of wood.
- The high survival rate of the young generation of fir indicates that despite the strong pressure of red deer, the fir regeneration mechanism based on inter-species competition of fungi may be an effective tool in the fight for the survival of this species in the Carpathians.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics
References
- Felton, A.M.; Wam, H.K.; Stolter, C.; Mathisen, K.M.; Wallgren, M. The complexity of interacting nutritional drivers behind food selection, a review of northern cervids. Ecosphere 2018, 9, 1–25. [Google Scholar] [CrossRef]
- Gill, R.M.A. A review of damage by mammals in north temperate forests 1. Deer. Forestry Int. J. For. Res. 1992, 65, 145–169. [Google Scholar] [CrossRef] [Green Version]
- Ando, M.; Yokota, H.; Shibata, E. Why do sika deer, Cervus Nippon, debark trees in summer on Mt. Ohdaigahara, central Japan? Mammal Study 2004, 29, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Côté, S.D.; Beguin, J.; de Bellefeuille, S.; Champagne, E.; Thiffault, N.; Tremblay, J.-P. Structuring Effects of Deer in Boreal Forest Ecosystems. Adv. Ecol. 2014, 2014, 917834. [Google Scholar] [CrossRef] [Green Version]
- Cukor, J.; Vacek, Z.; Linda, R.; Vacek, S.; Marada, P.; Šimůnek, V.; Havránek, F. Effects of Bark Stripping on Timber Production and Structure of Norway Spruce Forests in Relation to Climatic Factors. Forests 2019, 10, 320. [Google Scholar] [CrossRef] [Green Version]
- Domański, S. Próba fitopatologicznej oceny świerków ospałowanych przez zwierzynę w Karkonoszach. Folia For. Pol. Ser. A For. 1963, 12, 157–174. [Google Scholar]
- Vasiliauskas, R.; Stenlid, J.; Johansson, M. Fungi in bark peeling wounds of Picea abies in central Sweden. Eur. J. Forest Pathol. 1996, 26, 285–296. [Google Scholar] [CrossRef]
- Kwaśna, H.; Mazur, A.; Łabędzki, A.; Kuźminski, R.; Łakomy, P. Zbiorowiska grzybów w rozkładającym się drewnie dębu i sosny [Communities of fungi in decomposed wood of oak and pine]. Leśne Prace Badawcze (For. Res. Pap.) 2016, 77, 261–275. [Google Scholar]
- Arhipova, N.; Jansons, A.; Zaluma, A.; Gaitnieks, T.; Vasaitis, R. Bark stripping of Pinus contorta caused by moose and deer: Wounding patterns, discoloration of wood, and associated fungi. Can. J. For. Res. 2015, 45, 1434–1438. [Google Scholar] [CrossRef]
- Pach, M. Zasięg i dynamika rozprzestrzeniania się zgnilizny wewnątrz pni jodeł w wyniku ich spałowania przez jeleniowate [Extent and dynamics of wood decay spreading inward fir stems as a result of peeling by ungulates]. Sylwan 2005, 149, 23–35. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment; UN Food and Agriculture Organization: Rome, Italy, 2015. [Google Scholar]
- Jagodziński, A.; Dyderski, M.; Gęsikiewicz, K.; Horodecki, P. Tree and stand level estimations of Abies alba Mill. aboveground biomass. Ann. For. Sci. 2019, 76, 56. [Google Scholar] [CrossRef] [Green Version]
- Niemtur, S.; Chomicz, E.; Kapsa, M. Występowanie zgnilizny odziomkowej jodły pospolitej (Abies alba Mill.) w obszarach chronionych [Occurrence of the silver fir (Abies alba Mill.) butt rot in protected areas]. Leśne Prace Badawcze (For. Res. Pap.) 2014, 75, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Chwistek, K. Zmiany składu gatunkowego i struktury drzewostanów Gorczańskiego Parku Narodowego w latach 1992–2007 [Changes of the species composition and structure of stands of the Gorce National Park during the period 1992–2007]. Ochrona Beskidów Zachodnich 2010, 3, 79–92. [Google Scholar]
- Czarnota, P. Presja Jeleniowatych na Drzewostany Gorczańskiego Parku Narodowego—Raport z Monitoringu Uszkodzeń Drzewostanów przez Jeleniowate na Stałych Powierzchniach Badawczych GPN w roku 2017 [Deer Pressure on the Stands of the Gorce National Park—Report on the Monitoring of Damage to Stands by Deer on Permanent Research Plots of the GPN in 2017]; Unpublished Materials in Archive of GNP; Gorce National Park, GNP Library: Poręba Wielka, Poland, 2018. [Google Scholar]
- Barzdajn, W. Strategia restytucji jodły pospolitej (Abies alba Mill.) w Sudetach [A strategy for restitution of silver fir (Abies alba Mill.) in the Sudety Mountains]. Sylwan 2000, 144, 63–77. [Google Scholar]
- Dobrowolska, D. Dynamika luk w drzewostanach mieszanych rezerwatu Jata [Gap dynamics in mixed stands of Jata reserve]. Leśne Prace Badawcze (For. Res. Pap.) 2006, 4, 61–75. [Google Scholar]
- Valente, A.M.; Acevedo, P.; Figueiredo, A.M.; Martins, R.; Fonseca, C.; Torres, R.T.; Delibes-Mateos, M. Dear deer? Maybe for now. People’s perception on red deer (Cervus elaphus) populations in Portugal. Sci. Total Environ. 2020, 748, 141400. [Google Scholar] [CrossRef]
- Ludwig, M.; Grüninger, F.; Rothfuss, E.; Heurich, M. Discourse Analysis as an Instrument to Reveal the Pivotal Role of the Media in Local Acceptance or Rejection of a Wildlife Management Project: A Case Study from the Bavarian Forest National Park. Erdkunde 2012, 66, 143–156. Available online: http://www.jstor.org/stable/41759065 (accessed on 20 January 2021). [CrossRef]
- Miścicki, S.; Żurek, Z. Monitoring uszkodzeń spowodowanych przez jeleniowate w młodych drzewostanach i odnowieniach Gorczańskiego Parku Narodowego [Monitoring of the deer−related damage to the young stands and regeneration in the Gorczański National Park]. Sylwan 2015, 159, 505–515. [Google Scholar] [CrossRef]
- Jaworski, A. Fir regression in Polish mountains areas. For. Pathol. 1982, 12, 143–159. [Google Scholar] [CrossRef]
- Ficko, A.; Poljanec, A.; Boncina, A. Do changes in spatial distribution, structure and abundance of silver fir (Abies alba Mill.) indicate its decline? For. Ecol. Manag. 2011, 261, 844–854. [Google Scholar] [CrossRef]
- Senn, J.; Suter, W. Ungulate browsing on silver fir (Abies alba) in the Swiss Alps: Beliefs in search of supporting data. For. Ecol. Manag. 2003, 181, 151–154. [Google Scholar] [CrossRef]
- Häsler, H.; Senn, J. Ungulate browsing on European silver fir Abies alba: The role of occasions, food shortage and diet preferences. Wildl. Biol. 2012, 18, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Gazda, A.; Kościelniak, P.; Hardy, M.; Muter, E.; Kędra, K.; Bodziarczyk, J.; Frączek, M.; Chwistek, K.; Różański, W.; Szwagrzyk, J. Upward expansion of distribution ranges of tree species: Contrasting results from two national parks in Western Carpathians. Sci. Total Environ. 2018, 653, 920–929. [Google Scholar] [CrossRef]
- Dyderski, M.; Pawlik, Ł. Spatial distribution of tree species in mountain national parks depends on geomorphology and climate. For. Ecol. Manag. 2020, 474, 118366. [Google Scholar] [CrossRef]
- Vencurik, J.; Kucbel, S.; Saniga, M.; Jaloviar, P.; Sedmáková, D.; Pittner, J.; Parobeková, Z.; Bosela, M. Growth dynamics of the Norway spruce and silver fir understory in continuous cover forestry. IForest 2020, 13, 56–64. [Google Scholar] [CrossRef]
- Mohytych, V.; Sułkowska, M.; Klisz, M. Reproduction of silver fir (Abies alba Mill) forests in the Ukrainian Carpathians. Folia For. Pol. Ser. A For. 2019, 61, 156–158. [Google Scholar] [CrossRef]
- Pach, M. Tempo zarastania spał na jodle oraz niektóre czynniki na nie wpływające [The rate of bark−stripping wound closure in fir and some factors affecting it]. Sylwan 2008, 152, 46–57. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. Available online: https://www.megasoftware.net/home (accessed on 10 October 2020). [CrossRef] [Green Version]
- BLAST Database. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome (accessed on 10 October 2020).
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method—a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Catalogue of Life. Available online: http://www.catalogueoflife.org (accessed on 10 October 2020).
- Index Fungorum. Available online: http://www.indexfungorum.org/names/names.asp (accessed on 10 October 2020).
- Patejuk, K.; Pusz, W. Ocena zdrowotności igieł sosny błotnej (Pinus×rhaetica) w Parku Narodowym Gór Stołowych [Evaluation of the health status of peat−bog pine (Pinus×rhaetica) needlesin Góry Stołowe National Park]. Sylwan 2017, 161, 861–869. [Google Scholar] [CrossRef]
- Pusz, W.; Zwijacz-Kozica, T. Zbiorowiska grzybów mikroskopijnych zasiedlających martwe drewno świerkowe w Tatrzańskim Parku Narodowym [Composition of microscopic fungi associated with the spruce dead woodin the Tatra National Park]. Sylwan 2017, 161, 312–319. [Google Scholar] [CrossRef]
- Pusz, W.; Ogórek, R. Communities of Microscopic Fungi in Dead Spruce in Relation to the Season and Degree of Wood Decay in Poland’s Karkonosze Mountains. Pol. J. Environ. Stud. 2020, 29, 2503–2511. [Google Scholar] [CrossRef]
- Liesch, J.M.; Meinz, M.S.; Onishi, J.C.; Schwartz, R.E.; Bills, R.E. Antifungal Agent Produced by Arthrinium Arundinis ATCC 74359. U.S. Patent No. 5,712,109, 27 January 1998. [Google Scholar]
- Hosseyni-Moghaddam, M.S.; Soltani, J. Bioactivity of endophytic Trichoderma fungal species from the plant family Cupressaceae. Ann. Microbiol. 2014, 64, 753–761. [Google Scholar] [CrossRef]
- Robles, C.A.; Lopez, S.E.; McCargo, P.D.; Carmarán, C.C. Relationships between fungal endophytes and wood-rot fungi in wood of Platanus acerifolia in urban environments. Can. J. For. Res. 2015, 45, 929–936. [Google Scholar] [CrossRef]
- Longa, C.M.; Pertot, I.; Tosi, S. Ecophysiological requirements and survival of a Trichoderma atroviride isolate with biocontrol potential. J. Basic Microbiol. 2008, 48, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.; Fink, S.; Schwarze, F.W. Evaluation of Trichoderma spp. as a biocontrol agent against wood decay fungi in urban trees. Biol. Control. 2008, 45, 111–123. [Google Scholar] [CrossRef]
- Pellegrini, A.; Prodorutti, D.; Pertot, I. Use of bark mulch pre-inoculated with Trichoderma atroviride to control Armillaria root rot. Crop. Prot. 2014, 64, 104–109. [Google Scholar] [CrossRef]
- Błaszczyk, L.; Strakowska, J.; Chełkowski, J.; Gąbka-Buszek, A.; Kaczmarek, J. Trichoderma species occurring on wood with decay symptoms in mountain forests in Central Europe: Genetic and enzymatic characterization. J. Appl. Genet. 2016, 57, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Czarnota, P. Sprawozdanie z Działalności Naukowej i Monitoringowej Gorczańskiego Parku Narodowego za rok 2018 [Report on the Scientific and Monitoring Activity of the Gorce National Park for 2018]. 2018. Available online: https://www.gorczanskipark.pl/UserFiles/File/Nauka/sprawozdania/Sprawozdanie%20z%20dzialalnosci%20N-E%20GPN%20za%20rok%202018.pdf (accessed on 20 January 2021).
- John, S.; Scott, E.S.; Wicks, T.J.; Hunt, J.S. Interactions between Eutypalata and Trichoderma harzianum. Phytopathol. Mediterr. 2004, 43, 95–104. [Google Scholar]
- Heilmann-Clausen, J.; Walleyn, R. Some records of wood-inhabiting fungi on Fagus sylvatica in northern Spain. Rev. Catalana Micol. 2007, 29, 67–80. [Google Scholar]
- Brglez, A.; Piškur, B.; Humar, M.; Gričar, J.; Ogris, N. The Effect of Eutypella parasitica on the Wood Decay of Three Maple Species. Forests 2020, 11, 671. [Google Scholar] [CrossRef]
- Vasiliauskas, R.; Stenlid, J. Population structure and genetic variation in Cylindrobasidium evolvens. Mycol. Res. 1998, 102, 1453–1458. [Google Scholar] [CrossRef]
- Burneviča, N.; Jansons, A.; Zaļuma, A.; Kļaviņa, D.; Jansons, J.; Gaitnieks, T. Fungi inhabiting bark stripping wounds made by large game on stems of Picea abies (L.) Karst. in Latvia. Balt For. 2016, 22, 2–7. [Google Scholar]
- Orlikowski, L.B.; Duda, B.; Szkuta, G. Phytophthora citricola on European beech and silver fir in Polish forest nurseries. J. Plant. Prot. Res. 2004, 44, 57–64. [Google Scholar]
- Metzler, B.; Hecht, U.; Nill, M.; Brüchert, F.; Fink, S.; Kohnle, U. Comparing Norway spruce and silver fir regarding impact of bark wounds. For. Ecol. Manag. 2012, 274, 99–107. [Google Scholar] [CrossRef]
- Gramaje, D.; Agustí-Brisach, C.; Pérez-Sierra, A.; Moralejo, E.; Olmo, D.; Mostert, L.; Armengol, J. Fungal trunk pathogens associated with wood decay of almond trees on Mallorca (Spain). Persoonia 2012, 28, 1–13. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DBH [cm] | Fresh Wounds | Healed Wounds | Old Wounds |
|---|---|---|---|
| 4.0–6.9 | 13 | 13 | 12 |
| 7.0–9.9 | 11 | 12 | 12 |
| 10.0–12.9 | 12 | 15 | 12 |
| 13.0–16.9 | 5 | 10 | 14 |
| Sum | 41 | 50 | 50 |
| Control > 30.0 | 11 | ||
| In total | 152 | ||
| Fungal Taxa | Number | Frequency [%] | Accession Number in NCBI | Identity with NCBI Isolates (Accession Number) |
|---|---|---|---|---|
| Alternaria alternata (Fr.) Keissl. | 70 | 6.67 | MW090865 | MT644140, MT487778 |
| Arthrinium arundinis (Corda) Dyko & B. Sutton | 81 | 7.71 | MW090861 MW090870 MW113167 MW090903 MW113168 MW113226 MW113227 MW113228 | MT582801, MT446201 |
| Aureobasidium pullulans (de Bary & Löwenthal) G. Arnaud | 4 | 0.38 | MW090923 | MT363099, MN922125 |
| Botrytis cinerea Pers. | 8 | 0.76 | MW090881 | MT573470, MN448502 |
| Cadophora sp. | 23 | 2.19 | MW090880 MW090922 | MF782737, MF188972 |
| Chaetomium sp. | 2 | 0.19 | MW090810 | MH171491, KC963908 |
| Coniochaeta sp. | 1 | 0.10 | MW090815 | MH859071, MG905629 |
| Coprinellus micaceus (Bull.) Vilgalys, Hopple & Jacq. Johnson | 3 | 0.29 | MW090910 | MH179313, GU227721 |
| Cucurbitariaceae sp. | 1 | 0.10 | MW090913 | KC963916, MK460387 |
| Cylindrobasidium evolvens (Fr.) Jülich | 90 | 8.57 | MW113169 | MN947592, MH854673 |
| Cystobasidium larynges (Reiersöl) Yurkov, Kachalkin, H.M. Daniel, M. Groenew., Libkind, V. de García, Zalar, Gouliam., Boekhout & Begerow | 1 | 0.10 | MW090823 | MH047192, KY103134 |
| Cytospora sp. | 19 | 1.81 | MW090820 MW090909 | KY051899, KU516449 |
| Epicoccum nigrum Link | 34 | 3.24 | MW090813 | MT548679, LC543647 |
| Eutypa sp. | 129 | 12.29 | MW090805 MW090860 MW090912 MW090911 | AY620998, KF453561 |
| Fusarium acuminatum Ellis & Everh. | 6 | 0.57 | MW090924 | MT649858, MT635295 |
| Fusarium avenaceum (Fr.) Sacc. | 1 | 0.10 | MW090925 | MT446118, MT276139 |
| Fusarium tricinctum (Corda) Sacc. | 15 | 1.43 | MW090875 MW090907 MW090918 | MK934343, KC311496 |
| Helotiales sp. | 1 | 0.10 | MW090817 | DQ317330, MF494618 |
| Heterobasidion annosum (Fr.) Bref. | 1 | 0.10 | MW090819 | MK395162, KU727784 |
| Neonectria neomacrospora (C. Booth & Samuels) Mantiri & Samuels | 5 | 0.48 | MW090919 | MH580206, MG049669 |
| Nigrospora oryzae (Berk. & Broome) Petch | 9 | 0.86 | MW090868 | MT556421, MG661721 |
| Paraphaeosphaeria neglecta Verkley, Riccioni & Stielow | 87 | 8.29 | MW090864 MW090874 MW090905 MW090906 | MK646057, MN244542 |
| Penicillium chrysogenum Thom | 17 | 1.62 | MW090830 | MT524448, MK762610 |
| Peniophoraceae sp. | 6 | 0.57 | MW090901 | MH010048, MH857634 |
| Phomopsis sp. | 107 | 10.19 | MW090829 MW090876 | MN538335, MT877030 |
| Preussia minima (Auersw.) Arx | 3 | 0.29 | MW090811 | MG457827, MT645911 |
| Rhizosphaera macrospora Gourb. & M. Morelet | 4 | 0.38 | MW090806 | AM884745, MN538337 |
| Rhizosphaera oudemansii Maubl. | 1 | 0.10 | MW090827 | KU516578, EU700366 |
| Sarea difformis (Fr.) Fr. | 5 | 0.48 | MW090809 | MN699648, FR837921 |
| Schizophyllum commune Fr. | 9 | 0.86 | MW113225 | MT647523, MT601951 |
| Sordaria fimicola (Roberge ex Desm.) Ces. & De Not. | 1 | 0.10 | MW090812 | MK965099, KX986578 |
| Stereum sanguinolentum (Alb. & Schwein.) Fr. | 6 | 0.57 | MW090831 | AY618670, AF533962 |
| Sydowia polyspora (Bref. & Tavel) E. Müll. | 28 | 2.67 | MW090808 MW090828 MW090873 | KY659505, KU837235 |
| Tolypocladium sp. | 2 | 0.19 | MW090807 | MN096582, MH730171 |
| Trametes versicolor (L.) Lloyd | 2 | 0.19 | MW090818 MW090825 | KJ995921, EU661891 |
| Trametes sp. | 3 | 0.29 | MW090814 | MK343672, MK269115 |
| Trichoderma atroviride P. Karst | 248 | 23.62 | MW090869 | MT341775, MT023026 |
| Truncatella angustata (Pers.) S. Hughes | 5 | 0.48 | MW090908 | MT514378, MK647988 |
| Valsa sp. | 1 | 0.10 | MW090826 | HQ654894, KY051908 |
| Xylariales sp. | 10 | 0.95 | MW090804 MW090816 MW090821 MW090822 MW090878 | KC774617, KF415082 |
| Zalerion sp. | 1 | 0.10 | MW090824 | AY465470, AF169308 |
| Fungal Taxa | Wound | Control | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fresh | Healed | Old | ||||||||||
| O | M | H | O | M | H | O | M | H | O | M | H | |
| Alternaria alternata | 14 | 6 | 9 | 6 | 6 | 1 | 11 | 12 | 5 | |||
| Arthrinium arundinis | 19 | 12 | 9 | 4 | 3 | 1 | 3 | 14 | 7 | 6 | 3 | |
| Aureobasidium pullulans | 1 | 3 | ||||||||||
| Botrytis cinerea | 3 | 1 | 3 | 1 | ||||||||
| Cadophora sp. | 6 | 11 | 6 | |||||||||
| Chaetomium sp. | 2 | |||||||||||
| Coniochaeta sp. | 1 | |||||||||||
| Coprinellus micaceus | 3 | |||||||||||
| Cucurbitariaceae sp. | 1 | |||||||||||
| Cylindrobasidium evolvens | 7 | 6 | 6 | 23 | 24 | 24 | ||||||
| Cystobasidium laryngis | 1 | |||||||||||
| Cytospora sp. | 6 | 6 | 6 | 1 | ||||||||
| Epicoccum nigrum | 9 | 6 | 11 | 5 | 3 | |||||||
| Eutypa sp. | 51 | 42 | 36 | |||||||||
| Fusarium acuminatum | 6 | |||||||||||
| Fusarium avenaceum | 1 | |||||||||||
| Fusarium tricinctum | 3 | 6 | 1 | 2 | 3 | |||||||
| Helotiales sp. | 1 | |||||||||||
| Heterobasidion annosum | 1 | |||||||||||
| Neonectria neomacrospora | 5 | |||||||||||
| Nigrospora oryzae | 6 | 3 | ||||||||||
| Paraphaeosphaeria neglecta | 2 | 6 | 6 | 6 | 1 | 12 | 18 | 12 | 12 | 12 | ||
| Penicillium chrysogenum | 1 | 4 | 6 | 6 | ||||||||
| Peniophoraceae sp. | ||||||||||||
| Phomopsis sp. | 7 | 10 | 15 | 3 | 1 | 37 | 20 | 12 | 2 | |||
| Preussia minima | 2 | 6 | ||||||||||
| Rhizosphaera macrospora | 4 | |||||||||||
| Rhizosphaera oudemansii | 1 | |||||||||||
| Sarea difformis | 5 | |||||||||||
| Schizophyllum commune | 1 | 2 | 6 | |||||||||
| Sordaria fimicola | 1 | |||||||||||
| Stereum sanguinolentum | 6 | |||||||||||
| Sydowia polyspora | 8 | 4 | 8 | 5 | 1 | 1 | 1 | |||||
| Tolypocladium sp. | 1 | 1 | ||||||||||
| Trametes versicolor | 1 | 1 | ||||||||||
| Trametes sp. | 3 | |||||||||||
| Trichoderma atroviride | 6 | 6 | 7 | 42 | 42 | 43 | 6 | 8 | 7 | 24 | 27 | 30 |
| Truncatella angustata | 2 | 3 | ||||||||||
| Valsa sp. | 1 | |||||||||||
| Xylariales sp. | 1 | 1 | 1 | 1 | 4 | 1 | 1 | |||||
| Zalerion sp. | 1 | |||||||||||
| In total: | 86 | 70 | 93 | 70 | 63 | 79 | 159 | 157 | 118 | 43 | 55 | 56 |
| Fungal Taxa | DBH [cm] | ||||
|---|---|---|---|---|---|
| 4.0–6.9 | 7.0–9.9 | 10.0–12.9 | 13.0–16.9 | Control | |
| Alternaria alternata | 43 | 6 | 4 | 12 | 5 |
| Arthrinium arundinis | 23 | 19 | 30 | 9 | |
| Aureobasidium pullulans | 3 | 1 | |||
| Botrytis cinerea | 4 | 2 | 1 | 1 | |
| Cadophora sp. | 12 | 11 | |||
| Chaetomium sp. | 2 | ||||
| Coniochaeta sp. | 1 | ||||
| Coprinellus micaceus | 3 | ||||
| Cucurbitariaceae sp. | 1 | ||||
| Cylindrobasidium evolvens | 33 | 33 | 18 | 6 | |
| Cystobasidium laryngis | 1 | ||||
| Cytospora sp. | 18 | 1 | |||
| Epicoccum nigrum | 7 | 6 | 18 | 3 | |
| Eutypa sp. | 42 | 24 | 33 | 30 | |
| Fusarium acuminatum | 6 | ||||
| Fusarium avenaceum | 1 | ||||
| Fusarium tricinctum | 1 | 2 | 9 | 3 | |
| Helotiales sp. | 1 | ||||
| Heterobasidion annosum | 1 | ||||
| Neonectria neomacrospora | 5 | ||||
| Nigrospora oryzae | 9 | ||||
| Paraphaeosphaeria neglecta | 20 | 23 | 6 | 48 | |
| Penicillium chrysogenum | 1 | 10 | 6 | ||
| Peniophoraceae sp. | 6 | ||||
| Phomopsis sp. | 17 | 19 | 41 | 28 | 2 |
| Preussia minima | 2 | 6 | |||
| Rhizosphaera macrospora | 2 | 2 | |||
| Rhizosphaera oudemansii | 1 | ||||
| Sarea difformis | 5 | ||||
| Schizophyllum commune | 9 | ||||
| Sordaria fimicola | 1 | ||||
| Stereum sanguinolentum | 6 | ||||
| Sydowia polyspora | 12 | 10 | 1 | 5 | |
| Tolypocladium sp. | 2 | ||||
| Trametes versicolor | 1 | 1 | |||
| Trametes sp. | 3 | ||||
| Trichoderma atroviride | 54 | 1 | 33 | 91 | 81 |
| Truncatella angustata | 5 | ||||
| Valsa sp. | 1 | ||||
| Xylariales sp. | 1 | 2 | 7 | ||
| Zalerion sp. | 1 | ||||
| In total: | 306 | 113 | 215 | 271 | 172 |
| Location | Type of Wound | Part of Wood | α-Diversity Indexes | ||
|---|---|---|---|---|---|
| Shannon-Wiener (H) | Pielou (J) | Simpson (D) | |||
| J | old | outside | 0.0000 | 0.0000 | |
| J | fresh | inside | 0.4101 | 0.5917 | 0.2449 |
| J | fresh | outside | 0.6902 | 0.9957 | 0.4970 |
| J | fresh | middle | 0.4101 | 0.5917 | 0.2449 |
| K | control | inside | 0.9831 | 0.7091 | 0.5139 |
| K | control | outside | 0.7078 | 0.6443 | 0.4224 |
| K | control | middle | 1.0693 | 0.7713 | 0.5730 |
| K | old | inside | 1.6771 | 0.8618 | 0.7800 |
| K | old | outside | 1.8821 | 0.8566 | 0.8180 |
| K | old | middle | 1.7873 | 0.8595 | 0.7956 |
| K | healed | inside | 0.6229 | 0.4493 | 0.3033 |
| K | healed | outside | 0.4258 | 0.3876 | 0.2037 |
| K | healed | middle | 0.3488 | 0.5033 | 0.1975 |
| K | fresh | inside | 1.6408 | 0.9157 | 0.7846 |
| K | fresh | outside | 1.4594 | 0.8145 | 0.7160 |
| K | fresh | middle | 0.6931 | 1.0000 | 0.5000 |
| P | old | inside | 0.0000 | 0.0000 | |
| P | old | outside | 0.2062 | 0.2975 | 0.0997 |
| P | old | middle | 0.0000 | 0.0000 | |
| P | healed | inside | 2.2745 | 0.9153 | 0.8698 |
| P | healed | outside | 1.9073 | 0.9172 | 0.8194 |
| P | healed | middle | 1.2425 | 0.8962 | 0.6667 |
| P | fresh | inside | 0.0000 | 0.0000 | |
| P | fresh | outside | 1.0889 | 0.7855 | 0.5800 |
| S | control | inside | 1.2309 | 0.8879 | 0.6746 |
| S | control | outside | 0.5623 | 0.8113 | 0.3750 |
| S | control | middle | 0.8699 | 0.7918 | 0.5128 |
| S | old | inside | 1.6627 | 0.8545 | 0.7756 |
| S | old | outside | 1.7293 | 0.8316 | 0.7686 |
| S | old | middle | 1.8943 | 0.8621 | 0.8278 |
| S | healed | inside | 0.7356 | 0.6696 | 0.4063 |
| S | healed | outside | 0.6730 | 0.9710 | 0.4800 |
| S | healed | middle | 0.6365 | 0.9183 | 0.4444 |
| S | fresh | inside | 2.0577 | 0.9365 | 0.8549 |
| S | fresh | outside | 1.9568 | 0.8906 | 0.8375 |
| S | fresh | middle | 1.9437 | 0.9347 | 0.8443 |
| Control | Fresh | Healed | Old |
|---|---|---|---|
| 0.43 | 0.38 | ||
| 0.42 | |||
| 0.34 | |||
| 0.63 | |||
| 0.59 | |||
| DBH [d1,3; cm] in 1992 | Number of Silver Fir Trees in 1992 | Number of the Same Silver Fir Trees in 2017 | Mortality [N] | Survival Rate [%] |
|---|---|---|---|---|
| 7.0–7.9 | 164 | 153 | 11 (1C) | 93.3 |
| 8.0–8.9 | 116 | 110 | 6 (2C) | 94.8 |
| 9.0–9.9 | 113 | 110 | 3 (1C) | 97.3 |
| 10.0–10.9 | 71 | 69 | 2 (1C) | 97.2 |
| 11.0–11.9 | 71 | 70 | 1 | 98.6 |
| 12.0–12.9 | 82 | 80 | 2 (1BW) | 97.6 |
| 13.0–13.9 | 46 | 45 | 1 (1BW) | 97.8 |
| 14.0–14.9 | 40 | 39 | 1 | 97.5 |
| 15.0–15.9 | 44 | 44 | 0 | 100 |
| 16.0–16.9 | 38 | 38 | 0 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pusz, W.; Baturo-Cieśniewska, A.; Kaczmarek-Pieńczewska, A.; Patejuk, K.; Czarnota, P. Is the Survivability of Silver Fir under Condition of Strong Ungulate Pressure Related to Mycobiota of Bark-Stripping Wounds? Forests 2021, 12, 976. https://doi.org/10.3390/f12080976
Pusz W, Baturo-Cieśniewska A, Kaczmarek-Pieńczewska A, Patejuk K, Czarnota P. Is the Survivability of Silver Fir under Condition of Strong Ungulate Pressure Related to Mycobiota of Bark-Stripping Wounds? Forests. 2021; 12(8):976. https://doi.org/10.3390/f12080976
Chicago/Turabian StylePusz, Wojciech, Anna Baturo-Cieśniewska, Agata Kaczmarek-Pieńczewska, Katarzyna Patejuk, and Paweł Czarnota. 2021. "Is the Survivability of Silver Fir under Condition of Strong Ungulate Pressure Related to Mycobiota of Bark-Stripping Wounds?" Forests 12, no. 8: 976. https://doi.org/10.3390/f12080976
APA StylePusz, W., Baturo-Cieśniewska, A., Kaczmarek-Pieńczewska, A., Patejuk, K., & Czarnota, P. (2021). Is the Survivability of Silver Fir under Condition of Strong Ungulate Pressure Related to Mycobiota of Bark-Stripping Wounds? Forests, 12(8), 976. https://doi.org/10.3390/f12080976