
The Impact of Different Environmental Conditions during Vegetative Propagation on Growth, Survival, and Biochemical Characteristics in Populus Hybrids in Clonal Field Trial

Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.2. Methods
3. Results and Discussion
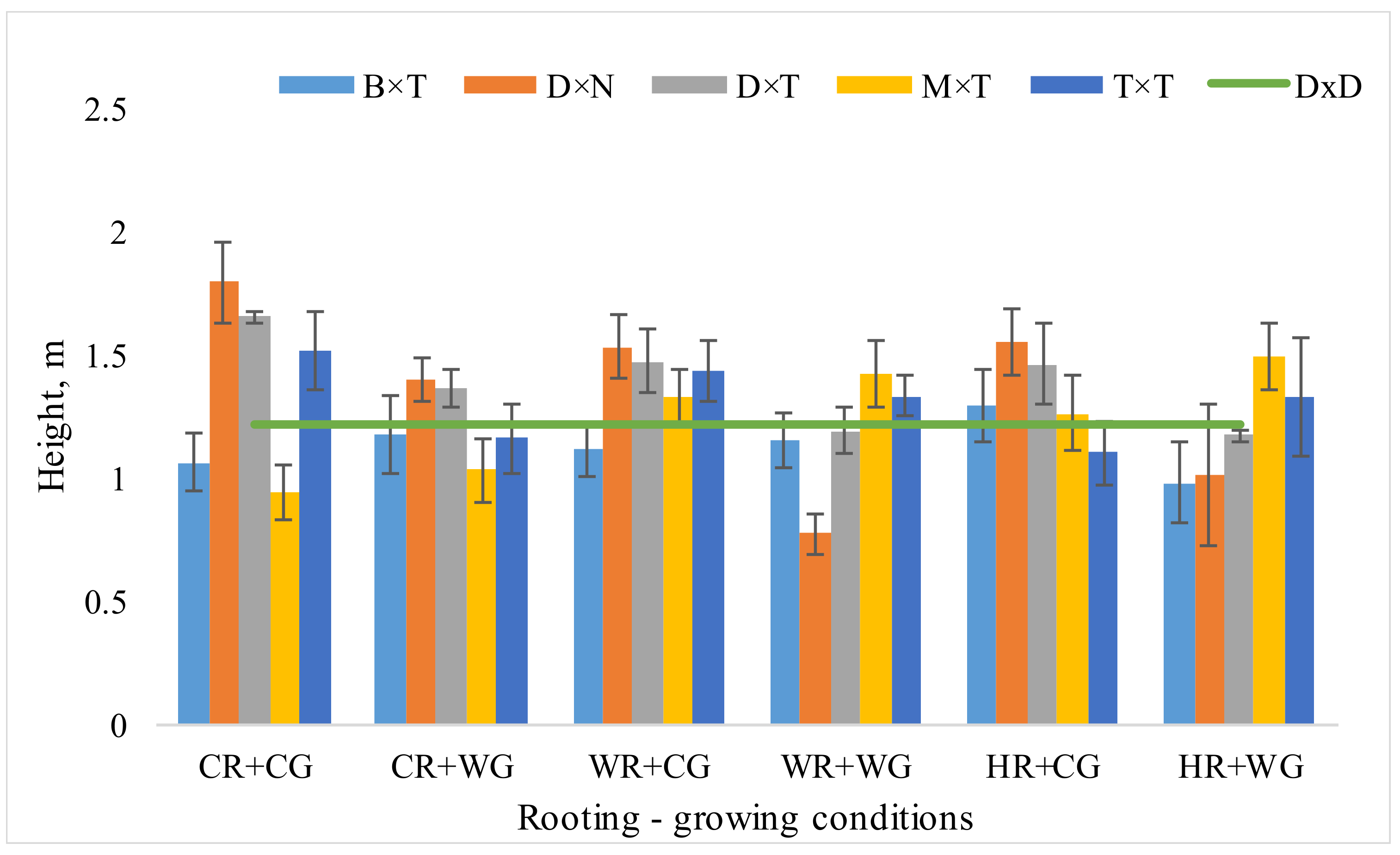
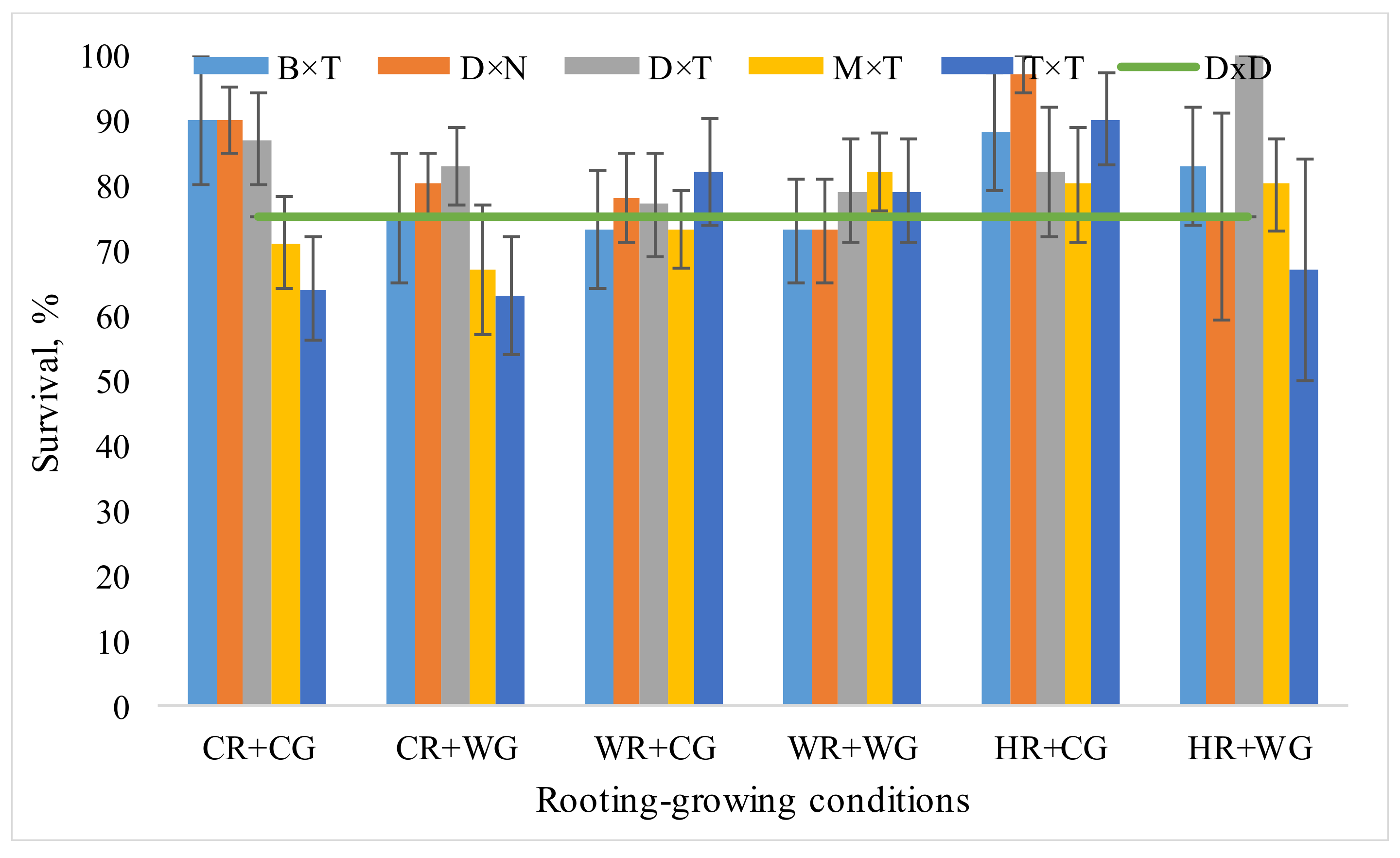
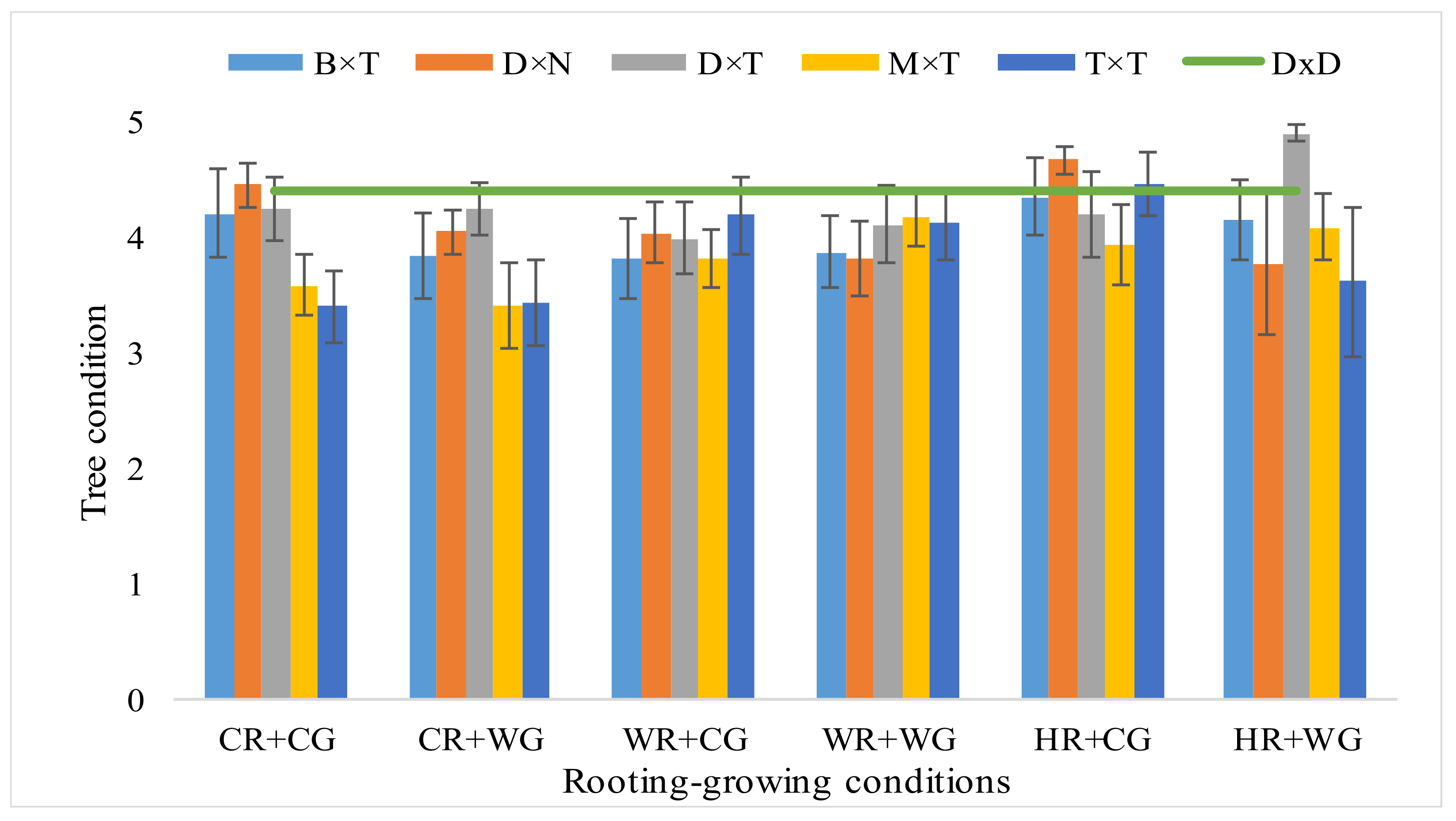
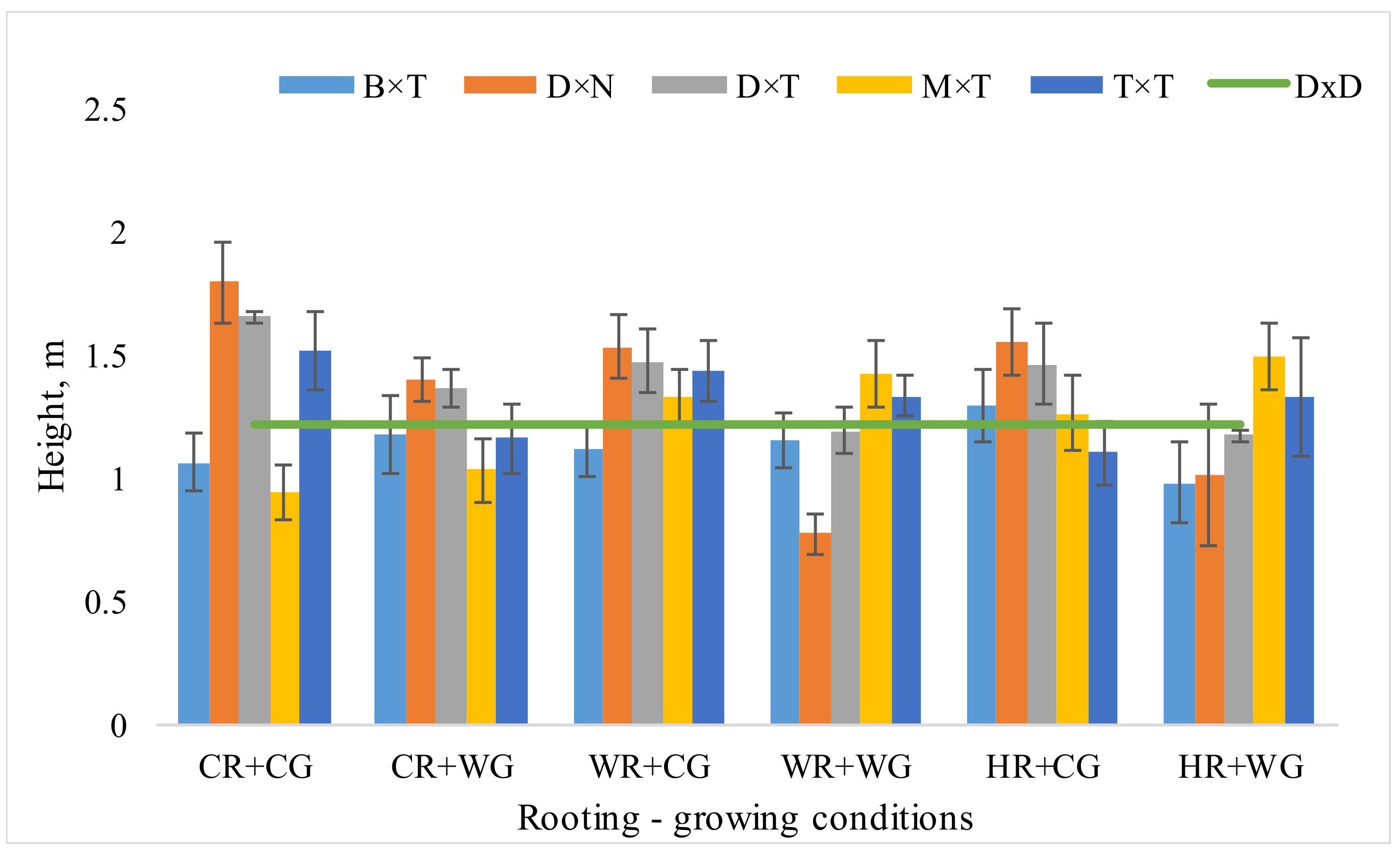
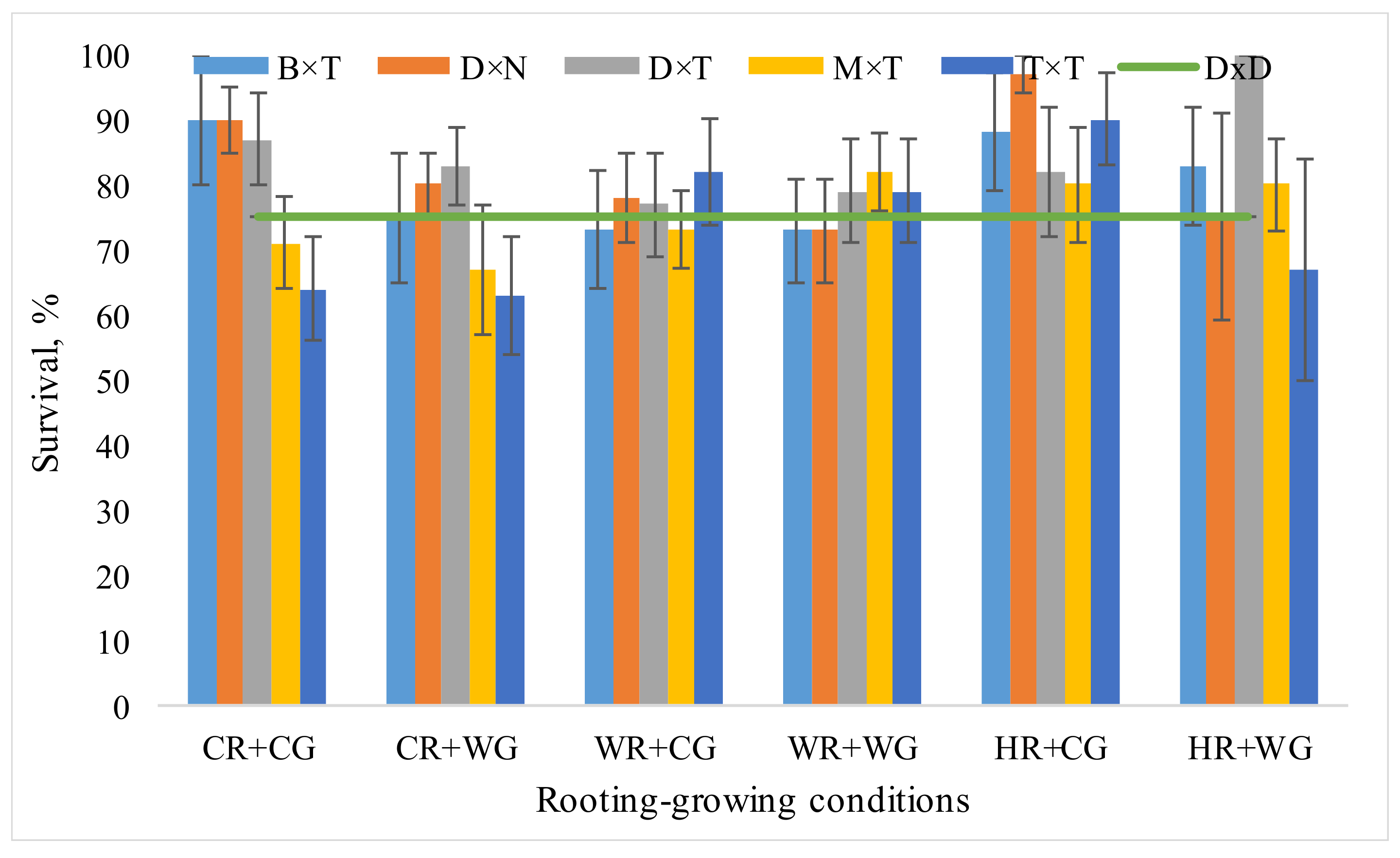
3.1. Dependence of Hybrid Populus Growth in Clonal Field Trial on the Vegatative Propagation (Rooting-Growing) Conditions
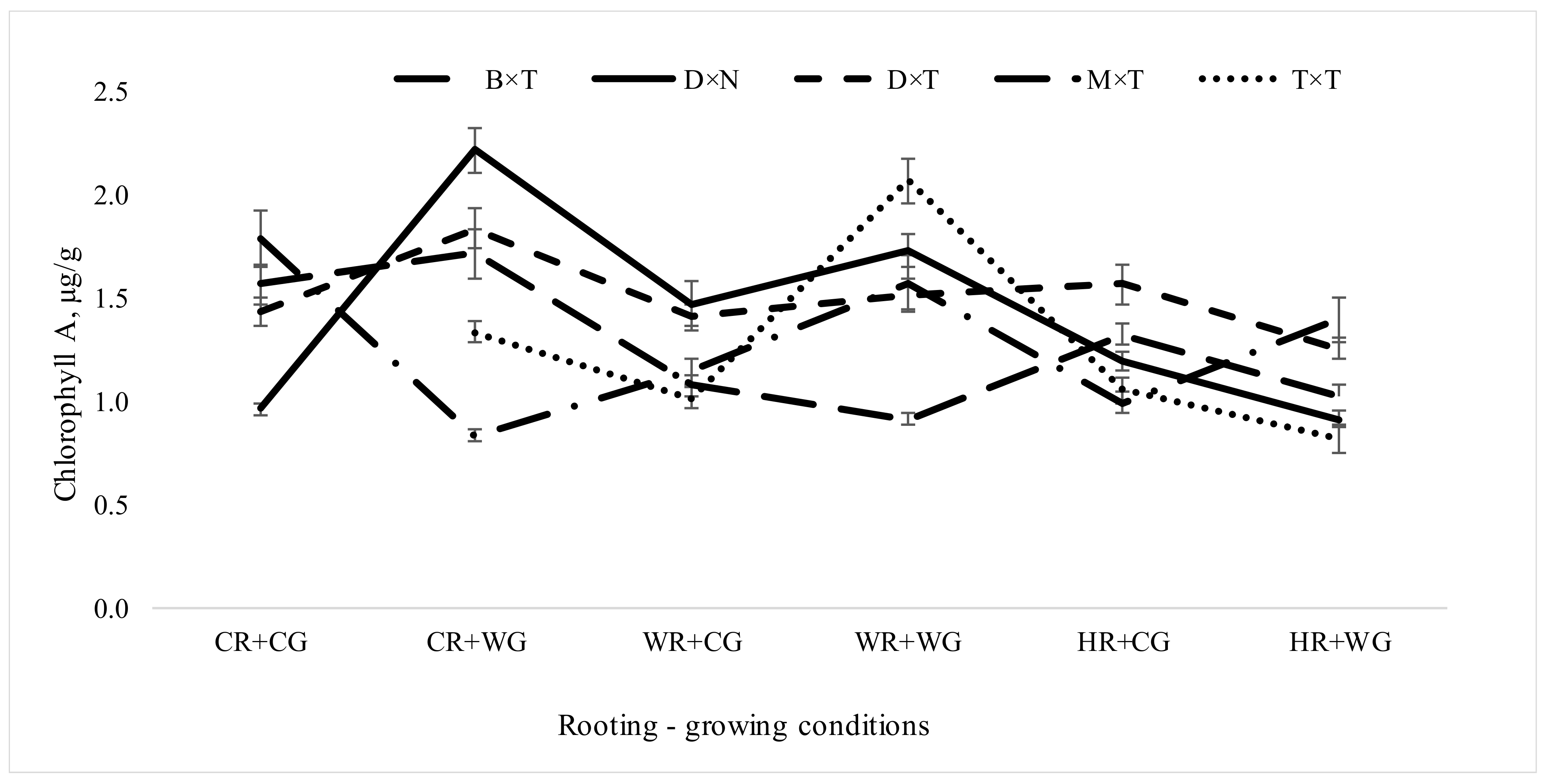
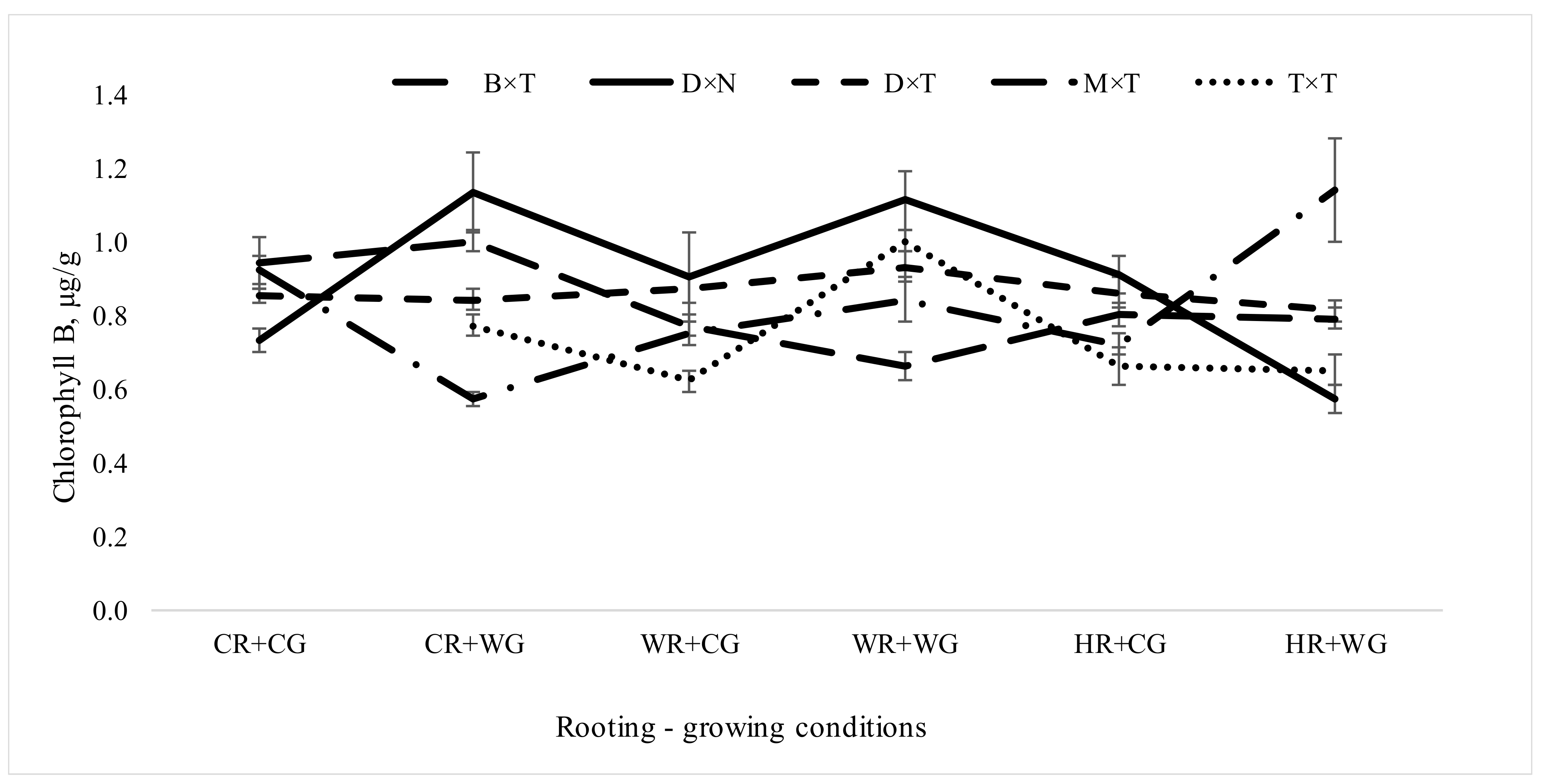
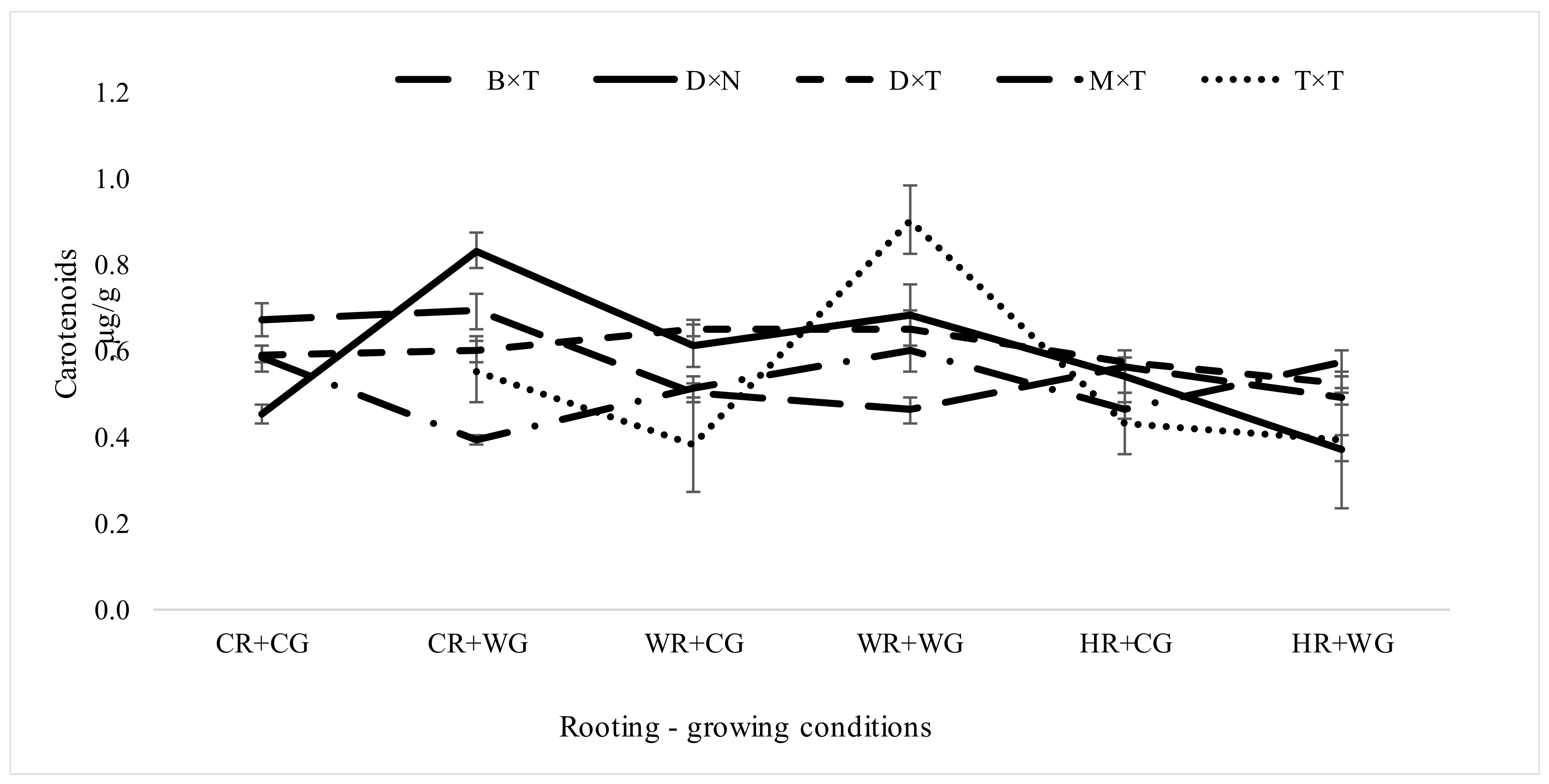
3.2. Dependence of Pigment Content in Leaves of Populus in Clonal Field Trial on Different Factors of Vegetative Propagation
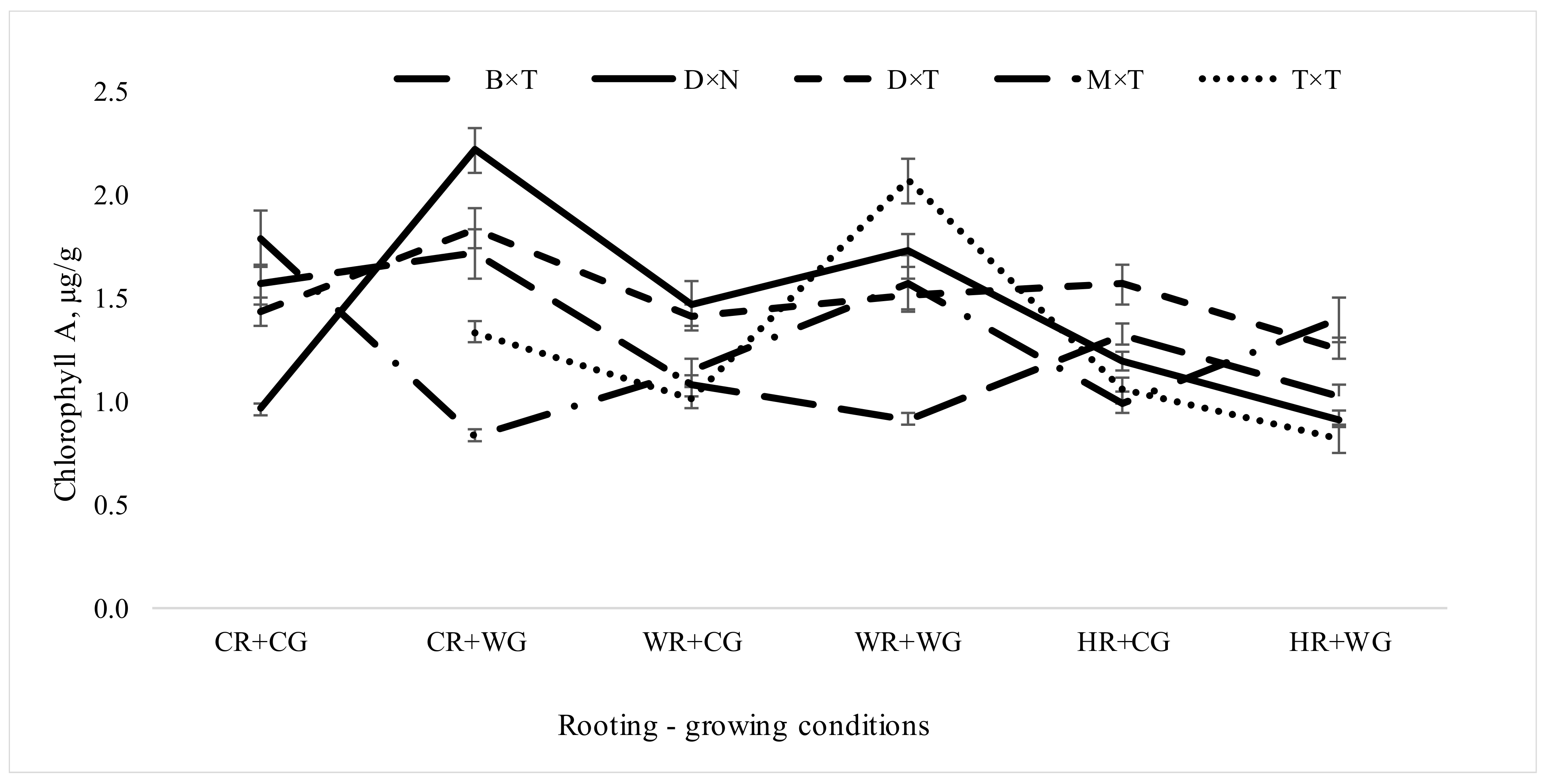
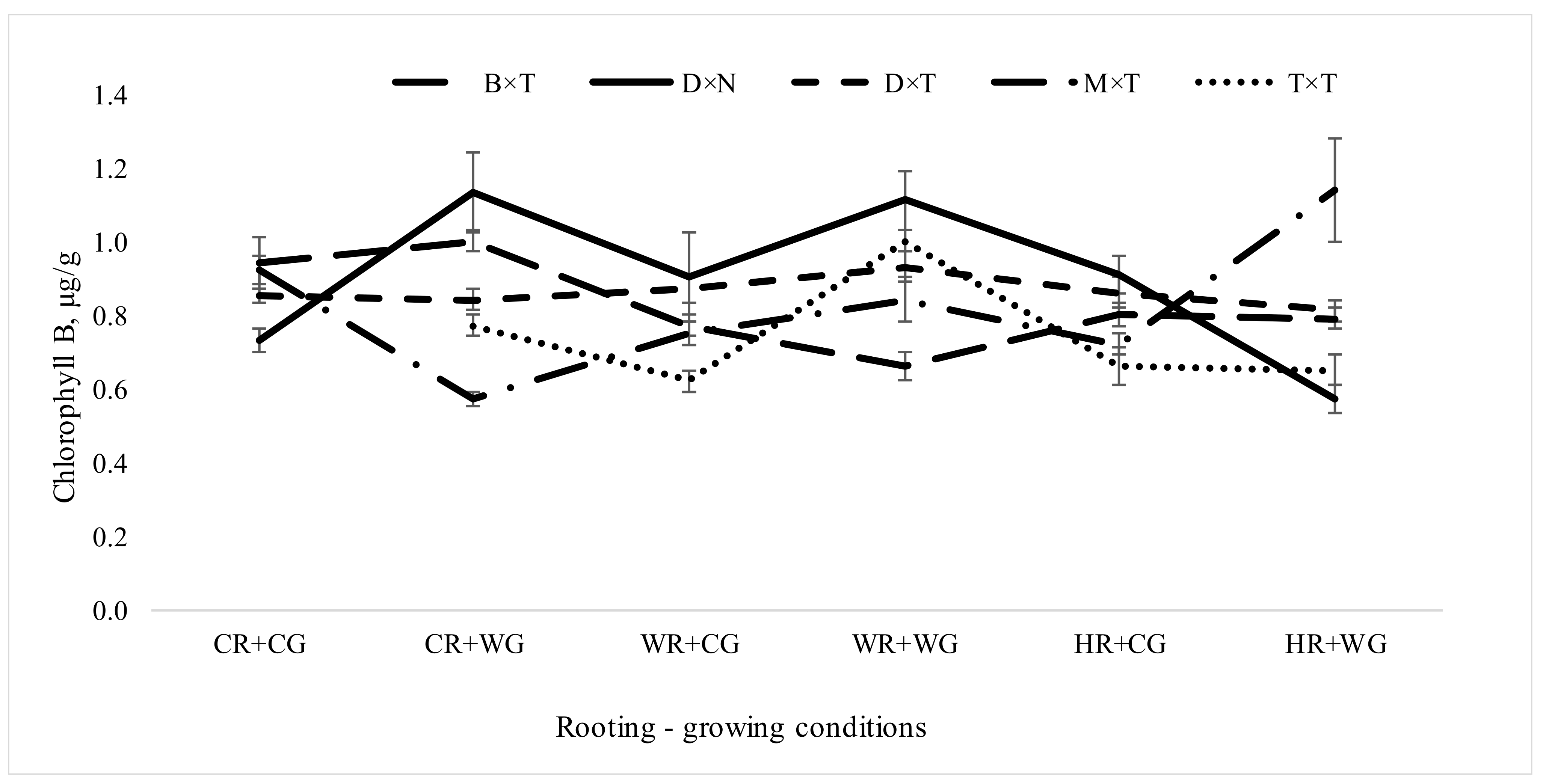
3.2.1. Impact of Hybrids and Treatments during Vegetative Propagation on Pigment Content in Hybrid Populus Leaves in Clonal Field Trial
3.2.2. Changes of Genetic Parameters of Different Traits of Trees in Clonal Field Trial Depending on Rooting-Growing Treatments during Vegetative Propagation in Phytotron Greenhouse
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Szczepańska, A.; Senetra, A. Forests as the key component of green belts surrounding urban areas. Balt. For. 2019, 25. [Google Scholar] [CrossRef]
- Sicard, P.; Agathokleous, E.; Araminiene, V.; Carrari, E.; Hoshika, Y.; De Marco, A.; Paoletti, E. Should we see urban trees as effective solutions to reduce increasing ozone levels in cities? Environ. Poll. 2018, 243, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.; Paula, L.; Dodd, T.; Németh, S.; Nanou, C.; Mega, V.; Campos, P. EU ambition to build the world’s leading bioe-conomy—Uncertain times demand innovative and sustainable solutions. New Biotechnol. 2018, 40, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Köhl, M.; Lasco, R.; Cifuentes, M.; Jonsson, Ö.; Korhonen, K.T.; Mundhenk, P.; Navar, J.D.J.; Stinson, G. Changes in forest production, biomass and carbon: Results from the 2015 UN FAO Global Forest Resource Assessment. For. Ecol. Manag. 2015, 352, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Payn, T.; Carnus, J.-M.; Freer-Smith, P.; Kimberley, M.; Kollert, W.; Liu, S.; Orazio, C.; Rodriguez, L.; Silva, L.N.; Wingfield, M.J. Changes in planted forests and future global implications. For. Ecol. Manag. 2015, 352, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Álvarez, P.; Pizarro, C.; Barrio-Anta, M.; Cámara-Obregón, A.; Bueno, J.L.M.; Álvarez, A.; Gutiérrez, I.; Burslem, D.F.R.P. Evaluation of Tree Species for Biomass Energy Production in Northwest Spain. Forests 2018, 9, 160. [Google Scholar] [CrossRef] [Green Version]
- Blonskaya, L.; Sultanova, R.; Muftakhova, S.; Martynova, M.; Konashova, S.; Sabirzyanov, I.; Odintsov, G. Biological indices of Bashkir Lombardy poplar (Populus nigra L. × Populus nigra var. italica Du Roi) in urban landscapes. Bulg. J. Agric. Sci. 2019, 25, 30–36. [Google Scholar]
- Chong, G.W.; Simonson, S.E.; Stohlgren, T.J.; Kalkhan, M.A. Biodiversity: Aspen stands have the lead, but will nonnative species take over. In Proceedings of the Sustaining Aspen in Western Landscapes Symposium, Grand Junction, CO, USA, 13–15 June 2000; p. 261. [Google Scholar]
- Nilsson, S.G.; Hedin, J.; Niklasson, M. Biodiversity and its Assessment in Boreal and Nemoral Forests. Scand. J. For. Res. 2001, 16, 10–26. [Google Scholar] [CrossRef] [Green Version]
- Latva-Karjanmaa, T.; Penttilä, R.; Siitonen, J. The demographic structure of European aspen (Populus tremula) populations in managed and old-growth boreal forests in eastern Finland. Can. J. For. Res. 2007, 37, 1070–1081. [Google Scholar] [CrossRef]
- De Rigo, D.; Enescu, C.M.; Houston Durrant, T.; Caudullo, G. Populus nigra in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; Publications Office of the European Union: 2 Rue Mercier, 2985 Luxembourg, 2016. [Google Scholar]
- Dickmann, D.I. An overview of the genus Populus. In Poplar Culture in North America; Part A; Canadian Science Publishing: Ottawa, ON, Canada, 2001. [Google Scholar]
- Isebrands, J.G.; Richardson, J. (Eds.) Poplars and Willows: Trees for Society and the Environment; CABI: Wallingford, UK, 2014. [Google Scholar]
- Blake, T.J.; Tschaplinski, T.; Eastham, A. Stomatal control of water use efficiency in poplar clones and hybrids. Can. J. Bot. 1984, 62, 1344–1351. [Google Scholar] [CrossRef]
- Eroğlu, E.; Acar, C. A Visual Assessment of Roadside Poplar Plantings in Turkey. J. Agric. Sci. 2018, 24, 185–198. [Google Scholar] [CrossRef]
- Gregg, J.W.; Jones, C.; Dawson, T.E. Urbanization effects on tree growth in the vicinity of New York City. Nat. Cell Biol. 2003, 424, 183–187. [Google Scholar] [CrossRef]
- Robinson, N. The Planting Design Handbook; Routledge: London, UK, 2016. [Google Scholar]
- Liu, B.; Liang, J.; Tang, G.; Wang, X.; Liu, F.; Zhao, D. Drought stress affects on growth, water use efficiency, gas exchange and chlorophyll fluorescence of Juglans rootstocks. Sci. Hortic. 2019, 250, 230–235. [Google Scholar] [CrossRef]
- Elliott, R.M.; Adkins, E.R.; Culligan, P.J.; Palmer, M.I. Stormwater infiltration capacity of street tree pits: Quantifying the influence of different design and management strategies in New York City. Ecol. Eng. 2018, 111, 157–166. [Google Scholar] [CrossRef]
- Gudynaitė-Franckevičienė, V.; Pliura, A.; Suchockas, V. Ecogenetic plasticity and genetic variation in Populus hybrids under the impact of simulated climate change related stressors. Balt. For. 2020, 26. [Google Scholar] [CrossRef]
- Je, S.-M.; Woo, S.Y.; Lee, S.H.; Kwak, M.J.; Lee, T.Y.; Kim, S.H. Combined effect of elevated CO2 concentration and drought on the photosynthetic apparatus and leaf morphology traits in seedlings of yellow poplar. Ecol. Res. 2017, 33, 403–412. [Google Scholar] [CrossRef]
- Pliūra, A.; Jankauskiene, J.; Lygis, V.; Suchockas, V.; Bajerkevičiene, G.; Verbylaite, R. Response of juvenile progeny of seven forest tree species and their populations to simulated climate change-related stressors, heat, elevated humidity and drought. iForest Biogeosc. For. 2018, 11, 374–388. [Google Scholar] [CrossRef] [Green Version]
- Pollastrini, M.; Puletti, N.; Selvi, F.; Iacopetti, G.; Bussotti, F. Widespread Crown Defoliation After a Drought and Heat Wave in the Forests of Tuscany (Central Italy) and Their Recovery—A Case Study from Summer 2017. Front. For. Glob. Chang. 2019, 2. [Google Scholar] [CrossRef] [Green Version]
- Sicard, P.; Augustaitis, A.; Belyazid, S.; Calfapietra, C.; de Marco, A.; Fenn, M.; Bytnerowicz, A.; Grulke, N.; He, S.; Matyssek, R.; et al. Global topics and novel approaches in the study of air pollution, climate change and forest ecosystems. Environ. Pollut. 2016, 213, 977–987. [Google Scholar] [CrossRef]
- Giesecke, T.; Brewer, S.; Finsinger, W.; Leydet, M.; Bradshaw, R.H. Patterns and dynamics of European vegetation change over the last 15,000 years. J. Biogeogr. 2017, 44, 1441–1456. [Google Scholar] [CrossRef] [Green Version]
- Kvaalen, H.; Johnsen, Ö. Timing of bud set in Picea abies is regulated by a memory of temperature during zygotic and somatic embryogenesis. New Phytol. 2007, 177, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Bräutigam, K.; Vining, K.J.; Lafon-Placette, C.; Fossdal, C.G.; Mirouze, M.; Gutiérrez Marcos, J.; Fluch, S.; Fernández Fraga, M.; Guevara, M.Á.; Abarca, D.; et al. Epigenetic regulation of adaptive responses of forest tree species to the environment. Ecol. Evolut. 2013, 3, 399–415. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Zhu, J.-K. Epigenetic regulation of stress responses in plants. Curr. Opin. Plant Biol. 2009, 12, 133–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyko, A.; Kovalchuk, I. Epigenetic control of plant stress response. Environ. Mol. Mutagen. 2008, 49, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Groszmann, M.; Greaves, I.K.; Albertyn, Z.I.; Scofield, G.N.; Peacock, W.J.; Dennis, E.S. Changes in 24-nt siRNA levels in Arabidopsis hybrids suggest an epigenetic contribution to hybrid vigor. Proc. Natl. Acad. Sci. USA 2011, 108, 2617–2622. [Google Scholar] [CrossRef] [Green Version]
- Mirouze, M.; Paszkowski, J. Epigenetic contribution to stress adaptation in plants. Curr. Opin. Plant Biol. 2011, 14, 267–274. [Google Scholar] [CrossRef]
- Bonasio, R.; Tu, S.; Reinberg, D. Molecular Signals of Epigenetic States. Science 2010, 330, 612–616. [Google Scholar] [CrossRef] [Green Version]
- Calarco, J.P.; Borges, F.; Donoghue, M.T.; Van Ex, F.; Jullien, P.E.; Lopes, T.; Gardner, R.; Berger, F.; Feijó, J.; Becker, J.; et al. Reprogramming of DNA Methylation in Pollen Guides Epigenetic Inheritance via Small RNA. Cell 2012, 151, 194–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnegan, E.J.; Genger, R.K.; Peacock, W.J.; Dennis, E.S. DNA methylation in plants. Ann. Rev. Plant Biol. 1998, 49, 223–247. [Google Scholar] [CrossRef]
- Niederhuth, C.E.; Schmitz, R.J. Covering Your Bases: Inheritance of DNA Methylation in Plant Genomes. Mol. Plant 2014, 7, 472–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- SAS Institute. SAS® 9.4 TS Level 1M4, X64_7PRO Platform; SAS Institute Inc.: Cary, NC, USA, 2020. [Google Scholar]
- Becker, W.A. Manual of Procedures in Quantitative Genetics, 4th ed.; Academic Enterprises: Pullman, WA, USA, 1984; 190p. [Google Scholar]
- Swiger, L.A.; Harvey, W.R.; Everson, D.O.; Gregory, K.E. The Variance of Intraclass Correlation Involving Groups with One Observation. Biometrics 1964, 20, 818. [Google Scholar] [CrossRef] [Green Version]
- Falconer, D.S. Introduction to Quantitative Genetics, 4th ed.; Trends in Genetics; Benjamin-Cummings Publishing: San Francisco, CA, USA, 1996; Volume 12, p. 280. [Google Scholar]
- Falconer, D.S. Introduction to Quantitative Genetics, 3rd ed.; Burnt Mill: Harlow, UK; Wiley: New York, NY, USA, 1989; p. 448. [Google Scholar]
- Ozkan, U.; Gökbulak, F. Effect of vegetation change from forest to herbaceous vegetation cover on soil moisture and temperature regimes and soil water chemistry. Catena 2017, 149, 158–166. [Google Scholar] [CrossRef]
- Tang, C.-S.; Shi, B.; Gao, L.; Daniels, J.L.; Jiang, H.-T.; Liu, C. Urbanization effect on soil temperature in Nanjing, China. Energy Build. 2011, 43, 3090–3098. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B.H. Hardwoods, Silvics of North America; US Department of Agriculture, Forest Service: Washington, DC, USA, 1990; p. 877.
- European Forest Genetic Resources Programme. 2017. Available online: www.euforgen.eu (accessed on 5 March 2021).
- Raj, S. Clone history shapes Populus drought responses. Proc. Natl. Acad. Sci. USA 2011, 108, 12521–12526. [Google Scholar] [CrossRef] [Green Version]
- Novák, V.; Lipiec, J. Water extraction by roots under environmental stresses. In Pollution and Water Resources, Columbia University Seminar Proceedings: Impact of Anthropogenic Activity and Climate Changes on the Environment of Central Europe and USA; Halasi-Kun, J., Stekauerová, V., Fodor, I., Nagy, V., Sinóros-Szabó, B., Lo Pinto, R., Eds.; Columbia University Press: New York, NY, USA, 2012. [Google Scholar]
- Cazzonelli, C.; Cuttriss, A.J.; Cossetto, S.B.; Pye, W.; Crisp, P.; Whelan, J.; Finnegan, E.J.; Turnbull, C.; Pogson, B.J. Regulation of Carotenoid Composition and Shoot Branching in Arabidopsis by a Chromatin Modifying Histone Methyltransferase, SDG8. Plant Cell 2009, 21, 39–53. [Google Scholar] [CrossRef] [Green Version]
- Heimann, L.; Horst, I.; Perduns, R.; Dreesen, B.; Offermann, S.; Peterhansel, C. A Common Histone Modification Code on C4 Genes in Maize and Its Conservation in Sorghum and Setaria italica. Plant Physiol. 2013, 162, 456–469. [Google Scholar] [CrossRef] [Green Version]
- Razaq, M.; Zhang, P.; Shen, H.-L. Salahuddin Influence of nitrogen and phosphorous on the growth and root morphology of Acer mono. PLoS ONE 2017, 12, e0171321. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Valiente, J.A.; Koehler, K.; Cavender-Bares, J. Climatic origins predict variation in photoprotective leaf pigments in response to drought and low temperatures in live oaks (Quercus series Virentes). Tree Physiol. 2015, 35, 521–534. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.; Je, S.; Kwak, M.; Akhmadi, K.; Tumurbaatar, E.; Khaine, I.; Lee, H.; Jang, J.; Kim, H.; Ahn, H.; et al. Physiological responses of Populus sibirica to different irrigation regimes for reforestation in arid area. S. Afr. J. Bot. 2017, 112, 329–335. [Google Scholar] [CrossRef]
- Kamble, P.N.; Giri, S.P.; Mane, R.S.; Tiwana, A. Estimation of chlorophyll content in young and adult leaves of some selected plants. Univ. J. Environ. Res. Technol. 2015, 5, 306–310. [Google Scholar]
- Ali, M.M.; Al-Ani, A.; Eamus, D.; Tan, D.K.Y. Leaf nitrogen determination using non-destructive techniques—A review. J. Plant Nutr. 2017, 40, 928–953. [Google Scholar] [CrossRef]
- Croft, H.; Chen, J.M.; Froelich, N.J.; Chen, B.; Staebler, R.M. Seasonal controls of canopy chlorophyll content on forest carbon uptake: Implications for GPP modeling. J. Geophys. Res. Biogeosci. 2015, 120, 1576–1586. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, H.; Uragami, C.; Cogdell, R.J. Carotenoids and Photosynthesis; Springer: Berlin/Heidelberg, Germany, 2016; Volume 79, pp. 111–139. [Google Scholar]
- Dhami, N.; Cazzonelli, C.I. Environmental impacts on carotenoid metabolism in leaves. Plant Growth Regul. 2020, 92, 455–477. [Google Scholar] [CrossRef]
- Morais, R.R.; Gonçalves, J.F.; Santos, U.M., Jr.; Dünisch, O.; Santos, A.L. Chloroplastid pigment contents and chlorophyll a fluorescence in amazonian tropical three species. Rev. Árvore 2007, 31, 959–966. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Smith, J.G.; Stringer, P.; Ennos, A.R. Effect of rooting conditions on the growth and cooling ability of Pyrus calleryana. Urban. For. Urban. Green. 2011, 10, 185–192. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Babani, F. Light Adaptation and Senescence of the Photosynthetic Apparatus. Changes in Pigment Composition, Chlorophyll Fluorescence Parameters and Photosynthetic Activity. Chlorophyll A Fluoresc. 2007, 19, 713–736. [Google Scholar] [CrossRef]
- Soujanya, P.; Kulkarni, B.S.; Harshavardhan, M. Genetic variability and heritability in rose. J. Pharm. Phytochem. 2019, 8, 2065–2067. [Google Scholar]
- Thulasiram, L.B.; Bhople, S.R.; Srikanth, M.; Nayak, B.R. Genetic variability and heritability studies in okra (Abelmoschus esculentus (L.) Moench). Plant Arch. 2017, 17, 907–910. [Google Scholar]
- Kumar, A.N.U.J.; Gaurav, S.S.; Bahuguba, D.K.; Sharma, P.; Singh, T.; Chand, P. Analysis of variability, heritability and genetic advance for yield and yield related trait in wheat (Triticum aestivum L.) genotypes. Inter. J. Agri. Sci. Res. 2017, 7, 4. [Google Scholar]
- Wang, Q.; Xie, W.; Xing, H.; Yan, J.; Meng, X.; Li, X.; Fu, X.; Xu, J.; Lian, X.; Yu, S.; et al. Genetic Architecture of Natural Variation in Rice Chlorophyll Content Revealed by a Genome-Wide Association Study. Mol. Plant 2015, 8, 946–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossdorf, O.; Richards, C.; Pigliucci, M. Epigenetics for ecologists. Ecol. Lett. 2007, 11, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Avramidou, E.; Ganopoulos, I.; Doulis, A.G.; Tsaftaris, A.S.; Aravanopoulos, F.A. Beyond population genetics: Natural epigenetic variation in wild cherry (Prunus avium). Tree Genet. Genomes 2015, 11, 1–9. [Google Scholar] [CrossRef]
- Markus, C.; Pecinka, A.; Karan, R.; Barney, J.N.; Merotto, A., Jr. Epigenetic regulation-contribution to herbicide resistance in weeds? Pest. Manag. Sci. 2018, 74, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.M.; Kim, M.; Ralph, P.J.; Marín-Guirao, L.; Pernice, M.; Procaccini, G. Stress Memory in Seagrasses: First Insight into the Effects of Thermal Priming and the Role of Epigenetic Modifications. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crossing Type | Hybrid Abbreviation | Crossing Combination | Clone Number or Cultivar Name Abbreviation |
|---|---|---|---|
| Inter-specific | D × N | P. deltoides × P. nigra | Gr-Comp, Gr-Xe-3, Nyd-Elle, UK-AgatF, UK-Robus, UK-Spitk |
| D × T | P. deltoides × P. trichocarpa | Isl-15, UK-Boela, UK-Donk | |
| M × T | P. maximowiczii × P. trichocarpa | SvSFPo2, SvSFPo6, SvSFPo7, UK-Andro | |
| B × T | P. balsamifera × P. trichocarpa | SvSFPo1, SvSFPo4, SvSFPo13 | |
| Intra- specific | T × T | P. trichocarpa × P. trichocarpa | SvSFPo14, SvSFPo15, UK-FrPau, SvSFPo9 |
| Effect | Num. DF | Den. DF | F Value | Prob F | p |
|---|---|---|---|---|---|
| Height | |||||
| Treatments | 5 | 606 | 2.15 | 0.058 | . |
| Hybrid | 4 | 606 | 1.13 | 0.393 | . |
| Interaction hybrid × treatments | 20 | 606 | 2.96 | <0.0001 | *** |
| Diameter | |||||
| Treatments | 5 | 606 | 2.48 | 0.031 | * |
| Hybrid | 4 | 606 | 3.79 | 0.004 | ** |
| Interaction hybrid × treatments | 20 | 606 | 2.89 | <0.0001 | *** |
| Effect | Num. DF | Den. DF | F Value | Prob F | p |
|---|---|---|---|---|---|
| Chlorophyll A | |||||
| Growing conditions | 1 | 824 | 35.20 | <0.0001 | *** |
| Rooting conditions | 2 | 824 | 30.61 | <0.0001 | *** |
| Interaction rooting × growing conditions | 2 | 824 | 10.91 | <0.0001 | *** |
| Hybrid | 4 | 811 | 8.14 | <0.0001 | *** |
| Interaction rooting × growing conditions × hybrid | 7 | 811 | 24.94 | <0.0001 | *** |
| Chlorophyll B | |||||
| Growing conditions | 1 | 824 | 14.84 | 0.0001 | *** |
| Rooting conditions | 2 | 824 | 3.96 | 0.0195 | * |
| Interaction rooting × growing conditions | 2 | 824 | 2.65 | 0.0715 | . |
| Hybrid | 4 | 811 | 5.99 | <0.0001 | *** |
| Interaction rooting × growing conditions × hybrid | 7 | 811 | 18.34 | <0.0001 | *** |
| Chlorophyll A and B ratio | |||||
| Growing conditions | 1 | 824 | 24.01 | <0.0001 | *** |
| Rooting conditions | 2 | 824 | 21.65 | <0.0001 | *** |
| Interaction rooting × growing conditions | 2 | 824 | 7.48 | 0.0006 | *** |
| Hybrid | 4 | 811 | 7.36 | <0.0001 | *** |
| Interaction rooting × growing conditions × hybrid | 7 | 811 | 18.44 | <0.0001 | *** |
| Carotenoids | |||||
| Growing conditions | 1 | 824 | 21.63 | <0.0001 | *** |
| Rooting conditions | 2 | 824 | 26.14 | <0.0001 | *** |
| Interaction rooting × growing conditions | 2 | 824 | 8.66 | 0.0002 | *** |
| Hybrid | 4 | 811 | 9.20 | <0.0001 | *** |
| Interaction rooting × growing conditions × hybrid | 7 | 811 | 13.03 | <0.0001 | *** |
| Trait | Treatment | CVG, % | Hi2 ± s.e, | Hm2± s.e | CVF, % | Trait Mean ± se | Clonal Variance Component, ±se | p |
|---|---|---|---|---|---|---|---|---|
| Chlorophyll A | CR + CG | 27.10 | 0.52 ± 0.16 | 0.94 ± 0.03 | 36.16 | 1.29 ± 0.04 | 0.12 ± 0.07 | . |
| CR + WG | 37.70 | 0.65 ± 0.11 | 0.97 ± 0.02 | 44.57 | 1.64 ± 0.05 | 0.38 ± 0.18 | * | |
| WR + CG | 26.22 | 0.40 ± 0.13 | 0.91 ± 0.04 | 40.64 | 1.26 ± 0.04 | 0.10 ± 0.05 | * | |
| WR + WG | 30.72 | 0.63 ± 0.13 | 0.96 ± 0.02 | 37.44 | 1.58 ± 0.05 | 0.23 ± 0.12 | * | |
| HR + CG | 32.05 | 0.60 ± 0.12 | 0.96 ± 0.02 | 39.28 | 1.25 ± 0.03 | 0.16 ± 0.07 | * | |
| HR + WG | 20.97 | 0.28 ± 0.13 | 0.85 ± 0.07 | 39.08 | 1.09 ± 0.03 | 0.05 ± 0.03 | . | |
| Chlorophyll B | CR + CG | 12.12 | 0.30 ± 0.14 | 0.87 ± 0.07 | 21.67 | 0.82 ± 0.01 | 0.00 ± 0.00 | . |
| CR + WG | 25.97 | 0.66 ± 0.11 | 0.97 ± 0.01 | 30.36 | 0.89 ± 0.02 | 0.05 ± 0.02 | * | |
| WR + CG | 23.07 | 0.50 ± 0.13 | 0.94 ± 0.02 | 31.91 | 0.81 ± 0.02 | 0.03 ± 0.01 | * | |
| WR + WG | 30.51 | 0.61 ± 0.13 | 0.96 ± 0.02 | 37.76 | 0.95 ± 0.03 | 0.08 ± 0.04 | * | |
| HR + CG | 20.37 | 0.41 ± 0.13 | 0.92 ± 0.04 | 30.50 | 0.83 ± 0.01 | 0.02 ± 0.01 | * | |
| HR + WG | 41.11 | 0.49 ± 0.15 | 0.93 ± 0.03 | 57.09 | 0.81 ± 0.04 | 0.11 ± 0.06 | * | |
| Carotenoids | CR + CG | 16.50 | 0.40 ± 0.16 | 0.91 ± 0.05 | 25.51 | 0.54 ± 0.01 | 0.00 ± 0.00 | . |
| CR + WG | 30.51 | 0.63 ± 0.12 | 0.97 ± 0.02 | 36.55 | 0.63 ± 0.01 | 0.03 ± 0.01 | * | |
| WR + CG | 32.13 | 0.53 ± 0.12 | 0.94 ± 0.03 | 42.86 | 0.55 ± 0.01 | 0.03 ± 0.01 | * | |
| WR + WG | 27.59 | 0.64 ± 0.13 | 0.96 ± 0.02 | 33.25 | 0.65 ± 0.01 | 0.03 ± 0.01 | * | |
| HR + CG | 19.93 | 0.41 ± 0.13 | 0.92 ± 0.04 | 30.15 | 0.53 ± 0.01 | 0.01 ± 0.00 | * | |
| HR + WG | 18.24 | 0.34 ± 0.14 | 0.88 ± 0.06 | 30.80 | 0.47 ± 0.01 | 0.00 ± 0.00 | * | |
| Chlorophyll A and B ratio | CR + CG | 14.97 | 0.35 ± 0.15 | 0.89 ± 0.06 | 24.76 | 1.56 ± 0.03 | 0.05 ± 0.03 | . |
| CR + WG | 18.67 | 0.52 ± 0.13 | 0.95 ± 0.02 | 24.74 | 1.81 ± 0.03 | 0.11 ± 0.05 | * | |
| WR + CG | 3.74 | 0.04 ± 0.05 | 0.36 ± 0.13 | 19.54 | 1.56 ± 0.02 | 0.00 ± 0.00 | . | |
| WR + WG | 12.38 | 0.31 ± 0.13 | 0.87 ± 0.06 | 21.86 | 1.70 ± 0.03 | 0.04 ± 0.02 | * | |
| HR + CG | 15.70 | 0.41 ± 0.13 | 0.92 ± 0.04 | 23.88 | 1.51 ± 0.02 | 0.05 ± 0.02 | * | |
| HR + WG | 17.77 | 0.35 ± 0.14 | 0.89 ± 0.05 | 29.27 | 1.45 ± 0.03 | 0.06 ± 0.04 | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gudynaitė-Franckevičienė, V.; Pliūra, A. The Impact of Different Environmental Conditions during Vegetative Propagation on Growth, Survival, and Biochemical Characteristics in Populus Hybrids in Clonal Field Trial. Forests 2021, 12, 892. https://doi.org/10.3390/f12070892
Gudynaitė-Franckevičienė V, Pliūra A. The Impact of Different Environmental Conditions during Vegetative Propagation on Growth, Survival, and Biochemical Characteristics in Populus Hybrids in Clonal Field Trial. Forests. 2021; 12(7):892. https://doi.org/10.3390/f12070892
Chicago/Turabian StyleGudynaitė-Franckevičienė, Valda, and Alfas Pliūra. 2021. "The Impact of Different Environmental Conditions during Vegetative Propagation on Growth, Survival, and Biochemical Characteristics in Populus Hybrids in Clonal Field Trial" Forests 12, no. 7: 892. https://doi.org/10.3390/f12070892
APA StyleGudynaitė-Franckevičienė, V., & Pliūra, A. (2021). The Impact of Different Environmental Conditions during Vegetative Propagation on Growth, Survival, and Biochemical Characteristics in Populus Hybrids in Clonal Field Trial. Forests, 12(7), 892. https://doi.org/10.3390/f12070892