
Winter Food Availability for Wild Herbivores Depending on the Type of Forest Regeneration


Abstract
:1. Introduction
Study Hypotheses
- The regeneration method will significantly affect the amount of available biomass, where the smallest amount of biomass will result from using the Clear Cutting method, and the relatively lowest number of woody plants per unit area will be expected.
- The largest amount of biomass available to herbivores will be young vegetation with low side branches.
- The highest browsing damage will occur when using the Clear Cutting method due to the small number of woody plants in the area.
2. Materials and Methods
2.1. Study Area
2.2. Biomass of Shoots
2.3. Woody Plants Damage Intensity
2.4. Presence of Herbivores
2.5. Statistical Methods
3. Results
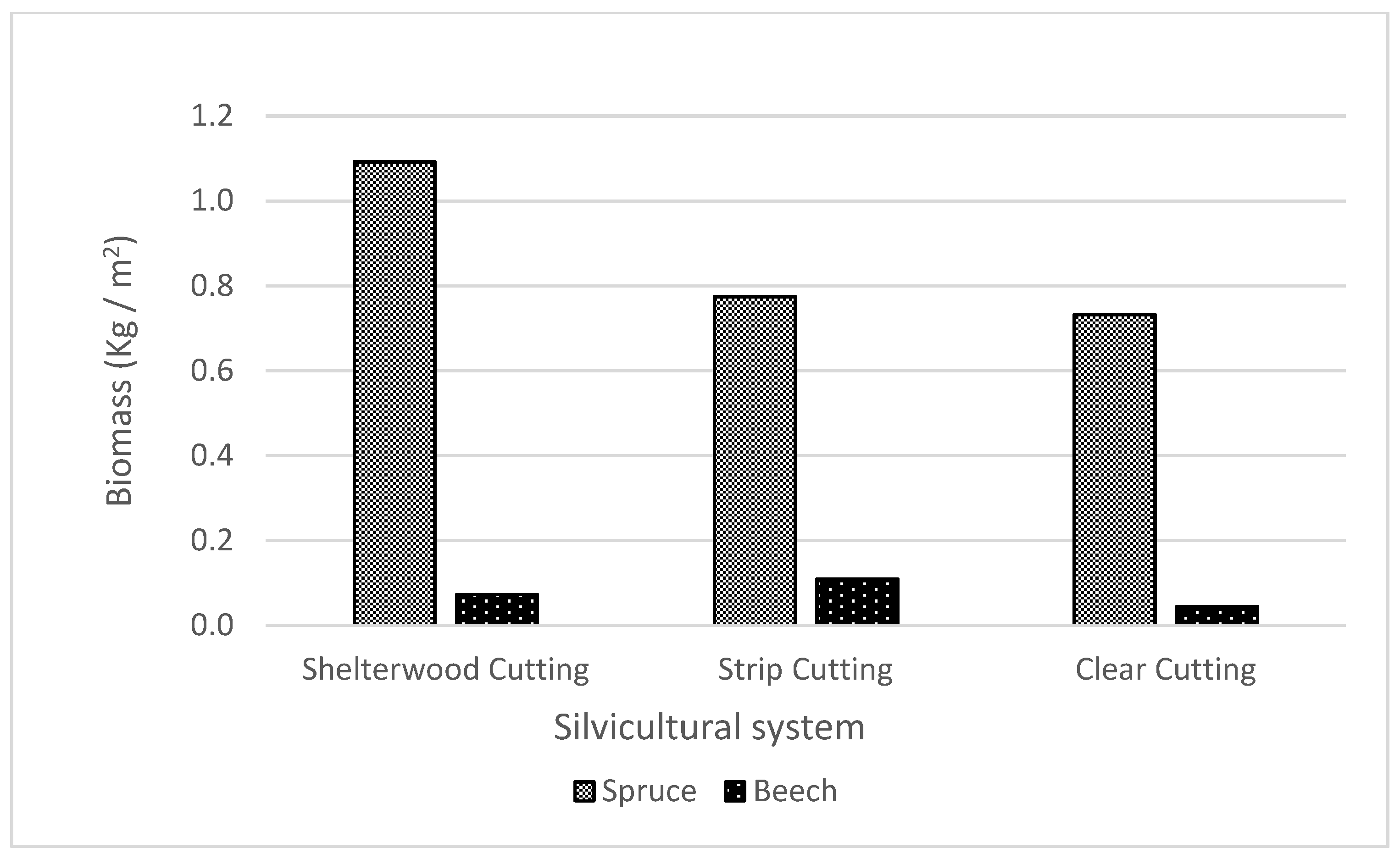
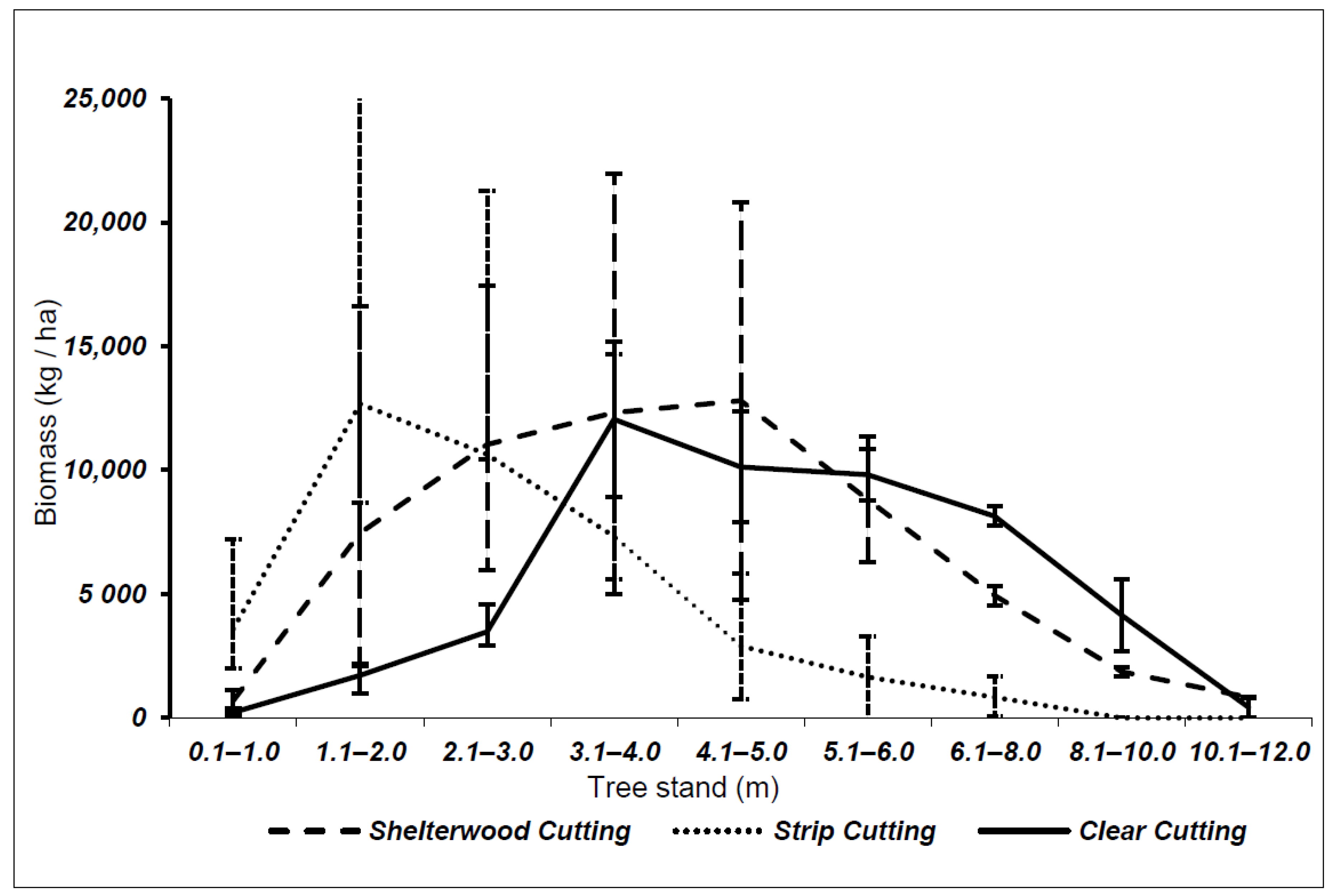
3.1. Biomass of Shoots
3.2. Woody Plants Damage Intensity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ramirez, J.I.; Jansen, P.A.; Poorter, L. Effects of wild ungulates on the regeneration, structure and functioning of temperate forests: A semi-quantitative review. For. Ecol. Manag. 2018, 424, 406–419. [Google Scholar] [CrossRef]
- Ramirez, J.I.; Jansen, P.A.; den Ouden, J.; Goudzwaard, L.; Poorter, L. Long-term effects of wild ungulates on the structure, composition and succession of temperate forests. For. Ecol. Manag. 2019, 432, 478–488. [Google Scholar] [CrossRef]
- Conover, M.R.; Butikofer, E.; Decker, D.J. Wildlife damage to crops: Perceptions of agricultural and wildlife leaders in 1957, 1987, and 2017. Wildl. Soc. Bull. 2018, 42, 551–558. [Google Scholar] [CrossRef]
- Hohbein, R.R.; Mengak, M.T. Cooperative extension agents as key informants in assessing wildlife damage trends in Georgia. Hum. Wildl. Interact. 2018, 12, 10. [Google Scholar]
- Schulze, E.D.; Bouriaud, O.; Wäldchen, J.; Eisenhauer, N.; Walentowski, H.; Seele, C.; Heinze, E.; Pruschitzki, U.; Dănilă, G.; Marin, G.; et al. Ungulate browsing causes species loss in deciduous forests independent of community dynamics and silvicultural management in Central and Southeastern Europe. Ann. For. Res. 2014, 57, 267–288. [Google Scholar] [CrossRef] [Green Version]
- Spake, R.; Bellamy, C.; Gill, R.; Watts, K.; Wilson, T.; Ditchburn, B.; Eigenbrod, F. Forest damage by hare depends on cross-scale interactions between climate, deer density and landscape structure. J. Appl. Ecol. 2020, 57, 1376–1390. [Google Scholar] [CrossRef]
- Faison, E.K.; DeStefano, S.; Foster, D.R.; Plotkin, A.B. Functional response of ungulate browsers in disturbed eastern hemlock forests. For. Ecol. Manag. 2016, 362, 177–183. [Google Scholar] [CrossRef]
- Velamazán, M.; San Miguel, A.; Escribano, R.; Perea, R. Compatibility of regeneration silviculture and wild ungulates in a Mediterranean pine forest: Implications for tree recruitment and woody plant diversity. Ann. For. Sci. 2018, 75, 35. [Google Scholar] [CrossRef] [Green Version]
- Ward, A.I.; White, P.C.; Walker, N.J.; Critchley, C.H. Conifer leader browsing by roe deer in English upland forests: Effects of deer density and understorey vegetation. For. Ecol. Manag. 2008, 256, 1333–1338. [Google Scholar] [CrossRef]
- Heinze, E.; Boch, S.; Fischer, M.; Hessenmöller, D.; Klenk, B.; Müller, J.; Prati, D.; Schulze, E.D.; Seele, C.; Socher, S.; et al. Habitat use of large ungulates in northeastern Germany in relation to forest management. For. Ecol. Manag. 2011, 261, 288–296. [Google Scholar] [CrossRef]
- Moser, B.; Schütz, M.; Hindenlang, K.E. Importance of alternative food resources for browsing by roe deer on deciduous trees: The role of food availability and species quality. For. Ecol. Manag. 2006, 226, 248–255. [Google Scholar] [CrossRef]
- Kuijper, D.P.; Cromsigt, J.P.; Churski, M.; Adam, B.; Jędrzejewska, B.; Jędrzejewski, W. Do ungulates preferentially feed in forest gaps in European temperate forest? For. Ecol. Manag. 2009, 258, 1528–1535. [Google Scholar] [CrossRef]
- Gerhardt, P.; Arnold, J.M.; Hackländer, K.; Hochbichler, E. Determinants of deer impact in European forests–A systematic literature analysis. For. Ecol. Manag. 2013, 310, 173–186. [Google Scholar] [CrossRef]
- Arnold, J.M.; Gerhardt, P.; Steyaert, S.M.; Hochbichler, E.; Hackländer, K. Diversionary feeding can reduce red deer habitat selection pressure on vulnerable forest stands, but is not a panacea for red deer damage. For. Ecol. Manag. 2018, 407, 166–173. [Google Scholar] [CrossRef]
- White, K.S.; Pendleton, G.W.; Hood, E. Effects of snow on Sitka black-tailed deer browse availability and nutritional carrying capacity in southeastern Alaska. J. Wildl. Manag. 2009, 73, 481–487. [Google Scholar] [CrossRef]
- Spitzer, R.; Felton, A.; Landman, M.; Singh, N.J.; Widemo, F.; Cromsigt, J.P. Fifty years of European ungulate dietary studies: A synthesis. Oikos 2020, 129, 1668–1680. [Google Scholar] [CrossRef]
- D’Aprile, D.; Vacchiano, G.; Meloni, F.; Garbarino, M.; Motta, R.; Ducoli, V.; Partel, P. Effects of Twenty Years of Ungulate Browsing on Forest Regeneration at Paneveggio Reserve, Italy. Forests 2020, 11, 612. [Google Scholar] [CrossRef]
- Nams, V.O.; Folkard, N.F.; Smith, J.N. Effects of nitrogen fertilization on several woody and nonwoody boreal forest species. Can. J. Bot. 1993, 71, 93–97. [Google Scholar] [CrossRef]
- Reimoser, F.; Gossow, H. Impact of ungulates on forest vegetation and its dependence on the silvicultural system. For. Ecol. Manag. 1996, 88, 107–119. [Google Scholar] [CrossRef]
- Homolka, M.; Heroldová, M. Vegetation as the food supply for game in a forest near Hostěnice. Přirodovědné Práce Ústavů Československé Akademie Věd v Brně 1990, 24, 40. [Google Scholar]
- Feuereisel, J.; Ernst, M. Verification of the food supply to game under conditions of the floodplain forest ecosystem. J. For. Sci. 2009, 55, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.S.; Hale, P.E.; Ford, W.M.; Wentworth, J.M.; French, J.R.; Anderson, O.F.; Pullen, G.B. White-tailed deer foraging in relation to successional stage, overstory type and management of southern Appalachian forests. Am. Midl. Nat. 1995, 133, 18–35. [Google Scholar] [CrossRef]
- Katreniak, J. Food Availability for Ruminants in Winter from 1st to 5th Forest Vegetation Stage. Folia Venatoria 1992, 22, 11–21. [Google Scholar]
- Bobek, B.; Merta, D.; Furtek, J. Winter food and cover refuges of large ungulates in lowland forests of south-western Poland. For. Ecol. Manag. 2016, 359, 247–255. [Google Scholar] [CrossRef]
- Storms, D.; Aubry, P.; Hamann, J.L.; Saïd, S.; Fritz, H.; Saint-Andrieux, C.; Klein, F. Seasonal variation in diet composition and similarity of sympatric red deer Cervus elaphus and roe deer Capreolus capreolus. Wildl. Biol. 2008, 14, 237–250. [Google Scholar] [CrossRef] [Green Version]
- Homolka, M. The food niches of three ungulate species in a woodland complex. Folia Zool. 1993, 42, 193–203. [Google Scholar]
- Heroldová, M. Trophic Niches of Three Ungulate Species in the Pálava Biosphere Reserve; Institute of Landscape Ecology, Academy of Sciences of the Czech Republic: Brno, Czech Republic, 1997. [Google Scholar]
- Homolka, M. Dietary analyses in the European elk (Alces alces) with a view to its prospects in southern Bohemia. Folia Zool. 1989, 38, 21–29. [Google Scholar]
- Arnold, W. Seasonal differences in the physiology of wild northern ruminants. Animal 2020, 14, s124–s132. [Google Scholar] [CrossRef] [Green Version]
- Turbill, C.; Ruf, T.; Mang, T.; Arnold, W. Regulation of heart rate and rumen temperature in red deer: Effects of season and food intake. J. Exp. Biol. 2011, 214, 963–970. [Google Scholar] [CrossRef] [Green Version]
- Green, K.; Davis, N.E.; Robinson, W.A.; McAuliffe, J.; Good, R.B. Diet selection by European hares (Lepus europaeus) in the alpine zone of the Snowy Mountains, Australia. Eur. J. Wildl. Res. 2013, 59, 693–703. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Silvicultural System | Shelterwood Cutting | Strip Cutting | Clear Cutting | Total |
|---|---|---|---|---|
| Beech | 28 | 27 | 20 | 86 |
| Spruce | 28 | 21 | 22 | 78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skoták, V.; Turek, K.; Kamler, J.; Kloz, J.; Novotná, P. Winter Food Availability for Wild Herbivores Depending on the Type of Forest Regeneration. Forests 2021, 12, 825. https://doi.org/10.3390/f12070825
Skoták V, Turek K, Kamler J, Kloz J, Novotná P. Winter Food Availability for Wild Herbivores Depending on the Type of Forest Regeneration. Forests. 2021; 12(7):825. https://doi.org/10.3390/f12070825
Chicago/Turabian StyleSkoták, Vlastimil, Kamil Turek, Jiří Kamler, Jiří Kloz, and Petra Novotná. 2021. "Winter Food Availability for Wild Herbivores Depending on the Type of Forest Regeneration" Forests 12, no. 7: 825. https://doi.org/10.3390/f12070825
APA StyleSkoták, V., Turek, K., Kamler, J., Kloz, J., & Novotná, P. (2021). Winter Food Availability for Wild Herbivores Depending on the Type of Forest Regeneration. Forests, 12(7), 825. https://doi.org/10.3390/f12070825