
Diverse Understory Vegetation Alleviates Nitrogen Competition with Crop Trees in Poplar Plantations


Abstract
1. Introduction
- (1)
- How does the presence of understory vegetation affect the growth and N utilization of poplar trees?
- (2)
- Does an increase in UVSR alleviate N competition with poplar trees and maintain poplar growth?
2. Materials and Methods
2.1. Study Site Description and Experimental Design
2.2. Tree Growth Monitoring
2.3. Poplar Leaf, Understory Vegetation, and Soil Sampling and Preparation
2.4. Measurement of Poplar Leaf, Understory Vegetation, and Soil Variables
2.5. Statistical Analyses
3. Results
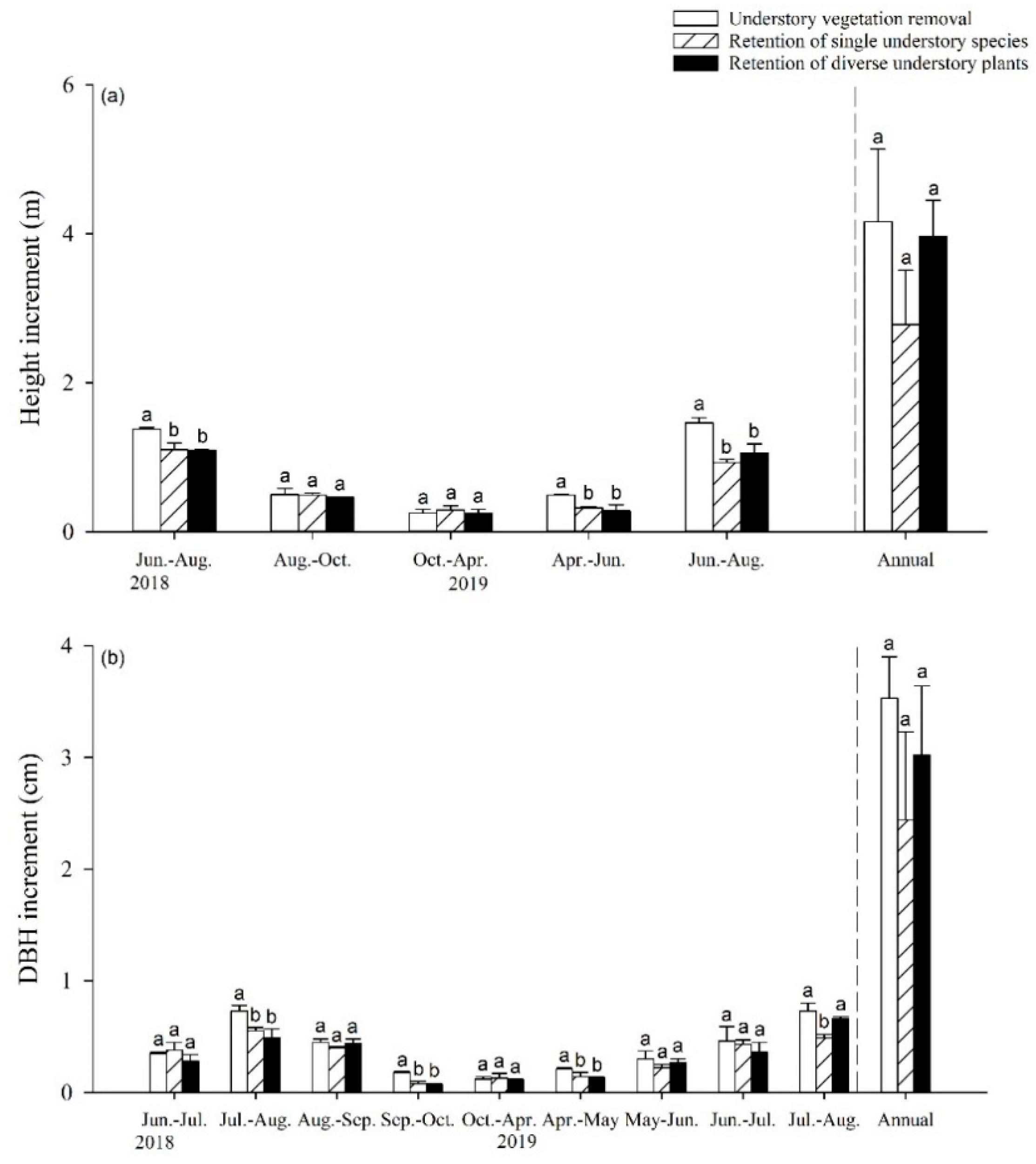
3.1. Impact of UVSR on Poplar Growth
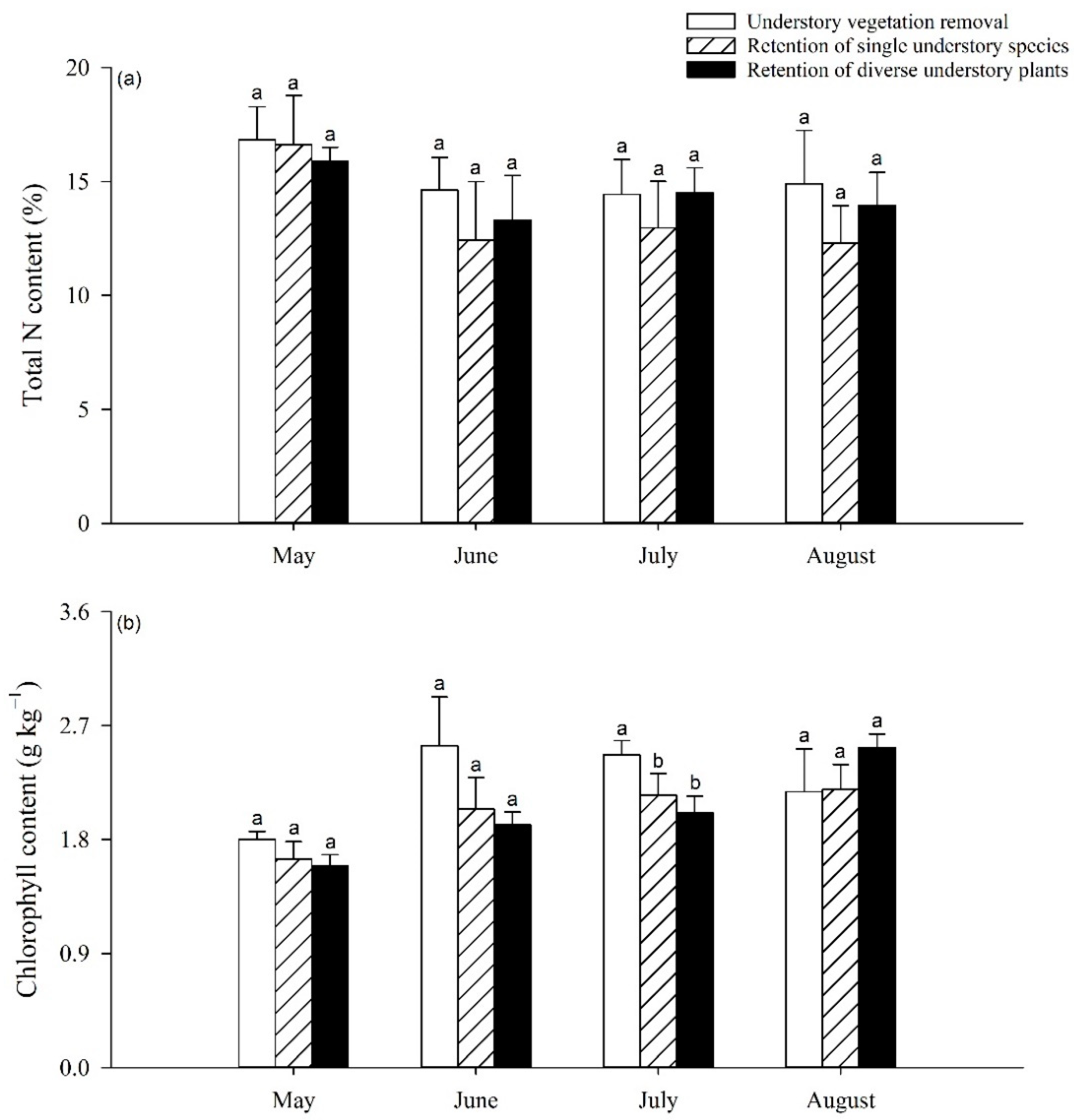
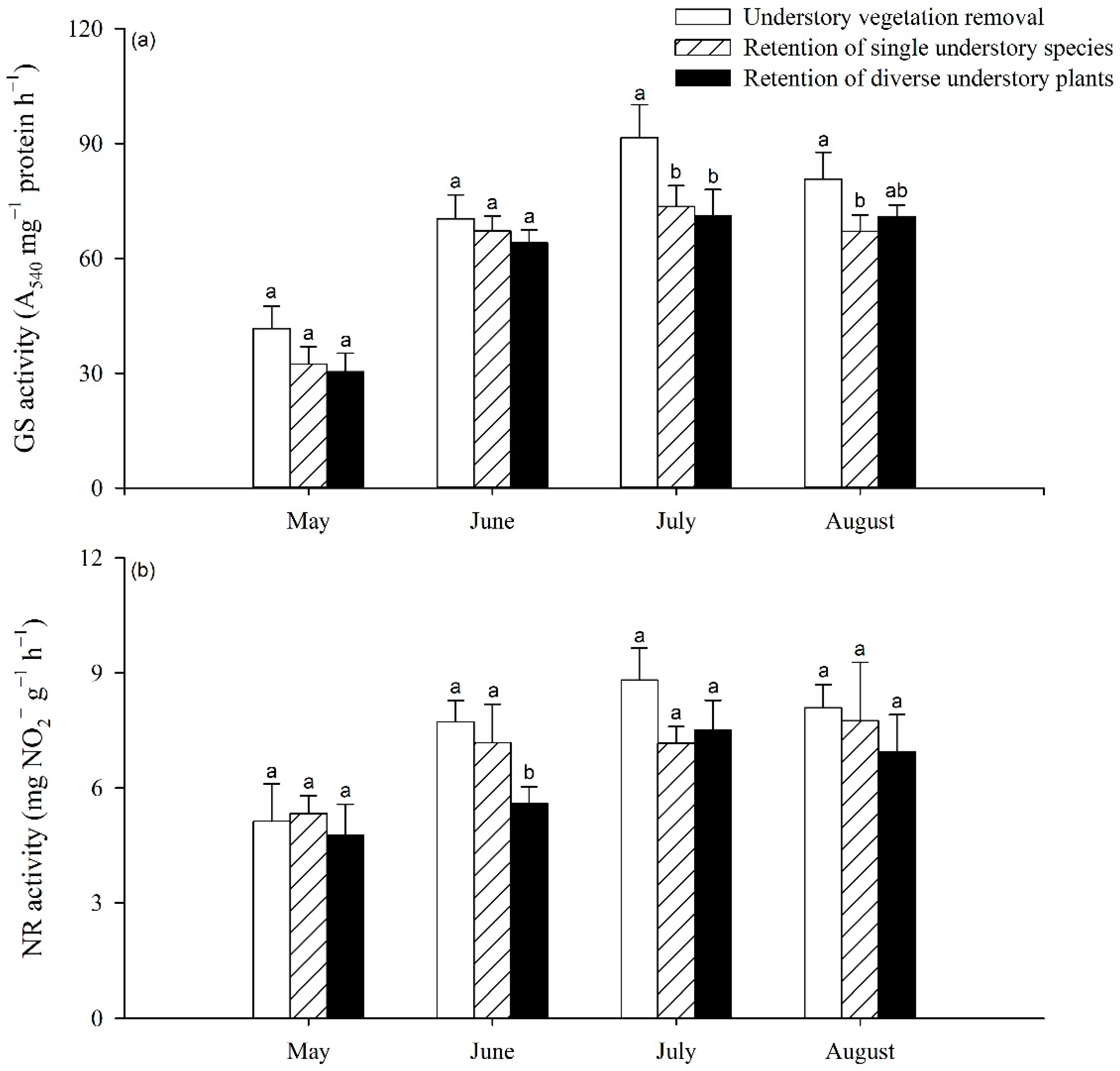
3.2. Impact of UVSR on Poplar Foliar N and Chlorophyll Content and Enzyme Activity
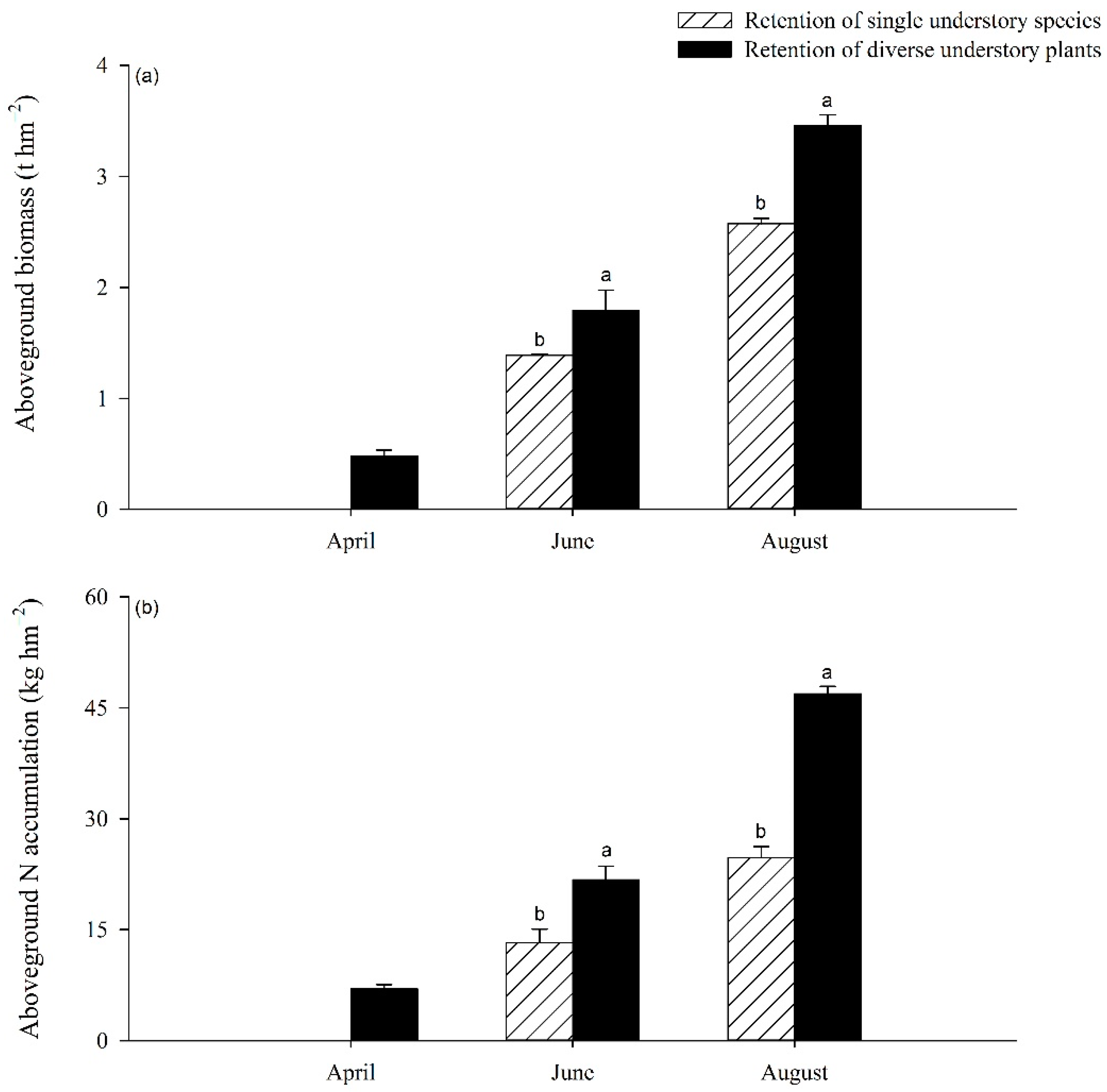
3.3. Biomass and N Accumulation in Understory Vegetation
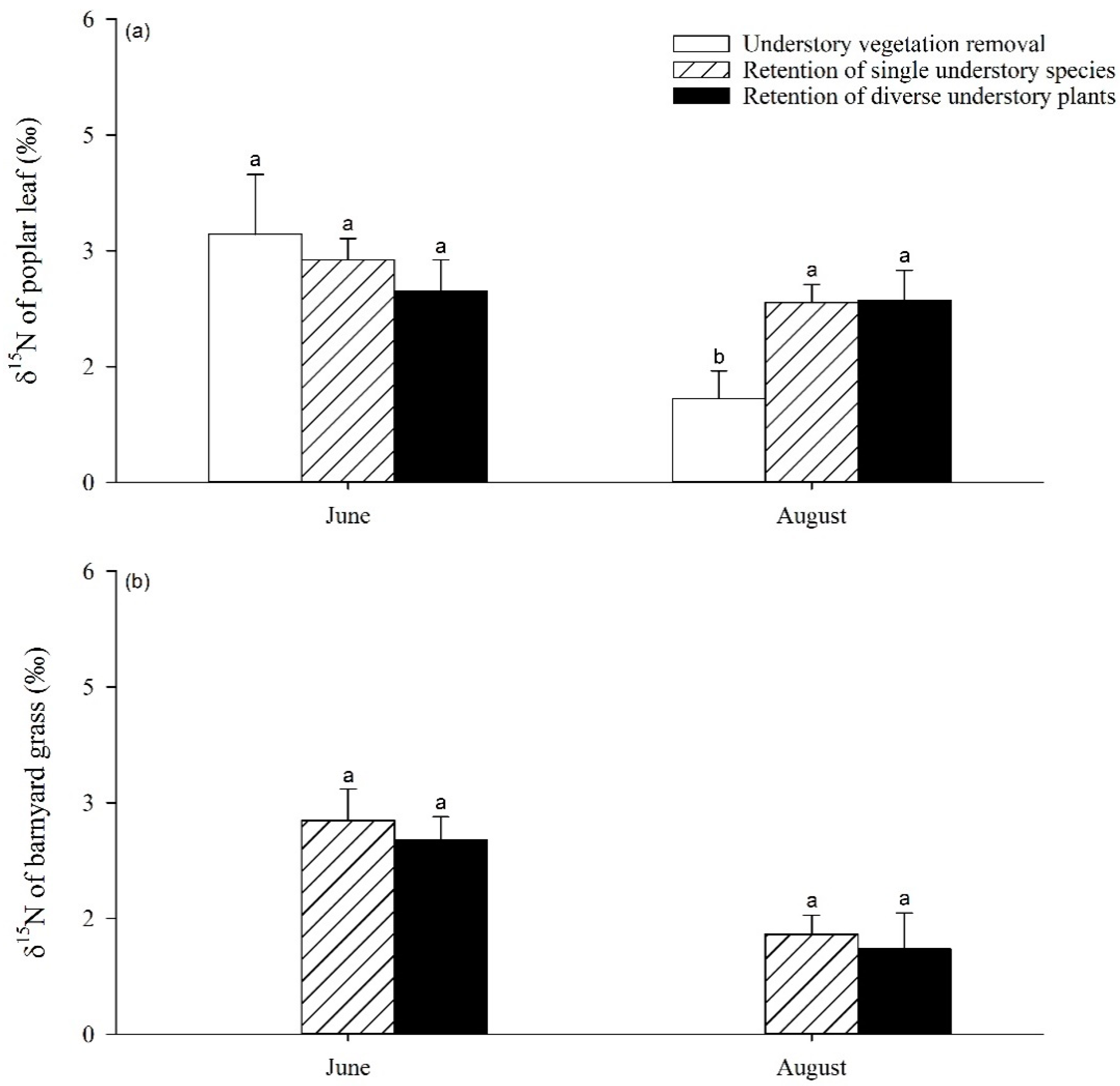
3.4. Linking δ15N of Plants and Soil Inorganic N Pool in Response to UVSR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nilsson, M.C.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Pablo, C.P.; Piotr, T.; Nicholas, C.C.; Ángela, R.L. Characterizing understory vegetation in Mediterranean forests using full-waveform airborne laser scanning data. Remote Sens. Environ. 2018, 217, 400–413. [Google Scholar]
- Gilliam, F.S. The ecological significance of the herbaceous layer in temperate forest ecosystems. BioScience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- Messier, C.; Coll, L.; Poitras-Larivière, A.P.; Jacques Brisson, N.B. Resource and non-resource root competition effects of grasses on early-versus late-successional trees. J. Ecol. 2009, 97, 548–554. [Google Scholar] [CrossRef]
- Lecerf, A.; Evangelista, C.; Cucherousset, J.; Boiché, A. Riparian overstory-understory interactions and their potential implications for forest-stream linkages. For. Ecol. Manag. 2016, 367, 112–119. [Google Scholar] [CrossRef]
- Liao, Y.C.; Fan, H.B.; Wei, X.H.; Wu, J.P.; Duan, H.L.; Fu, X.L.; Liu, W.F.; Wang, H.M.; Zhan, X.W.; Tang, P.; et al. Competition increased fine root biomass in Chinese fir (Cunninghamia lanceolata) plantations in subtropical China. For. Ecol. Manag. 2019, 435, 151–157. [Google Scholar] [CrossRef]
- Giuggiola, A.; Zweifel, R.; Feichtinger, L.; Vollenweider, P.; Bugmann, H.; Haeni, M.; Rigling, A. Competition for water in a xeric forest ecosystem-Effects of understory removal on soil micro-climate, growth and physiology of dominant Scots pine trees. For. Ecol. Manag. 2018, 409, 241–249. [Google Scholar] [CrossRef]
- Osburn, E.D.; Elliottt, K.J.; Knoepp, J.D.; Miniat, C.F.; Barrett, J.E. Soil microbial response to Rhododendron understory removal in southern Appalachian forests: Effects on extracellular enzymes. Soil Biol. Biochem. 2018, 127, 50–59. [Google Scholar] [CrossRef]
- Zhang, J.J.; Li, Y.F.; Chang, S.X.; Jiang, P.K.; Zhou, G.M.; Liu, J.; Wu, J.S.; Shen, Z.M. Understory vegetation management affected greenhouse gas emissions and labile organic carbon pools in an intensively managed Chinese chestnut plantation. Plant Soil 2014, 376, 363–375. [Google Scholar] [CrossRef]
- Gutierrez-Coarite, R.; Mollinedo, J.; Cho, A.; Wright, M.G. Canopy management of macadamia trees and understory plant diversification to reduce macadamia felted coccid (Eriococcus ironsidei) populations. Crop Prot. 2018, 113, 75–83. [Google Scholar] [CrossRef]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef] [PubMed]
- Wardle, D.A.; Zackrisson, O. Effects of species and functional group loss on island ecosystem properties. Nature 2005, 435, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P. Benefits of plant diversity to ecosystem: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Díaz, S.; Lavorel, S.; de Bello, F.; Quétier, F.; Grigulis, K.; Robson, M. Incorporating plant functional diversity effects in ecosystem service assessments. Proc. Natl. Acad. Sci. USA 2007, 104, 20684–20689. [Google Scholar] [CrossRef]
- Swenson, N.G. Plant functional diversity: Organism traits, community structure, and ecosystem properties. Ecology 2016, 97, 3556–3558. [Google Scholar] [CrossRef]
- Elliott, K.J.; Vose, J.M.; Knoepp, J.D.; Clinton, B.D.; Kloeppel, B.D. Functional role of the herbaceous layer in eastern deciduous forest ecosystems. Ecosystems 2015, 18, 221–236. [Google Scholar] [CrossRef]
- Potvin, C.; Gotelli, N.J. Biodiversity enhances individual performance but does not affect survivorship in tropical trees. Ecol. Lett. 2008, 11, 217–223. [Google Scholar] [CrossRef]
- Archaux, F.; Chevalier, R.; Berthelot, A. Towards practices favourable to plant diversity in hybrid poplar plantations. For. Ecol. Manag. 2010, 259, 2410–2417. [Google Scholar] [CrossRef]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: From temperate to boreal forests. Glob. Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, Y.; Castro-Izaguirre, N.; Baruffol, M.; Brezzi, M.; Lang, A.N.; Li, Y.; Hardtle, W.; Oheimb, G.; Yang, X.; et al. Impacts of species richness on productivity in a large-scale subtropical forest experiment. Science 2018, 362, 80–83. [Google Scholar] [CrossRef]
- McEwan, R.W.; Muller, R.N. Dynamics, diversity, and resource gradient relationships in the herbaceous layer of an old-growth Appalachian forest. Plant Ecol. 2011, 212, 1179–1191. [Google Scholar] [CrossRef]
- Chomel, M.; Guittonny-Larchevêque, M.; DesRochers, A.; Baldy, V. Effect of mixing herbaceous litter with tree litters on decomposition and N release in boreal plantations. Plant Soil 2016, 398, 229–241. [Google Scholar] [CrossRef]
- Santonja, M.; Rancon, A.; Fromin, N.; Baldy, V.; Hättenschwiler, S.; Fernandez, C.; Montès, N.; Mirleau, P. Plant litter diversity increases microbial abundance, fungal diversity, and carbon and nitrogen cycling in a Mediterranean shrubland. Soil Biol. Biochem. 2017, 111, 124–134. [Google Scholar] [CrossRef]
- Gavinet, J.; Prévosto, B.; Fernandez, C. Do shrubs facilitate oak seedling establishment in Mediterranean pine forest understory? For. Ecol. Manag. 2016, 381, 289–296. [Google Scholar] [CrossRef]
- Pennekamp, F.; Pontarp, M.; Tabi, A.; Altermatt, F.; Alther, R.; Choffat, Y.; Fronhofer, E.A.; Ganesanandamoorthy, P.; Garnier, A.; Griffiths, J.I.; et al. Biodiversity increases and decreases ecosystem stability. Nature 2018, 563, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Kawarasaki, S.; Hori, Y. Flowering phenology of understory herbaceous species in a cool temperate deciduous forest in Ogawa forest reserve, central Japan. J. Plant Res. 2001, 114, 19–23. [Google Scholar] [CrossRef]
- Gherardi, L.A.; Sala, O.E.; Yahdjian, L. Preference for different inorganic nitrogen forms among plant functional types and species of the Patagonian steppe. Oecologia 2013, 173, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved-A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- Matonis, M.S.; Binkley, D. Not just about the trees: Key role of mosaic-meadows in restoration of ponderosa pine ecosystems. For. Ecol. Manag. 2018, 411, 120–131. [Google Scholar] [CrossRef]
- Gallart, M.; Love, J.; Meason, D.F.; Coker, G.; Clinton, P.W.; Xue, J.M.; Jameson, P.E.; Klápště, J.; Turnbull, M.H. Field-scale variability in site conditions explain phenotypic plasticity in response to nitrogen source in Pinus radiata D. Don. Plant Soil 2019, 443, 353–368. [Google Scholar] [CrossRef]
- Tateno, R.; Nakayama, M.; Yano, M.; Fukuzawa, K.; Inagaki, Y.; Koba, K.; Ugawa, S. Nitrogen source utilization in co-existing canopy tree and dwarf bamboo in a northern hardwood forest in Japan. TreesStruct. Funct. 2020, 34, 1047–1057. [Google Scholar] [CrossRef]
- Qiao, Y.F.; Miao, S.J.; Silva, L.C.R.; Horwath, W.R. Understory species regulate litter decomposition and accumulation of C and N in forest soils: A long-term dual-isotope experiment. For. Ecol. Manag. 2014, 329, 318–327. [Google Scholar] [CrossRef]
- Ge, X.M.; Tian, Y.; Tang, L.Z. Nutrient distribution indicated whole-tree harvesting as a possible factor restricting the sustainable productivity of a poplar plantation system in China. PLoS ONE 2015, 10, e0125303. [Google Scholar] [CrossRef]
- Sui, J.; Ji, C.; Wang, X.; Liu, Z.; Sa, R.; Hu, Y.; Wang, C.; Li, Q.; Liu, X. A plant growth-promoting bacterium alters the microbial community of continuous cropping poplar trees’ rhizosphere. J. Appl. Microbiol. 2019, 126, 1209–1220. [Google Scholar] [CrossRef]
- Ventura, M.; Panzacchi, P.; Muzzi, E.; Magnani, F.; Tonon, G. Carbon balance and soil carbon input in a poplar short rotation coppice plantation as affected by nitrogen and wood ash application. New For. 2019, 50, 969–990. [Google Scholar] [CrossRef]
- Tanaka-Oda, A.; Kenzo, T.; Inoue, Y.; Yano, M.; Koba, K.; Ichie, T. Variation in leaf and soil 15N in diverse tree species in a lowland dipterocarp rainforest, Malaysia. Trees Struct. Funct. 2016, 30, 509–522. [Google Scholar] [CrossRef]
- Debiasi, T.V.; Calzavara, A.K.; da Silva, L.M.I.; da Silva, J.G.; Bianchini, E.; Pimenta, J.A.; Stolf-Moreira, R.; Aidar, M.P.M.; Sodek, L.; Oliveira, H.C. Nitrogen metabolism of Neotropical tree seedlings with contrasting ecological characteristics. Acta. Physiol. Plant. 2019, 41, 131. [Google Scholar] [CrossRef]
- Muruganandam, S.; Israel, D.W.; Robarge, W.P. Nitrogen transformations and microbial communities in soil aggregates from three tillage systems. Soil Sci. Soc. Am. J. 2010, 74, 120–129. [Google Scholar] [CrossRef]
- Craine, J.M.; Brookshire, E.N.J.; Cramer, M.D.; Hasselquist, N.J.; Koba, K.; Marin-Spiotta, E.; Wang, L.X. Ecological interpretations of nitrogen isotope ratios of terrestrial plants and soils. Plant Soil 2015, 396, 1–26. [Google Scholar] [CrossRef]
- Philben, M.; Billings, S.A.; Edwards, K.A.; Podrebarac, F.A.; van Biesen, G.; Ziegler, S.E. Amino acid delta 15N indicates lack of N isotope fractionation during soil organic nitrogen decomposition. Biogeochemistry 2018, 138, 69–83. [Google Scholar] [CrossRef]
- Ohashi, Y.; Sahri, M.H.; Yoshizawa, N.; Itoh, T. Annual rhythm of xylem growth in rubberwood (Hevea brasiliensis) trees grown in Malaysia. Holzforschung 2001, 55, 151–154. [Google Scholar] [CrossRef]
- Just, M.G.; Frank, S.D. Evaluation of an easy-to-install, low-cost dendrometer band for citizen-science tree research. J. For. 2019, 117, 317–322. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Gitelson, A.; Lang, M. Non-destructive determination of chlorophyll content of leaves of a green and an aurea mutant of tobacco by reflectance measurements. Plant Physiol. 1996, 148, 483–493. [Google Scholar] [CrossRef]
- Aldarini, N.; Alhasawi, A.A.; Thomas, S.C.; Appanna, V.D. The role of glutamine synthetase in energy production and glutamine metabolism during oxidative stress. Antonie Leeuwenhoek 2017, 110, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.J.; Vale, D.; Cunha, L.; Melo, P. Role of the C-terminal extension peptide of plastid located glutamine synthetase from Medicago truncatula: Crucial for enzyme activity and needless for protein import into the plastids. Plant Physiol. Biochem. 2017, 111, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Radin, J.W. In vivo assay of nitrate reductase in cotton leaf discs. Plant Physiol. 1973, 51, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Maevskaya, S.N.; Egorova, E.A.; Bukhov, N.G. Effect of elevated temperature on nitrite and nitrate reduction in leaves and intact chloroplasts. Russ. J. Plant Physiol. 2003, 50, 599–603. [Google Scholar] [CrossRef]
- Gao, W.; Yang, J.; Ren, S.R.; Liu, H.L. The trend of soil organic carbon, total nitrogen, and wheat and maize productivity under different long-term fertilizations in the upland fluvo-aquic soil of North China. Nutr. Cycl. Agroecosyst. 2015, 103, 61–73. [Google Scholar] [CrossRef]
- Norman, R.J.; Edberg, J.C.; Stucki, J.W. Determination of nitrate in soil extracts by dual-wavelength ultraviolet spectrophotometry. Soil Sci. Soc. Am. J. 1985, 49, 1182–1185. [Google Scholar] [CrossRef]
- Griesheim, K.L.; Mulvaney, R.L. Improving the accuracy of diffusion for inorganic 15N analyses of soil extracts and water. Commun. Soil Sci. Plant Anal. 2019, 50, 1161–1169. [Google Scholar] [CrossRef]
- Matsushima, M.; Choi, W.J.; Chang, S.X. White spruce foliar δ13C and δ15N indicate changed soil N availability by understory removal and N fertilization in a 13-year-old boreal plantation. Plant Soil 2012, 361, 375–384. [Google Scholar] [CrossRef]
- Littke, K.M.; Harrington, T.B.; Slesak, R.A.; Holub, S.M.; Hatten, J.A.; Gallo, A.C.; Littke, W.R.; Harrison, R.B.; Turnblom, E.C. Impacts of organic matter removal and vegetation control on nutrition and growth of Douglas-fir at three Pacific Northwestern Long-Term Soil Productivity sites. For. Ecol. Manag. 2020, 468, 118176. [Google Scholar] [CrossRef]
- Dubbert, M.; Piayda, A.; Cuntz, M.; Correia, A.C.; Silva, F.C.E.; Pereira, J.S.; Werner, C. Stable oxygen isotope and flux partitioning demonstrates understory of an oak savanna contributes up to half of ecosystem carbon and water exchange. Front. Plant Sci. 2014, 5, 530. [Google Scholar] [CrossRef]
- Mazzochini, G.G.; Camargo, J.L.C. Understory plant interactions along a successional gradient in central Amazon. Plant Soil 2020, 450, 81–92. [Google Scholar] [CrossRef]
- Gilliam, F. The Herbaceous Layer in Forests of Eastern North America; Oxford Scholarship Online; Oxford University: Oxford, UK, 2014; pp. 1–24. [Google Scholar]
- De Long, J.R.; Dorrepaal, E.; Kardol, P.; Nilsson, M.C.; Teuber, L.M.; Wardle, D.A. Understory plant functional groups and litter species identity are stronger drivers of litter decomposition than warming along a boreal forest post-fire successional gradient. Soil Biol. Biochem. 2016, 98, 159–170. [Google Scholar] [CrossRef]
- Trentini, C.P.; Villagra, M.; Pámies, D.G.; Laborde, V.B.; Bedano, J.C.; Campanello, P.I. Effect of nitrogen addition and litter removal on understory vegetation, soil mesofauna, and litter decomposition in loblolly pine plantations in subtropical Argentina. For. Ecol. Manag. 2018, 429, 133–142. [Google Scholar] [CrossRef]
- Wardle, D.A.; Nilsson, M.C.; Zackrisson, O.; Gallet, C. Determinants of litter mixing effects in a Swedish boreal forest. Soil Biol. Biochem. 2003, 35, 827–835. [Google Scholar] [CrossRef]
- Pei, Z.Q.; Leppert, K.N.; Eichenberg, D.; Bruelheide, H.; Niklaus, P.A.; Buscot, F.; Gutknecht, J.L.M. Leaf litter diversity alters microbial activity, microbial abundances, and nutrient cycling in a subtropical forest ecosystem. Biogeochemistry 2017, 134, 163–181. [Google Scholar] [CrossRef]
- Zimonick, B.J.; Simard, S.W.; Roach, W.J. Selective removal of paper birch increases growth of juvenile Douglas-fir while minimizing impacts on the plant community. Scand. J. For. Res. 2017, 32, 708–716. [Google Scholar] [CrossRef]
- Otsing, E.; Barantal, S.; Anslan, S.; Koricheva, J.; Tedersoo, L. Litter species richness and composition effects on fungal richness and community structure in decomposing foliar and root litter. Soil Biol. Biochem. 2018, 125, 328–339. [Google Scholar] [CrossRef]
- Xiao, W.Y.; Chen, H.Y.H.; Kumar, P.; Chen, C.; Guan, Q.W. Multiple interactions between tree composition and diversity and microbial diversity underly litter decomposition. Geoderma 2019, 341, 161–171. [Google Scholar] [CrossRef]
- Jones, A.R.; Dalal, R.C. Enrichment of natural 15N abundance during soil N losses under 20 years of continuous cereal cropping. Sci. Total Environ. 2017, 574, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Li, F.L.; McCormack, M.L.; Liu, X.; Hu, H.; Feng, D.F.; Bao, W.K. Vertical fine-root distributions in five subalpine forest types shifts with soil properties across environmental gradients. Plant Soil 2020, 456, 129–143. [Google Scholar] [CrossRef]
- Zhang, C.; Stratópoulos, L.M.F.; Xu, C.; Pretzsch, H.; Rötzer, T. Development of fine root biomass of two contrasting urban tree cultivars in response to drought stress. Forests 2020, 11, 108. [Google Scholar] [CrossRef]
- Zhang, H.Q.; Zhao, X.Q.; Chen, Y.L.; Zhang, L.Y.; Shen, R.F. Case of a stronger capability of maize seedlings to use ammonium being responsible for the higher 15N recovery efficiency of ammonium compared with nitrate. Plant Soil 2019, 440, 293–309. [Google Scholar] [CrossRef]
- Iqbal, A.; Dong, Q.; Wang, X.R.; Gui, H.P.; Zhang, H.H.; Pang, N.C.; Zhang, X.L.; Song, M.Z. Nitrogen preference and genetic variation of cotton genotypes for nitrogen use efficiency. J. Sci. Food Agric. 2020, 100, 2761–2773. [Google Scholar] [CrossRef]
- Veres, Z.; Kotroczó, Z.; Fekete, I.; Tóth, J.A.; Lajtha, K.; Townsend, K.; Tóthmérész, B. Soil extracellular enzyme activities are sensitive indicators of detrital inputs and carbon availability. Appl. Soil Ecol. 2015, 92, 18–23. [Google Scholar] [CrossRef]
- Yan, H.M.; Yang, F.; Gao, J.M.; Peng, Z.H.; Chen, W.M. Subsoil microbial community responses to air exposure and legume growth depend on soil properties across different depths. Sci. Rep. 2019, 9, 18536. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Z.; Bao, X.L.; Yang, Y.L.; Zhao, Y.; Liang, C.; Xie, H.T. Comparison of soil phosphorus and phosphatase activity under long-term no-tillage and maize residue management. Plant Soil Environ. 2019, 65, 408–415. [Google Scholar] [CrossRef]
- Barel, J.M.; Kuyper, T.W.; de Boer, W.; De Deyn, G.B. Plant presence reduces root and shoot litter decomposition rates of crops and wild relatives. Plant Soil 2019, 438, 313–327. [Google Scholar] [CrossRef]
- Chen, F.S.; Wang, G.G.; Fang, X.M.; Wan, S.Z.; Zhang, Y.; Liang, C. Nitrogen deposition effect on forest litter decomposition is interactively regulated by endogenous litter quality and exogenous resource supply. Plant Soil 2019, 437, 413–426. [Google Scholar] [CrossRef]
- Midwood, A.J.; Hannam, K.D.; Forge, T.A.; Neilsen, D.; Emde, D.; Jones, M.D. Importance of drive-row vegetation for soil carbon storage in woody perennial crops: A regional study. Geoderma 2020, 377, 114591. [Google Scholar] [CrossRef]
- Rivaie, A.A. The effects of understory vegetation on P availability in Pinus radiata forest stands: A review. J. For. Res. 2014, 25, 489–500. [Google Scholar] [CrossRef]
- Kitagawa, R.; Ueno, M.; Masaki, T. Initial effects of thinning and concomitant disturbance on the understory woody community in Japanese cedar plantation. J. For. Res. 2018, 23, 120–128. [Google Scholar] [CrossRef]
- Casals, P.; Valor, T.; Besalu, A.; Molina-Terren, D. Understory fuel load and structure eight to nine years after prescribed burning in Mediterranean pine forests. For. Ecol. Manag. 2016, 362, 156–168. [Google Scholar] [CrossRef]
- Dagan, U.; Izhaki, I. Understory vegetation in planted pine forests governs bird community composition and diversity in the eastern Mediterranean region. For. Ecosyst. 2019, 6, 29. [Google Scholar] [CrossRef]
- Duguid, M.C.; Frey, B.R.; Ellum, D.S.; Kelty, M.; Ashton, M.S. The influence of ground disturbance and gap position on understory plant diversity in upland forests of southern New England. For. Ecol. Manag. 2013, 303, 148–159. [Google Scholar] [CrossRef]
- De Stefano, A.; Blazier, M.A.; Comer, C.E.; Dean, T.J.; Wigley, T.B. Understory vegetation richness and diversity of Eucalyptus benthamii and Pinus elliottii plantations in the mid-south US. For. Sci. 2020, 66, 66–81. [Google Scholar]
- Grime, J.; Mackey, J.M.L.; Hillier, S.H.; Read, D.J. Floristic diversity in a model system using experimental microcosms. Nature 1987, 328, 420–422. [Google Scholar] [CrossRef]
- Hansson, M.; Fogelfors, H. Management of a semi-natural grassland; results from a 15-year-old experiment in southern Sweden. J. Veg. Sci. 2000, 11, 31–38. [Google Scholar] [CrossRef]
- Antonsen, H.; Olsson, P. Relative importance of burning, mowing and species translocation in the restoration of a former boreal hayfield: Responses of plant diversity and the microbial community. J. Appl. Ecol. 2005, 42, 337–347. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| June | August | ||||||
|---|---|---|---|---|---|---|---|
| Soil Layer (0–5 cm) | Soil Layer (5–10 cm) | Soil Layer (10–20 cm) | Soil Layer (0–5 cm) | Soil Layer (5–10 cm) | Soil Layer (10–20 cm) | ||
| NH4+-N content (mg kg−1) | UR | 2.23 ± 0.22 b | 1.39 ± 0.22 b | 1.06 ± 0.18 b | 2.05 ± 0.28 a | 2.01 ± 0.28 a | 1.79 ± 0.26 a |
| RS | 4.34 ± 0.45 a | 4.60 ± 1.06 a | 2.73 ± 0.55 a | 2.46 ± 0.62 a | 2.89 ± 0.49 a | 1.47 ± 0.06 a | |
| RD | 3.97 ± 1.04 a | 3.81 ± 0.65 a | 1.77 ± 0.30 b | 2.43 ± 0.40 a | 2.11 ± 0.43 a | 1.61 ± 0.12 a | |
| NO3−-N content (mg kg−1) | UR | 26.17 ± 5.67 a | 24.73 ± 1.86 a | 15.35 ± 3.28 a | 11.63 ± 2.49 a | 12.32 ± 1.95 a | 6.88 ± 0.81 a |
| RS | 26.28 ± 5.26 a | 10.15 ± 1.92 b | 8.01 ± 1.59 b | 9.78 ± 1.99 a | 12.16 ± 1.66 a | 4.62 ± 0.95 b | |
| RD | 24.53 ± 4.44 a | 10.19 ± 1.40 b | 6.74 ± 1.92 b | 10.66 ± 0.67 a | 9.15 ± 1.14 a | 5.69 ± 0.34 ab | |
| δ15N of NH4+-N (‰) | UR | 4.75 ± 0.01 a | 4.75 ± 0.03 a | 6.78 ± 0.05 a | 2.62 ± 0.07 a | 2.65 ± 0.02 a | 3.25 ± 0.01 a |
| RS | 4.52 ± 0.28 ab | 4.44 ± 0.26 b | 6.56 ± 0.10 a | 2.48 ± 0.15 a | 2.47 ± 0.14 a | 3.62 ± 0.09 a | |
| RD | 4.26 ± 0.07 b | 4.30 ± 0.03 b | 6.53 ± 0.31 a | 2.57 ± 0.21 a | 2.62 ± 0.03 a | 3.48 ± 0.24 a | |
| δ15N of NO3−-N (‰) | UR | 2.71 ± 0.07 a | 2.85 ± 0.02 a | 3.69 ± 0.09 a | −3.33 ± 0.26 a | −3.46 ± 0.17 a | 0.54 ± 0.02 b |
| RS | 2.48 ± 0.03 b | 2.79 ± 0.05 a | 3.66 ± 0.18 a | −3.58 ± 0.10 a | −3.65 ± 0.03 a | 0.63 ± 0.01 a | |
| RD | 2.67 ± 0.08 a | 2.44 ± 0.17 b | 3.65 ± 0.19 a | −3.67 ± 0.07 a | −3.68 ± 0.09 a | 0.64 ± 0.01 a | |
| Variable Y | Standardized Coefficient | r2 | p Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| x1 | x2 | x3 | x4 | x5 | x6 | ||||
| June | δ15N of poplar leaf under UR treatment | - | - | 0.38 | - | - | 0.62 | 0.99 | 0.02 |
| δ15N of poplar leaf under RS treatment | - | - | 0.49 | - | - | 0.51 | 0.99 | 0.02 | |
| δ15N of poplar leaf under RD treatment | - | - | 0.59 | - | - | 0.41 | 0.99 | 0.00 | |
| δ15N of barnyard grass under RS treatment | - | - | 0.34 | - | - | 0.67 | 0.99 | 0.01 | |
| δ15N of barnyard grass under RD treatment | - | - | 0.33 | - | - | 0.67 | 0.99 | 0.00 | |
| August | δ15N of poplar leaf under UR treatment | 0.04 | - | - | - | - | 0.97 | 0.99 | 0.02 |
| δ15N of poplar leaf under RS treatment | - | - | 1.45 | - | 0.45 | - | 0.99 | 0.02 | |
| δ15N of poplar leaf under RD treatment | 1.13 | - | - | - | 0.13 | - | 0.99 | 0.02 | |
| δ15N of barnyard grass under RS treatment | - | - | - | - | 0.13 | 1.13 | 0.99 | 0.01 | |
| δ15N of barnyard grass under RD treatment | 0.02 | - | - | - | - | 0.98 | 0.99 | 0.00 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Qin, G.; Zhai, Z.; Zhou, S.; Tang, L.; Tian, Y. Diverse Understory Vegetation Alleviates Nitrogen Competition with Crop Trees in Poplar Plantations. Forests 2021, 12, 705. https://doi.org/10.3390/f12060705
Zhang J, Qin G, Zhai Z, Zhou S, Tang L, Tian Y. Diverse Understory Vegetation Alleviates Nitrogen Competition with Crop Trees in Poplar Plantations. Forests. 2021; 12(6):705. https://doi.org/10.3390/f12060705
Chicago/Turabian StyleZhang, Jiayu, Guangzhen Qin, Zheng Zhai, Shichao Zhou, Luozhong Tang, and Ye Tian. 2021. "Diverse Understory Vegetation Alleviates Nitrogen Competition with Crop Trees in Poplar Plantations" Forests 12, no. 6: 705. https://doi.org/10.3390/f12060705
APA StyleZhang, J., Qin, G., Zhai, Z., Zhou, S., Tang, L., & Tian, Y. (2021). Diverse Understory Vegetation Alleviates Nitrogen Competition with Crop Trees in Poplar Plantations. Forests, 12(6), 705. https://doi.org/10.3390/f12060705