
Morphology, Molecular Phylogeny, and Pathogenicity of Neofusicoccum parvum, Associated with Leaf Spot Disease of a New Host, the Japanese Bay Tree (Machilus thunbergii)

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Isolation
2.2. Morphology
2.3. DNA Extraction, PCR Amplification and Sequencing
2.4. Molecular Phylogeny
2.5. Pathogenicity
3. Results
3.1. Fungus Isolation and Morphological Characterization
3.2. Molecular Phylogeny
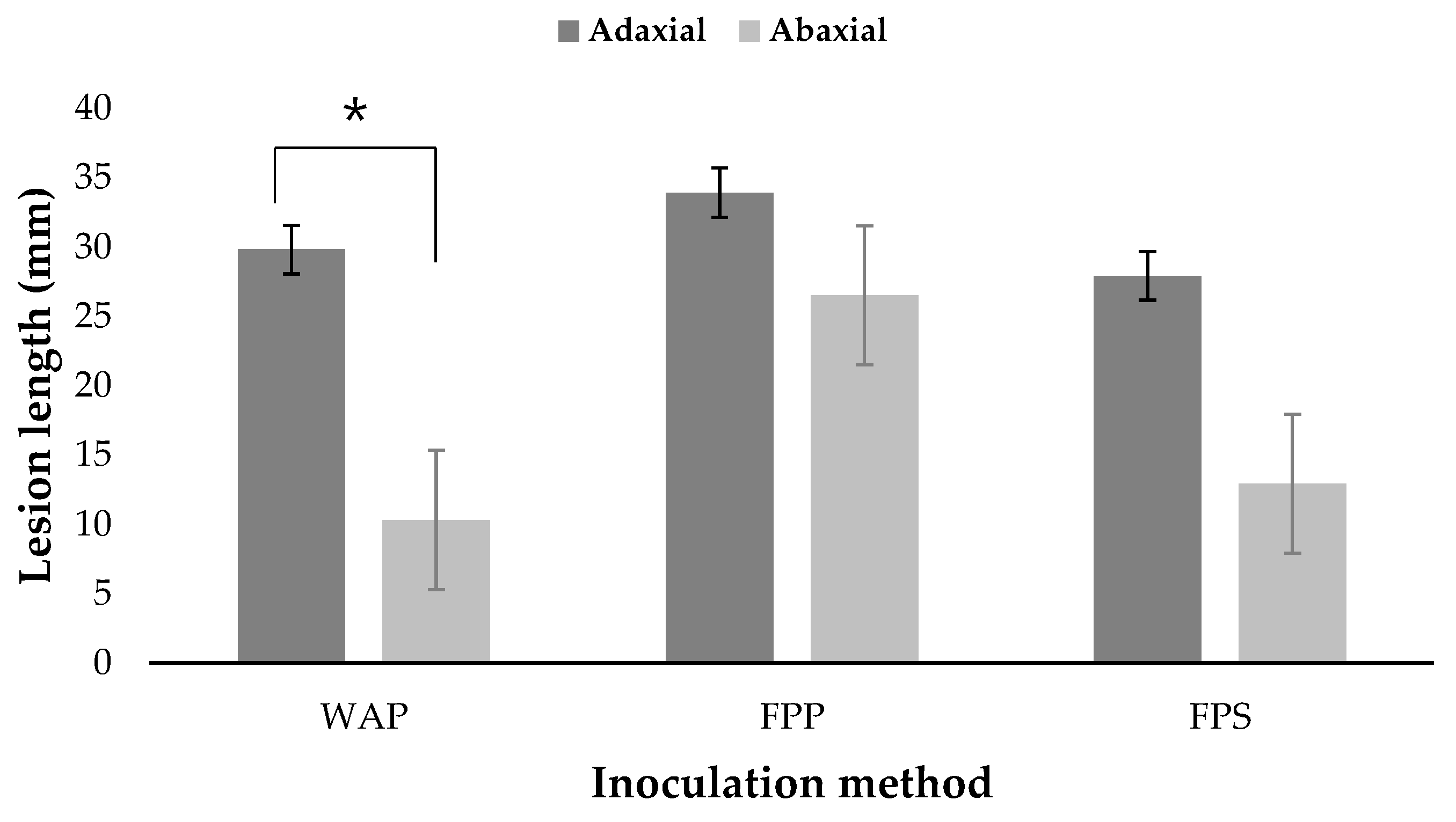
3.3. Pathogenicity Tests
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ren, Q.; Wu, D.; Wu, C.; Wang, Z.; Jiao, J.; Jiang, B.; Zhu, J.; Huang, Y.; Li, T.; Yuan, W. Modeling the potential distribution of Machilus thunbergii; under the climate change patterns in China. OJF 2020, 10, 217–231. [Google Scholar] [CrossRef]
- Wu, S.H.; Hwang, C.Y.; Lin, T.P.; Chung, J.D.; Cheng, Y.P.; Hwang, S.Y. Contrasting phylogeographical patterns of two closely related species, Machilus thunbergii and Machilus kusanoi (Lauraceae), in Taiwan. J. Biogeogr. 2006, 33, 936–947. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Raven, P.H.; Hong, D.Y. Flora of China. In Menispermaceae through Capparaceae; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Missouri Botanical Garden Press: St. Louis, MO, USA, 2008. [Google Scholar]
- Li, G.; Ju, H.K.; Chang, H.W.; Jahng, Y.; Lee, S.H.; Son, J.K. Melanin biosynthesis inhibitors from the bark of Machilus thunbergii. Biol. Pharm. Bull. 2003, 26, 1039–1041. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shimomura, H.; Sashida, Y.; Oohara, M. Lignans from Machilus thunbergii. Phytochemestry 1987, 3, 1513–1515. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, Systematic Mycology and Microbiology Laboratory; USDA: Washington, DC, USA, 2018. [Google Scholar]
- Sakalidis, M.L.; Slippers, B.; Wingfield, B.D.; Hardy, G.S.J.; Burgess, T.I. The Challenge of understanding the origin, pathways and extent of fungal invasions: Global populations of the Neofusicoccum parvum-N. ribis species complex. Divers. Distrib. 2013, 19, 873–883. [Google Scholar] [CrossRef]
- Pennycook, S.R.; Samuels, G.J. Botryosphaeria and Fusicoccum species associated with ripe fruit rot of Actinidia deliciosa (Kiwifruit) in New Zealand. Mycotaxon 1985, 24, 445–458. [Google Scholar]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Slippers, B.; Smit, W.A.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Taxonomy, phylogeny and identification of Botryosphaeriaceae associated with pome and stone fruit trees in South Africa and other regions of the world. Plant Pathol. 2007, 56, 128–139. [Google Scholar] [CrossRef]
- Espinoza, J.G.; Briceño, E.X.; Chávez, E.R.; Úrbez-Torres, J.R.; Latorre, B.A. Neofusicoccum spp. associated with stem canker and dieback of blueberry in Chile. Plant Dis. 2009, 93, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Laveau, C.; Letouze, A.; Louvet, G.; Bastien, S.; Guérin-Dubrana, L. Differential aggressiveness of fungi implicated in esca and associated diseases of grapevine in France. Phytopathol. Mediterr. 2009, 48, 15. [Google Scholar]
- Golzar, H.; Burgess, T.I. Neofusicoccum Parvum, a causal agent associated with cankers and decline of Norfolk Island pine in Australia. Australas. Plant Pathol. 2011, 40, 484–489. [Google Scholar] [CrossRef]
- Thomidis, T.; Michailides, T.J.; Exadaktylou, E. Neofusicoccum parvum associated with fruit rot and shoot blight of peaches in Greece. Eur. J. Plant Pathol. 2011, 131, 661–668. [Google Scholar] [CrossRef]
- Vakalounakis, D.J.; Ntougias, S.; Kavroulakis, N.; Protopapadakis, E. Neofusicoccum parvum and Diaporthe foeniculina associated with twig and shoot blight and branch canker of citrus in Greece. J. Phytopathol. 2019, 167, 527–537. [Google Scholar] [CrossRef]
- Chilvers, M.I.; Jones, S.; Meleca, J.; Peever, T.L.; Pethybridge, S.J.; Hay, F.S. Characterization of mating type genes supports the hypothesis that Stagonosporopsis chrysanthemi is homothallic and provides evidence that Stagonosporopsis tanaceti is heterothallic. Curr. Genet. 2014, 60, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Cubero, O.F.; Crespo, A.; Fatehi, J.; Bridge, P.D. DNA Extraction and PCR amplification method suitable for fresh, herbarium-stored, lichenized, and other fungi. Plant. Syst. Evol. 1999, 216, 243–249. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, A.M., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- O’Donnell, K. Molecular phylogeny of the Nectria haematococca-Fusarium solani species complex. Mycologia 2000, 92, 919–938. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.C.; Nam, S.S.; Kachroo, A.; Kim, Y.H.; Yang, J.W. Characterization and pathogenicity of sweet potato (Ipomoea batatas) black rot caused by Ceratocystis fimbriata in Korea. Eur. J. Plant Pathol. 2018, 152, 833–840. [Google Scholar] [CrossRef]
- Yang, J.W.; Nam, S.S.; Lee, H.U.; Choi, K.H.; Hwang, S.G.; Paul, N.C. Fusarium root rot caused by Fusarium solani on sweet potato (Ipomoea batatas) in South Korea. Can. J. Plant Pathol. 2018, 40, 90–95. [Google Scholar] [CrossRef]
- Choi, Y.P.; Paul, N.C.; Lee, H.B.; Yu, S.H. First record of Alternaria simsimi causing leaf spot on sesame (Sesamum indicum L.) in Korea. Mycobiology 2014, 42, 405–408. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [PubMed]
- Slippers, B.; Crous, P.W.; Denman, S.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Combined multiple gene genealogies and phenotypic characters differentiate several species previously identified as Botryosphaeria dothidea. Mycologia 2004, 96, 83–101. [Google Scholar] [CrossRef] [PubMed]
- Karikome, H.; Mimaki, Y.; Sashida, Y. A Butanolide and phenolics from Machilus thunbergii. Phytochemistry 1991, 30, 315–319. [Google Scholar] [CrossRef]
- Iturritxa, E.; Slippers, B.; Mesanza, N.; Wingfield, M.J. First report of Neofusicoccum parvum causing canker and die-back of eucalyptus in Spain. Australas. Plant Dis. Notes 2011, 6, 57–59. [Google Scholar] [CrossRef]
- Choi, I.Y.; Sharma, P.K.; Cheong, S.S. First report of Neofusicoccum parvum associated with bark dieback of blueberry in Korea. Plant Pathol. J. 2012, 28, 217. [Google Scholar] [CrossRef]
- Alberti, I.; Prodi, A.; Nipoti, P.; Grassi, G. First report of Neofusicoccum parvum causing stem and branch canker on Cannabis Sativa in Italy. J Plant. Dis. Prot. 2018, 125, 511–513. [Google Scholar] [CrossRef]
- Lopes, U.P.; Zambolim, L.; Pinho, D.B.; Barros, A.V.; Costa, H.; Pereira, O.L. Postharvest rot and mummification of strawberry fruits caused by Neofusicoccum parvum and N. kwambonambiense in Brazil. Trop. Plant Pathol. 2014, 39, 178–183. [Google Scholar] [CrossRef]
- Manca, D.; Bregant, C.; Maddau, L.; Pinna, C.; Montecchio, L.; Linaldeddu, B. First report of canker and dieback caused by Neofusicoccum parvum and Diplodia olivarum on oleaster in Italy. Ital. J. Mycol. 2020, 85–91. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Isolate ID | Host | Origin | Accession No. | |
|---|---|---|---|---|---|
| ITS | EF1-Alpha | ||||
| Neofusicoccum parvum | CMML20-01 * | Machilus thunbergii Sieb. & Zucc. | South Korea | MW142215 | MW142216 |
| Neofusicoccum parvum | CMML20-03 | M. thunbergii | South Korea | MW535296 | MW535298 |
| Neofusicoccum parvum | CMML20-04 | M. thunbergii | South Korea | MW535297 | MW535299 |
| Neofusicoccum umdonicola Pavlic, Slippers & M.J. Wingf. | CBS 123644 | Syzygium cordatum Hochst. ex Krauss | South Africa | KX464226 | KX464759 |
| Neofusicoccum occulatum Sakalidis & T. Burgess | CBS 128008T | Eucalyptus grandis W. Hill. hybrid | Australia | MH864743 | EU339509 |
| Neofusicoccum sinoeucalypti G.Q. Li & S.F. Chen | CERC 2005T | Eucalyptus urophylla S.T. Blake × E. grandis W. Hill. | China | KX278061 | KX278166 |
| Neofusicoccum kwambonambiense Pavlic, Slippers & M.J. Wingf. | CSF 6037 | E. urophylla S.T. Blake × E. grandis | China | MT028610 | MT028776 |
| Neofusicoccum cordaticola Pavlic, Slippers | CMW 14124 | Syzygium cordatum Hochst. ex Krauss | South Africa | EU821925 | EU821895 |
| Neofusicoccum hongkongense G.Q. Li & S.F. Chen | CERC 2973T | Araucaria cunninghamii Mudie | China | KX278052 | KX278157 |
| Neofusicoccum parvum | CBS 123652 | S. cordatum Hochst. ex Krauss | South Africa | KX464184 | KX464710 |
| Neofusicoccum parvum | CMW 9080 | Populus nigra (L.) | New Zealand | AY236942 | AY236887 |
| Neofusicoccum parvum | CERC 3503 | E. urophylla × E. grandis | China | KX278059 | KX278164 |
| Neofusicoccum andinum Mohali, Slippers & M.J. Wingf. | CBS 117,921 | Eucalyptus sp. | Venezuela | KX464152 | KX464647 |
| Neofusicoccum macroclavatum (Burgess, Barber & Hardy) | CBS 114149 | Grevillea sp. R. Br. Ex. Knight | Australia | KX464174 | KX464694 |
| Neofusicoccum stellenboschiana Tao Yang & Crous | CBS 282.70 | Arum italicum Mill. | Spain | KX464225 | KX464758 |
| Neofusicoccum mangroviorum J.A.Osorio, Jol.Roux & Z.W.de Beer | CMW 41365T | Avicennia marina (Forssk.) Vierh | South Africa | NR_147360 | KP860702 |
| Neofusicoccum cryptoaustrale Pavlic, Maleme, Slippers & M.J. Wingf. | CMW 23785T | Eucalyptus sp. L’Hér | South Africa | NR_137718 | FJ752713 |
| Neofusicoccum viticlavatum (Niekerk & Crous) Crous | CBS 123532 | Oldenburgia sp. Less. | South Africa | KX464228 | KX464761 |
| Neofusicoccum pistaciarum Tao Yang & Crous | CBS 113083T | Pistacia vera (L.) | USA | NR_147367 | KX464712 |
| Neofusicoccum ursorum Pavlic, Maleme, Slippers & M.J. Wingf. | CBS 122812 | Eucalyptus sp. L’Hér | South Africa | KX464227 | KX464760 |
| Neofusicoccum terminaliae | CBS 125264 | Terminalia sericea Burch. Ex. DC. | South Africa | GQ471805 | GQ471783 |
| Neofusicoccum buxi Crous. | CBS 113714 | Buxus sempervirens (L.) | Sweden | KX464164 | KX464677 |
| Botryosphaeria dothidea (Moug. ex Fr.) Ces. & De Not. | CBS 110302 | Vitis vinifera (L.) | Portugal | EU673174 | AY573218 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.; Paul, N.C.; Lee, K.-H.; Kim, H.-J.; Sang, H. Morphology, Molecular Phylogeny, and Pathogenicity of Neofusicoccum parvum, Associated with Leaf Spot Disease of a New Host, the Japanese Bay Tree (Machilus thunbergii). Forests 2021, 12, 440. https://doi.org/10.3390/f12040440
Choi S, Paul NC, Lee K-H, Kim H-J, Sang H. Morphology, Molecular Phylogeny, and Pathogenicity of Neofusicoccum parvum, Associated with Leaf Spot Disease of a New Host, the Japanese Bay Tree (Machilus thunbergii). Forests. 2021; 12(4):440. https://doi.org/10.3390/f12040440
Chicago/Turabian StyleChoi, Sungyu, Narayan Chandra Paul, Kye-Han Lee, Hyun-Jun Kim, and Hyunkyu Sang. 2021. "Morphology, Molecular Phylogeny, and Pathogenicity of Neofusicoccum parvum, Associated with Leaf Spot Disease of a New Host, the Japanese Bay Tree (Machilus thunbergii)" Forests 12, no. 4: 440. https://doi.org/10.3390/f12040440
APA StyleChoi, S., Paul, N. C., Lee, K.-H., Kim, H.-J., & Sang, H. (2021). Morphology, Molecular Phylogeny, and Pathogenicity of Neofusicoccum parvum, Associated with Leaf Spot Disease of a New Host, the Japanese Bay Tree (Machilus thunbergii). Forests, 12(4), 440. https://doi.org/10.3390/f12040440