
Anthropogenic Disturbances Shape Soil Capillary and Saturated Water Retention Indirectly via Plant Functional Traits and Soil Organic Carbon in Temperate Forests


 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
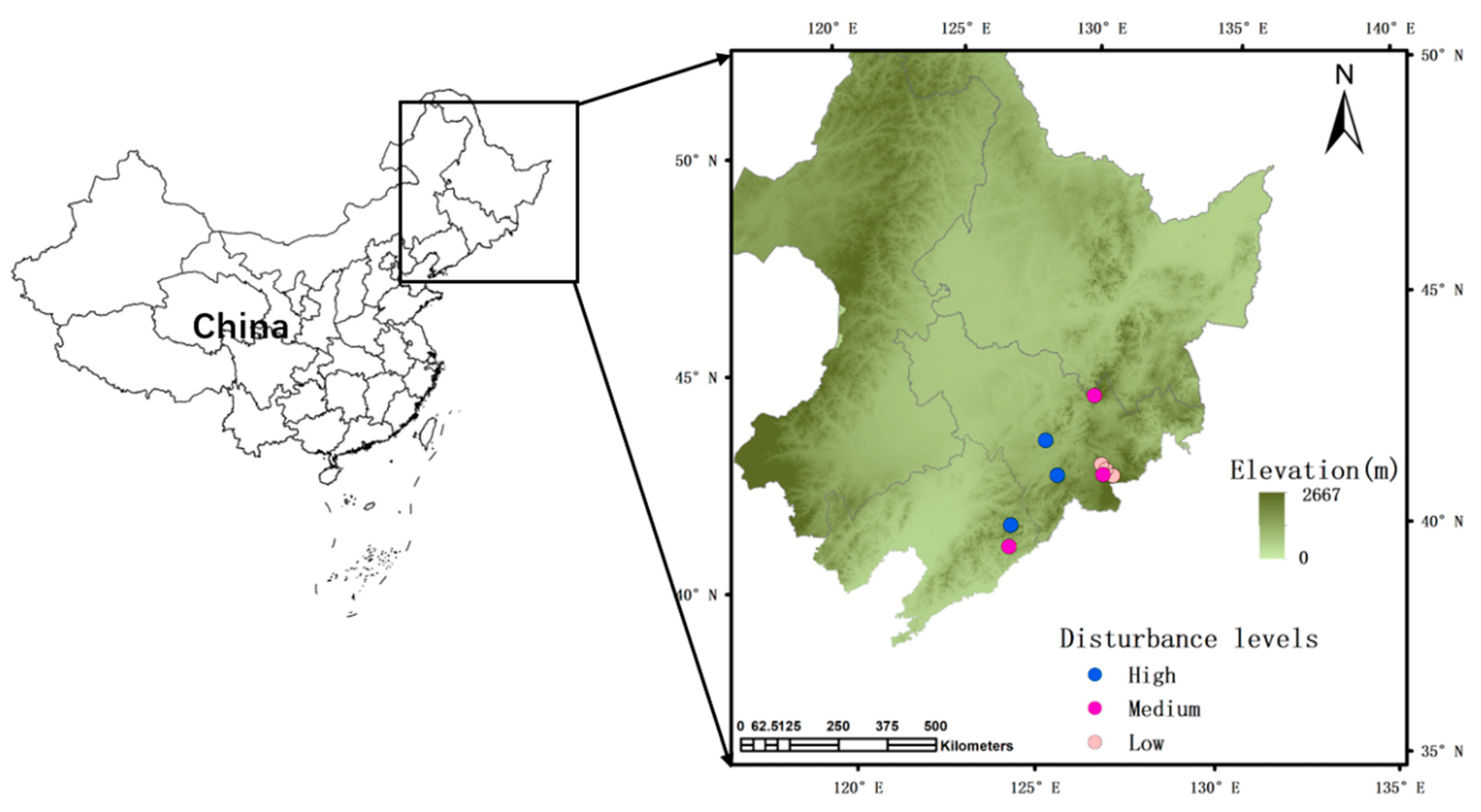
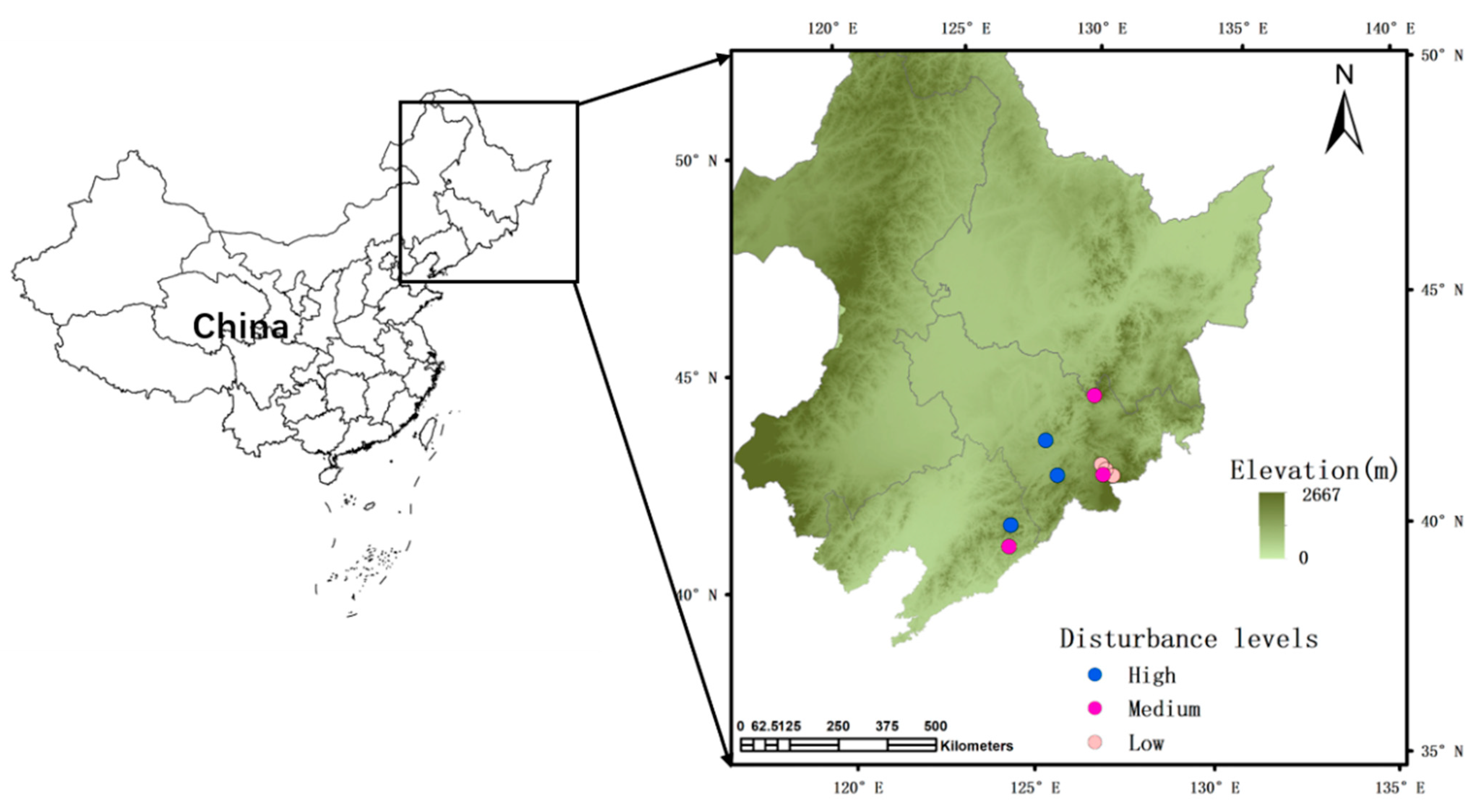
2.1. Study Sites and Forest Inventories
2.2. Assessment of Disturbance Intensities
2.3. Quantification of Plant Functional Trait Compositions and Diversity Attributes
2.4. Quantification of Soil Basic Properties and Soil Water-Physical Properties
2.5. Statistical Analyses
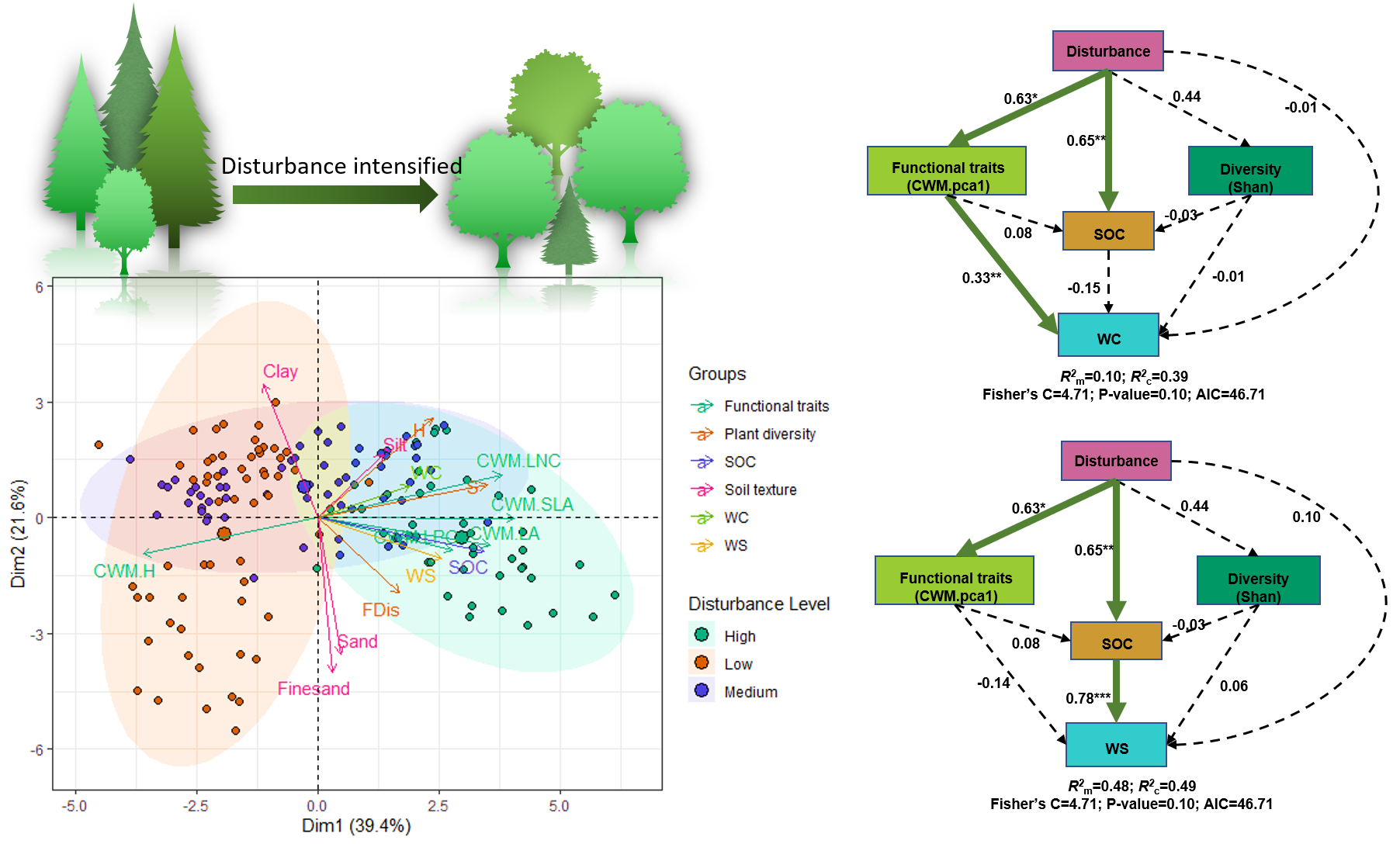
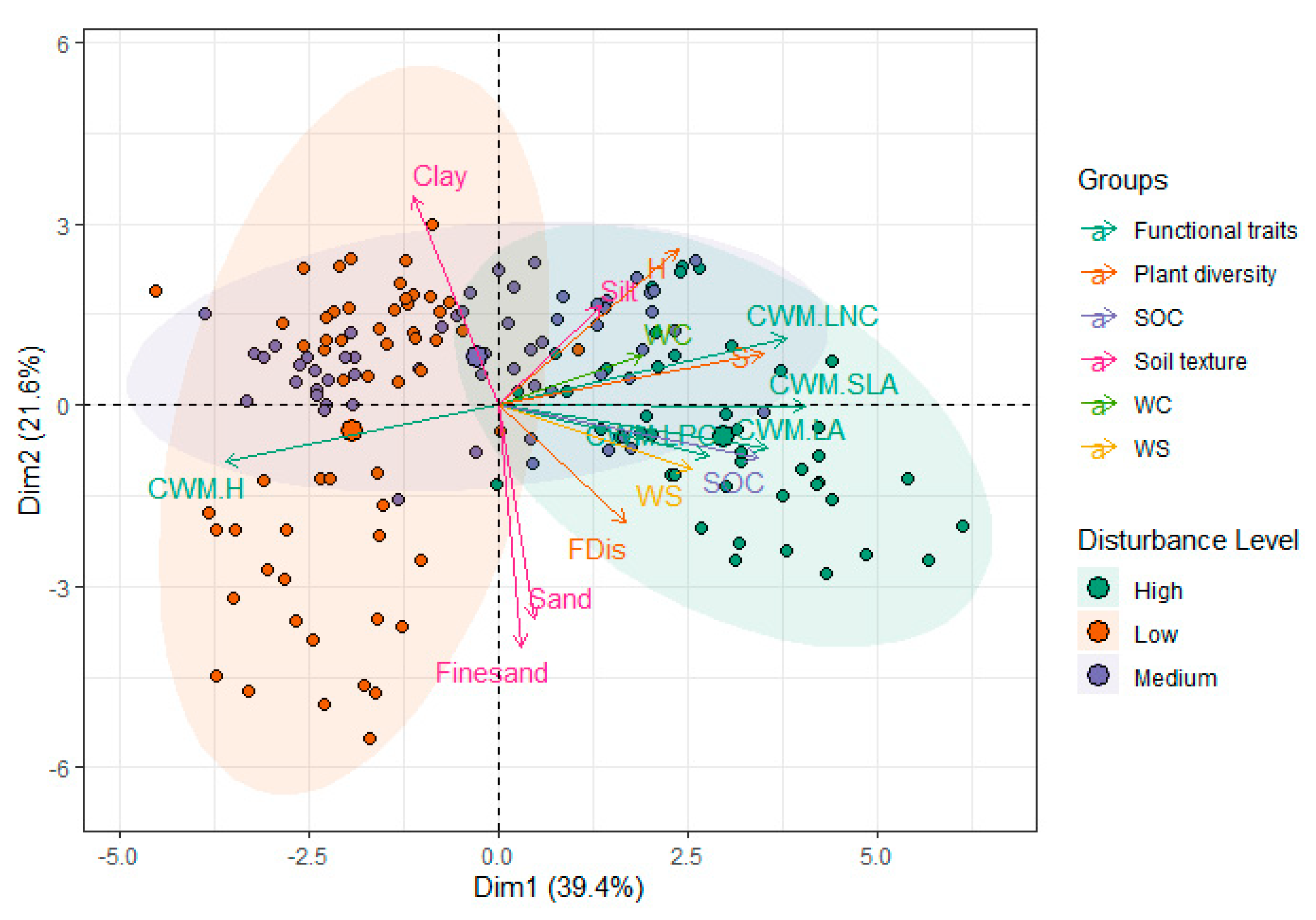
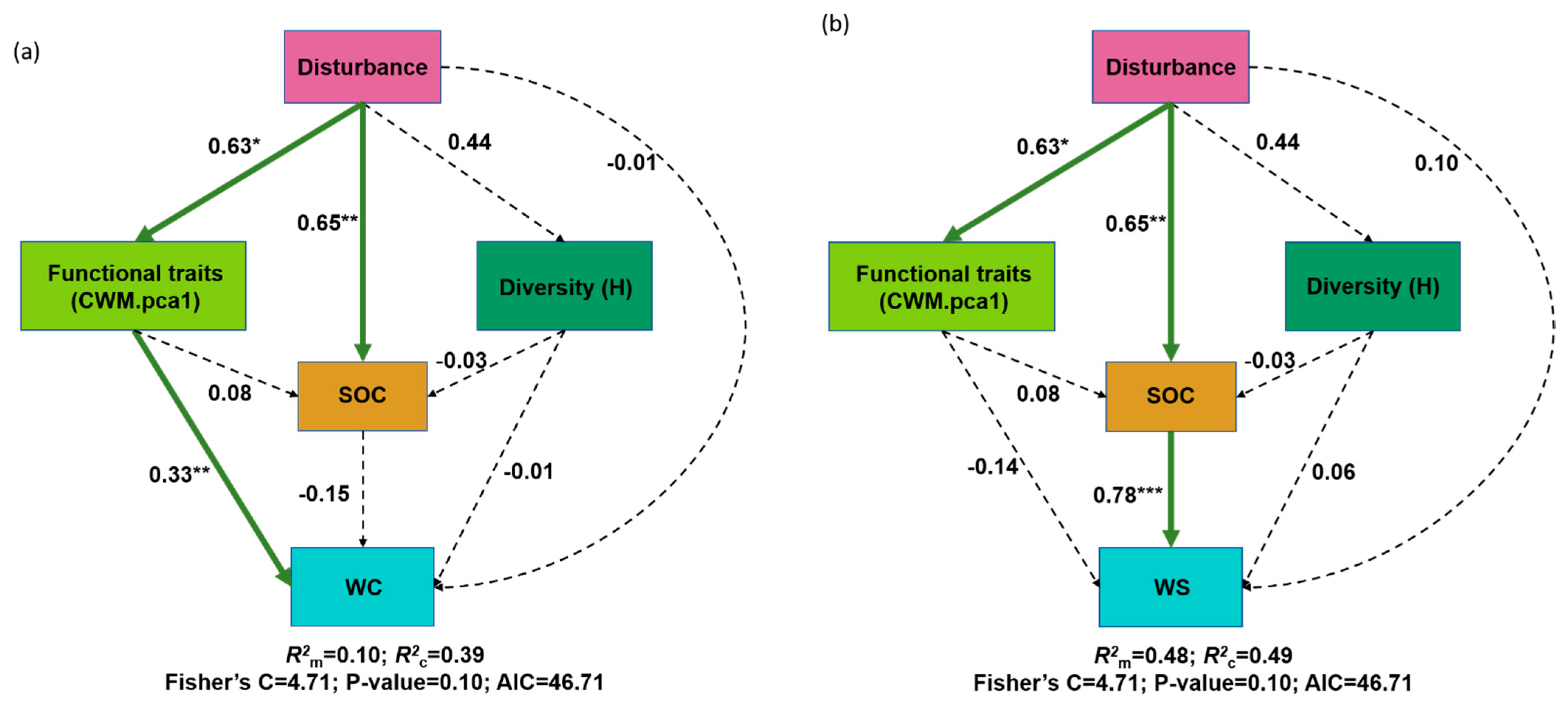
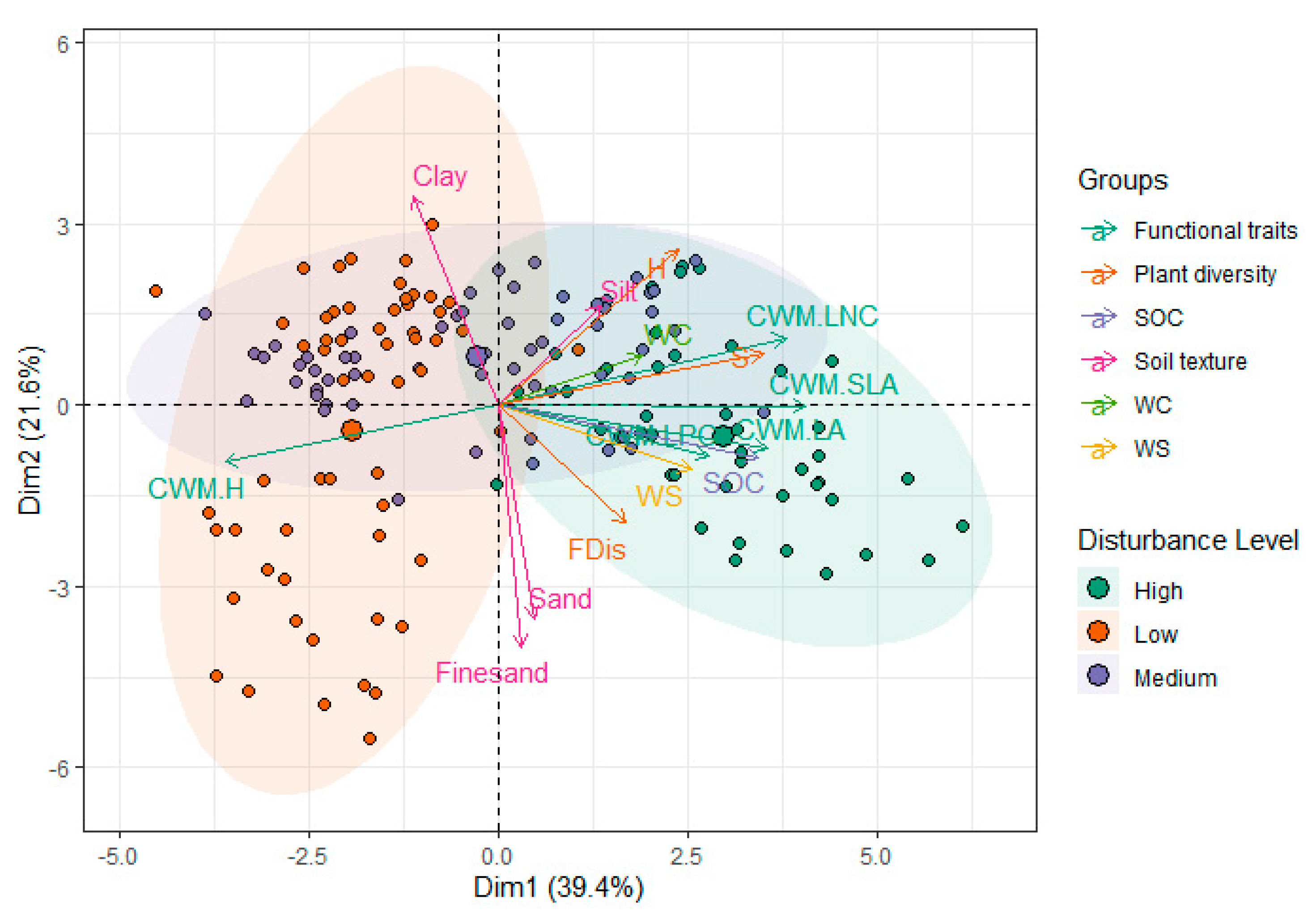
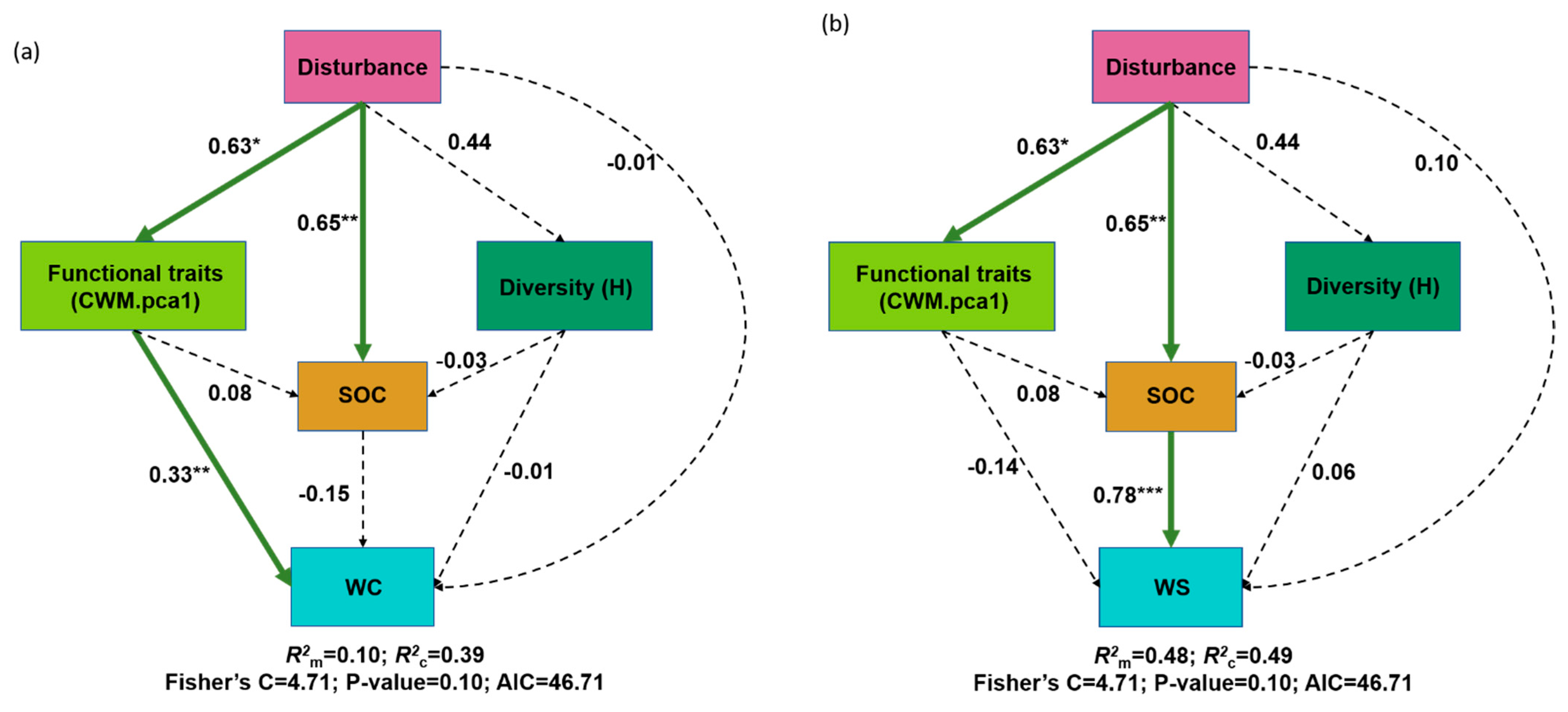
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lü, Y.H.; Hu, J.; Sun, F.X.; Zhang, L.W. Water retention and hydrological regulation: Harmony but not the same in terrestrial hydrological ecosystem services. Acta Ecol. Sin. 2015, 35, 5191–5196. [Google Scholar]
- Zhang, X.; Zhao, W.; Wang, L.; Liu, Y.; Liu, Y.; Feng, Q. Relationship between soil water content and soil particle size on typical slopes of the Loess Plateau during a drought year. Sci. Total Environ. 2019, 648, 943–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.P.; Xia, J.B.; Zhao, X.M.; Zhuge, Y.P. Soil moisture ecological characteristics of typical shrub and grass vegetation on Shell Island in the Yellow River Delta, China. Geoderma 2019, 348, 45–53. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, G.L.; Yang, J.L.; Li, D.C.; Zhao, Y.G.; Liu, F.; Yang, R.M.; Yang, F. Organic matter controls of soil water retention in an alpine grassland and its significance for hydrological processes. J. Hydrol. 2014, 519, 3086–3093. [Google Scholar] [CrossRef]
- Liu, Y.; Cui, Z.; Huang, Z.; Miao, H.-T.; Wu, G.-L. The influence of litter crusts on soil properties and hydrological processes in a sandy ecosystem. Hydrol. Earth Syst. Sci. 2019, 23, 2481–2490. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Miao, H.T.; Chang, X.F.; Wu, G.L. Higher species diversity improves soil water infiltration capacity by increasing soil organic matter content in semiarid grasslands. Land Degrad. Dev. 2019, 30, 1599–1606. [Google Scholar] [CrossRef]
- Fischer, C.; Leimer, S.; Roscher, C.; Ravenek, J.; de Kroon, H.; Kreutziger, Y.; Baade, J.; Bessler, H.; Eisenhauer, N.; Weigelt, A.; et al. Plant species richness and functional groups have different effects on soil water content in a decade-long grassland experiment. J. Ecol. 2019, 107, 127–141. [Google Scholar] [CrossRef] [Green Version]
- Saxton, K.E.; Rawls, W.J. Soil water characteristic estimates by texture and organic matter for hydrologic solutions. Soil Sci. Soc. Am. J. 2006, 70, 1569–1578. [Google Scholar] [CrossRef] [Green Version]
- Wösten, J.H.M.; Pachepsky, Y.A.; Rawls, W.J. Pedotransfer functions: Bridging the gap between available basic soil data and missing soil hydraulic characteristics. J. Hydrol. 2001, 251, 123–150. [Google Scholar] [CrossRef]
- Rawls, W.J.; Pachepsky, Y.A.; Ritchie, J.C.; Sobecki, T.M.; Bloodworth, H. Effect of soil organic carbon on soil water retention. Geoderma 2003, 116, 61–76. [Google Scholar] [CrossRef]
- Bucka, F.B.; Kölbl, A.; Uteau, D.; Peth, S.; Kögel-Knabner, I. Organic matter input determines structure development and aggregate formation in artificial soils. Geoderma 2019, 354, 113881. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, G.; Heathman, G.C.; Wang, Y.; Huang, C.-H. Fractal features of soil particle-size distribution as affected by plant communities in the forested region of Mountain Yimeng, China. Geoderma 2009, 154, 123–130. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, A.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Chen, S.; Wang, W.; Xu, W.; Wang, Y.; Wan, H.; Chen, D.; Tang, Z.; Tang, X.; Zhou, G.; Xie, Z.; et al. Plant diversity enhances productivity and soil carbon storage. Proc. Natl. Acad. Sci. USA 2018, 115, 4027–4032. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.; Tischer, J.; Roscher, C.; Eisenhauer, N.; Ravenek, J.; Gleixner, G.; Attinger, S.; Jensen, B.; de Kroon, H.; Mommer, L.; et al. Plant species diversity affects infiltration capacity in an experimental grassland through changes in soil properties. Plant Soil 2015, 397, 1–16. [Google Scholar] [CrossRef]
- Tobner, C.M.; Paquette, A.; Gravel, D.; Reich, P.B.; Williams, L.J.; Messier, C. Functional identity is the main driver of diversity effects in young tree communities. Ecol. Lett. 2016, 19, 638–647. [Google Scholar] [CrossRef]
- Dawud, S.M.; Raulund-Rasmussen, K.; Ratcliffe, S.; Domisch, T.; Finér, L.; Joly, F.X.; Hättenschwiler, S.; Vesterdal, L.; Ostertag, R. Tree species functional group is a more important driver of soil properties than tree species diversity across major European forest types. Funct. Ecol. 2017, 31, 1153–1162. [Google Scholar] [CrossRef]
- Grandy, A.S.; Neff, J.C. Molecular C dynamics downstream: The biochemical decomposition sequence and its impact on soil organic matter structure and function. Sci. Total Environ. 2008, 404, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Cotrufo, M.F.; Ranalli, M.G.; Haddix, M.L.; Six, J.; Lugato, E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 2019, 12, 989–994. [Google Scholar] [CrossRef]
- Teixeira, L.H.; Oliveira, B.F.; Krah, F.-S.; Kollmann, J.; Ganade, G. Linking plant traits to multiple soil functions in semi-arid ecosystems. J. Arid Environ. 2020, 172, 104040. [Google Scholar] [CrossRef]
- Tilman, D.; Lehman, C.L.; Thomson, K.T. Plant diversity and ecosystem productivity: Theoretical considerations. Proc. Natl. Acad. Sci. USA 1997, 94, 1857–1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.P.; Tobias, J.A.; Sheil, D.; Meijaard, E.; Laurance, W.F. Maintaining ecosystem function and services in logged tropical forests. Trends Ecol. Evol. 2014, 29, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Wang, S.; Ali, A.; Gazol, A.; Ruiz-Benito, P.; Wang, X.; Lin, F.; Ye, J.; Hao, Z.; Loreau, M. Aboveground carbon storage is driven by functional trait composition and stand structural attributes rather than biodiversity in temperate mixed forests recovering from disturbances. Ann. For. Sci. 2018, 75, 2–13. [Google Scholar] [CrossRef] [Green Version]
- de Avila, A.L.; van der Sande, M.T.; Dormann, C.F.; Peña-Claros, M.; Poorter, L.; Mazzei, L.; Ruschel, A.R.; Silva, J.N.M.; de Carvalho, J.O.P.; Bauhus, J.; et al. Disturbance intensity is a stronger driver of biomass recovery than remaining tree-community attributes in a managed Amazonian forest. J. Appl. Ecol. 2018, 55, 1647–1657. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B. The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Buzzard, V.; Michaletz, S.T.; Deng, Y.; He, Z.; Ning, D.; Shen, L.; Tu, Q.; Van Nostrand, J.D.; Voordeckers, J.W.; Wang, J.; et al. Continental scale structuring of forest and soil diversity via functional traits. Nat. Ecol. Evol. 2019, 3, 1298–1308. [Google Scholar] [CrossRef]
- Diaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Prentice, I.C.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef]
- Handa, I.T.; Aerts, R.; Berendse, F.; Berg, M.P.; Bruder, A.; Butenschoen, O.; Chauvet, E.; Gessner, M.O.; Jabiol, J.; Makkonen, M.; et al. Consequences of biodiversity loss for litter decomposition across biomes. Nature 2014, 509, 218–221. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Chen, Y.; Castro-Izaguirre, N.; Baruffol, M.; Brezzi, M.; Lang, A.; Li, Y.; Hardtle, W.; von Oheimb, G.; Yang, X.; et al. Impacts of species richness on productivity in a large-scale subtropical forest experiment. Science 2018, 362, 80–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidl, R.; Spies, T.A.; Peterson, D.L.; Stephens, S.L.; Hicke, J.A. Searching for resilience: Addressing the impacts of changing disturbance regimes on forest ecosystem services. J. Appl. Ecol. 2016, 53, 120–129. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.B.; Zhao, Z.G.; Fang, Y. Soil hydro-physical characteristics and water retention function of typical shrubbery stands in the Yellow River Delta of China. Catena 2017, 156, 315–324. [Google Scholar] [CrossRef]
- Hao, Z.Q.; Zhang, J.; Song, B.; Ye, J.; Li, B.H. Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest. For. Ecol. Manag. 2007, 252, 1–11. [Google Scholar] [CrossRef]
- Yang, H.; Li, F. Distribution patterns of dominant tree species on northern slope of Changbai Moutain. Res. For. Ecosyst. 1985, 5, 1–14. [Google Scholar]
- Dai, L.M.; Chen, G.; Deng, H.B.; Ji, L.Z.; Hao, Z.Q.; Wang, Q.L. Structure characteristics and health distance assessment of various disturbed communities of Korean pine and broadleaved mixed forest in Changbai Mountains. Chin. J. Appl. Ecol. 2004, 15, 1750–1754. [Google Scholar]
- Harms, K.E.; Condit, R.; Hubbell, S.P.; Foster, R.B. Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. J. Ecol. 2001, 89, 947–959. [Google Scholar] [CrossRef]
- Kahl, T.; Bauhus, J. An index of forest management intensity based on assessment of harvested tree volume, tree species composition and dead wood origin. Nat. Conserv. 2014, 7, 15. [Google Scholar] [CrossRef]
- Shao, G.F.; Schall, P.; Weishampel, J.F. Dynamic simulations of mixed broadleaved-Pinus koraiensis forests in the Changbaishan biosphere reserve of China. For. Ecol. Manag. 1994, 70, 169–181. [Google Scholar] [CrossRef]
- Yuan, Z.; Ali, A.; Jucker, T.; Ruiz-Benito, P.; Wang, S.; Jiang, L.; Wang, X.; Lin, F.; Ye, J.; Hao, Z.; et al. Multiple abiotic and biotic pathways shape biomass demographic processes in temperate forests. Ecology 2019, 100, e02650. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Wang, S.; Gazol, A.; Mellard, J.; Lin, F.; Ye, J.; Hao, Z.; Wang, X.; Loreau, M. Multiple metrics of diversity have different effects on temperate forest functioning over succession. Oecologia 2016, 182, 1175–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Yan, E.-R.; Chang, S.X.; Cheng, J.-Y.; Liu, X.-Y. Community-weighted mean of leaf traits and divergence of wood traits predict aboveground biomass in secondary subtropical forests. Sci. Total Environ. 2017, 574, 654–662. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Lu, R. Analytical Methods of Soil and Agricultural Chemistry; China Agricultural Science and Technology Press: Beijing, China, 1999; pp. 107–240. [Google Scholar]
- Lefcheck, J.S. PIECEWISESEM: Piecewise structural equation modelling in R for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Shipley, B. Confirmatory path analysis in a generalized multilevel context. Ecology 2009, 90, 363–368. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H.; O’Hara, R.B. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Thom, D.; Seidl, R. Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biol. Rev. 2016, 91, 760–781. [Google Scholar] [CrossRef]
- Molino, J.F.; Sabatier, D. Tree diversity in tropical rain forests: A validation of the intermediate disturbance hypothesis. Science 2001, 294, 1702–1704. [Google Scholar] [CrossRef] [Green Version]
- Hiltner, U.; Huth, A.; Braeuning, A.; Herault, B.; Fischer, R. Simulation of succession in a neotropical forest: High selective logging intensities prolong the recovery times of ecosystem functions. For. Ecol. Manag. 2018, 430, 517–525. [Google Scholar] [CrossRef]
- Wu, Z.L.; Zhou, X.N.; Zheng, L.F.; Gao, S.; Luo, J.Z.; Cai, R.T.; Fang, W.C.; Wang, X.M. Study on soil physic chemical properties in natural forest selective cutting arae after 10 years. J. Mt. Sci. 2008, 26, 180–184. [Google Scholar]
- Dexter, A.R.; Richard, G.; Arrouays, D.; Czyż, E.A.; Jolivet, C.; Duval, O. Complexed organic matter controls soil physical properties. Geoderma 2008, 144, 620–627. [Google Scholar] [CrossRef]
- Huang, Z.; Clinton, P.W.; Davis, M.R. Post-harvest residue management effects on recalcitrant carbon pools and plant biomarkers within the soil heavy fraction in Pinus radiata plantations. Soil Biol. Biochem. 2011, 43, 404–412. [Google Scholar] [CrossRef]
- Mo, F.; Li, X.Y.; He, S.X.; Wang, X.X. Evaluation of soil and water conservation capacity of different forest types in Dongling Mountain. Acta Ecol. Sin. 2011, 31, 5009–5016. [Google Scholar]
- Chen, Y.L.; Li, C.R. Soil Moisture Properties under Pure and Mixed Plantations of Pinus Koraiensi and Fraxinus Mandshurica. Bull. Soil Water Conserv. 2011, 31, 85–87+116. [Google Scholar]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.; Xu, S.; Ciais, P.; Manzoni, S.; Fang, J.; Yu, G.; Tang, X.; Zhou, P.; Wang, W.; Yan, J.; et al. Climate and litter C/N ratio constrain soil organic carbon accumulation. Natl. Sci. Rev. 2019, 6, 746–757. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.K.; Wang, S.L. Forming and Stable Mechanism of Soil Aggregate and Influencing Factors. Chin. J. Soil Sci. 2005, 36, 415–421. [Google Scholar] [CrossRef]
- Gould, I.J.; Quinton, J.N.; Weigelt, A.; De Deyn, G.B.; Bardgett, R.D. Plant diversity and root traits benefit physical properties key to soil function in grasslands. Ecol. Lett. 2016, 19, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setala, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Wang, X.; Huang, Z.; Hong, M.M.; Zhao, Y.F.; Ou, Y.S.; Zhang, J. A comparison of the effects of natural vegetation regrowth with a plantation scheme on soil structure in a geological hazard-prone region. Eur. J. Soil Sci. 2019, 70, 674–685. [Google Scholar] [CrossRef]
- Kammer, P.M.; Schöb, C.; Eberhard, G.; Gallina, R.; Meyer, R.; Tschanz, C. The relationship between soil water storage capacity and plant species diversity in high alpine vegetation. Plant Ecol. Divers. 2013, 6, 457–466. [Google Scholar] [CrossRef]
- Green, J.K.; Seneviratne, S.I.; Berg, A.M.; Findell, K.L.; Hagemann, S.; Lawrence, D.M.; Gentine, P. Large influence of soil moisture on long-term terrestrial carbon uptake. Nature 2019, 565, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Regelink, I.C.; Stoof, C.R.; Rousseva, S.; Weng, L.; Lair, G.J.; Kram, P.; Nikolaidis, N.P.; Kercheva, M.; Banwart, S.; Comans, R.N.J. Linkages between aggregate formation, porosity and soil chemical properties. Geoderma 2015, 247, 24–37. [Google Scholar] [CrossRef]
- Meurer, K.; Barron, J.; Chenu, C.; Coucheney, E.; Fielding, M.; Hallett, P.; Herrmann, A.M.; Keller, T.; Koestel, J.; Larsbo, M.; et al. A framework for modelling soil structure dynamics induced by biological activity. Glob. Chang. Biol. 2020, 26, 5382–5403. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.Q.; Xie, C.K.; Cai, S.Z.; Chen, Y.; Lv, Y.P.; Mo, Z.L.; Liu, T.L.; Yang, Z.W. Effects of Tree Root Density on Soil Total Porosity and Non-Capillary Porosity Using a Ground-Penetrating Tree Radar Unit in Shanghai, China. Sustainability 2018, 10, 4640. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Yuan, Z.; Ali, A.; Sanaei, A.; Mao, Z.; Ding, F.; Zheng, D.; Fang, S.; Jia, Z.; Tao, Z.; et al. Anthropogenic Disturbances Shape Soil Capillary and Saturated Water Retention Indirectly via Plant Functional Traits and Soil Organic Carbon in Temperate Forests. Forests 2021, 12, 1588. https://doi.org/10.3390/f12111588
Liu S, Yuan Z, Ali A, Sanaei A, Mao Z, Ding F, Zheng D, Fang S, Jia Z, Tao Z, et al. Anthropogenic Disturbances Shape Soil Capillary and Saturated Water Retention Indirectly via Plant Functional Traits and Soil Organic Carbon in Temperate Forests. Forests. 2021; 12(11):1588. https://doi.org/10.3390/f12111588
Chicago/Turabian StyleLiu, Shufang, Zuoqiang Yuan, Arshad Ali, Anvar Sanaei, Zikun Mao, Fan Ding, Di Zheng, Shuai Fang, Zhaojie Jia, Zhao Tao, and et al. 2021. "Anthropogenic Disturbances Shape Soil Capillary and Saturated Water Retention Indirectly via Plant Functional Traits and Soil Organic Carbon in Temperate Forests" Forests 12, no. 11: 1588. https://doi.org/10.3390/f12111588
APA StyleLiu, S., Yuan, Z., Ali, A., Sanaei, A., Mao, Z., Ding, F., Zheng, D., Fang, S., Jia, Z., Tao, Z., Lin, F., Ye, J., Wang, X., & Hao, Z. (2021). Anthropogenic Disturbances Shape Soil Capillary and Saturated Water Retention Indirectly via Plant Functional Traits and Soil Organic Carbon in Temperate Forests. Forests, 12(11), 1588. https://doi.org/10.3390/f12111588