
Colonization Pattern of Abandoned Croplands by Quercus pyrenaica in a Mediterranean Mountain Region



Abstract
1. Introduction
2. Materials and Methods
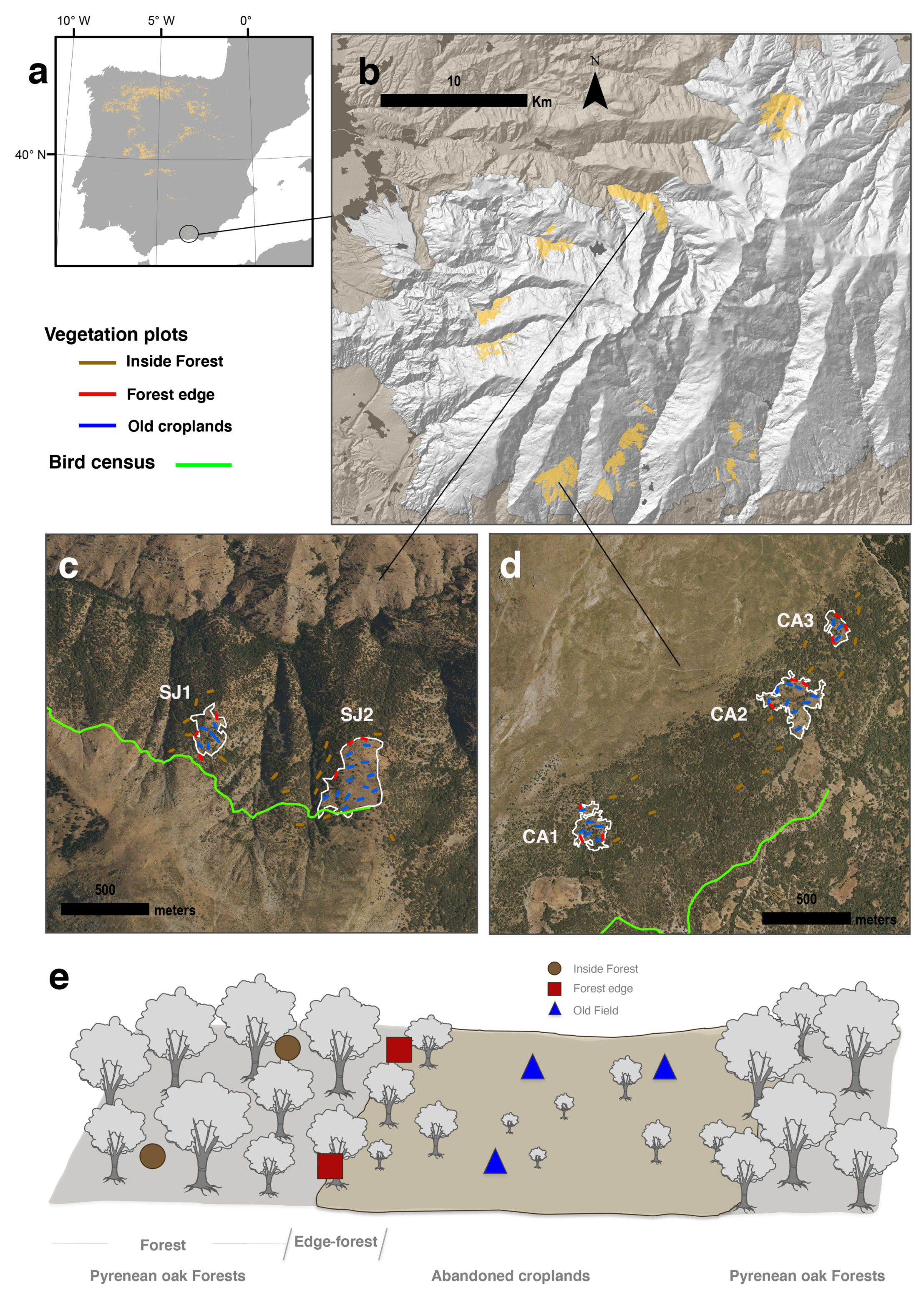
2.1. Sampling Description
2.2. Data Analysis
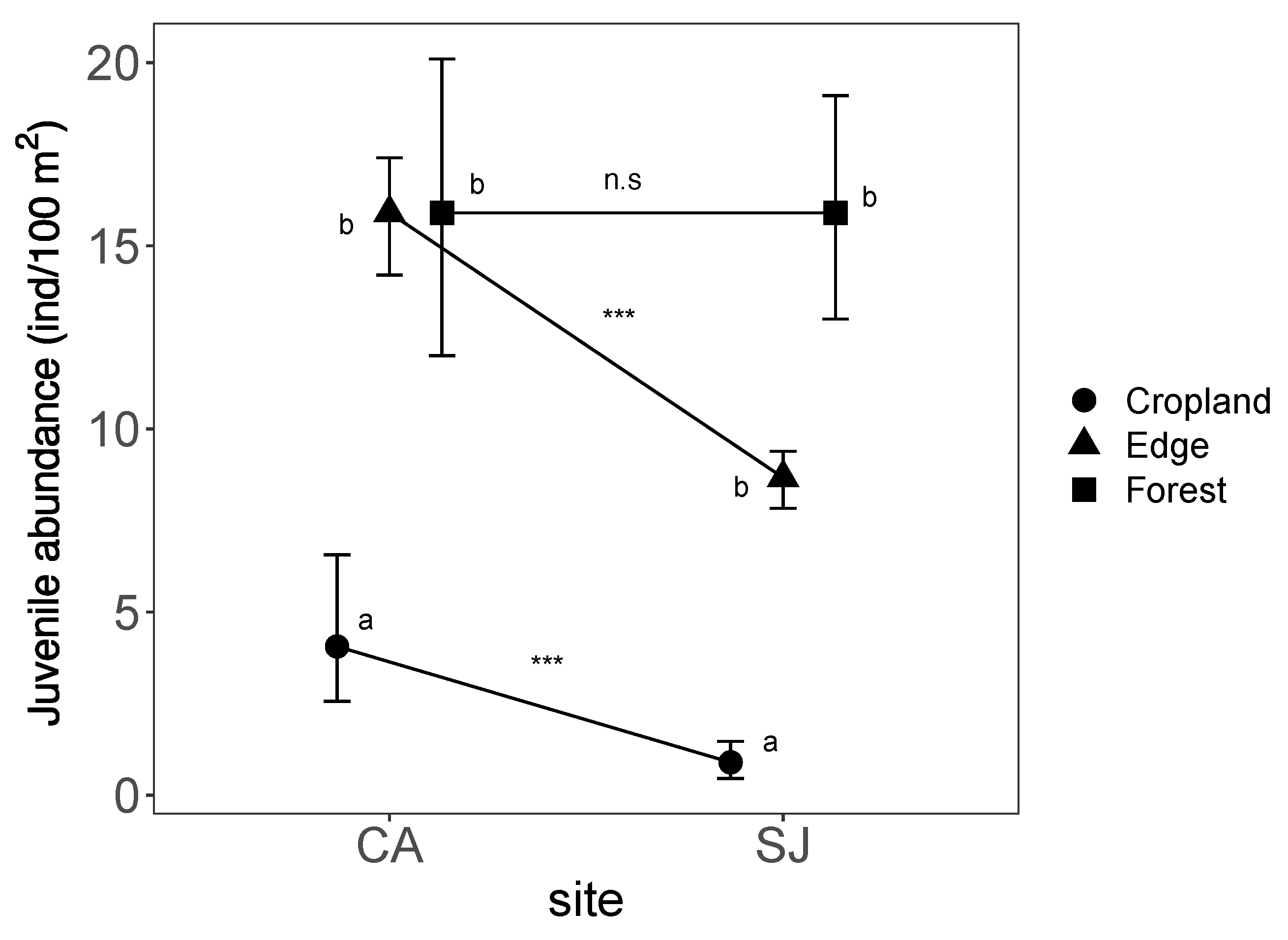
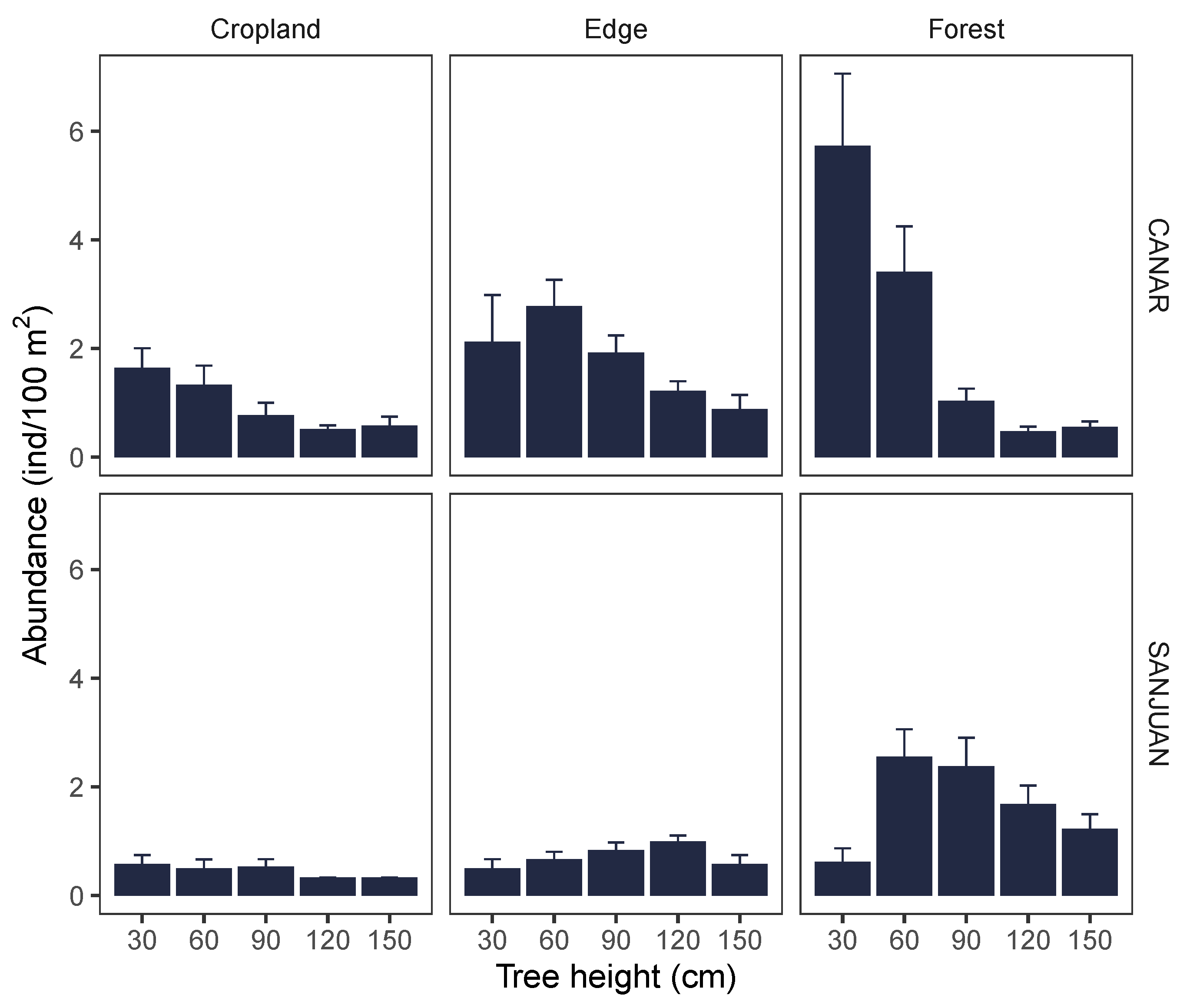
3. Results
4. Discussion
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butchart, S.H.M.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global Biodiversity: Indicators of Recent Declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef]
- Winkler, K.; Fuchs, R.; Rounsevell, M.; Herold, M. Global land use changes are four times greater than previously estimated. Nat. Commun. 2021, 12, 2501. [Google Scholar] [CrossRef]
- Sala, O.E. Global Biodiversity Scenarios for the Year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Lindenmayer, D.; Cunningham, S.; Young, A. Land Use Intensification: Effects on Agriculture, Biodiversity and Ecological Processes; CSIRO Publishing: Melbourne, Australia, 2012. [Google Scholar]
- Hasan, S.S.; Zhen, L.; Miah, M.G.; Ahamed, T.; Samie, A. Impact of land use change on ecosystem services: A review. Environ. Dev. 2020, 34, 100527. [Google Scholar] [CrossRef]
- Rey Benayas, J.M. Abandonment of agricultural land: An overview of drivers and consequences. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2007, 2, 1–14. [Google Scholar] [CrossRef]
- Pías, B.; Escribano-Avila, G.; Virgós, E.; Sanz-Pérez, V.; Escudero, A.; Valladares, F. The colonization of abandoned land by Spanish juniper: Linking biotic and abiotic factors at different spatial scales. For. Ecol. Manag. 2014, 329, 186–194. [Google Scholar] [CrossRef]
- Valbuena-Carabaña, M.; de Heredia, U.L.; Fuentes-Utrilla, P.; González-Doncel, I.; Gil, L. Historical and recent changes in the Spanish forests: A socio-economic process. Rev. Palaeobot. Palynol. 2010, 162, 492–506. [Google Scholar] [CrossRef]
- Martínez-Fernández, J.; Ruiz-Benito, P.; Zavala, M.A. Recent land cover changes in Spain across biogeographical regions and protection levels: Implications for conservation policies. Land Use Policy 2015, 44, 62–75. [Google Scholar] [CrossRef]
- Rounsevell, M.; Reginster, I.; Araújo, M.; Carter, T.; Dendoncker, N.; Ewert, F.; House, J.; Kankaanpää, S.; Leemans, R.; Metzger, M.; et al. A coherent set of future land use change scenarios for Europe. Agric. Ecosyst. Environ. 2006, 114, 57–68. [Google Scholar] [CrossRef]
- Perpiña Castillo, C.; Jacobs-Crisioni, C.; Diogo, V.; Lavalle, C. Modelling agricultural land abandonment in a fine spatial resolution multi-level land-use model: An application for the EU. Environ. Model. Softw. 2021, 136, 104946. [Google Scholar] [CrossRef]
- Sheffer, E. A review of the development of Mediterranean pine–oak ecosystems after land abandonment and afforestation: Are they novel ecosystems? Ann. For. Sci. 2012, 69, 429–443. [Google Scholar] [CrossRef]
- Debussche, M.; Lepart, J.; Dervieux, A. Mediterranean landscape changes: Evidence from old postcards. Glob. Ecol. Biogeogr. 1999, 8, 3–15. [Google Scholar] [CrossRef]
- Peñuelas, J.; Boada, M. A global change-induced biome shift in the Montseny mountains (NE Spain): A global change-induced biome shift. Glob. Chang. Biol. 2003, 9, 131–140. [Google Scholar] [CrossRef]
- Álvarez-Martínez, J.M.; Suárez-Seoane, S.; Stoorvogel, J.J.; de Luis Calabuig, E. Influence of land use and climate on recent forest expansion: A case study in the Eurosiberian—Mediterranean limit of north-west Spain. J. Ecol. 2014, 102, 905–919. [Google Scholar] [CrossRef]
- de Natale, F.; Gasparini, P.; Carriero, A. A study on tree colonization of abandoned land in the Italian Alps: Extent and some characteristics of new forest stands in Trentino. In Sustainable Forestry: From Monitoring and Modelling to Knowledge Management and Policy Science; Reynolds, K.M., Thomson, A.J., Köhl, M., Shannon, M.A., Ray, D., Rennolls, K., Eds.; CABI: Wallingford, UK, 2007; pp. 269–284. [Google Scholar]
- Piussi, P. Expansion of European mountain forests. In Forests in Sustainable Mountain Development: A State of Knowledge Report for 2000. Task Force on Forests in Sustainable Mountain Development; CABI: Wallingford, UK, 2000; pp. 19–28. [Google Scholar]
- MacDonald, D.; Crabtree, J.; Wiesinger, G.; Dax, T.; Stamou, N.; Fleury, P.; Gutierrez Lazpita, J.; Gibon, A. Agricultural abandonment in mountain areas of Europe: Environmental consequences and policy response. J. Environ. Manag. 2000, 59, 47–69. [Google Scholar] [CrossRef]
- Jiménez-Olivencia, Y.; Porcel, L.; Caballero, A. Medio siglo en la evolución de los paisajes naturales y agrarios de Sierra Nevada (España). Boletín de la Asociación de Geógrafos Españoles 2015, 68, 205–232. [Google Scholar] [CrossRef]
- European Environment Agency. Europe’s Ecological Backbone: Recognising the True Value of Our Mountains; Publications Office: København, Denmark, 2010. [Google Scholar]
- Rutherford, G.N.; Bebi, P.; Edwards, P.J.; Zimmermann, N.E. Assessing land-use statistics to model land cover change in a mountainous landscape in the European Alps. Ecol. Model. 2008, 212, 460–471. [Google Scholar] [CrossRef]
- Zimmermann, P.; Tasser, E.; Leitinger, G.; Tappeiner, U. Effects of land-use and land-cover pattern on landscape-scale biodiversity in the European Alps. Agric. Ecosyst. Environ. 2010, 139, 13–22. [Google Scholar] [CrossRef]
- Mietkiewicz, N.; Kulakowski, D.; Rogan, J.; Bebi, P. Long-term change in sub-alpine forest cover, tree line and species composition in the Swiss Alps. J. Veg. Sci. 2017, 28, 951–964. [Google Scholar] [CrossRef]
- García, I.; Jiménez, P. 9230 Robledales de Quercus pyrenaica y robledales de Quercus robur y Quercus pyrenaica del noroeste ibérico. In Bases Ecológicas Preliminares para la Conservación de los Tipos de Hábitat de Interés Comunitario en España; VV.AA., Ed.; Ministerio de Medio Ambiente, y Medio Rural y Marino: Madrid, Spain, 2009; pp. 1–66. [Google Scholar]
- Alba-Sánchez, F.; Abel-Schaad, D.; López-Sáez, J.A.; Sabariego-Ruiz, S.; Pérez-Díaz, S.; Luelmo-Lautenschlaeger, R.; Garrido-García, J.A. Early anthropogenic change in western Mediterranean mountains (Sierra Nevada, SE Spain). Anthropocene 2021, 33, 100278. [Google Scholar] [CrossRef]
- Gavilán, R.G.; Escudero, A.; Rubio, A. Effects of disturbance on floristic patterns of Quercus pyrenaica forests in Central Spain. In Vegetation Science in Retrospect and Perspective—Proceedings 41st IAVS Symposium; International Association for Vegetation Science: Montpellier, France, 2000; pp. 226–229. [Google Scholar]
- Calvo, L.; Tárrega, R.; de Luis, E. Post-fire succession in two Quercus pyrenaica communities with different disturbance histories. Ann. For. Sci. 1999, 56, 441–447. [Google Scholar] [CrossRef]
- Tárrega, R.; Calvo, L.; Marcos, E.; Taboada, A. Forest structure and understory diversity in Quercus pyrenaica communities with different human uses and disturbances. For. Ecol. Manag. 2006, 227, 50–58. [Google Scholar] [CrossRef]
- Ruiz de la Torre, J. Flora Mayor; Organismo Autónomo Parques Nacionales: Madrid, Spain, 2006. [Google Scholar]
- Sánchez Palomares, O.; Roig, S.; del Río, M.; Rubio-Sánchez, A.; Gandullo, J.M. Las Estaciones Ecológicas Actuales y Potenciales de los Rebollares Españoles; Number nº 17 in Monografias INIA, Serie Forestal; Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria: Madrid, Spain, 2008. [Google Scholar]
- Herrera Calvo, P.M. El uso pastoral como alternativa de gestión de los hábitats vinculados a los rebollares ibéricos. Rev. Pastos 2016, 46, 6–23. [Google Scholar]
- Álvarez, R.; García-Duro, J.; Muñoz, J.; Pesqueira, X.; Rivas, M.; Reyes, O.; Casal, M. Cambios en la estructura y diversidad, y en los modelos de combustible del bosque de Quercus pyrenaica tras fuego. In Proceedings of the V Congreso Forestal Español, Ávila, Spain, 21–25 September 2009. [Google Scholar]
- Valbuena-Carabaña, M.; Gil, L. Centenary coppicing maintains high levels of genetic diversity in a root resprouting oak (Quercus pyrenaica Willd.). Tree Genet. Genomes 2017, 13, 28. [Google Scholar] [CrossRef]
- Cañellas, I.; Del-Río, M.; Roig, S.; Montero, G. Growth response to thinning in Quercus pyrenaica Willd. coppice stands in Spanish central mountain. Ann. For. Sci. 2004, 61, 243–250. [Google Scholar] [CrossRef]
- Bravo, J.A.; Roig, S.; Serrada, R. Selvicultura en montes bajos y medios de Quercus ilex L., Q. pyrenaica Willd. y Q. faginea Lam. In Compendio de Selvicultura Aplicada en España; Serrada, R., Montero, G., Reque, J.A., Eds.; INIA: Madrid, Spain, 2008; pp. 657–744. [Google Scholar]
- Valbuena-Carabaña, M.; Gil, L. Efectos de la gestión selvícola pasada y presente sobre la diversidad genética actual y futura de Quercus pyrenaica Willd. en Sierra Nevada. Ecosistemas 2014, 23, 90–98. [Google Scholar]
- Piqué, M.; Vericat, P. Evolution and perspectives of coppice forests in European countries and South Africa. In FPS COST Action FP1301: Innovative Management and Multifunctional Utilization of Traditional Coppice Forests? An Answer to Future Ecological, Economic and Social Challenges in the European Forestry Sector (EuroCoppice); Bartlett, D., Nicolescu, V.N., Pyttel, P., Eds.; 2015; pp. 38–40. Available online: https://gala.gre.ac.uk/id/eprint/14009/ (accessed on 1 May 2018).
- Piqué, M.; Laina, R.; Vericat Grau, P.; Beltrán, M.; Busquets, E.; Tolosana, E. Spain. In Coppice Forests in Europe; Unrau, A., Becker, G., Spinelli, R., Lazdina, D., Magagnotti, N., Nicolescu, V., Buckley, P., Barlett, D., Kofman, P., Eds.; Albert Ludwig University of Freiburg: Freiburg im Breisgau, Germany, 2018; pp. 331–336. [Google Scholar]
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef]
- Fady, B.; Aravanopoulos, F.A.; Alizoti, P.; Mátyás, C.; von Wühlisch, G.; Westergren, M.; Belletti, P.; Cvjetkovic, B.; Ducci, F.; Huber, G.; et al. Evolution-based approach needed for the conservation and silviculture of peripheral forest tree populations. For. Ecol. Manag. 2016, 375, 66–75. [Google Scholar] [CrossRef]
- Jump, A.S.; Cavin, L.; Hunter, P.D. Monitoring and managing responses to climate change at the retreating range edge of forest trees. J. Environ. Monit. 2010, 12, 1791–1798. [Google Scholar] [CrossRef]
- Benavides, R.; Rabasa, S.G.; Granda, E.; Escudero, A.; Hódar, J.A.; Martínez-Vilalta, J.; Rincón, A.M.; Zamora, R.; Valladares, F. Direct and indirect effects of climate on demography and early growth of Pinus sylvestris at the rear edge: Changing roles of biotic and abiotic factors. PLoS ONE 2013, 8, e59824. [Google Scholar] [CrossRef]
- Matías, L.; Linares, J.C.; Sánchez-Miranda, A.; Jump, A.S. Contrasting growth forecasts across the geographical range of Scots pine due to altitudinal and latitudinal differences in climatic sensitivity. Glob. Chang. Biol. 2017, 23, 4106–4116. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M. Seedling establishment of a boreal tree species (Pinus sylvestris) at its southernmost distribution limit: Consequences of being in a marginal Mediterranean habitat. J. Ecol. 2004, 92, 266–277. [Google Scholar] [CrossRef]
- Jiménez-Olivencia, Y.; Porcel-Rodríguez, L.; Caballero-Calvo, A.; Bonet, F.J. Evolución de los usos del suelo en Sierra Nevada en los últimos 50 años y cambios en el paisaje. In La Huella del Cambio Global en Sierra Nevada: Retos Para la Conservación; Zamora, R., Pérez-Luque, A.J., Bonet, F., Barea-Azcón, J., Aspizua, R., Eds.; Junta de Andalucía, Consejería de Medio Ambiente y Ordenación del Territorio: Seville, Spain, 2015; pp. 54–56. [Google Scholar]
- Pérez-Luque, A.; Gea-Izquierdo, G.; Zamora, R. Land-use legacies and climate change as a double challenge to oak forest resilience: Mismatches of geographical and ecological rear edges. Ecosystems 2021, 24, 755–773. [Google Scholar] [CrossRef]
- Pérez-Luque, A.; Benito, B.M.; Bonet-García, F.J.; Zamora, R. Ecological Diversity within Rear-Edge: A Case Study from Mediterranean Quercus pyrenaica Willd. Forests 2021, 12, 10. [Google Scholar] [CrossRef]
- CMA. Mapa de usos y Coberturas de Andalucía 1956–1999–2003, 1:25,000; Consejería de Medio Ambiente, Junta de Andalucía: Sevilla, Spain, 2007. [Google Scholar]
- Navarro-González, I.; Bonet-García, F.; Pérez-Luque, A. Cartografía histórica de la vegetación mediante ortofotos. In Observatorio de Cambio Global de Sierra Nevada. Metodologías de Seguimiento; Aspizua, R., Barea-Azcón, J., Bonet, F., Pérez-Luque, A., Zamora, R., Eds.; Consejería de Medio Ambiente, Junta de Andalucía: Sevilla, Spain, 2012; pp. 16–17. [Google Scholar]
- Moreno-Llorca, R.A.; Pérez-Luque, A.J.; Bonet, F.J.; Pérez-Pérez, R.; Zamora, R. Caracterización de fuentes de información para la reconstrucción histórica de la vegetación. Un caso de estudio en Sierra Nevada. In Proceedings of the XII Congreso Nacional de Medio Ambiente (CONAMA 2014), Madrid, Spain, 24–27 November 2014. [Google Scholar]
- Moreno-Llorca, R.; Pérez-Luque, A.; Bonet, F.; Zamora, R. Historical analysis of socio-ecological changes in the municipality of Cáñar (Alpujarra, Sierra Nevada) over the last 5 centuries. In Global Change Impacts in Sierra Nevada: Challenges for Conservation; Zamora, R., Pérez-Luque, A., Bonet, F., Barea-Azcón, J., Aspizua, R., Eds.; Consejería de Medio Ambiente y Ordenación del Territorio, Junta de Andalucía: Sevilla, Spain, 2016; pp. 59–62. [Google Scholar]
- Plieninger, T.; Rolo, V.; Moreno, G. Large-Scale Patterns of Quercus ilex, Quercus suber, and Quercus pyrenaica Regeneration in Central-Western Spain. Ecosystems 2010, 13, 644–660. [Google Scholar] [CrossRef]
- Pérez-Luque, A.J.; Zamora, R.; Bonet, F.J.; Pérez-Pérez, R. Dataset of MIGRAME project (global change, altitudinal range shift and colonization of degraded habitats in Mediterranean mountains). PhytoKeys 2015, 56, 61–81. [Google Scholar] [CrossRef] [PubMed]
- Barea-Azcón, J.; Martín-Jaramillo, J.; López-Sanjuan, R. Passeriformes y otras aves. In Observatorio de Cambio Global de Sierra Nevada. Metodologías de Seguimiento; Aspizua, R., Barea-Azcón, J., Bonet, F., Pérez-Luque, A., Zamora, R., Eds.; Consejería de Medio Ambiente, Junta de Andalucía: Sevilla, Spain, 2012; pp. 86–87. [Google Scholar]
- Pérez-Luque, A.J.; Barea-Azcón, J.M.; Álvarez-Ruiz, L.; Bonet-García, F.J.; Zamora, R. Dataset of Passerine bird communities in a Mediterranean high mountain (Sierra Nevada, Spain). ZooKeys 2016, 552, 137–154. [Google Scholar] [CrossRef]
- Gómez, J.M. Spatial patterns in long-distance dispersal of Quercus ilex acorns by jays in a heterogeneous landscape. Ecography 2003, 26, 573–584. [Google Scholar] [CrossRef]
- Pons, J.; Pausas, J.G. Acorn dispersal estimated by radio-tracking. Oecologia 2007, 153, 903–911. [Google Scholar] [CrossRef]
- Zamora, R.; Pérez-Luque, A.J.; Benito, B.M.; Bonet-García, F.J.; Navarro, I.; Pérez-Pérez, R.; Hódar, J.A.; Matías, L. Cambio Global, Migración Altitudinal y Colonización de Hábitats Degradados en Montañas Mediterráneas (MIGRAME); Invitación a la Ecología; Asociación Española de Ecología Terrestre: Pamplona, Spain, 2013. [Google Scholar]
- Zamora, R.; Barea-Azcón, J.M. Long-term changes in mountain passerine bird communities in the Sierra Nevada (southern Spain): A 30-year case study. Ardeola 2015, 62, 3–18. [Google Scholar] [CrossRef]
- Dunn, P.K.; Smyth, G.K. Tweedie GLMs. In Generalized Linear Models With Examples in R; Springer Texts in Statistics; Springer: New York, NY, USA, 2018; pp. 457–490. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Ieno, E.N.; Zuur, A.F. A Beginner’S Guide to Data Exploration and Visualization with R; Highland Statistics Ltd.: Newburgh, UK, 2015. [Google Scholar]
- Dunn, P.K.; Smyth, G.K. Series evaluation of Tweedie exponential dispersion models. Stat. Comput. 2005, 15, 267–280. [Google Scholar] [CrossRef]
- Dunn, P.K. Tweedie: Evaluation of Tweedie Exponential Family Models, R package version 2.3.3; 2017. Available online: https://cran.r-project.org/web/packages/tweedie/tweedie.pdf (accessed on 7 September 2021).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2010. [Google Scholar]
- Gómez, J.; García, D.; Zamora, R. Impact of vertebrate acorn- and seedling-predators on a Mediterranean Quercus pyrenaica forest. For. Ecol. Manag. 2003, 180, 125–134. [Google Scholar] [CrossRef]
- Perea, R.; San Miguel, A.; Gil, L. Interacciones planta-animal en la regeneración de Quercus pyrenaica: Ecología y gestión. Ecosistemas 2014, 23, 18–26. [Google Scholar]
- Ameztegui, A.; Coll, L.; Brotons, L.; Ninot, J.M. Land-use legacies rather than climate change are driving the recent upward shift of the mountain tree line in the Pyrenees. Glob. Ecol. Biogeogr. 2016, 25, 263–273. [Google Scholar] [CrossRef]
- Améztegui, A.; Brotons, L.; Coll, L. Land-use changes as major drivers of mountain pine (Pinus uncinata Ram.) expansion in the Pyrenees: Land-use changes drive Pinus uncinata expansion. Glob. Ecol. Biogeogr. 2010, 19, 632–641. [Google Scholar] [CrossRef]
- Lasanta-Martínez, T.; Vicente-Serrano, S.M.; Cuadrat-Prats, J.M. Mountain Mediterranean landscape evolution caused by the abandonment of traditional primary activities: A study of the Spanish Central Pyrenees. Appl. Geogr. 2005, 25, 47–65. [Google Scholar] [CrossRef]
- Kozak, J. Forest cover change in the western carpathians in the past 180 years: A case study in the Orawa region in Poland. Mt. Res. Dev. 2003, 23, 369–375. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Lasanta, T.; Romo, A. Analysis of spatial and temporal evolution of vegetation cover in the spanish central pyrenees: Role of human management. Environ. Manag. 2004, 34, 802–818. [Google Scholar] [CrossRef]
- European Environment Agency. European Forest Ecosystems State and Trends; EUR-OP: København, Denmark, 2016. [Google Scholar]
- Puerta-Piñero, C.; Espelta, J.M.; Sánchez-Humanes, B.; Rodrigo, A.; Coll, L.; Brotons, L. History matters: Previous land use changes determine post-fire vegetation recovery in forested Mediterranean landscapes. For. Ecol. Manag. 2012, 279, 121–127. [Google Scholar] [CrossRef]
- Hermy, M.; Verheyen, K. Legacies of the past in the present-day forest biodiversity: A review of past land-use effects on forest plant species composition and diversity. Ecol. Res. 2007, 22, 361–371. [Google Scholar] [CrossRef]
- Cruz-Alonso, V.; Ruiz-Benito, P.; Villar-Salvador, P.; Rey-Benayas, J.M. Long-term recovery of multifunctionality in Mediterranean forests depends on restoration strategy and forest type. J. Appl. Ecol. 2019, 56, 745–757. [Google Scholar] [CrossRef]
- Minotta, G.; Degioanni, D. Naturally regenerated English oak (Quercus robur L.) stands on abandoned agricultural lands in Rilate valley (Piedmont Region, NW Italy). IForest Biogeosci. For. 2011, 4, 31–37. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Cañellas, I. Local climate forces instability in long-term productivity of a Mediterranean oak along climatic gradients. Ecosystems 2014, 17, 228–241. [Google Scholar] [CrossRef]
- Nathan, R. Long-distance dispersal of plants. Science 2006, 313, 786–788. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, N.; Kellman, M. Tree seed dispersal among forest fragments: II. Dispersal abilities and biogeographical controls. J. Biogeogr. 2002, 29, 351–363. [Google Scholar] [CrossRef]
- Kurek, P.a.; Dobrowolska, D.; Wiatrowska, B. Dispersal distance and burial mode of acorns in eurasian jays garrulus glandarius in european temperate forests. Acta Ornithol. 2019, 53, 155–162. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Cañellas, I.; Montero, G. Acorn production in Spanish holm oak woodlands. Investig. Agrar. Sist. y Recur. For. 2006, 15, 339–354. [Google Scholar] [CrossRef]
- Leal, J. Análisis del Crecimiento Vegetativo e Inversión Reproductiva de Quercus pyrenaica Willd. en tres localidades de Sierra Nevada. Master’s Thesis, Universidad de Granada, Granada, Spain, 2013. [Google Scholar]
- Gómez, J.; Gómez-Aparicio, L.; Zamora, R.; Montes, J. Problemas de Regeneración de Especies Forestales Autóctonas en el Espacio Natural Protegido de Sierra Nevada. In Proceedings of the III Congreso Forestal Español, Granada, Spain, 25–28 September 2001. [Google Scholar]
- Navarro-González, I.; Pérez-Luque, A.J.; Bonet, F.J.; Zamora, R. The weight of the past: Land-use legacies and recolonization of pine plantations by oak trees. Ecol. Appl. 2013, 23, 1267–1276. [Google Scholar] [CrossRef]
- Milder, A.I.; Fernández-Santos, B.; Martínez-Ruiz, C. Colonization patterns of woody species on lands mined for coal in Spain: Preliminary insights for forest expansion. Land Degrad. Dev. 2013, 24, 39–46. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Rey Benayas, J.M.; Castro, J. Shifting demographic conflicts across recruitment cohorts in a dynamic post-disturbance landscape. Ecology 2016, 97, 2628–2639. [Google Scholar] [CrossRef] [PubMed]
- Perring, M.P.; De Frenne, P.; Baeten, L.; Maes, S.L.; Depauw, L.; Blondeel, H.; Carón, M.M.; Verheyen, K. Global environmental change effects on ecosystems: The importance of land-use legacies. Glob. Chang. Biol. 2016, 22, 1361–1371. [Google Scholar] [CrossRef] [PubMed]
- Titos, M. (Ed.) Las minas de la Estrella. In La Aventura de Sierra-Nevada 1717–1915; Editorial Universidad de Granada: Granada, Spain, 1990; pp. 226–236. [Google Scholar]
- Mesa-Torres, M. Cáñar: Balcón de la Alpujarra; Fundación Caja General de Ahorros de Granada: Granada, Spain, 2009. [Google Scholar]
- Moreno-Llorca, R.; Zamora, R. Caracterización de la Carga Ganadera en Sierra Nevada en la Segunda Mitad del Siglo XX. Proyecto MIGRAME; Project Report; IISTA-CEAMA; University of Granada: Granada, Spain, 2012. [Google Scholar]
- Robles-Cruz, A.B. “En el conjunto de las Sierras Béticas”: Pastos, producción, diversidad y cambio global. In Pastos, Clave en la Gestión de los Territorios: Integrando Disciplinas; Fernández Rebollo, P., Gómez Cabrera, A., Guerrero, J.E., Garrido Varo, A., Calzado Martínez, C., García Moreno, A.M., Carbonero Muñoz, M.D., Blázquez Carrasco, A., Escuín Royo, S., Castillo Carrión, S., Eds.; Junta de Andalucía, Consejería de Agricultura y Pesca: Sevilla, Spain, 2008; pp. 30–51. [Google Scholar]
- Castro, J.; Zamora, R.; Hódar, J.A. Restoring Quercus pyrenaica forests using pioneer shrubs as nurse plants. Appl. Veg. Sci. 2006, 9, 137–142. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Zamora, R.; Gómez, J.M.; Hódar, J.A.; Castro, J.; Baraza, E. Applying plant facilitation to forest restoration: A meta-analysis of the use of shrubs as nurse plants. Ecol. Appl. 2004, 14, 1128–1138. [Google Scholar] [CrossRef]
- Costa, A.; Villa, S.; Alonso, P.; García-Rodríguez, J.; Martín, F.; Martínez-Ruiz, C.; Fernández-Santos, B. Can native shrubs facilitate the early establishment of contrasted co-occurring oaks in Mediterranean grazed areas? J. Veg. Sci. 2017, 28, 1047–1056. [Google Scholar] [CrossRef]
- Baraza, E.; Gómez, J.M.; Hódar, J.A.; Zamora, R. Herbivory has a greater impact in shade than in sun: Response of Quercus pyrenaica seedlings to multifactorial environmental variation. Can. J. Bot. 2004, 82, 357–364. [Google Scholar] [CrossRef]
- Muñoz, J. Bosques autóctonos y matorrales de media y alta montaña. In Observatorio de Cambio Global de Sierra Nevada: Metodologías de Seguimiento; Aspizua, R., Barea-Azcón, J., Bonet, F., Pérez-Luque, A., Zamora, R., Eds.; Consejería de Medio Ambiente, Junta de Andalucía: Sevilla, Spain, 2014; pp. 74–75. [Google Scholar]
- Chazdon, R.L.; Lindenmayer, D.; Guariguata, M.R.; Crouzeilles, R.; Rey Benayas, J.M.; Lazos Chavero, E. Fostering natural forest regeneration on former agricultural land through economic and policy interventions. Environ. Res. Lett. 2020, 15, 043002. [Google Scholar] [CrossRef]
- Stritih, A.; Senf, C.; Seidl, R.; Grêt-Regamey, A.; Bebi, P. The impact of land-use legacies and recent management on natural disturbance susceptibility in mountain forests. For. Ecol. Manag. 2021, 484, 118950. [Google Scholar] [CrossRef]
- Pausas, J.; Ribeiro, E.; Dias, S.; Pons, J.; Beseler, C. Regeneration of a marginal Quercus suber forest in the eastern Iberian Peninsula. J. Veg. Sci. 2006, 17, 729–738. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Cropland | Elevation (m) | Area (ha) | Number of Plots | |||
|---|---|---|---|---|---|---|---|
| Code | Abandonment Age (Years) | Cropland | Edge | Forest | |||
| Robledal de Cáñar | CA1 | >60 | 1796–1866 | 3.29 | 6 | 3 | 4 |
| CA2 | <30 | 1789–1858 | 5.80 | 9 | 3 | 7 | |
| CA3 | 40–60 | 1851–1892 | 1.56 | 3 | 3 | 4 | |
| Robledal de San Juan | SJ1 | 40–60 | 1507–1674 | 3.47 | 6 | 3 | 6 |
| SJ2 | 30–40 | 1575–1746 | 10.36 | 13 | 3 | 10 | |
| Variable | Southern Site (CA) | Northern Site (SJ) | U Statistic | p Value |
|---|---|---|---|---|
| % of Q. pyrenaica | 96.11 ± 1.28 | 100 ± 0 | 3.688 | 0.0001 |
| Tree density (ind/ha) | 1671.11 ± 229.21 | 1587.5 ± 161.67 | 4.808 | 0.9369 |
| Juvenile abundance (ind/ha) | 1004.44 ± 195.72 | 883.33 ± 127.18 | 4.852 | 0.7667 |
| Adult abundance (ind/ha) | 584.44 ± 80.47 | 704.17 ± 63.31 | 4.448 | 0.1780 |
| Maximum tree height (m) | 13.93 ± 0.65 | 13.75 ± 0.71 | 4.824 | 0.8736 |
| Tree height mean (m) | 4.32 ± 0.6 | 5.09 ± 0.37 | 4.330 | 0.0855 |
| Tree height median (m) | 3.19 ± 0.83 | 3.57 ± 0.66 | 4.564 | 0.3527 |
| Tree height 75 percentile (m) | 5.73 ± 1.02 | 8.29 ± 0.6 | 4.343 | 0.0922 |
| Tree height 90 percentile (m) | 10.07 ± 0.95 | 11.22 ± 0.54 | 4.605 | 0.4399 |
| Basal Area (m/ha) | 37.56 ± 4.23 | 33.58 ± 3.6 | 4.912 | 0.5400 |
| Model | df | logLik | AICc | AICc | Nagelkerke R2 |
|---|---|---|---|---|---|
| Habitat type + Site + Habitat type × Site | 6 | −221.89 | 457.78 | 0 | 0.970 |
| Habitat type + Site | 4 | −231.83 | 473.65 | 15.87 | 0.955 |
| Habitat type | 3 | −236.21 | 480.42 | 22.64 | 0.946 |
| Site | 2 | −291.06 | 588.12 | 130.34 | 0.116 |
| null model | 1 | −293.09 | 590.19 | 132.41 | 0 |
| Variable | SS | df | F | p-Value |
|---|---|---|---|---|
| Habitat type | 233.89 | 2 | 72.95 | 0.0001 |
| Site | 13.09 | 1 | 8.16 | 0.0054 |
| Habitat type × Site | 27.19 | 2 | 8.48 | 0.0004 |
| Residuals | 123.43 | 77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Luque, A.J.; Bonet-García, F.J.; Zamora, R. Colonization Pattern of Abandoned Croplands by Quercus pyrenaica in a Mediterranean Mountain Region. Forests 2021, 12, 1584. https://doi.org/10.3390/f12111584
Pérez-Luque AJ, Bonet-García FJ, Zamora R. Colonization Pattern of Abandoned Croplands by Quercus pyrenaica in a Mediterranean Mountain Region. Forests. 2021; 12(11):1584. https://doi.org/10.3390/f12111584
Chicago/Turabian StylePérez-Luque, Antonio J., Francisco J. Bonet-García, and Regino Zamora. 2021. "Colonization Pattern of Abandoned Croplands by Quercus pyrenaica in a Mediterranean Mountain Region" Forests 12, no. 11: 1584. https://doi.org/10.3390/f12111584
APA StylePérez-Luque, A. J., Bonet-García, F. J., & Zamora, R. (2021). Colonization Pattern of Abandoned Croplands by Quercus pyrenaica in a Mediterranean Mountain Region. Forests, 12(11), 1584. https://doi.org/10.3390/f12111584