1. Introduction
The size of the assimilatory apparatus in trees may be characterised using different indicators, from the simplest, but at the same time less accurate dimensional traits of the crown, such as crown projection area and crown volume, to more accurate quantitative traits, e.g., weight of leaved twigs, needle weight, volume of leaved wigs and needle volume [
1].
The size of the assimilatory apparatus of the crown and its efficiency in the process of organic matter production has a primary effect on tree increment. Lemke [
2] stated a very strong correlation between the size of the assimilatory apparatus in pines and an increment in stem volume. This conclusion is essential for the rationalisation of decisions taken in forest economy, particularly breeding, forest management and utilisation. It is an indication for the practical application of such procedures, which by maintaining an optimal stand structure, taking into consideration the efficiency of the assimilatory apparatus—results in the maximum increment in tree volume. Growth efficiency depends on the size of the tree and the volume of needles. When determining the prescribed yield, we also need to take into consideration, next to the growing stock, the increment predicted for the coming period. The size of tree crowns is an element in the forecast of the increment capacity of trees and stands [
3].
Most studies concerning the assimilatory apparatus conducted to date focused on the determination of this parameter in the aspect of the stand, and not a single tree. Publications on forest productivity referred to productive biomass per hectare depending on, e.g., mean height, site index, age or degree of stocking. In this respect, several studies may be mentioned concerning the biomass of different stands, in a global aspect [
4,
5,
6,
7,
8], in the USSR [
9,
10], in Canada [
11], in the United States [
12,
13,
14,
15], for the following specific tree species: Norway spruce [
16], Scots pine [
17], compared to deciduous (beech) and coniferous (spruce) species [
18]. Attempts to determine the complicated structure of forest biomass using linear regression equations were undertaken by Usoltsev [
19] as well as Onuchin and Borisov [
20]. Later models took into consideration the total crown biomass [
21,
22]. Other components of biomass, such as the leaves and roots, were included when the models were made more precise by combining them with average characteristics of the stand [
23,
24]. Interesting studies on needle biomass in pine were conducted in Finland by Vanninen et al. [
25], Mäkelä and Vanninen [
26], Vanninen and Mäkelä [
27], Helmisari et al. [
28], Lehtonen [
29] and Muukkonen [
30]. Chroust [
31] developed empirical equations for leaf biomass in stands located in the former Czechoslovakia, while Heinsdorf and Krauß [
32] prepared dry biomass tables for Scots pine in Germany. Among other methods used to determine crown biomass, we need to mention a model proposed by Shinozaki et al. [
33,
34], using the stem diameter at crown base. Dry biomass in kg/ha for leaves (needles), twigs and the number of trees per hectare jointly and in 4-centimeter diameter subclasses, depending on site index and age of the stand, are given in “Tables of crown biomass for even-aged stands of
Picea abies in Switzerland” [
24]. The above-mentioned authors also evaluated crown biomass in forest tree species from the Ural Mountains and Kazakhstan [
35]. Studies on the determination of tree crown biomass were also conducted by Danilov [
36], Smirnov [
37], Babich [
38], Sokolov et al. [
39] and Sokolov and Kurbanov [
40].
The first tables facilitating an estimation of fresh needle mass in Scots pine were prepared by Burger [
41], on the basis of 210 trees coming from 58 stands. Studies on the size of the assimilatory apparatus, although on a much lesser scale, were also undertaken by Tiren [
42], Dengler [
43] and Badoux [
44]. The size of the assimilatory apparatus was typically characterised using the weight of fresh or dry needles, the number of needles or their upper area [
45]. Currently, research is underway on carbon content by tree species, biomass components and the impact of environmental factors on these values [
46,
47].
In Poland, an attempt to estimate the size of the assimilatory apparatus, approximated as the volume of leaved twigs in pine stands, was undertaken by Lemke [
48,
49]. His further studies [
50,
51] resulted in the development of “Tables for the estimation of needle weight and needled twigs in Scots pine” on sites typical of this species. These studies, conducted over a period of many years on a rich empirical material of 1808 trees from 33 stands, were crowned with the development of “Tables of the assimilatory apparatus in Scots pine (
Pinus sylvestris L.)” [
52] using regression equations as variables of the simple dimensional traits of trees (diameter at breast height and height) and their age. Socha and Wężyk [
53], on the basis of investigations conducted in forests located in Niepołomice, proposed allometric equations for the estimation of needle biomass in Scots pine.
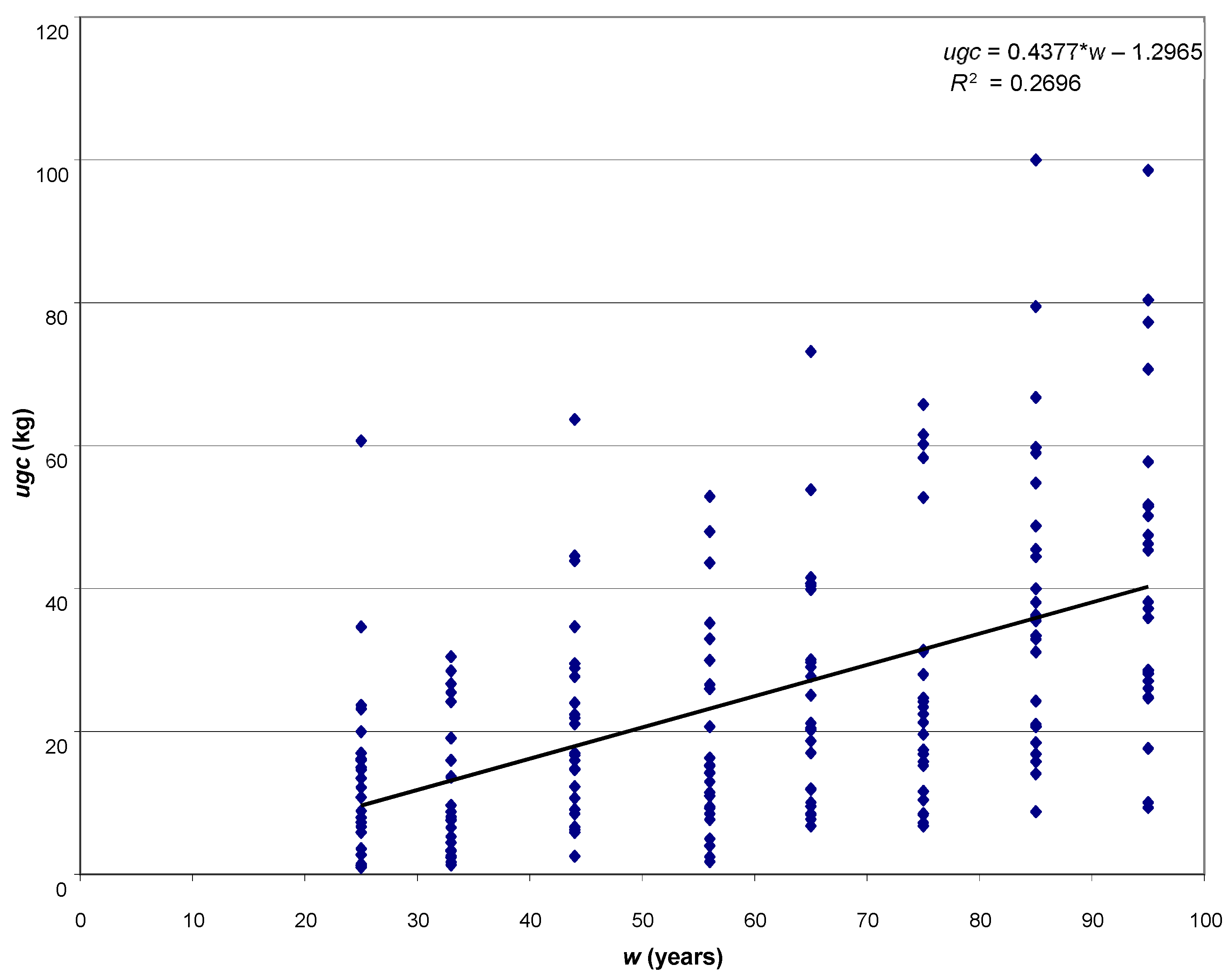
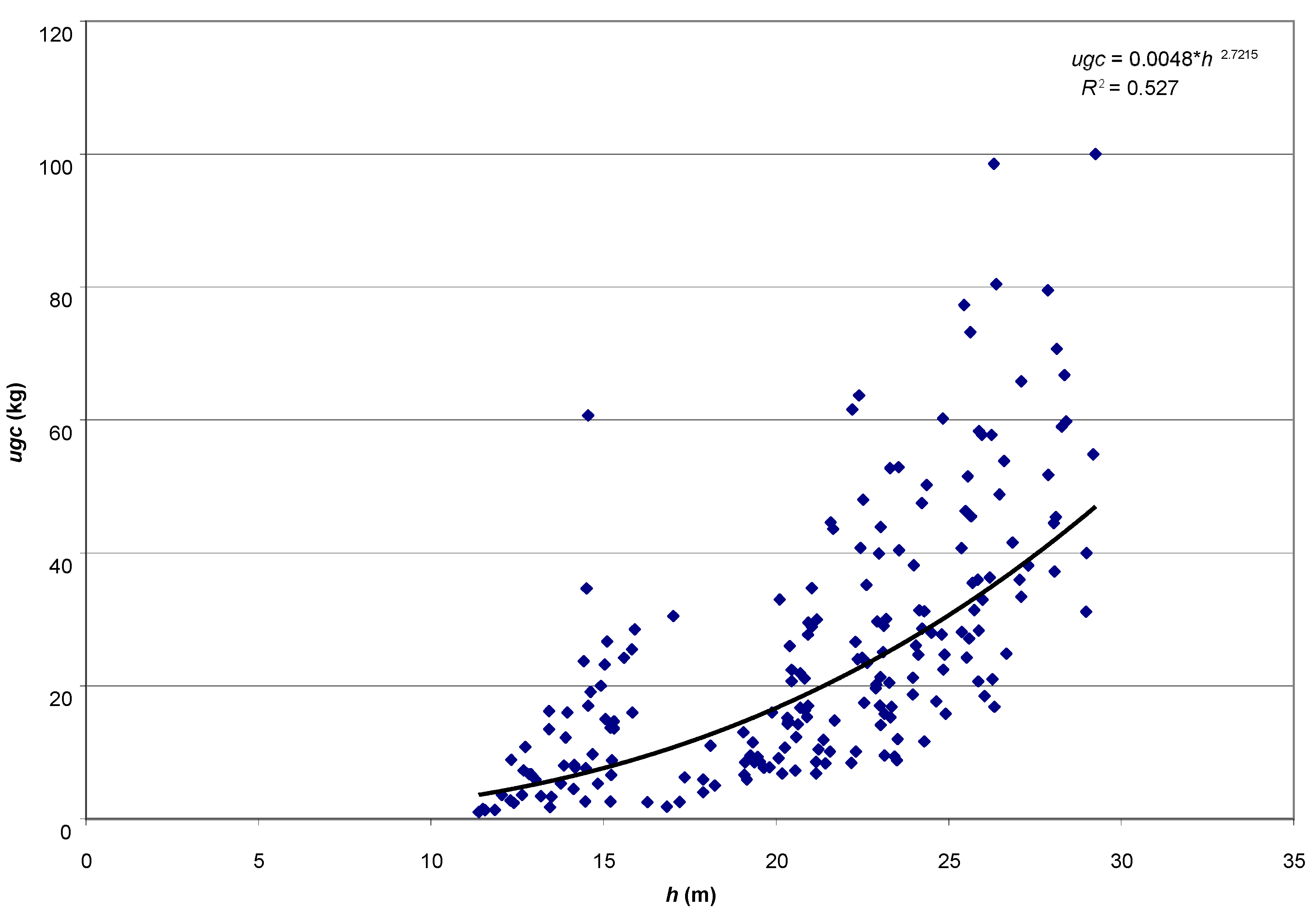
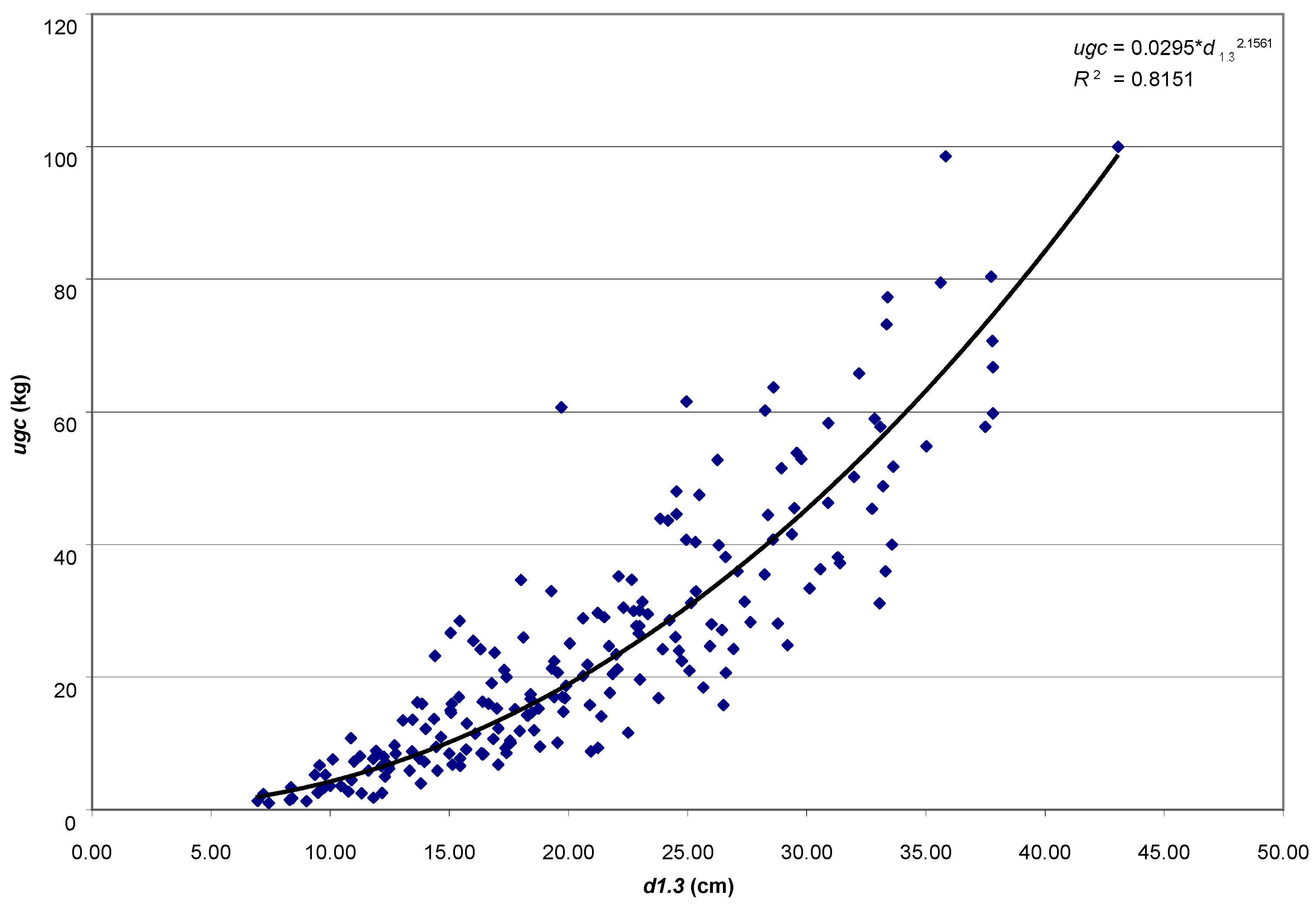
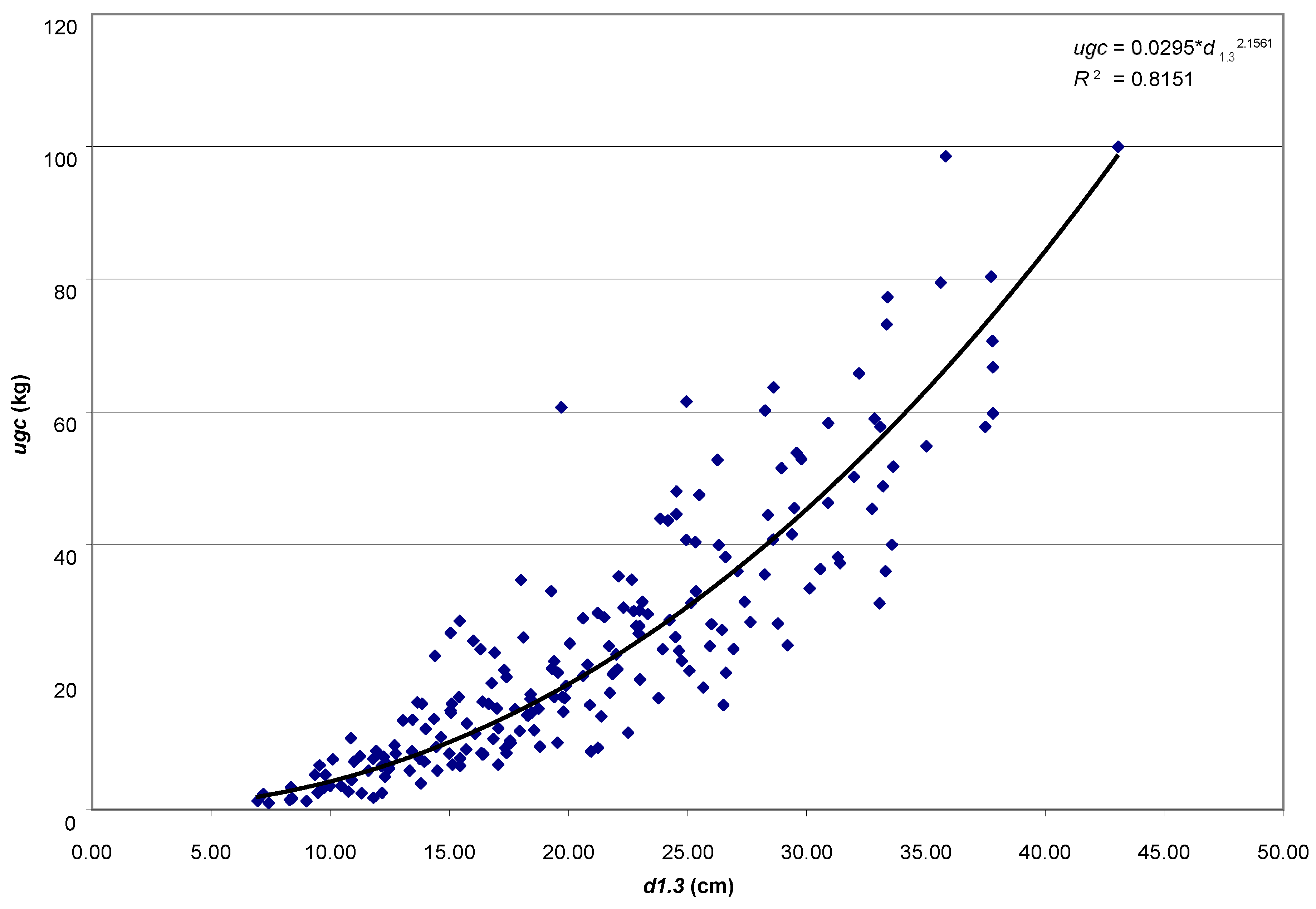
The aim of the study was to determine the size of the assimilatory apparatus of single pines: weight of leaved twigs (ugc), needle weight (ic), the volume of leaved twigs (ugo) and needle volume (io), and its relationship with selected dendrometric and increment traits of trees. A research hypothesis was made that the basic taxation features (age, height and diameter at breast height) will be strongly related to the weight of leaved twigs, thanks to which it will be possible to use them in the future to develop a local model of the Scots pine biomass fraction.
2. Material and Methods
The analytical material comprises results of measurements for 200 trees coming from 8 pure pine stands aged 25–95 years, growing on the fresh mixed coniferous forest habitat, in the Murowana Goślina Forest Experimental Station (52°34′ N, 17°00′ E) in western Poland. Average temperature in the five years before the study was 8.02 °C, and average annual rainfall was 544.2 mm.
In each stand, one plot with 200–350 trees was chosen. The location of a plot in a stand was supposed to minimize the non-age-related differences between trees. All plots created a trajectory of changes resulted from the chronosequence. On trees from all the 8 mean sample plots, the position of breast heights and the direction of north were permanently marked. The breast height diameter of the outside bark was measured crosswise from two directions of NS and EW rounded to 1 mm. The average diameter at breast height was calculated as the arithmetic mean of both these measurements rounded to 1 mm. Tree heights were measured in terms of diameter subclasses and levelled heights were determined on the basis of the height curve. For each stand, a total of 25 mean sample trees were selected. The number and dimensions of mean sample trees in diameter subclasses were established in accordance with the Draudt method [
54]. The number of model trees falling into each diameter subclass was proportional to the number of trees in each diameter subclass. Mean sample trees with appropriately shaped medium-sized crowns were selected. The size of crowns and their conditions status were estimated by sight. Total stem length was measured on felled trees accurate to 1 cm. Next, discs were cut from the length of 0.0 m, 1.30 m, as well as 0.5 m, 1.5 m, 2.5 m, and the end of the last complete section. If a verticil swelling or another deformation was found in the position from which a disc was to be collected, a disc was cut from a regular fragment located in the nearest vicinity. Such collected analytical material was adequately secured to be transported to the laboratory. The weight of the leaved twigs of the crown was determined on the basis of a direct measurement of this trait. From each crown, all needle-covered twigs were collected, and they were next weighed immediately after being cut off. For 200 mean sample trees, after the sample from each tree had been weighed, additionally needle volume, together with twigs, was determined using xylometry. Measurements were always taken at the date of sample collection. Samples of fresh twigs with needles were collected for laboratory analyses. From 25–56-, and 65–95-years-old trees, 20 or 10% of twigs were collected for laboratory analyses, respectively. The sample mass twigs with needles, not less than 2 kg, depended on the stand age. Prior to measurement samples labelled with information on the number of the mean sample plot and the number of the tree—were stored in plastic bags to prevent them from drying. Next, in the course of laboratory studies using a DPM 001 electronic increment meter (Vienna, Austria) according to K. Johann, the number of annual growth rings was determined, and diameters were measured accurate to 0.01 mm in 5-year periods on all discs coming from 200 mean sample trees. To analyse the stems, increment values of the following dendrometric traits were collected within 5- and 10-year periods: height, diameter at breast height, basal area at breast height and volume. Within the age classes, Pearson’s linear and multiple correlation and the corresponding significance levels were determined between the features of the assimilation apparatus (weight of leaved twigs (
ugc), needle weight (
ic), the volume of leaved twigs (
ugo), needle volume (
io)) and the basic dendrometric traits (height—
h and diameter at breast height—
d1.3) and with selected increments (height—
Ih5 and
Ih10, diameter at breast height—
Id5 and
Id10, basal area at breast height—
Ig5 and
Ig10, volume—
Iv5 and
Iv10). For all empirical material, the functional dependencies of the weight of leaved twigs on the age, height and diameter at breast height of trees were presented (power function, coefficient of determination, correlation coefficient). All analyses were performed in the Statistica ver. 13 (TIBCO Software Inc. 2017).
4. Discussion
The increment of trees depends, primarily, on the size of the assimilation apparatus of the crown and its efficiency to produce organic matter. The growth of individual forest trees declines with advancing age and the decline results from reduced resource use efficiency [
55]. Lemke [
2] stated a very strong correlation between the size of the assimilatory apparatus in pines and an increment in stem volume. When comparing results in this study with those reported from investigations conducted by Lemke [
48,
56] and Lemke and Kaźmierczak [
3], in pine stands of a similar age from 15 to 114, growing in the same habitat, fresh mixed coniferous forest, comparable dependencies of the size of the assimilatory apparatus with basic dendrometric traits and increment were observed in the trees. In the cited studies, a stronger relationship with the assimilatory apparatus was recorded in relation to the diameter at breast height in terms of height (correlation from 0.781 to 0.962), which was slightly weaker, as in this study (from 0.836 to 0.968). The strongest relationship with the weight of leaved twigs and needle weight was recorded for the increment in volume (correlation from 0.707 to 0.980), in this study (from 0.843 to 0.979); it was slightly weaker for the increment in basal area at breast height (from 0.656 to 0.959), in this study (from 0.852 to 0.984); followed by the increment in diameter at breast height (from 0.563 to 0.887), in this study (from 0.791 to 0.945) and the increment in height (from −0.115 to 0.816), in this study (from 0.021 to 0.758). In the case of the latter of the above-mentioned traits, the dependencies were statistically non-significant in some stands. The age of the stand generally did not have any effect on the power of the analysed dependencies.
The basic taxation features are strongly related to the weight of leaved twigs, thanks to which it is possible to use them to develop a local model of the Scots pine biomass fraction. The equations, based on universal features of standing trees (age, height and diameter at breast height), are simple in use and applicable for trees growing in stands, especially on sites typical for pine. The majority of allometric equations that can be found in the literature concern the dry biomass of the crown; only very few models predict the fresh mass of twigs with needles of individual pine trees [
52]. The only ones, based on the same independent variables as the one discussed above, are multinomial equations proposed by Lemke [
51] for use with different parameter sets for different age classes. In a publication, Socha and Wężyk [
53] proposed a wide set of equations based on many more independent variables e.g., basal area increment, absolute crown length and relative crown length.
The increasing demand for credible biomass and carbon estimates at widely varying spatial scales requires circumspect evaluations of the sampling and modelling practices used for biomass equation development. The accuracy of forest biomass estimates can be improved, including estimates of the aboveground live tree components as well as the dead and nontree fractions [
45].
5. Conclusions
A strong relationship between the size of the assimilatory apparatus and the basic dendrometric traits (height and diameter at breast height) confirms their applicability as variables in the development of allometric equations. In order to enhance the accuracy of regression equations, they should additionally include the age of trees as an independent variable. The application of these traits is of great practical importance, since they are determined within the framework of general stand valuations.
Taking into consideration the relationship of the size of the assimilatory apparatus with increment traits, we need to stress, in this context, a strong correlation with the increment in basal area at breast height (correlation coefficient of 0.8731 ÷ 0.9836). This trait is much easier to determine in the course of measurement work and at the same time it is much more objective in terms of the expected accuracy when compared with an increase in volume. In relation to the diameter at breast height increment (correlation coefficient of 0.7835 ÷ 0.9581), after the measurement of the diameter at breast height and its increment, it requires only simple mathematical transformations. For the above-mentioned reasons, the increment in breast height section area is predisposed to be commonly used in the determination of the efficiency of the assimilatory apparatus of trees. In contrast, increment in height may not be used for this purpose.
Characteristics of the assimilatory apparatus show a similar relationship with the analysed types of increment in trees. Thus, due to the lower consumption of labour and expenses, studies should be based on the weight of leaved twigs.


 ,
, 

{kind=link}
{kind=link}
{kind=link}