
Analytical Calculations of Some Effects of Tidal Forces on Plants on the International Space Station


{kind=link}
{kind=link}
Abstract
1. Why Can Variations in Gravity Affect Plants on the International Space Station?
1.1. Review of Some Experiments on Plants on the ISS
1.2. The ISS Can Be Seen as a Giant Swing along Its Motion in Earth-Orbit
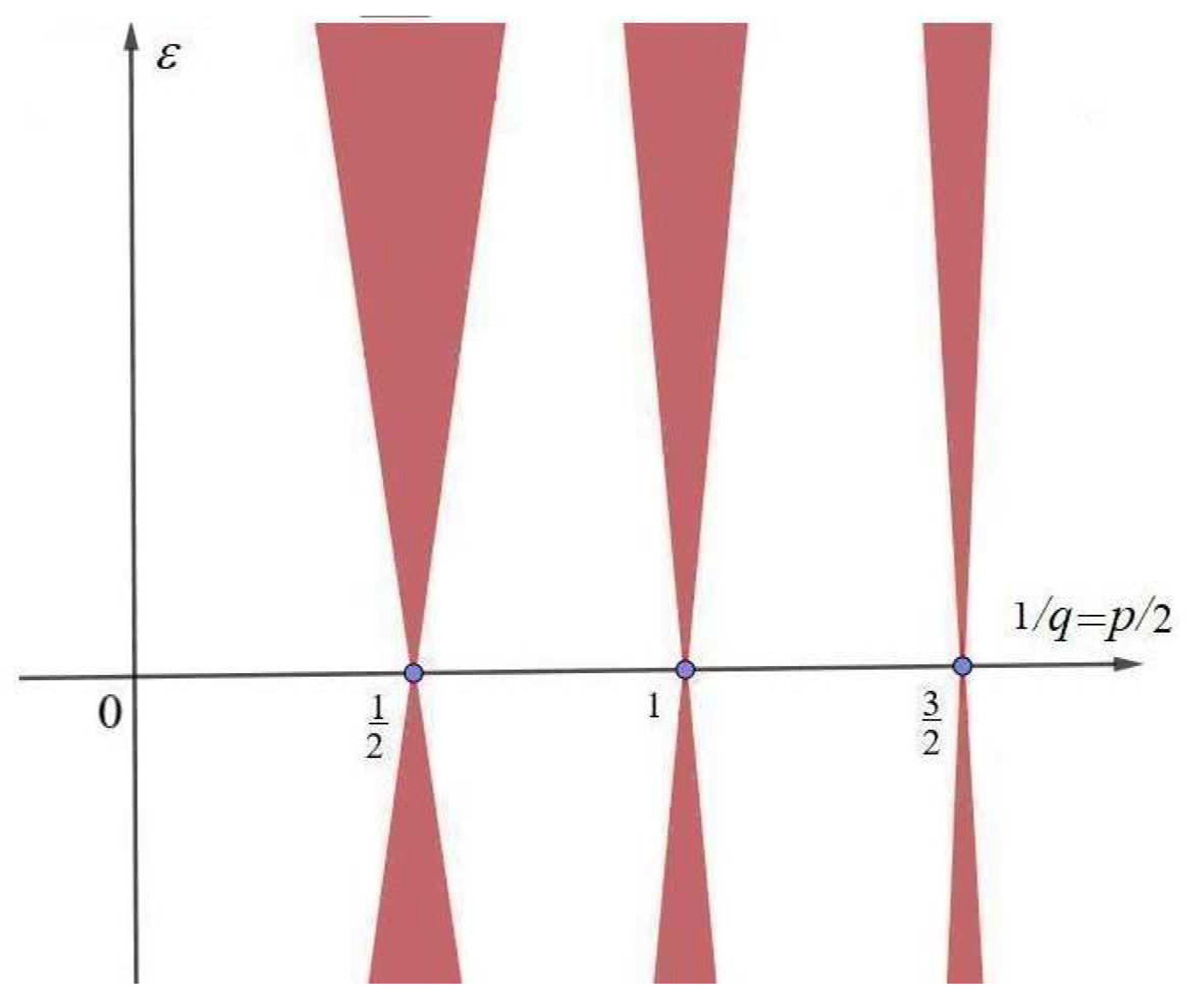
2. Analytical Method and Parametric Resonance
2.1. Parametric Resonance
2.2. Plant Experiments on the Earth
2.3. Plant Experiments Inside the ISS
2.4. Remarks and Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moran, N. Osmoregulation of leaf motor cells. FEBS Lett. 2007, 581, 2337–2347. [Google Scholar] [CrossRef]
- Miles, L.E.M.; Raynal, D.M.; Wilson, M.A. Blind man living in normal society has circadian rhythms of 24.9 h. Science 1977, 198, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Gluckman, B.J.; Netoff, T.I.; Neel, E.J.; Ditto, W.L.; Spano, M.L.; Schiff, S.J. Stochastic resonance in neuronal network from mammalian brain. Phys. Rev. Lett. 1996, 77, 4098–4101. [Google Scholar] [CrossRef]
- Bevington, M. Lunar biological effects and the magnetosphere. Pathophysiology 2015, 22, 211–222. [Google Scholar] [CrossRef]
- Bünning, E. The Physiological Clock. In Circadian Rhythms and Biological Chronometry, 3rd ed.; English Universities Press: London, UK, 1973. [Google Scholar]
- Klein, G. Farewell to the Internal Clock. In A Contribution in the Field of Chronobiology; Springer: New York, NY, USA, 2007. [Google Scholar]
- Fisahn, J.; Yazdanbakhsh, N.; Klingelé, E.; Barlow, P.W. Arabidopsis root growth kinetics and lunisolar tidal acceleration. New Phytol. 2012, 195, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Konopliv, A.S.; Binder, A.B.; Hood, L.L.; Kucinskas, A.B.; Sjogren, L.; Williams, J.G. Improved gravity field of the moon from lunar prospector. Science 1998, 281, 1476–1480. [Google Scholar] [CrossRef] [PubMed]
- Sorbetti-Guerri, F.; Michel, D. Tree stem diameters fluctuate with tide. Nature 1998, 392, 665–666. [Google Scholar]
- James, A.B.; Monreal, J.A.; Nimmo, G.A.; Kelly, C.L.; Herzyk, P.; Jenkins, G.I.; Nimmo, H.G. The circadian clock in arabidopsis roots is a simplified slave version of the clock in shoots. Science 2008, 322, 1832–1835. [Google Scholar] [CrossRef]
- Zajaczhowska, U.; Barlow, P.W. The effect of lunisolar tidal acceleration upon stem elongation growth, nutations and leaf movements in peppermint (Menta × piperita L.). Plant Biol. 2017, 19, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Crossley, D.; Hinderer, J.; Boy, J.P. Time variations of the European gravity field from superconducting gravimeters. Geophys. J. Int. 2005, 161, 257–264. [Google Scholar] [CrossRef]
- Barlow, P.W.; Fisahn, J. Lunisolar tidal force and the growth of plant roots, and some other of its effects on plant movements. Ann. Bot. 2012, 110, 301–318. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fisahn, J. Control of plant leaf movements by the lunisolar tidal force. Ann. Bot. 2018, 121, e1–e6. [Google Scholar] [CrossRef]
- Fisahn, J.; Barlow, P.; Dorda, G. A proposal to explain how the circatidal rhythm of the Arabidopsis thalania root elongation rate could be mediated by the lunisolar gravitational force: A quantum physical approach. Ann. Bot. 2018, 122, 725–733. [Google Scholar]
- Fisahn, J.; Klingelé, E.; Barlow, P.W. Lunar gravity affects leaf movements of Arabidopsis thalania in the International Space Station. Planta 2015, 241, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Brinckmann, E. ESA hardware for plant research on the International Space Station. Adv. Space Res. 2005, 36, 1162–1166. [Google Scholar] [CrossRef]
- Gouin, H. Introduction to Mathematical Methods of Analytical Mechanics; Elsevier & ISTE Editions: London, UK, 2020. [Google Scholar]
- Arnold, V.I. Ordinary Differential Equations; Springer: Berlin, Germany, 1992. [Google Scholar]
- de Gennes, P.G.; Brochard-Wyart, F.; Quéré, D. Capillarity and Wetting Phenomena: Drops, Bubbles, Pearls, Waves; Springer: New York, NY, USA, 2004. [Google Scholar]
- Mattia, D.; Gogotsi, Y. Review: Static and dynamic behavior of liquids inside carbon nanotube. Microfluid-Nanofluid 2008, 5, 289–305. [Google Scholar] [CrossRef]
- Taylor, G.I. The instability of liquid surfaces when accelerated in a direction perpendicular to their planes. Proc. R. Soc. Lond. 1950, A201, 192–196. [Google Scholar]
- Mellor, G.L. Introduction to Physical Oceanography; Springer: Berlin, Germany, 1996. [Google Scholar]
- Tavenner, E. The Roman Farmer and the Moon, Transactions and Proceedings of the American Philological Association; The Johns Hopkins University Press: Baltimore, MD, USA, 1918; Volume 49, pp. 67–82. [Google Scholar]
- Gouin, H. The watering of tall trees—Embolization and recovery. J. Theor. Biol. 2015, 369, 42–50. [Google Scholar] [CrossRef]
- Brouwer, D.; Clemence, G.M. Methods of Celestial Mechanics; Academic Press: New York, NY, USA, 1961. [Google Scholar]
- Olson, D.H.; Jacobs, J.W. Experimental study of Rayleigh-Taylor instability with a complex initial perturbation. Phys. Fluids 2009, 21, 034103. [Google Scholar] [CrossRef]
- Mishra, A.A.; Hasan, N.; Sanghi, S.; Kumar, R. Two-dimensional buoyancy driven thermal mixing in a horizontally partitioned adiabatic enclosure. Phys. Fluids 2008, 20, 063601. [Google Scholar] [CrossRef]
- Gouin, H. Influence of lunisolar tides on plants. Parametric resonance induced by periodic variations of gravity. Phys. Fluids 2020, 32, 101907. [Google Scholar] [CrossRef]
- Longman, I.M. Formulas for computing the tidal accelerations due to the Moon and the Sun. J. Geophys. 1959, 64, 2351–2355. [Google Scholar] [CrossRef]
- Boffetta, G.; Mazzino, A. Incompressible Rayleigh–Taylor turbulence. Annu. Rev. Fluid Mech. 2017, 49, 119. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gouin, H. Analytical Calculations of Some Effects of Tidal Forces on Plants on the International Space Station. Forests 2021, 12, 1443. https://doi.org/10.3390/f12111443
Gouin H. Analytical Calculations of Some Effects of Tidal Forces on Plants on the International Space Station. Forests. 2021; 12(11):1443. https://doi.org/10.3390/f12111443
Chicago/Turabian StyleGouin, Henri. 2021. "Analytical Calculations of Some Effects of Tidal Forces on Plants on the International Space Station" Forests 12, no. 11: 1443. https://doi.org/10.3390/f12111443
APA StyleGouin, H. (2021). Analytical Calculations of Some Effects of Tidal Forces on Plants on the International Space Station. Forests, 12(11), 1443. https://doi.org/10.3390/f12111443