Abstract
Quantifying the phenological variations of Populus euphratica Olivier (P. euphratica) resulting from climate change is vital for desert ecosystems. There has previously been great progress in the influence of climate change on vegetation phenology, but knowledge of the variations in P. euphratica phenology is lacking in extremely arid areas. In this study, a modified method was proposed to explore P. euphratica phenology and its response to climate change using 18-year Global Land Surface Satellite (GLASS) leaf area index (LAI) time series data (2000–2017) in the upper Tarim River basin. The start of the growing season (SOS), length of the growing season (LOS), and end of the growing season (EOS) were obtained with the dynamic threshold method from the reconstructed growth time series curve by using the Savitzky–Golay filtering method. The grey relational analysis (GRA) method was utilized to analyze the influence between the phenology and the key climatic periods and factors. Importantly, we also revealed the positive and negative effects between interannual climate factors and P. euphratica phenology using the canonical correlation analysis (CCA) method, and the interaction between the SOS in spring and EOS in autumn. The results revealed that trends of P. euphratica phenology (i.e., SOS, EOS, and LOS) were not significant during the period from 2000–2017. The spring temperature and sunshine duration (SD) controlled the SOS, and the EOS was mainly affected by the temperature and SD from June–November, although the impacts of average relative humidity (RH) and precipitation (PR) on the SOS and EOS cannot be overlooked. Global warming may lead to SOS advance and EOS delay, and the increase in SD and PR may lead to earlier SOS and later EOS. Runoff was found to be a more key factor for controlling P. euphratica phenology than PR in this region.
1. Introduction
Vegetation phenology is the research of events in the plant life cycle, and how these events adapt to climate changes [1,2], which makes it the most intuitive and sensitive biological indicator of climate change. Climate change, such as modified precipitation, increasing temperature, and increasing frequency of extreme weather events, will change the vegetation phenology [3], which will have extensive effects on plant community structure, plant distribution, energy cycles, vegetation ecosystems, and primary production of vegetation [4,5,6,7,8]. Altered phenological timing, furthermore, may have negative effects on species interactions in the food chain, such as migration, survival, reproduction, and occupying feeding habits [9,10]. Therefore, a better understanding of how vegetation phenology responds to climate change can improve the accuracy of phenological simulation models and provide a scientific reference for future management of vegetation ecosystems.
Plant growth, including leaf unfolding, flowering, fruiting, and leaf senescence, has been recorded to reflect the phenological characteristics of vegetation based on ground monitoring sites of phenology. However, this method has some limitations in the spatio-temporal scale and biome scale [11]. Therefore, the remote sensing technique for phenological monitoring, which is used to simulate various logistic regression models through long-term series remote sensing data and to extract vegetation phenology (i.e., the start of the growing season (SOS), length of the growing season (LOS), and end of the growing season (EOS)) from suitable models, has been widely applied to explore the spatial–temporal variations in vegetation phenology at the regional and global scales [12,13,14,15,16,17]. However, there is a wide range of mixed pixels in satellite remote sensing data, and the spatial resolution of remote sensing data will affect the accuracy of vegetation phenological feature inversion, especially in the extreme arid areas where the vegetation distribution is more sparse and the coverage is lower. In addition, previous studies on vegetation phenology have focused on plant phenology change trends [18,19,20], phenology changes in different vegetation types [21], temperature [22,23,24], rainfall [25,26,27], aspect [28], altitude [23], solar radiation [29], and other single climatic factors affecting vegetation phenology. Therefore, there is still a need to demonstrate the relationship of the preseason, interannual, and multi-climatic factors with phenology, and what kind of interaction exists between the SOS and EOS.
Arid and semi-arid ecosystems are important parts of Earth’s climate system, covering 41% of the Earth’s land surface [30,31,32]. These regions are characterized by harsh environments and little rainfall, especially in extremely arid areas, where the average annual rainfall is often <60–100 mm [33]. The Tarim River basin is located in the hinterland of Eurasia, an extremely arid area. This region is highly vulnerable and sensitive to climate change due to the particular features comprising mountains and a mosaic of inner-mountain basins [34]. Populus euphratica (Salicaceae), an ancient, precious, endangered species, is a constructive tree in extremely arid areas [34]. P. euphratica is a vital part of the extreme drought ecosystem in this region due to its advantageous characteristics such as cold resistance, heat resistance, drought resistance, wind–sand resistance, and salt–alkali resistance [35]. In the upper Tarim River basin, since there it is less affected by the Ecological Water Conveyance Project than the lower Tarim River, the results on the response of P. euphratica phenology to climate change are more believable. However, research in this field mainly focuses on the relationship between vegetation and groundwater [34,36], changes in rainfall and temperature under climate change and global warming, its impact on water resources [37,38,39,40,41], and the impact of water on physical and chemical indicators of vegetation growth [30,32,42,43]. To date, few, if any, studies have revealed the dynamics of P. euphratica phenology and its response to climate change in arid areas.
The novelty of this study is that a modified method was proposed to explore P. euphratica phenology and its response to climate change using 18-year Global Land Surface Satellite (GLASS) leaf area index (LAI) time series data (2000–2017) in the upper Tarim River basin. A sandwich spatial sampling strategy, which can reduce the accuracy error caused by many mixed pixels in satellite remote sensing data, was used to extract the LAI information of P. euphratica from the GLASS LAI time series data, and the phenological information (i.e., SOS, LOS, EOS) was obtained with the dynamic threshold method from the reconstructed growth time series curve by using the Savitzky–Golay filtering method. The grey relational analysis (GRA) and canonical correlation analysis (CCA) methods were utilized to explore the relationship between phenology and multi-climatic factors. Specifically, the main objectives of this study were to (1) quantitatively reveal the change trends of P. euphratica phenology; (2) identify the key meteorological factors and key periods that affect the phenology of P. euphratica, and (3) confirm whether the beginning and the end of the P. euphratica growing season influence each other. The results provide insights into quantitative assessment of interannual variations of P. euphratica phenology under climate change and provide a scientific reference for future management of the desert ecosystem.
2. Study Area and Data
2.1. Study Area
Through field investigation, P. euphratica was found to be mainly distributed along the upper reaches of the Tarim River and the Yerqiang River. Therefore, this investigation mainly focused on the upper Tarim River and the Yeerqiang River basin, which was divided into two regions (Figure 1b): Bachu (BC) and Shaya (SY). The study area experiences a typical temperate arid continental climate, with long sunshine duration, strong annual evaporation, and low annual rainfall. The extreme minimum temperature is −25 °C, the extreme maximum temperature is 39.4 °C, and the annual average temperature is 10.4 °C [34]. The annual average precipitation is 30–50 mm, and the annual potential evaporation is 2000–2900 mm [43].
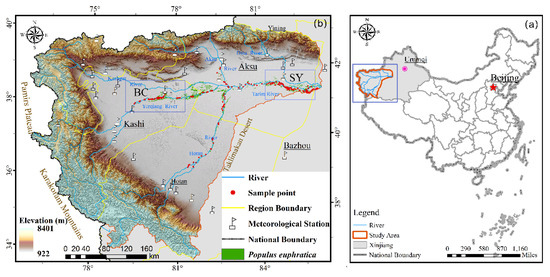
Figure 1.
(a) Study area. (b) Sample points, meteorological stations, and typical sample area of Bachu (BC) and Saya (SY) regions.
2.2. Data
The Global Land Surface Satellite (GLASS) leaf area index (LAI) has a temporal resolution of eight days and spans the period from 1981–2017. The LAI product from 1982–1999 was developed from AVHRR reflectance with a spatial resolution of 0.05°, and that from 2000–2017 with a spatial resolution of 1 km was derived from MODIS reflectance [44]. The GLASS LAI data product is a national key project, and its dataset is superior to other similar products in terms of spatial and temporal accuracy [45]. This product feature has significant advantages in the continuity of its time series and the integrity of its spatial scope [46]. Moreover, the GLASS LAI dataset has passed the verification analysis of the measurement data of the common international site, with the results demonstrating that the verification accuracy is higher than other similar products [47,48]. Since P. euphratica has a small distribution area and low vegetation coverage, this study used the GLASS 1 km resolution data, which were from the National Earth System Science Data Sharing Infrastructure, National Science & Technology Infrastructure of China (http://www.geodata.cn (accessed on 14 October 2019)). A total of eighteen years of data from 2000–2017 were obtained, and all the LAI values were the original values of the GLASS LAI dataset, which were equal to 10 times the actual values.
This study used climatic data, including sunshine duration (SD), average air temperature (AT), average land surface temperature (ST), effective accumulated temperature (EA), average relative humidity (RH), and precipitation (PR). These data were derived from http://data.cma.cn/ (accessed on 14 July 2021) from 1999–2018. Annual runoff data were derived from the Statistical Bulletin of the Ministry of Water Resources of the People’s Republic of China (http://www.mwr.gov.cn/sj/#tjgb (accessed on 14 July 2021)). Additionally, in this study, based on Landsat TM/OLI long time series remote sensing data with 30 m resolution from 1990–2015, the distribution data of P. euphratica for 1990, 1995, 2000, 2005, 2010, and 2015 were extracted using the object-oriented classification method. These data were then used to select typical sample sites of P. euphratica.
3. Methods
The technique flowchart of this study is shown in Figure 2, the research steps were as follows. (1) P. euphratica samples were selected and extracted from LAI time series data using the sandwich spatial sampling method. The growth time series curve of P. euphratica was then reconstructed using the Savitzky–Golay filtering method. (2) The phenological features of P. euphratica, including the SOS, EOS, and LOS, were extracted using the dynamic threshold method. (3) The grey relational analysis (GRA) method was used to analyze the correlations between climatic factors and the SOS during three periods (spring, SOS preseason, and annual), and the EOS during three periods (autumn, EOS preseason, and annual). (4) The canonical correlation analysis (CCA) method was used to explore the positive and negative effects between phenology (SOS, EOS) and interannual climatic factors (SD, AT, ST, EA, RH, and PR).
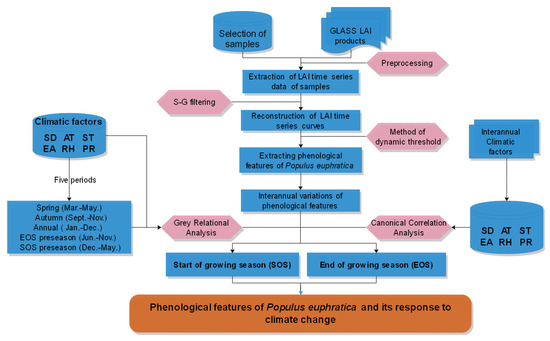
Figure 2.
Schematic diagram of the extraction processes of henological features of P. euphratica. Abbreviations: Leaf area index (LAI), Global Land Surface Satellite (GLASS), sunshine duration (SD), average air temperature (AT), average land surface temperature (ST), effective accumulated temperature (EA), average relative humidity (RH), precipitation (PR), start of the growing season (SOS), end of the growing season (EOS), Savitzky-Golay (S-G) filtering.
3.1. Extract LAI Features
Based on the distribution data of P. euphratica in 1990, 1995, 2005, 2010, and 2015, the regional composition sample data of P. euphratica, which fully met the 1 km2 area criterion, were automatically identified by a 1 km × 1 km square window. The typical sample points were selected from high-resolution imagery (e.g., Google Earth imagery, Version 2018) by the Sandwich Spatial Sampling tool, which is developed based on spatial optimization sampling strategy of Trinity theory. The LAI information was extracted from each sample using the GLASS LAI products from 2000–2017.
3.2. Filtering and Reconstruction of LAI Curves
Although the GLASS LAI products have been processed to remove noise, the time series information of the extracted sample points still exhibits the “pseudo” peak phenomenon. Thus, the Savitzky-Golay (S-G) filtering method was utilized to remove the noise and reconstruct the LAI curves of P. euphratica. S-G filtering can be used with the time series index data of different time scales and is not limited by sensor differences, so the noise in the original data can be removed using the minimum root mean square error [49].
where Yj is the resultant LAI value, Yj is the original LAI value, Ci is the coefficient for the LAI value of the filter, and N is the number of convoluting integers and is equal to the smoothing window size (2 m + 1).
3.3. Dynamic Threshold Method
The dynamic threshold method is widely utilized to extract phenological features of vegetation because it has better spatio-temporal applicability [50,51]. In this method, the SOS is defined as the time when the vegetation LAI increases by a certain percentage, while the EOS is defined as the time when the vegetation LAI decreases by a certain percentage. The threshold used in this method is not a specific value, but a dynamic ratio. When the dynamic threshold method was proposed to extract phenological features, Jönsson and Eklundh [52] suggested that the threshold at the SOS and EOS is 0.2, which has been subsequently used by many scholars in phenological research. However, the choices of threshold values depend on different plant species and different regions, so we had to proceed cautiously and rigorously when selecting thresholds. In this study, we combined remote sensing data with 2016 ground-based monitoring data and used the following procedure illustrated by the flow chart in Figure 3. The SOS and EOS values were obtained using various dynamic thresholds (i.e., A1, A2) ranging from 0.1–0.4 at 0.02 intervals. Then, the results were verified by using the ground-based monitoring data (Table 1). In terms of these tests, we opted to use the thresholds of A = 0.2 and A2 = 0.3 to extract the SOS and EOS of P. euphratica phenology in the study area from 2000 to 2017, respectively.
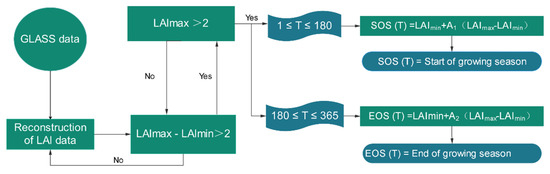
Figure 3.
Flow chart of the dynamic threshold method used to extract the phenological features of P. euphratica.

Table 1.
Phenology (i.e., SOS and EOS) extracted from different thresholds (A1/A2) and ground-based monitoring data in 2016.
3.4. Grey Relational Analysis (GRA)
The relationship between many factors falls within a gray area, due to the limitations of people’s understanding and the complexity of geographical phenomena, and it is difficult to objectively measure the degree of relevance by the relative correlation coefficient method [53]. The grey relational analysis method makes up for the above shortcomings. GRA is not limited by sample type and probability distribution, so it is a new method to investigate uncertain problems under limited information and data [54,55]. Details of the principles and computation steps of the PNPI were presented in Li et al. [53]. In this study, the SOS is used as the reference factor sequences from 2000 to 2017, and the comparative factor sequence consists of climatic factors (i.e., SD, AT, ST, EA, RH, and PR) during three periods (spring, SOS preseason, and annual) from 2000 to 2017. The EOS from 2000 to 2017 is used as the reference factor sequence, and the comparative factor sequence consists of climatic factors during three periods (autumn, EOS preseason, and annual). The correlations between climatic factors and the P. euphratica phenology during different periods were explored.
3.5. Canonical Correlation Analysis (CCA)
Canonical correlation is a statistical analysis method used to study the correlation between two groups of variables and was first proposed by Hotelling in 1935 [56]. It is also a dimensionality reduction technology, which is widely used in various fields [57]. Vegetation phenology is not only related to climatic factors, but also the physiological characteristics of vegetation. The change in climatic factors may not lead to one-way phenological changes due to the interaction effects between the SOS in spring and the EOS in autumn [8,58]. It is not rigorous to use simple climatic data to describe these changes. Therefore, the first set of variables (X1) consists of the EOS and SOS from 2000 to 2017 in this study, and the interannual climatic factors (SD, AT, ST, EA, RH, and PR) during the period of 2000–2017 as the second set of variables (Y1). The CCA method not only revealed the relationship between SOS and EOS, but also explored the correlations between the P. euphratica phenology and interannual climate factors. The principle and computation steps are detailed in Hotelling [56].
4. Results
4.1. Monthly and Interannual Variations of Climate Variables
Figure 4 presents the monthly and interannual variations in SD, AT, ST, EA, RH, and PR time series from 1999–2018 in the upper Tarim River basin (i.e., BC and SY). Generally, the monthly and interannual variations trend of SD, AT, ST, EA, RH, and PR are consistent in BC and SY regions, which have a typical temperate arid continental climate with strong annual evaporation and low annual rainfall. The PR was scarce and mostly concentrated from June to September with the highest value of 79.00 mm in July 2002 and 57.80 mm in August 2016 in BC and SY regions, respectively. The annual PR of BC and SY ranged from 8.70 mm (1999) to 183.90 mm (2010) and 24.80 mm (2007) to 107.00 mm (2016), respectively. The RH peaked in December, and the annual average RH was 44.42% (2018) and 59.83% (2003). The AT peaked in July, and the highest average AT of BC and SY was 28.27 °C (July 2015) and 28.28 °C (July 2015), respectively. From 1999–2018, the annual average AT was the highest (13.44 °C) in 2016 and the lowest (11.27 °C) in 2012 in this study area. The annual average ST ranged from 14.90 °C (2011) to 17.12 °C (2006). The annual SD ranged from 2574.60 h (2010) to 3077.00 h (2011) in the upper Tarim River basin, and the annual EA was 4420.50–5055.90 °C. The study showed that there was no significant trend in annual PR, SD, and EA, and annual average AT, EA, and ST from 1999–2018.
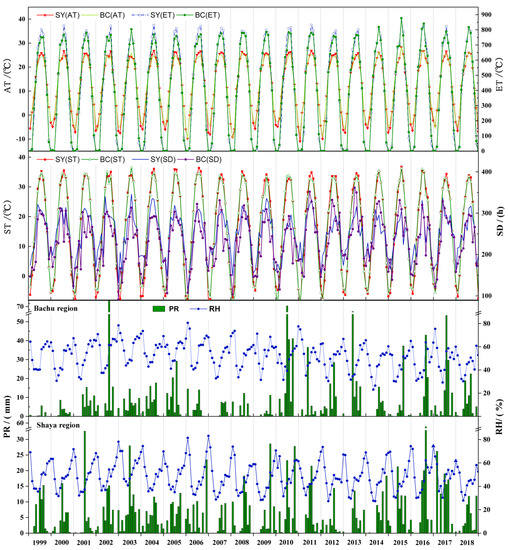
Figure 4.
Sunshine duration (SD), average air temperature (AT), average land surface temperature (ST), effective accumulated temperature (EA), average relative humidity (RH), and precipitation (PR) time series from 1999–2018 in Bachu region (BC) and Shaya region (SY).
4.2. Interannual Variations of P. euphratica Phenology
Interannual changes and P. euphratica phenology (SOS, LOS, EOS) are shown in Figure 5 in the upper Tarim River basin from 2000–2017. Generally, the upper Tarim River basin showed advancing SOS from 2000–2017 and the EOS of P. euphratica was advanced, but a slight lengthening in LOS was observed in the study area. However, it was found that the interannual variation trends of the EOS, LOS, and SOS derived from GLASS LAI data were not significant in the study time series. It is reasonable that a longer study time series for GLASS LAI data should be considered to research P. euphratica phenology under global climate change.
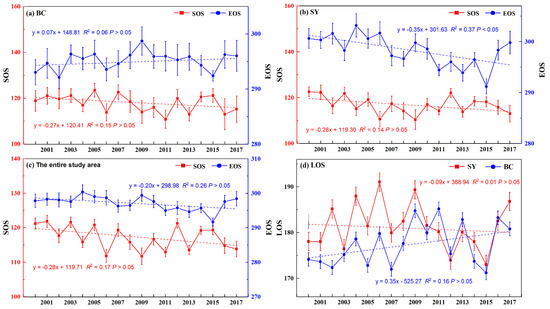
Figure 5.
Interannual changes in P. euphratica phenology (SOS, LOS, and EOS) in the upper Tarim River basin from 2000–2017. (a) Bachu region (BC), (b) Shaya region (SY), (c) entire study area, (d) LOS of BC and SY.
As demonstrated in Figure 5a–c, the SOS of P. euphratica of BC and SY was advanced from 2000–2017, the average SOS peaked in 2005 (124th, early May), and the troughs of average SOS appeared in 2011 (111st, late April) in the BC regions. In the SY regions, however, the mean SOS of P. euphratica was the latest (123rd, early May) in 2000 and the earliest (111st, late April) in 2009. The 18-year average SOS of P. euphratica was on the 117th day (late April) in the study area. BC showed delayed EOS, but this was different from SY. The mean EOS of P. euphratica was the latest (299th, late October) in 2009 and the earliest (292nd, late October) in 2002 in BC regions from 2000–2017. In the SY regions, the average EOS of P. euphratica peaked in 2004 (303rd, early November) and the troughs of average EOS appeared in 2015 (291st, late October). The 18-year average EOS of P. euphratica was on the 297th day (late October) in the upper Tarim River basin. In Figure 5d, the LOS of P. euphratica differs among the different regions (BC and SY) with the rank order based on P. euphratica LOS being SY > BC, and the 18-year average LOS of BC and SY regions were 177 and 181 days, respectively.
Overall, the spatial heterogeneity of the P. euphratica phenology was higher, and the changing trend of P. euphratica phenology varied in different regions (BC and SY). BC showed delayed EOS, but in SY it was significantly advanced. The P. euphratica phenology varied among different years in the same region, and the spatial heterogeneity of P. euphratica phenology in the same year and region (BC or SY) was higher. This is reasonable since climate, which is variable and complex, fundamentally determines the phenology and interannual change trends of phenology.
4.3. Interannual Variations in P. euphratica Phenology
4.3.1. Effects of Key Climatic Periods and Factors
The GRA between phenology (SOS and EOS) and climatic factors (SD, AT, ST, EA, RH, and PR) is presented in Figure 6. The SOS of P. euphratica was mainly affected by spring climatic factors, while the EOS of P. euphratica was mainly affected by the EOS preseason (June–November) meteorological factors. The SOS of P. euphratica was mainly affected by the climatic factors (0.81) in spring (March–May) (Figure 6a), and less affected by the annual (January–December) climatic factors (0.75) or the SOS preseason (December–May) climatic factors (0.67) (Figure 6c,e). Moreover, the influence of spring climatic factors in the different regions (BC and ALE) on the SOS of P. euphratica was greater than that of the annual climatic factors or the SOS preseason climatic factors.
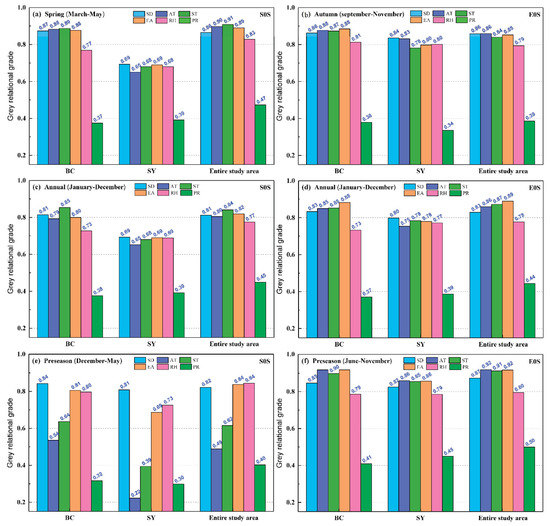
Figure 6.
Grey relational analysis between phenology (SOS and EOS) and climatic factors (SD, AT, ST, EA, RH, and PR). (a) GRA between the SOS and climatic factors in spring (March–May), (b) GRA between the EOS and climatic factors in autumn (September–November), (c) GRA between the SOS and the annual (January–December) climatic factors, (d) GRA between the EOS and the annual (January–December) climatic factors, (e) GRA between the SOS and the SOS preseason (December–May) climatic factors, (f) GRA between the EOS and the EOS preseason (June–November) climatic factors.
The EOS of P. euphratica was mainly affected by the EOS preseason (June–November) meteorological factors (0.82) (Figure 6f), and less affected by the annual (January–December) climatic factors (0.78) or the meteorological factors (0.77) in autumn (September–November) (Figure 6b,d). Moreover, the influence of autumn climatic factors on the EOS of P. euphratica was greater than that of the annual climatic factors or the EOS preseason meteorological factors in the BC and SY regions. It indicated that spring climatic factors (March–May) influence the SOS of P. euphratica more than climatic factors in other periods (i.e., annual and preseason), and the EOS is more affected by the variations in climatic factors from June to November.
Different climatic factors had different effects on phenology (Figure 6). The effects of average ST (0.91), average AT (0.90), average EA (0.89), and SD (0.86) on the SOS of P. euphratica were greater than those of average RH (0.82) and precipitation (0.47). The effects of average AT (0.86), SD (0.86), average EA (0.85), and average ST (0.84) on the EOS of P. euphratica were greater than those of average RH (0.79) and precipitation (0.39). Therefore, we could infer that temperature (i.e., AT, ST, and EA) and SD controlled P. euphratica phenology, although the impacts of RH and PR on P. euphratica phenology cannot be overlooked in the upper Tarim River basin.
4.3.2. Exploration of the Relationships between SOS, EOS, and Interannual Climate
Figure 7 shows the CCA patterns between phenology (SOS and EOS) and climatic factors (SD, AT, ST, EA, RH, and PR). A total of two pairs of canonical variables were extracted, the first of which (U1, V1) was statistically significant (p < 0.05). The results showed that the first pair of canonical correlation variables revealed a clear correlation between phenology and climatic factors, as indicated by the canonical correlation coefficient of 0.63. The canonical variable U1 had a significant positive correlation with SOS but a significant negative correlation with EOS, with typical load coefficients of −0.26 and 0.99, respectively. The typical variable V1 had positive correlations with average RH with the typical load coefficient of 0.24. V1 had negative correlations with PR, average ST, average AT, EA, and SD, however, with typical load coefficients of −0.26, −0.22, −0.63, −0.87, −0.17, respectively. These results generally indicated that the SOS had negative correlations with PR, SD, and temperature (i.e., AT, ST, and EA), and a positive correlation with RH. The EOS had positive correlations with PR and temperature, and negative correlations with relative humidity.
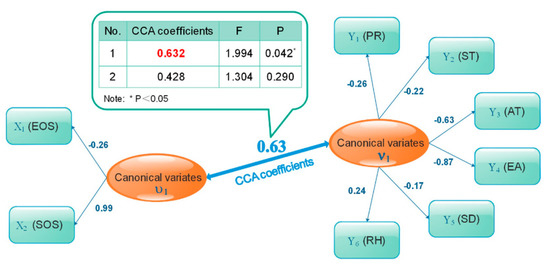
Figure 7.
Canonical correlation analysis patterns between phenology (SOS and EOS) and interannual climatic factors (SD, AT, ST, EA, RH, and PR).
The canonical redundancy analysis (Table 2) revealed that the first pair of canonical variables U1 could explain 53.14% of the intragroup variation and 8.97% of the other group variation. The typical variable V1, however, could explain 22.51% of the intragroup variation and 21.20% of the other group variation. Therefore, we could infer that the SOS and EOS of P. euphratica affect each other. The phenology (SOS, EOS) of P. euphratica is not only related to meteorological factors but also the physiological characteristics of P. euphratica. Therefore, it was found that the SOS of P. euphratica was strongly related to EOS with a high canonical correlation coefficient of 0.63, implying that a later SOS may generally be accompanied by a later EOS.
Table 2.
Canonical redundancy analyses between U1 and V1.
5. Discussion
5.1. Validation of P. euphratica Phenology
Remote sensing monitoring of vegetation phenology is based on vegetation growth characteristics (LAI, EVI, NDVI, and other features), and the vegetation phenology is simulated using a mathematical model. This method is different from the traditional ground observation phenology method [11], but previous studies showed that the vegetation phenology phase determined by remote sensing data was closely related to those obtained from the ground phenology monitoring points [59]. This study was based on one year’s worth of data from the ground phenology monitoring points of P. euphratica, and established a mathematical model of P. euphratica phenology (Table 1). The phenological features of P. euphratica, including the SOS, EOS, and LOS, were extracted using the dynamic threshold method. To verify the accuracy of the research results and the applicability of the method, based on the GLASS LAI and higher resolution of MODIS NDVI (MOD09Q1 V6) data in 2006, 2010, and 2014, 25 samples were randomly selected to compare and analyze the results of key phenological phase extraction of P. euphratica through the process flow chart steps in Figure 2. As shown in Figure 8, The results revealed that using the GLASS LAI and MODIS NDVI to extract the phenology of P. euphratica yielded higher correlations (SOS: R2 = 0.861, p < 0.001; EOS: R2 = 0.824, p < 0.001). Overall, this method could quickly identify and extract the key phenological period of P. euphratica in a large area, and this study’s findings are reliable and have scientific reference value for the study of vegetation phenology, especially in extremely arid areas.
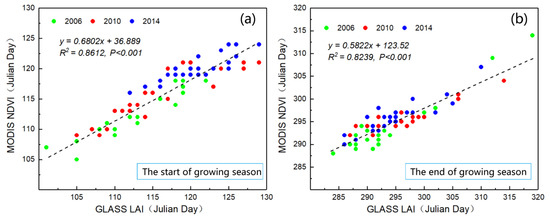
Figure 8.
Extraction of P. euphratica phenology based on MODIS NDVI and GLASS LAI data. (a) Start of the growing season (SOS), (b) end of the growing season (EOS).
In addition, phenological research of P. euphratica in the Tarim River basin has been rare, and other researchers [60,61,62,63,64,65,66] believe that air temperature is the main factor affecting plant phenology in arid and semi-arid areas, and the accumulated temperature should therefore be the condition used to judge P. euphratica phenology (Table 3). For example, Zhao et al. [61,66], Liu et al. [64,65], Wu et al. [63], and Zhang et al. [62] monitored the phenology of P. euphratica by taking the date the daily average temperature reached ≥5 °C as the SOS of P. euphratica, and the date the daily average temperature reached ≤5 °C as the EOS in Hexi Corridor. These results showed that the SOS of P. euphratica was early March, which was earlier than the findings of this study. The EOS of P. euphratica was late October, which was consistent with the findings of this study. However, our study showed that the temperature (i.e., AT, ST, and EA) and SD were strongly related to phenology (SOS, EOS) in the upper Tarim River basin. Moreover, vegetation phenology is affected by many factors such as geographical factors and climatic factors. Therefore, using a single accumulated temperature as the criterion for judging the phenology of P. euphratica cannot directly reflect this cycle, and this method is not rigorous.
Table 3.
Canonical redundancy comparative study on the phenology of P. euphratica [60,61,62,63,64,65,66].
5.2. Relationships between Phenology of P. euphratica and Climatic Factors
Previous researchers showed that the SOS of vegetation was advanced and EOS was delayed with climate warming [67,68,69,70]. In this study, however, the SOS and EOS of P. euphratica had an earlier trend, but the change was not significant in the upper Tarim River basin from 2000–2017. Wu et al. [71] reported that there was no significant advance trend in SOS from 1982–2016 in six temperate areas, and Zhang et al. [11] found that there was no advanced trend of spring phenology and the rate of earlier SOS was slow with variations in climate from 2006–2017. Zhang et al.’s [11] study results implied that the trend of EOS was not significant from 2006–2017, which was consistent with the findings of Li et al. [72] and Shen et al. [8]. It is reasonable that the SOS displays a later trend and EOS shows an earlier trend as the microclimate of this region is different. In addition, vegetation phenology is regional, which is closely related to species, geographical factors, and climatic factors. The response of vegetation phenology to climate change is not universally identical in different regions [73]. Therefore, it is necessary to explore the variations in P. euphratica phenology and its response to climate change in extremely arid areas.
In this study, we found that temperature (i.e., AT, ST, and EA) and SD controlled P. euphratica phenology, although the impacts of RH and PR on P. euphratica phenology cannot be overlooked in the upper Tarim River basin. Importantly, Figure 9 revealed that the SOS had negative correlations with PR, SD, and temperature (i.e., AT, ST, and EA), but a positive correlation with RH, while the relationships between the EOS and meteorological factors were the opposite. Air temperature is one of the main factors affecting the phenology of P. euphratica, but precipitation has the least effect on phenology compared to other climatic factors (i.e., AT, EA, ST, SD, RH) in the study area. The temperature increase may lead to earlier SOS, which is attributed to the fact that increasing temperature can promote the chemical reaction rate of vegetation to meet the thermal requirement for budburst and leaf unfolding, and it can accelerate the rate of earlier SOS [8,11,74,75].
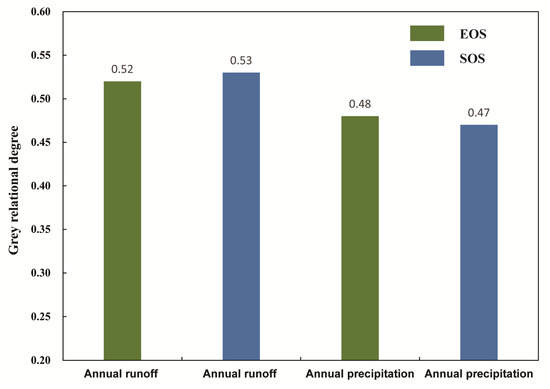
Figure 9.
Grey relational analysis between phenology (SOS and EOS) and water (annual runoff and annual precipitation).
Meanwhile, as reported in previous studies (e.g., Che et al. [73]; Dreesen et al. [76]; Anderegg et al. [77]) higher temperatures in spring and autumn will increase surface evapotranspiration and decrease water availability, preventing plant photosynthesis and growth, increasing plant death and chlorophyll degradation, and resulting in earlier EOS. This study found that a temperature increase may lead to earlier SOS and later EOS, which was similar to the findings of Wang et al. [67] and Tao et al. [78], who reported that a temperature decrease may lead to later SOS and earlier EOS among different vegetation types. In addition, the results of this study revealed that the SOS of P. euphratica was strongly related to EOS with a high canonical correlation coefficient of 0.63, indicating that a later SOS may generally be accompanied by a later EOS. This result was similar to the findings of Wu et al. [58], who reported that SOS was positively correlated with EOS from 1997–2014.
5.3. Which has a Stronger Influence on Phenology, Runoff, or Precipitation?
Water is a key climatic factor restricting vegetation growth in arid areas [79,80]. The phenological characteristics of P. euphratica in the Tarim River basin are affected by rainfall and runoff. Rainfall directly increases soil moisture and provides direct absorption by the roots of P. euphratica [81,82]. On the other hand, runoff compensates the groundwater so the groundwater depth becomes shallow and is absorbed by the roots of P. euphratica [34,43]. Based on the annual runoff and precipitation data, this study used the grey correlation method to explore the grey correlation grade between P. euphratica phenology and annual rainfall and runoff (Figure 9). The results demonstrated that the grey correlation grade between runoff and phenology is higher than that between rainfall and phenology. Precipitation is scarce in arid areas, and P. euphratica growth is not sensitive to precipitation events, which is consistent with previous research results [83,84]. However, runoff affects the growth of P. euphratica by replenishing the groundwater table, which is then absorbed by the roots of P. euphratica, hence there is a certain lag effect [85]. Therefore, the grey correlation grade of rainfall and runoff for the phenology of P. euphratica is low, but runoff is still the dominant factor, which is easy to regulate and control. The regulation of runoff and the stable control or reduction in shallow groundwater depth may weaken the impact of global climate change on the phenology of P. euphratica.
5.4. Limitations
Through field investigation, the concentrated distribution of P. euphratica has been found to be decreased [86,87]. P. euphratica has gradually declined due to climate changes and disturbance by human activities, and its vegetation coverage is low [34,85]. The retrieval accuracy of the spatio-temporal patterns of P. euphratica phenology using 1 km × 1 km GLASS LAI remote sensing data is not high. Moreover, considering that there is a wide range of mixed pixels in satellite remote sensing data, the spatial resolution of remote sensing data will affect the accuracy of P. euphratica phenological feature inversion. Therefore, to solve the above two problems, this study utilized a grid spatial sampling strategy to select typical sample areas in order to obtain P. euphratica phenological information, thereby reducing the impact of the above two problems to explore the changing trend of P. euphratica phenology in arid areas and its response to climate change. This paper has provided an approach in which the satellite product data are used to reveal the interannual variations of the phenology of the vegetation, plant community, and a specific tree species, of which the spatial aggregation and coverage are low.
Yang et al. [88] found that the preseasons of EOS were defined as 1–6 months before early November, and the preseason temperature is the major control factor of the dormancy onset date. Fu et al. [89] defined May to October as the EOS preseason, and then revealed the influences of climate change and spring phenology on autumn phenology from 1982–2015. Ren et al. [90] found that the phenology was most correlated with climatic factors in the period of about three months to phenology (SOS, EOS). However, the key periods that affect the phenology may vary among different areas [91,92,93]. In this study, the climatic factors could be divided into five temporal periods, which have a certain subjectivity. We should use a suitable method in the future, such as linear correlation analysis, partial correlation analysis, and Pearson correlation analysis, to determine the optimal periods, and make the climatic factors in the period have a strong correlation with phenology.
Since the study area mainly consists of modern alluvial plain and delta plain, the topography is characterized by monotonous flatness and a slight slope of the entire plain, and the range of elevation is small, the impact of topographical factors on P. euphratica phenology was ignored. Due to the limitation of vegetation coverage and aggregation degree of P. euphratica in arid areas, this study needs a longer time series and higher spatial resolution remote sensing data to reveal the phenological characteristics of P. euphratica. Meanwhile, this study established a model based on a single year of ground monitoring data, and the accuracy of the simulation was insufficient, indicating that further improvement is needed in simulation and validation. Thus, these questions await further investigation.
6. Conclusions
Vegetation phenology is very sensitive to climate change in extremely arid regions, and is thus the best indicator of climate change as well as an essential indicator of ecosystem function. Using the Global Land Surface Satellite leaf area index time series data from 2000 to 2017, the phenological information (i.e., SOS, LOS, and EOS) was obtained with the dynamic threshold method from the reconstructed growth time series curve by using the Savitzky–Golay filtering method. The interannual variation trends of P. euphratica phenology were not significant from 2000 to 2017 because there was no significant trend in annual climatic factors. P. euphratica phenology was remarkably changed with respect to its high spatial heterogeneity. The spring climatic factors had a great influence on the SOS of P. euphratica, while the EOS was mainly affected by the EOS preseason (June–November) meteorological factors. The temperature and SD controlled P. euphratica phenology, although the impacts of RH and PR on P. euphratica phenology cannot be overlooked in the upper Tarim River basin, implying that it is not easy to characterize phenological change with simple climate data. This difficulty is due to the interaction between the SOS in spring and the EOS in autumn, as well as the comprehensive influence of multiple climatic factors on vegetation phenology change, which has not been adequately considered in current vegetation phenology research and ecosystem models. This study also revealed that global warming may lead to SOS advancement and EOS delay, and the increase in SD may lead to earlier SOS and later EOS. Due to the scarcity of rainfall in extremely arid regions, vegetation growth primarily depends on groundwater recharge, so the phenology of P. euphratica has a higher correlation with runoff than rainfall in the upper reaches of the Tarim River. The results provide insights into quantitative assessment of spatio-temporal variations in P. euphratica phenology under climate change and provide a scientific reference for future management of the desert ecosystem.
Author Contributions
Conceptualization, H.L. and L.B.; methodology, H.L.; software, J.F.; validation, J.F. and J.Z.; formal analysis, H.L. and J.F.; investigation, H.L. and L.B.; writing—original draft preparation, H.L.; writing—review and editing, L.B. and J.F.; funding acquisition, L.B. and J.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by China’s Xinjiang Production and Construction Corps’s (XPCC) Key Science and Technology Project (No. 2019AB002), the Agricultural Science and Technology Innovation Program (ASTIP) of the Chinese Academy of Agricultural Sciences (CAAS-ASTIP-2016-AII), and the Strategic Priority Research Program of the Chinese Academy of Sciences (No. XDA19030302).
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We would like to thank the National Earth System Science Data Center, National Science & Technology Infrastructure of China (http://www.geodata.cn accessed on 14 July 2021) for their data support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, Q.; Kong, D.; Shi, P.; Singh, V.P.; Sun, P. Vegetation phenology on the Qinghai-Tibetan Plateau and its response to climate change (1982–2013). Agric. For. Meteorol. 2018, 248, 408–417. [Google Scholar] [CrossRef]
- Nilsson, A.L.K.; Slagsvold, T.; Rostad, O.W.; Knudsen, E.; Jerstad, K.; Cadahia, L.; Reitan, T.; Helberg, M.; Walseng, B.; Stenseth, N.C. Territory location and quality, together with climate, affect the timing of breeding in the white-throated dipper. Sci. Rep. 2019, 9, 7671. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Jin, N.; Yu, Q. Impacts of climate change and crop management practices on soybean phenology changes in China. Sci. Total Environ. 2020, 707, 135638. [Google Scholar] [CrossRef]
- Gonsamo, A.; Chen, J.M.; Price, D.T.; Kurz, W.A.; Liu, J.; Boisvenue, C.; Hember, R.A.; Wu, C.; Chang, K.-H. Improved assessment of gross and net primary productivity of Canada’s landmass. J. Geophys. Res. Biogeosci. 2013, 118, 1546–1560. [Google Scholar] [CrossRef]
- Meng, F.; Niu, H.; Suonan, J.; Zhang, Z.; Wang, Q.; Li, B.; Lv, W.; Wang, S.; Duan, J.; Liu, P. Divergent responses of community reproductive and vegetative phenology to warming and cooling: Asymmetry vs symmetry. Front. Plant Sci. 2019, 10, 1310. [Google Scholar] [CrossRef] [PubMed]
- Pabon-Moreno, D.E.; Musavi, T.; Migliavacca, M.; Reichstein, M.; Mahecha, M.D.J.B. Ecosystem physio-phenology revealed using circular statistics. Biogeoences 2020, 17, 3991–4006. [Google Scholar] [CrossRef]
- Schwieder, M.; Leitao, P.J.; Bustamante, M.M.D.; Ferreira, L.G.; Rabe, A.; Hostert, P. Mapping Brazilian savanna vegetation gradients with Landsat time series. Int. J. Appl. Earth Obs. Geoinf. 2016, 52, 361–370. [Google Scholar] [CrossRef]
- Shen, M.; Jiang, N.; Peng, D.; Rao, Y.; Huang, Y.; Fu, Y.H.; Yang, W.; Zhu, X.; Cao, R.; Chen, X.; et al. Can changes in autumn phenology facilitate earlier green-up date of northern vegetation? Agric. For. Meteorol. 2020, 291, 108077. [Google Scholar] [CrossRef]
- Misra, G.; Asam, S.; Menzel, A. Ground and satellite phenology in alpine forests are becoming more heterogeneous across higher elevations with warming. Agric. For. Meteorol. 2021, 303, 108383. [Google Scholar] [CrossRef]
- Burgess, M.D.; Smith, K.W.; Evans, K.L.; Leech, D.; Pearce-Higgins, J.W.; Branston, C.J.; Briggs, K.; Clark, J.R.; Feu, C.R.d.; Lewthwaite, K.; et al. Tritrophic phenological match-mismatch in space and time. Nat. Ecol. Evol. 2018, 2, 945–970. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Tong, X.; Zhang, J.; Meng, P.; Li, J.; Liu, P. Dynamics of phenology and its response to climatic variables in a warm-temperate mixed plantation. For. Ecol. Manag. 2021, 483, 118785. [Google Scholar] [CrossRef]
- Araghi, A.; Martinez, C.J.; Adamowski, J.; Olesen, J.E. Associations between large-scale climate oscillations and land surface phenology in Iran. Agric. For. Meteorol. 2019, 278, 107682. [Google Scholar] [CrossRef]
- Bornez, K.; Descals, A.; Verger, A.; Penuelas, J. Land surface phenology from VEGETATION and PROBA-V data. Assessment over deciduous forests. Int. J. Appl. Earth Obs. Geoinf. 2020, 84, 11. [Google Scholar] [CrossRef]
- Geng, X.; Zhou, X.; Yin, G.; Hao, F.; Zhang, X.; Hao, Z.; Singh, V.P.; Fu, Y.H. Extended growing season reduced river runoff in Luanhe River basin. J. Hydrol. 2020, 582, 124538. [Google Scholar] [CrossRef]
- Norman, S.P.; Hargrove, W.W.; Christie, W.M. Spring and Autumn Phenological Variability across Environmental Gradients of Great Smoky Mountains National Park, USA. Remote Sens. 2017, 9, 407. [Google Scholar] [CrossRef] [Green Version]
- Tong, X.Y.; Tian, F.; Brandt, M.; Liu, Y.; Zhang, W.M.; Fensholt, R. Trends of land surface phenology derived from passive microwave and optical remote sensing systems and associated drivers across the dry tropics 1992–2012. Remote Sens. Environ. 2019, 232, 12. [Google Scholar] [CrossRef]
- Xu, X.J.; Zhou, G.M.; Du, H.Q.; Mao, F.J.; Xu, L.; Li, X.J.; Liu, L.J. Combined MODIS land surface temperature and greenness data for modeling vegetation phenology, physiology, and gross primary production in terrestrial ecosystems. Sci. Total Environ. 2020, 726, 11. [Google Scholar] [CrossRef]
- Delbart, N.; Beaubien, E.; Kergoat, L.; le Toan, T. Comparing land surface phenology with leafing and flowering observations from the PlantWatch citizen network. Remote Sens. Environ. 2015, 160, 273–280. [Google Scholar] [CrossRef]
- Lim, C.H.; Jung, S.H.; Kim, A.R.; Kim, N.S.; Lee, C.S. Monitoring for Changes in Spring Phenology at Both Temporal and Spatial Scales Based on MODIS LST Data in South Korea. Remote Sens. 2020, 12, 3282. [Google Scholar] [CrossRef]
- Suepa, T.; Qi, J.; Lawawirojwong, S.; Messina, J.P. Understanding spatio-temporal variation of vegetation phenology and rainfall seasonality in the monsoon Southeast Asia. Environ. Res. 2016, 147, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.X.; Zhao, L.; Lin, A.W.; Wang, L.C.; Li, Q.J.; She, D.X.; Qu, S. Impacts of preseason drought on vegetation spring phenology across the Northeast China Transect. Sci. Total Environ. 2020, 738, 10. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Liu, Q.; Du, W.; Zhou, G.; Qin, L.; Sun, Z. Modelling leaf phenology of some trees with accumulated temperature in a temperate forest in northeast China. For. Ecol. Manag. 2021, 489, 119085. [Google Scholar] [CrossRef]
- Zou, F.; Li, H.; Hu, Q. Responses of vegetation greening and land surface temperature variations to global warming on the Qinghai-Tibetan Plateau, 2001–2016. Ecol. Indicators 2020, 119, 106867. [Google Scholar] [CrossRef]
- Ma, J.; Xiao, X.; Li, R.; Zhao, B.; Myint, S.W. Enhanced spring phenological temperature sensitivity explains the extension of carbon uptake period in temperate forest protected areas. For. Ecol. Manag. 2020, 455, 117679. [Google Scholar] [CrossRef]
- Meier, G.A.; Brown, J.F.; Evelsizer, R.J.; Vogelmann, J.E. Phenology and climate relationships in aspen (Populus tremuloides Michx.) forest and woodland communities of southwestern Colorado. Ecol. Indicators 2015, 48, 189–197. [Google Scholar] [CrossRef]
- Li, X.; Zhang, L.; Luo, T. Rainy season onset mainly drives the spatiotemporal variability of spring vegetation green-up across alpine dry ecosystems on the Tibetan Plateau. Sci. Rep. 2020, 10, 18797. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zheng, X.; Ma, L.; Wang, H.; Huang, Q.; Leng, G.; Meng, E.; Guo, Y. Quantitative contribution of climate change and human activities to vegetation cover variations based on GA-SVM model. J. Hydrol. 2020, 584, 124687. [Google Scholar] [CrossRef]
- An, S.; Zhang, X.; Chen, X.; Yan, D.; Henebry, G. An Exploration of Terrain Effects on Land Surface Phenology across the Qinghai–Tibet Plateau Using Landsat ETM+ and OLI Data. Remote Sens. 2018, 10, 1069. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Wang, Y.; Zhang, Z.; Magliulo, V.; Jiang, H.; Cheng, M. Phenology Plays an Important Role in the Regulation of Terrestrial Ecosystem Water-Use Efficiency in the Northern Hemisphere. Remote Sens. 2017, 9, 664. [Google Scholar] [CrossRef] [Green Version]
- Yuan, G.; Zhang, P.; Shao, M.-a.; Luo, Y.; Zhu, X. Energy and water exchanges over a riparian Tamarix spp. stand in the lower Tarim River basin under a hyper-arid climate. Agric. For. Meteorol. 2014, 194, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Schimel, D.S. Drylands in the Earth System. Science 2010, 327, 418–419. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Feng, Q.; Si, J.; Xi, H.; Su, Y.; Mitchell, P.J.; Pinkard, E.A. Flooding constrains tree water use of a riparian forest in the lower Heihe River Basin, Northwest China. Sci. Total Environ. 2021, 760, 144069. [Google Scholar] [CrossRef] [PubMed]
- Noy-Meir, I. Desert ecosystems: Environment and producers. Annu. Rev. Ecol. Syst. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Zhou, H.H.; Chen, Y.N.; Zhu, C.G.; Li, Z.; Fang, G.H.; Li, Y.P.; Fu, A.H. Climate change may accelerate the decline of desert riparian forest in the lower Tarim River, Northwestern China: Evidence from tree-rings of Populus euphratica. Ecol. Indicators 2020, 111, 105997. [Google Scholar] [CrossRef]
- Li, H.; Bai, L.; Feng, J.; Gao, H.; Ran, Q.; Yu, T.; Gao, W. Analysis of spatio-temporal characteristics of Populus euphratica forests in the Yarkand River Basin, Xinjiang. Acta Ecol. Sin. 2019, 39, 5080–5094. (In Chinese) [Google Scholar] [CrossRef]
- Gou, S.; Miller, G. A groundwater-soil-plant-atmosphere continuum approach for modelling water stress, uptake, and hydraulic redistribution in phreatophytic vegetation. Ecohydrology 2014, 7, 1029–1041. [Google Scholar] [CrossRef]
- Chen, Y.; Li, Z.; Fan, Y.; Wang, H.; Deng, H. Progress and prospects of climate change impacts on hydrology in the arid region of northwest China. Environ. Res. 2015, 139, 11–19. [Google Scholar] [CrossRef]
- Wu, K.; Xu, W.; Yang, W. Effects of precipitation changes on soil bacterial community composition and diversity in the Junggar desert of Xinjiang, China. PeerJ 2020, 8, e8433. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Li, W.; Ji, M.; Lu, F.; Dong, S. A comprehensive approach to characterization of the nonlinearity of runoff in the headwaters of the Tarim River, western China. Hydrol. Process. 2010, 24, 136–146. [Google Scholar] [CrossRef]
- Keram, A.; Halik, Ü.; Keyimu, M.; Aishan, T.; Mamat, Z.; Rouzi, A. Gap dynamics of natural Populus euphratica floodplain forests affected by hydrological alteration along the Tarim River: Implications for restoration of the riparian forests. For. Ecol. Manag. 2019, 438, 103–113. [Google Scholar] [CrossRef]
- Lang, P.; Ahlborn, J.; Schäfer, P.; Wommelsdorf, T.; Jeschke, M.; Zhang, X.; Thomas, F.M. Growth and water use of Populus euphratica trees and stands with different water supply along the Tarim River, NW China. For. Ecol. Manag. 2016, 380, 139–148. [Google Scholar] [CrossRef]
- Eusemann, P.; Petzold, A.; Thevs, N.; Schnittler, M. Growth patterns and genetic structure of Populus euphratica Oliv. (Salicaceae) forests in NW China – Implications for conservation and management. For. Ecol. Manag. 2013, 297, 27–36. [Google Scholar] [CrossRef]
- Zhou, H.H.; Chen, Y.N.; Li, W.H.; Chen, Y.P. Photosynthesis of Populus euphratica in relation to groundwater depths and high temperature in arid environment, northwest China. Photosynthetica 2010, 48, 257–268. [Google Scholar] [CrossRef]
- Hu, Y.; Li, H.; Wu, D.; Chen, W.; Zhao, X.; Hou, M.; Li, A.; Zhu, Y. LAI-indicated vegetation dynamic in ecologically fragile region: A case study in the Three-North Shelter Forest program region of China. Ecol. Indicators 2021, 120, 106932. [Google Scholar] [CrossRef]
- Li, J.; Xiao, Z. Evaluation of the version 5.0 global land surface satellite (GLASS) leaf area index product derived from MODIS data. Int. J. Remote Sens. 2020, 41, 9140–9160. [Google Scholar] [CrossRef]
- Xiang, Y.; Xiao, Z.; Ling, S.; Wang, J.; Song, J. Validation of Global LAnd Surface Satellite (GLASS) leaf area index product. J. Remote Sens. 2014, 18, 573–584. [Google Scholar] [CrossRef]
- Zhao, Y.; Bai, L.; Feng, J.; Lin, X.; Wang, L.; Xu, L.; Ran, Q.; Wang, K. Spatial and Temporal Distribution of Multiple Cropping Indices in the North China Plain Using a Long Remote Sensing Data Time Series. Sensors 2016, 16, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Li, J.; Liu, Q.H.; Zhong, B.; Wu, S.L.; Xia, C.F. Analysis of Differences in Phenology Extracted from the Enhanced Vegetation Index and the Leaf Area Index. Sensors 2017, 17, 1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.; Jönsson, P.; Jin, H.; Eklundh, L. Performance of Smoothing Methods for Reconstructing NDVI Time-Series and Estimating Vegetation Phenology from MODIS Data. Remote Sens. 2017, 9, 1271. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Yang, X.; Niu, J.; Jin, Y.; Xu, B.; Shen, G.; Zhang, W.; Zhao, F.; Zhang, Y. Remote sensing monitoring of green-up dates in the Xilingol grasslands of northern China and their correlations with meteorological factors. Int. J. Remote Sens. 2019, 40, 2190–2211. [Google Scholar] [CrossRef]
- Hu, L.; Fan, W.; Ren, H.; Liu, S.; Cui, Y.; Zhao, P. Spatiotemporal Dynamics in Vegetation GPP over the Great Khingan Mountains Using GLASS Products from 1982 to 2015. Remote Sens. 2018, 10, 488. [Google Scholar] [CrossRef] [Green Version]
- Jönsson, P.; Eklundh, L. Seasonality extraction by function fitting to time-series of satellite sensor data. IEEE Trans. Geosci. Comput. Remote Sens. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Li, P.; He, Z.; He, D.; Xue, D.; Wang, Y.; Cao, S. Fractional vegetation coverage response to climatic factors based on grey relational analysis during the 2000–2017 growing season in Sichuan Province, China. Int. J. Remote Sens. 2019, 41, 1170–1190. [Google Scholar] [CrossRef]
- He, D.; Yi, G.; Zhang, T.; Miao, J.; Li, J.; Bie, X. Temporal and Spatial Characteristics of EVI and Its Response to Climatic Factors in Recent 16 years Based on Grey Relational Analysis in Inner Mongolia Autonomous Region, China. Remote Sens. 2018, 10, 961. [Google Scholar] [CrossRef] [Green Version]
- Gao, B.; Li, J.; Wang, X.S. Impact of frozen soil changes on vegetation phenology in the source region of the Yellow River from 2003 to 2015. Theor. Appl. Climatol. 2020, 141, 1219–1234. [Google Scholar] [CrossRef]
- Hotelling, H. Relations Between Two Sets of Variates. Biometrika 1935, 28, 321–377. [Google Scholar] [CrossRef]
- Ivanova, Y.; Kovalev, A.; Yakubailik, O.; Soukhovolsky, V. The Use of Satellite Information (MODIS/Aqua) for Phenological and Classification Analysis of Plant Communities. Forests 2019, 10, 561. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Hou, X.; Peng, D.; Gonsamo, A.; Xu, S. Land surface phenology of China’s temperate ecosystems over 1999–2013: Spatial-temporal patterns, interaction effects, covariation with climate and implications for productivity. Agric. For. Meteorol. 2016, 216, 177–187. [Google Scholar] [CrossRef]
- Peng, D.; Zhang, X.; Wu, C.; Huang, W.; Gonsamo, A.; Huete, A.R.; Didan, K.; Tan, B.; Liu, X.; Zhang, B. Intercomparison and evaluation of spring phenology products using National Phenology Network and AmeriFlux observations in the contiguous United States. Agric. For. Meteorol. 2017, 242, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Abdurahman, M.; Kurban, A.; Ablat, A.; Adilla, R.; Duan, H.; Ablekim, A.; Halik, U. Study on Phenological Characters of Populus euphratica Oliv. in the Lower Reaches of the Tarim River. Arid Zone Res. 2008, 25, 525–530. (In Chinese) [Google Scholar] [CrossRef]
- Zhao, M.; Liu, P.; Zhu, X.; Zhang, K. Respond of Populus euphratica Oliv. Phenology to climate Warming in the Oasis Lower Reaches of Heihe Rvier from 1960 to 2010. Acta Bot. Boreali-Occident. Sin. 2012, 32, 2108–2115. (In Chinese) [Google Scholar]
- Zhang, W.; Liu, P.; Feng, Q.; Wang, T.; Wang, T. The spatiotemporal responses of Populus euphratica to global warming in Chinese oases between 1960 and 2015. J. Geogr. Sci. 2018, 28, 579–594. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Zhang, S.; Ji, Y. Response of Populus euphratica Oliv to Climate Change in Minqin Oasis from 1955 to 2009. Res. Soil Water Conserv. 2015, 22, 123–127. (In Chinese) [Google Scholar] [CrossRef]
- Liu, P.; Zhu, X.; Zhao, M.; Yao, Y.; Chen, L. Response of Annual Growing Season of Populus euphraticato Climate Change in the Jiuquan Oasis during the Period from 1955 to 2010. Arid Zone Res. 2013, 30, 101–108. (In Chinese) [Google Scholar] [CrossRef]
- Liu, P.; Yang, Q. Response of the Annual Growing Season of Populus Euphratica to Climate Change in Dunhuang Oasis from 1955 to 2010. Resour. Sci. 2012, 34, 566–571. (In Chinese) [Google Scholar]
- Zhao, M.; Liu, P.; Zhu, X.; Zhang, K.; Hou, C. Response of Populus Euphratica’s Annual Growth Period to Climate Change in Ejina Banner’s Oasis over the Last 51 Years. Bull. Soil Water Conserv. 2012, 32, 205–209. (In Chinese) [Google Scholar] [CrossRef]
- Wang, Y.; Luo, Y.; Shafeeque, M. Interpretation of vegetation phenology changes using daytime and night-time temperatures across the Yellow River Basin, China. Sci. Total Environ. 2019, 25, 693. [Google Scholar] [CrossRef]
- Liang, S.; Shi, P.; Li, H. Urban spring phenology in the middle temperate zone of China: Dynamics and influence factors. Int. J. Biometeorol. 2016, 60, 531–544. [Google Scholar] [CrossRef]
- Karami, M.; Hansen, B.U.; Westergaard-Nielsen, A.; Abermann, J.; Lund, M.; Schmidt, N.M.; Elberling, B. Vegetation phenology gradients along the west and east coasts of Greenland from 2001 to 2015. Ambio 2017, 46, S94–S105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, C.; Tang, X.; Gu, Q.; Wang, T.; Wei, J.; Song, L.; Ma, M. Climatic anomaly and its impact on vegetation phenology, carbon sequestration and water-use efficiency at a humid temperate forest. J. Hydrol. 2018, 565, 150–159. [Google Scholar] [CrossRef]
- Wu, X.C.; Liu, H.Y. Consistent shifts in spring vegetation green-up date across temperate biomes in China, 1982–2006. Glob. Chang. Biol. 2013, 19, 870–880. [Google Scholar] [CrossRef]
- Li, P.; Peng, C.; Wang, M.; Luo, Y.; Li, M.; Zhang, K.; Zhang, D.; Zhu, Q. Dynamics of vegetation autumn phenology and its response to multiple environmental factors from 1982 to 2012 on Qinghai-Tibetan Plateau in China. Sci. Total Environ. 2018, 637–638, 855. [Google Scholar] [CrossRef]
- Che, M.; Chen, B.; Innes, J.L.; Wang, G.; Dou, X.; Zhou, T.; Zhang, H.; Yan, J.; Xu, G.; Zhao, H. Spatial and temporal variations in the end date of the vegetation growing season throughout the Qinghai–Tibetan Plateau from 1982 to 2011. Agric. For. Meteorol. 2014, 189–190, 81–90. [Google Scholar] [CrossRef]
- Chen, L.; Huang, J.-G.; Ma, Q.; Hanninen, H.; Tremblay, F.; Bergeron, Y. Long-term changes in the impacts of global warming on leaf phenology of four temperate tree species. Glob. Chang. Biol. 2019, 25, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Asse, D.; Chuine, I.; Vitasse, Y.; Yoccoz, N.G.; Delpierre, N.; Badeau, V.; Delestrade, A.; Randin, C.F. Warmer winters reduce the advance of tree spring phenology induced by warmer springs in the Alps. Agric. For. Meteorol. 2018, 252, 220–230. [Google Scholar] [CrossRef]
- Dreesen, F.E.; Boeck, H.D.; Janssens, I.A.; Nijs, I. Do successive climate extremes weaken the resistance of plant communities? An experimental study using plant assemblages. Biogeosciences 2014, 10, 9149–9177. [Google Scholar] [CrossRef] [Green Version]
- Anderegg, W.R.L.; Plavcova, L.; Anderegg, L.D.L.; Hacke, U.G.; Berry, J.A.; Field, C.B. Drought’s legacy: Multiyear hydraulic deterioration underlies widespread aspen forest die-off and portends increased future risk. Glob. Chang. Biol. 2013, 19, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.X.; Wang, H.J.; Liu, Y.C.; Xu, Y.J.; Dai, J.H. Phenological response of different vegetation types to temperature and precipitation variations in northern China during 1982–2012. Int. J. Remote Sens. 2017, 38, 3236–3252. [Google Scholar] [CrossRef]
- Gao, X.; Wang, A.; Zhao, Y.; Zhao, X.; Sun, M.; Du, J.; Gang, C. Study on Water Suitability of Apple Plantations in the Loess Plateau under Climate Change. Int. J. Environ. Res. Public Health 2018, 15, 2504. [Google Scholar] [CrossRef] [Green Version]
- Ying, W.; Chunxia, W.; Jukui, Z.; Chunqing, W. The reproductive strategy in a Chloris virgata population in response to precipitation regimes. R. Soc. Open Sci. 2018, 5, 180607. [Google Scholar] [CrossRef]
- Song, X.L.; Wang, Y.H.; Lv, X.M. Responses of plant biomass, photosynthesis and lipid peroxidation to warming and precipitation change in two dominant species (Stipa grandis and Leymus chinensis) from North China Grasslands. Ecol. Evol. 2016, 6, 1871–1882. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.Y.; Zuo, X.A.; Li, Y.L.; Zhang, T.H.; Zhang, R.; Chen, J.L.; Lv, P.; Zhao, X.Y. Community carbon and water exchange responses to warming and precipitation enhancement in sandy grassland along a restoration gradient. Ecol. Evol. 2019, 9, 10938–10949. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.-Q.; Li, Y.; Xu, H. Seasonal variation in plant hydraulic traits of two co-occurring desert shrubs, Tamarix ramosissima and Haloxylon ammodendron, with different rooting patterns. Ecol. Res. 2011, 26, 1071–1080. [Google Scholar] [CrossRef]
- Xu, H.; Li, Y. Water-use strategy of three central Asian desert shrubs and their responses to rain pulse events. Plant Soil. 2006, 285, 5–17. [Google Scholar] [CrossRef]
- Liao, S.M.; Xue, L.Q.; Dong, Z.C.; Zhu, B.L.; Zhang, K.; Wei, Q.; Fu, F.B.; Wei, G.H. Cumulative ecohydrological response to hydrological processes in arid basins. Ecol. Indicators 2020, 111, 106005. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, Y.; Zhang, Z.; Zhang, H.; Guo, X.; Yu, S.; Du, W.; Huang, F. The Variations of Land Surface Phenology in Northeast China and Its Responses to Climate Change from 1982 to 2013. Remote Sens. 2016, 8, 400. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, D.; Chen, J.; Wang, C.; Shen, M. The mixed pixel effect in land surface phenology: A simulation study. Remote Sens. Environ. 2018, 211, 338–344. [Google Scholar] [CrossRef]
- Yang, Y.T.; Guan, H.D.; Shen, M.G.; Liang, W.; Jiang, L. Changes in autumn vegetation dormancy onset date and the climate controls across temperate ecosystems in China from 1982 to 2010. Glob. Chang. Biol. 2015, 21, 652–665. [Google Scholar] [CrossRef]
- Fu, Y.Y.; He, H.S.; Zhao, J.J.; Larsen, D.R.; Zhang, H.Y.; Sunde, M.G.; Duan, S.W. Climate and Spring Phenology Effects on Autumn Phenology in the Greater Khingan Mountains, Northeastern China. Remote Sens. 2018, 10, 499. [Google Scholar] [CrossRef] [Green Version]
- Ren, P.X.; Liu, Z.L.; Zhou, X.L.; Peng, C.H.; Xiao, J.F.; Wang, S.H.; Li, X.; Li, P. Strong controls of daily minimum temperature on the autumn photosynthetic phenology of subtropical vegetation in China. For. Ecosyst. 2021, 8, 1–21. [Google Scholar] [CrossRef]
- Yuan, M.X.; Wang, L.C.; Lin, A.W.; Liu, Z.J.; Li, Q.J.; Qu, S. Vegetation green up under the influence of daily minimum temperature and urbanization in the Yellow River Basin, China. Ecol. Indicators 2020, 108, 105760. [Google Scholar] [CrossRef]
- Jeong, S.; Ho, C.; Gim, H.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Chang. Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Du, J.; He, Z.; Piatek, K.B.; Chen, L.; Lin, P.; Zhu, X. Interacting effects of temperature and precipitation on climatic sensitivity of spring vegetation green-up in arid mountains of China. Agric. For. Meteorol. 2019, 269, 71–77. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).