Developing Growth Models of Stand Volume for Subtropical Forests in Karst Areas: A Case Study in the Guizhou Plateau




Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection and Treatment
2.2.1. Forest Inventory Spatial Dataset
2.2.2. Climate Spatial Dataset
2.3. Methods
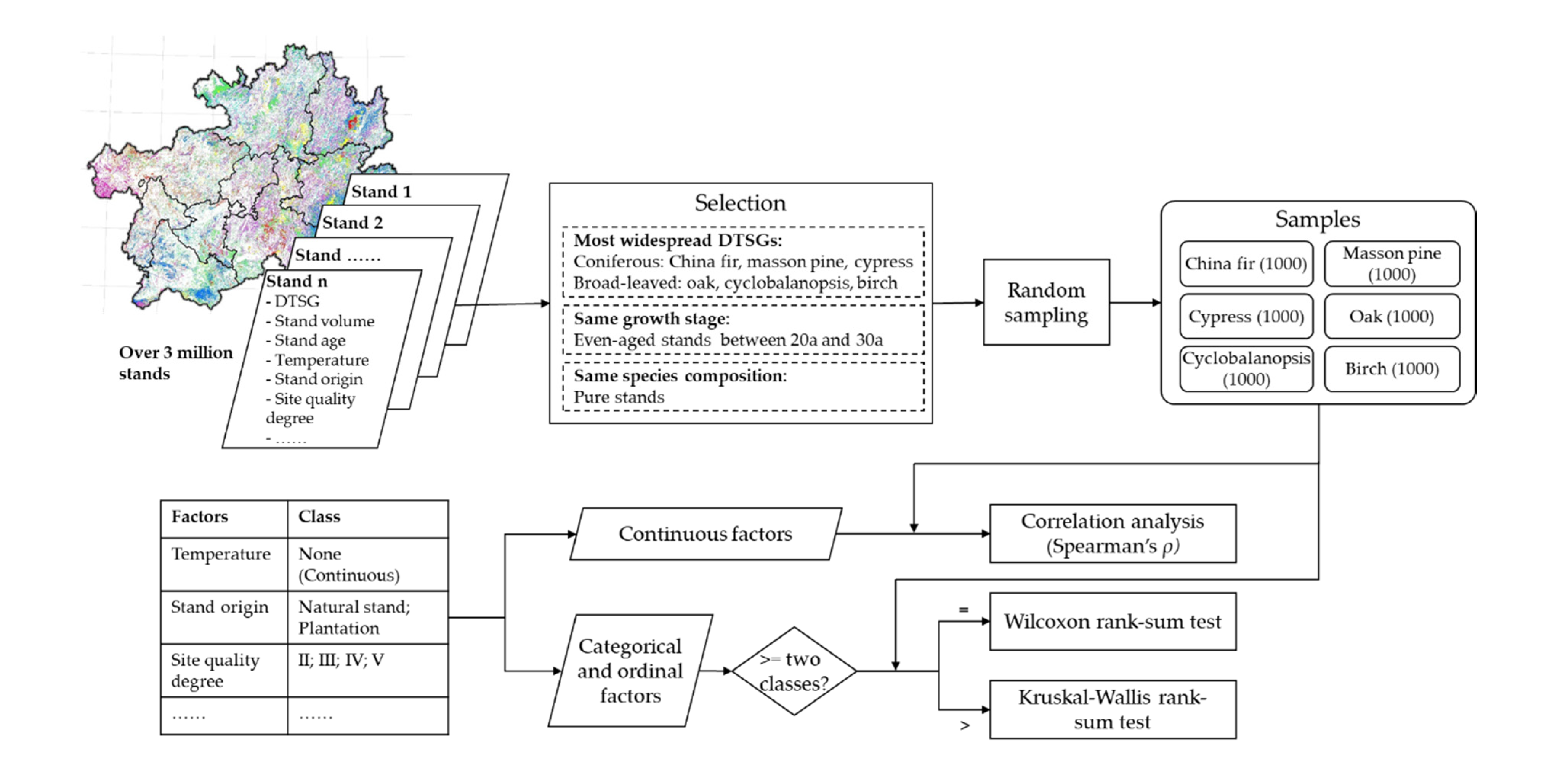
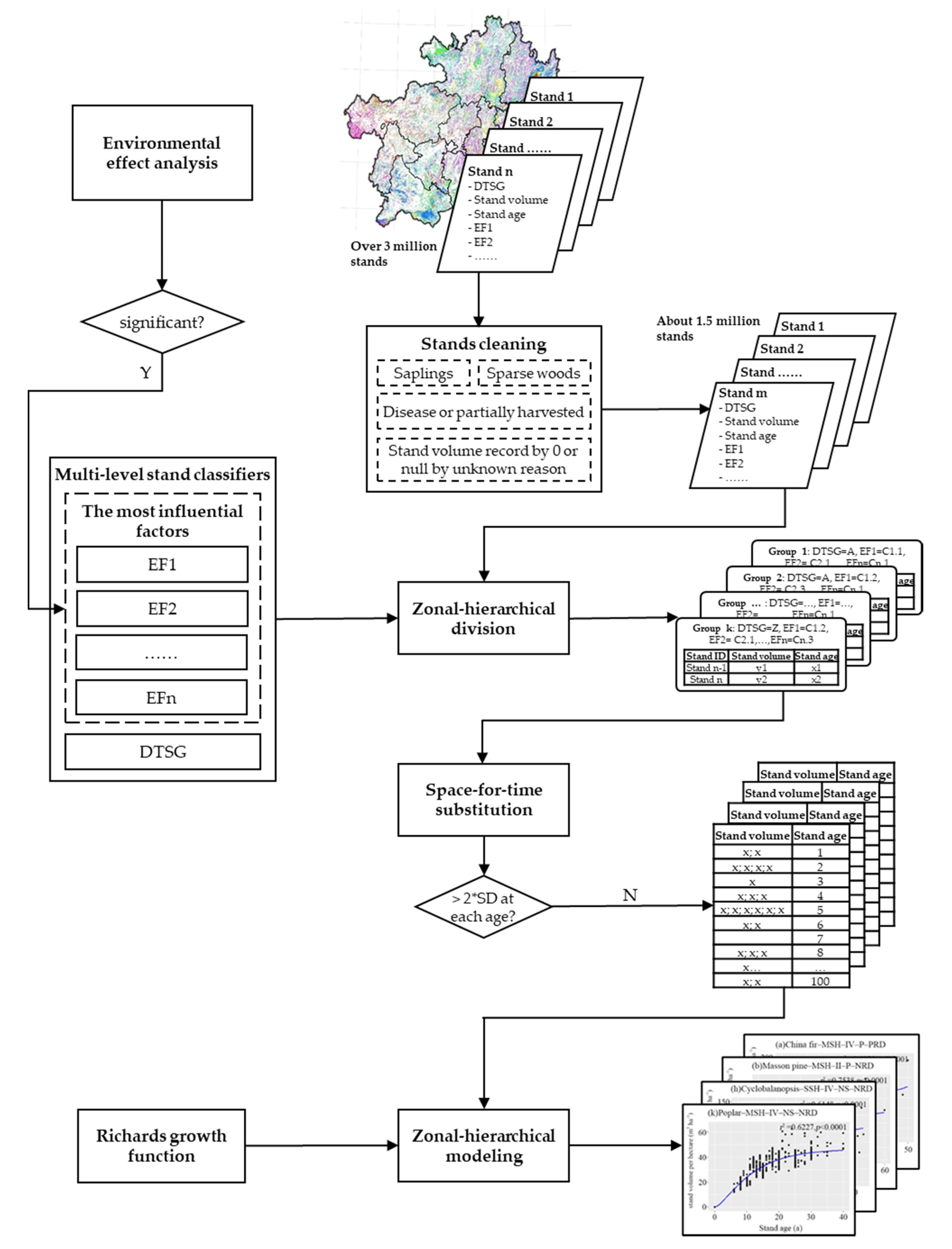
2.3.1. Environmental Effect Analysis
Environmental Factor Selection
Stand Selection and Sampling for Environmental Effect Analysis
Statistical Analysis for Revealing the Effect of Environment Factors on Stand Volume
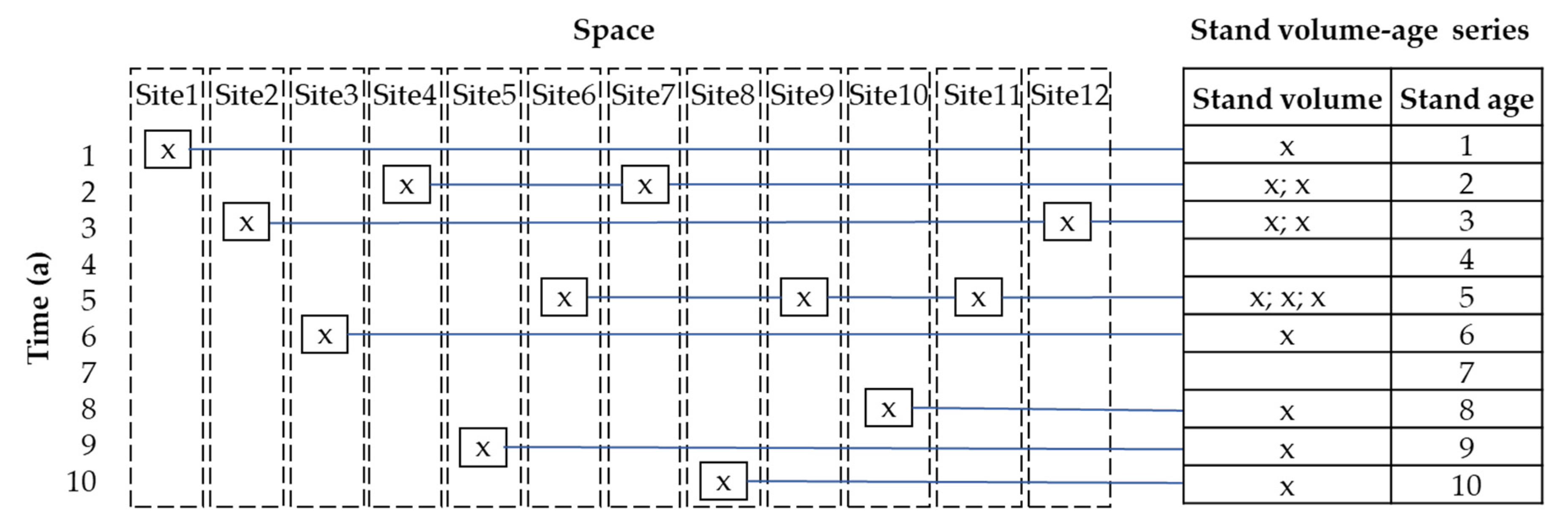
2.3.2. Stand Volume Growth Modeling
Model Fitting
Model Evaluation
3. Results
3.1. Statistics of Stand Volume in 2016
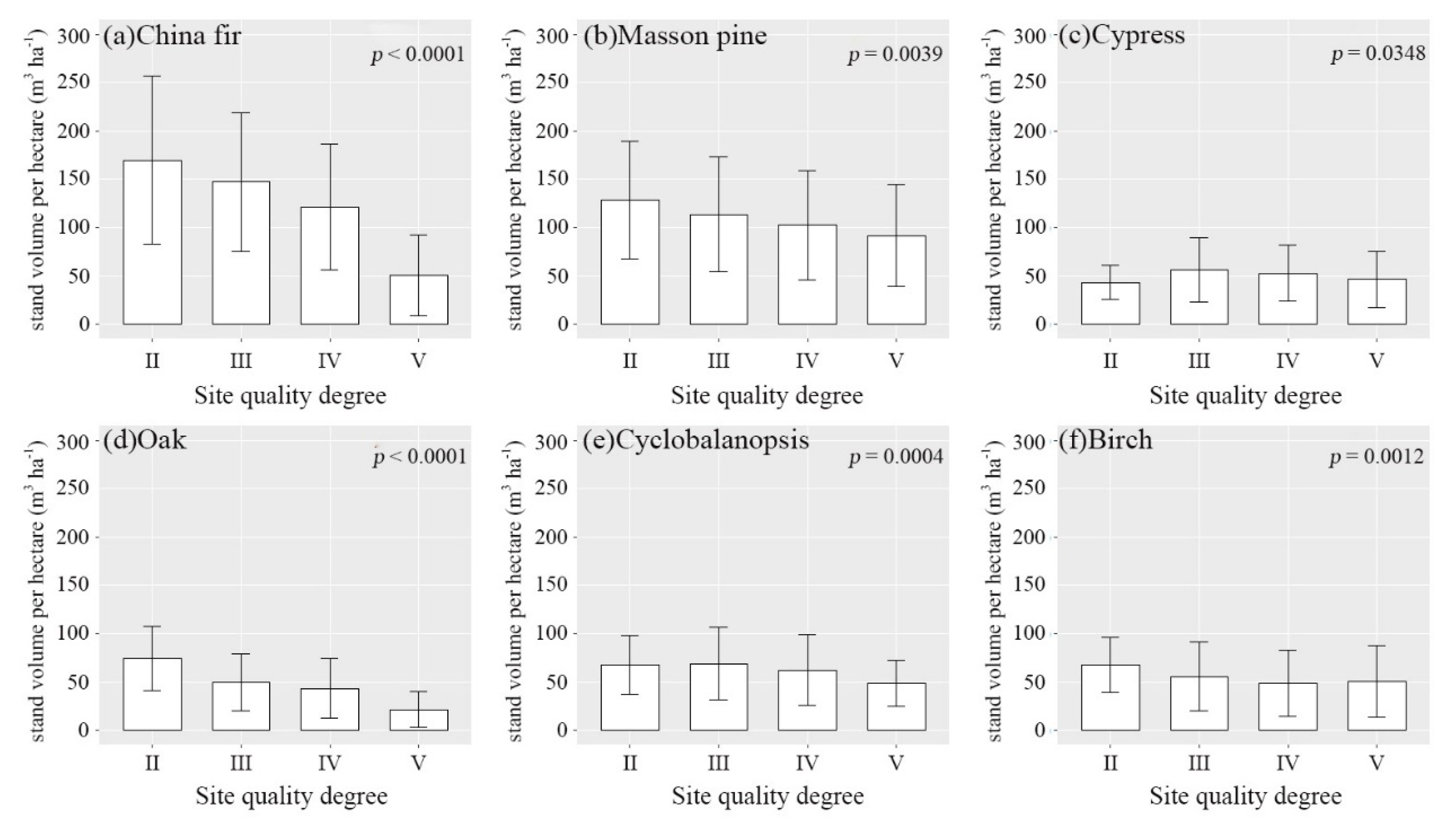
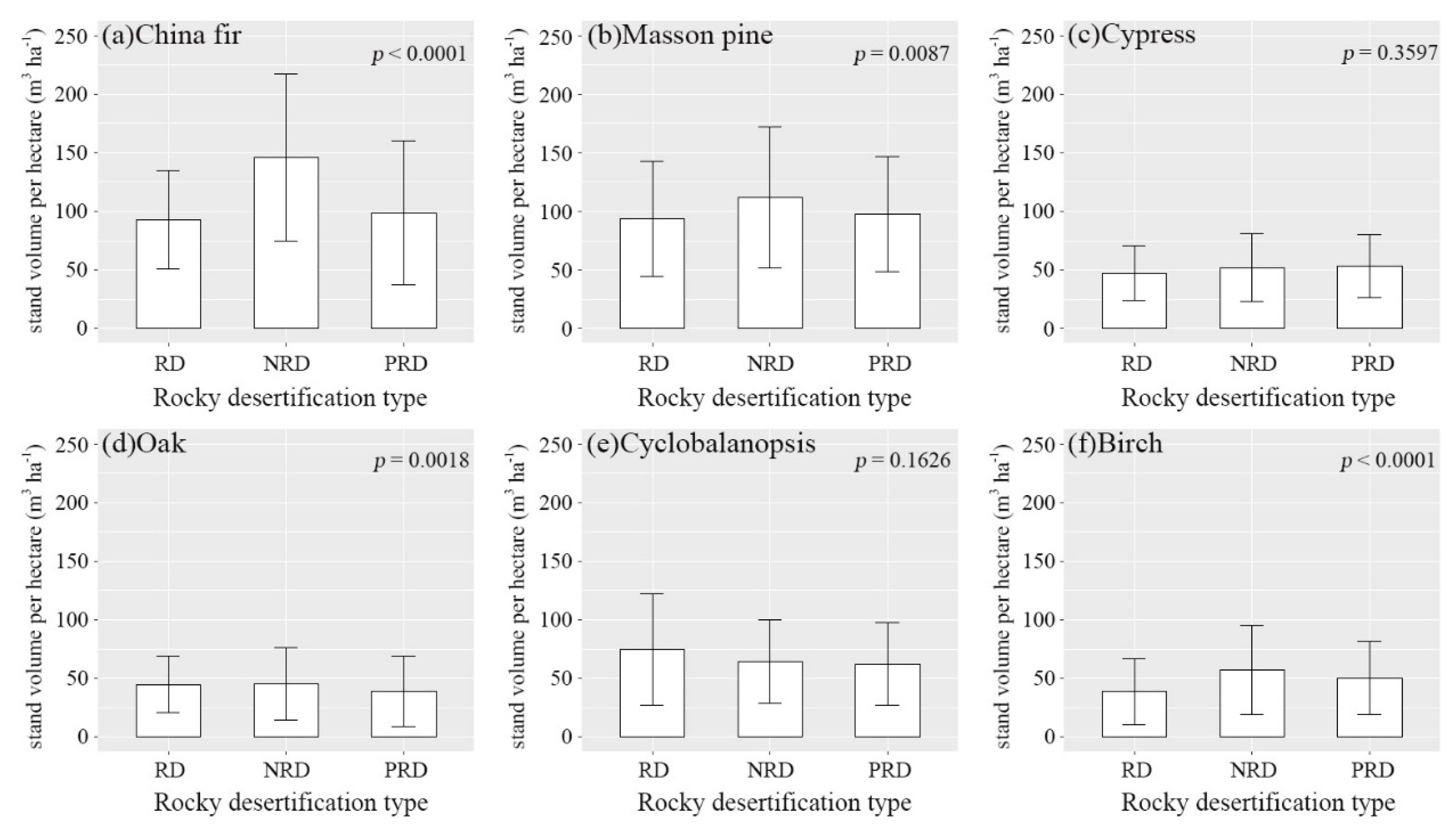
3.2. Environmental Effect on Stand Volume
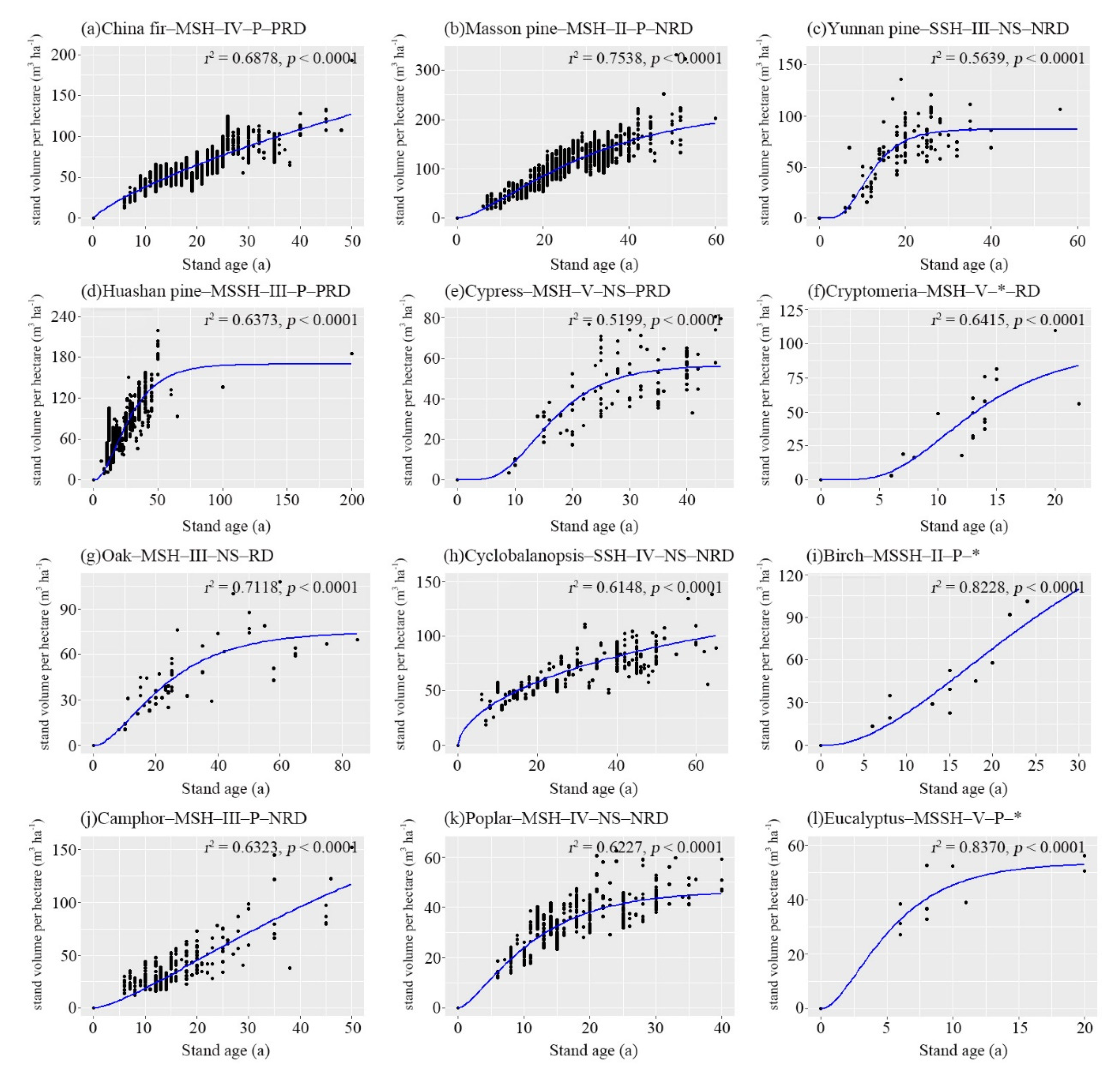
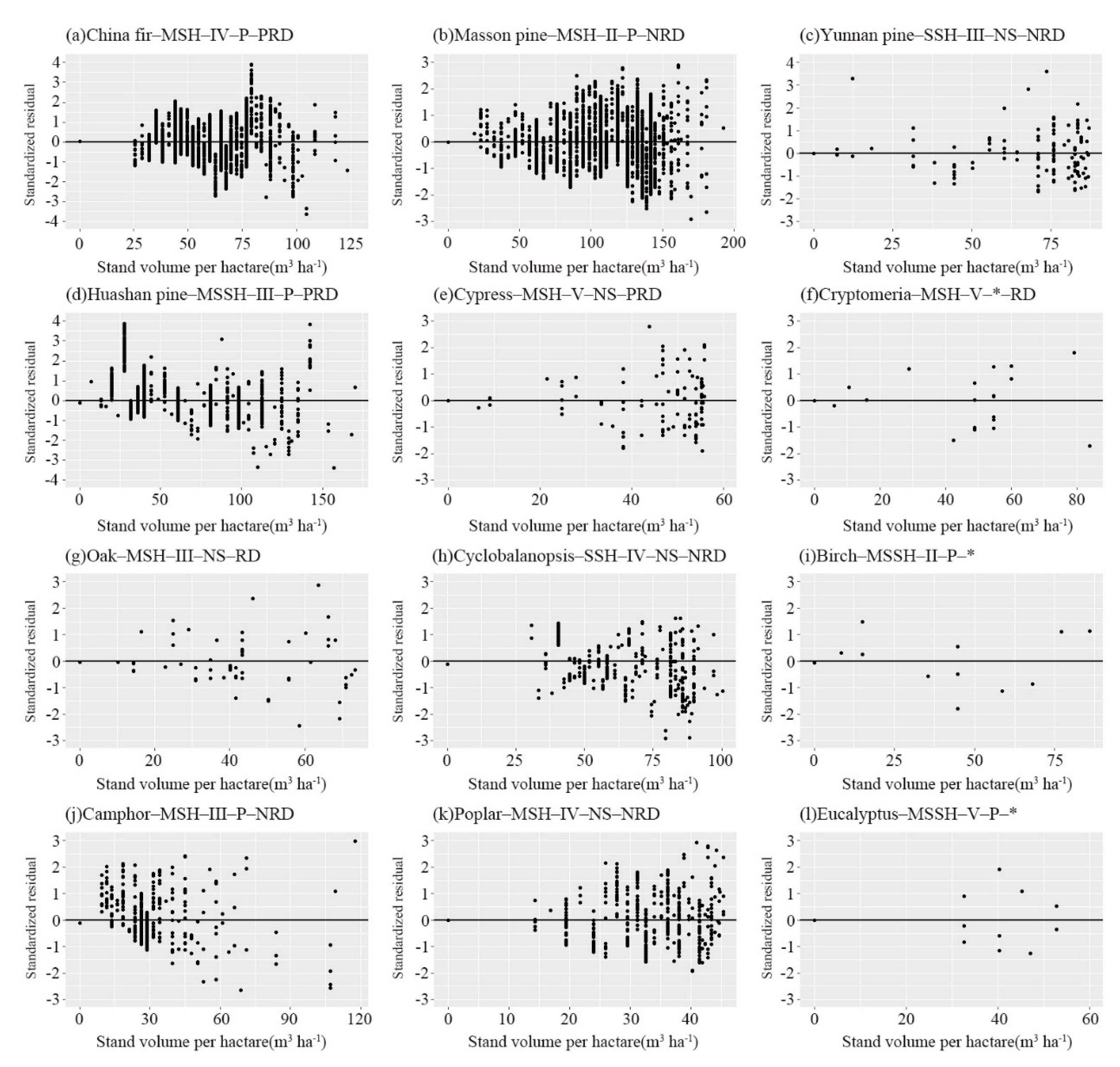
3.3. Growth Modeling of Stand Volume
3.3.1. Goodness-of-Fit
3.3.2. Residual Analysis
3.3.3. Validation
4. Discussion
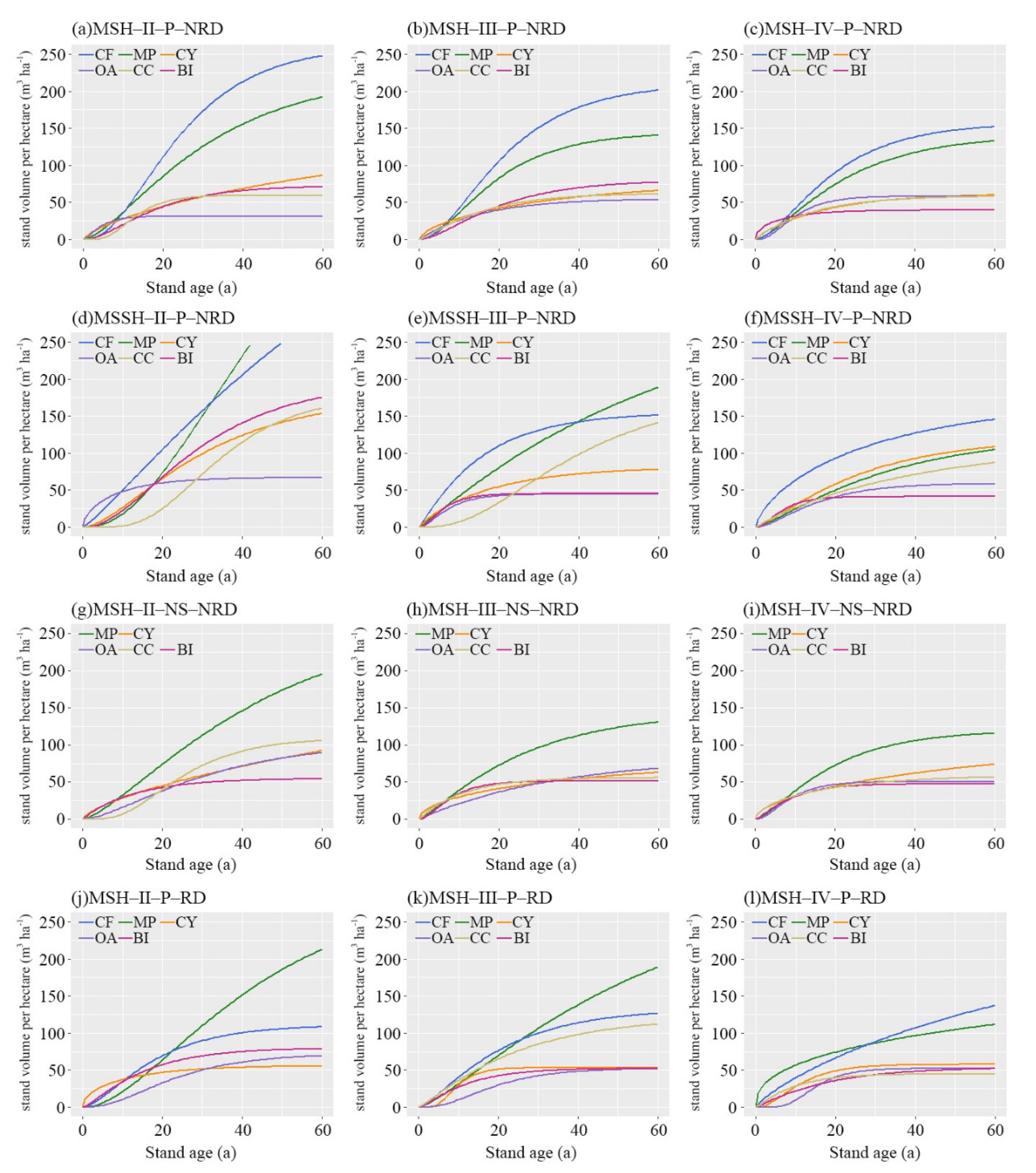
4.1. Growth of Stand Volume in Guizhou Plateau
4.1.1. Growth Rate
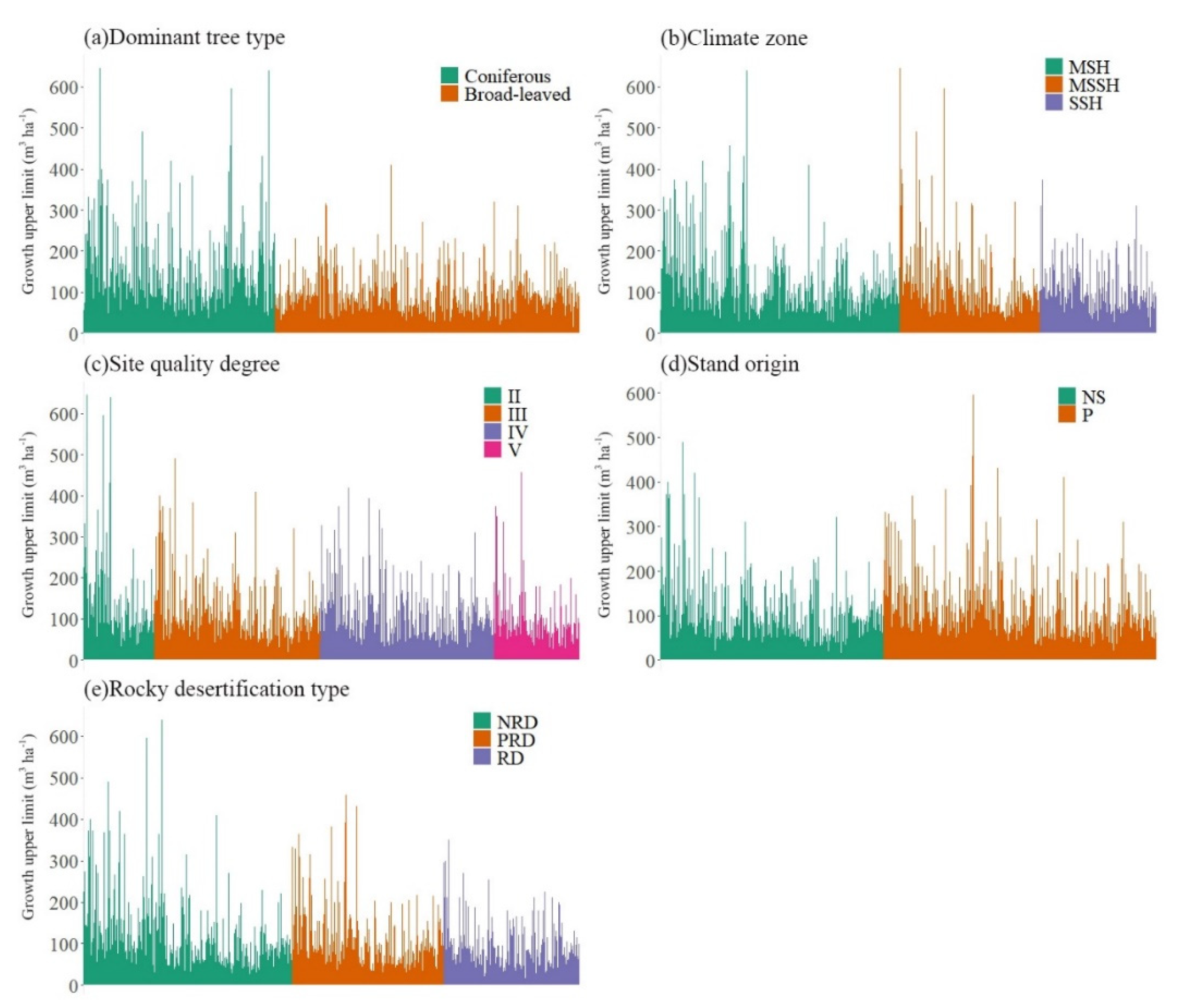
4.1.2. Growth Upper Limit
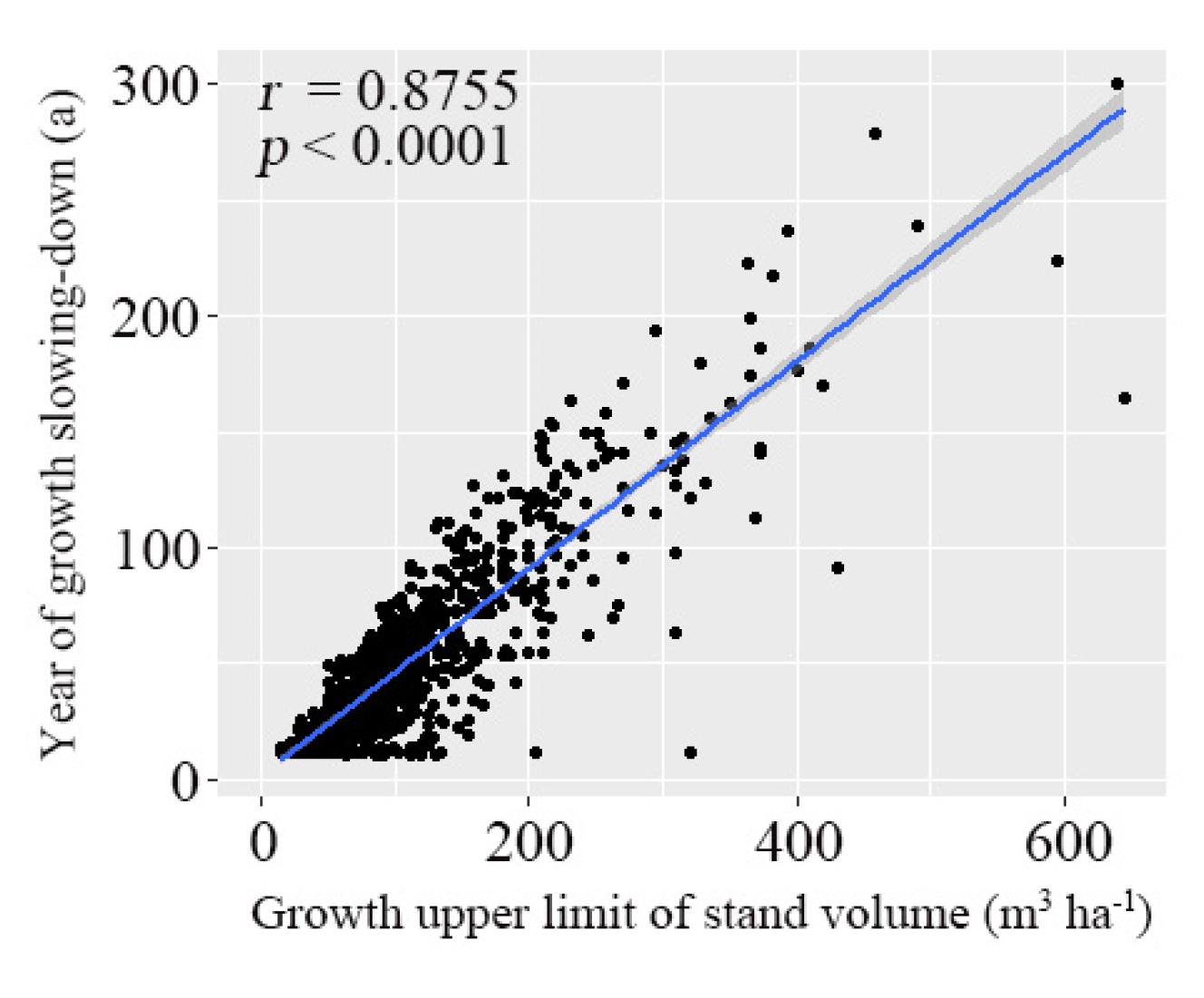
4.1.3. Years of Growth Slowing Down
4.2. Environmental Effect on Stand Volume in Guizhou Plateau
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dube, T.; Sibanda, M.; Shoko, C.; Mutanga, O. Stand-volume estimation from multi-source data for coppiced and high forest Eucalyptus spp. silvicultural systems in KwaZulu-Natal, South Africa. ISPRS J. Photogramm. 2017, 132, 162–169. [Google Scholar] [CrossRef]
- Xu, Y.; Li, C.; Sun, Z.; Jiang, L.; Fang, J. Tree height explains stand volume of closed-canopy stands: Evidence from forest inventory data of China. For. Ecol. Manag. 2019, 438, 51–56. [Google Scholar] [CrossRef]
- Gara, T.W.; Murwira, A.; Chivhenge, E.; Dube, T.; Bangira, T. Estimating wood volume from canopy area in deciduous woodlands of Zimbabwe. South. For. J. For. Sci. 2014, 76, 237–244. [Google Scholar] [CrossRef]
- Akindele, S.O.; LeMay, V.M. Development of tree volume equations for common timber species in the tropical rain forest area of Nigeria. For. Ecol. Manag. 2006, 226, 41–48. [Google Scholar] [CrossRef]
- Gibbs, H.K.; Brown, S.; Niles, J.O.; Foley, J.A. Monitoring and estimating tropical forest carbon stocks: Making REDD a reality. Environ. Res. Lett. 2007, 2, 045023. [Google Scholar] [CrossRef]
- Losi, C.J.; Siccama, T.G.; Condit, R.; Morales, J.E. Analysis of alternative methods for estimating carbon stock in young tropical plantations. For. Ecol. Manag. 2003, 184, 355–368. [Google Scholar] [CrossRef]
- Malimbwi, R.E.; Solberg, B.; Luogak, E. Estimation of biomass and volume in miombo woodland at Kitulangalo Forest Reserve, Tanzania. J. Trop. For. Sci. 1994, 7, 230–242. [Google Scholar]
- Wang, X.; Li, Z.; Liu, X.; Deng, G.; Jiang, Z. Estimating Stem Volume Using QuickBird Imagery and Allometric Relationships for OpenPopulus xiaohei Plantations. J. Integr. Plant Biol. 2007, 49, 1304–1312. [Google Scholar] [CrossRef]
- Peng, C. Growth and yield models for uneven-aged stands: Past, present and future. For. Ecol. Manag. 2000, 132, 259–279. [Google Scholar] [CrossRef]
- Burkhart, H.E.; Tomé, M. Chapter 15: Growth and Yield Models for Uneven-Aged Stands. In Modeling Forest Trees and Stands; Burkhart, H.E., Tomé, M., Eds.; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Ford, D.; Williams, P. Karst Hydrogeology and Geomorphology; John Wiley & Sons Ltd.: Chichester, UK, 2007. [Google Scholar]
- Jiang, Z.; Lian, Y.; Qin, X. Rocky desertification in Southwest China: Impacts, causes, and restoration. Earth Sci. Rev. 2014, 132, 1–12. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Guo, K.; Wang, S.; Liu, H.; Zhao, H.; Qiao, X.; Hou, D.; Li, S. Aboveground carbon stock, allocation and sequestration potential during vegetation recovery in the karst region of southwestern China: A case study at a watershed scale. Agric. Ecosyst. Environ. 2016, 235, 91–100. [Google Scholar] [CrossRef]
- Yuan, D.X. Karst of China, 1st ed.; Geological Publishing House: Beijing, China, 1991. (In Chinese) [Google Scholar]
- Guizhou Forest Editorial Committee. Guizhou Forest, 1st ed.; Guizhou Science and Technology Press; China Forestry Press: Guiyang, China, 1992. (In Chinese) [Google Scholar]
- Zhu, H.; Wang, H.; Li, B.; Sirirugsa, P. Biogeography and Floristic Affinities of the Limestone Flora in Southern Yunnan, China. Ann. Mo. Bot. Gard. 2003, 90, 444–465. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, M.; Bussmann, W.R.; Liu, H.; Liu, Y.; Peng, Y.; Zu, K.; Zhao, Y.; Liu, Z.; Yu, S. Species richness and conservation gap analysis of karst areas: A case study of vascular plants from Guizhou, China. Glob. Ecol. Conserv. 2018, 16, e00460. [Google Scholar] [CrossRef]
- Peng, W.; Song, T.; Zeng, F.; Wang, K.; Du, H.; Lu, S. Relationships between woody plants and environmental factors in karst mixed evergreen-deciduous broadleaf forest, southwest China. J. Food Agric. Environ. 2012, 10, 890–896. [Google Scholar]
- Zhou, Y.; He, X.; Xie, Y.; Wang, M.; Wu, M.; Wu, D. Type Classification for Vegetation Restoration of Karst Mountains in Bijie. Sci. Silvae Sin. 2008, 44, 123–128. (In Chinese) [Google Scholar]
- Xu, G. A Brief Study on Guizhou’s Historical Change of Forest and Vegetation and its Consequences. J. Guizhou Univ. Natl. Philos. Soc. Sci. 2010, 69–73. (In Chinese) [Google Scholar] [CrossRef]
- Wang, L.C.; Lee, D.W.; Zuo, P.; Zhou, Y.K.; Xu, Y.P. Karst environment and eco-poverty in southwestern China: A case study of Guizhou Province. Chin. Geogr. Sci. 2004, 14, 21–27. [Google Scholar] [CrossRef]
- Li, H.; Cai, Y.; Chen, R.; Chen, Q.; Yan, X. Effect assessment of the project of Grain for Green in the karst region in southwestern China: A case study of Bijie Prefecture. Acta Ecol. Sin. 2011, 31, 3255–3264. (In Chinese) [Google Scholar]
- Tang, Y.Z.; Shao, Q.Q. Dataset of Water Conservation of Forest Ecosystem in the Upper Reaches of Wujiang River, China. J. Glob. Chang. Data Discov. 2018, 2, 428–436. [Google Scholar]
- Chi, X.; Zhang, Z.; Xu, X.; Zhang, X.; Zhao, Z.; Liu, Y.; Wang, Q.; Wang, H.; Li, Y.; Yang, G.; et al. Threatened medicinal plants in China: Distributions and conservation priorities. Biol. Conserv. 2017, 210, 89–95. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, G.; Zhu, J.; Ni, J. Aggregated spatial distributions of species in a subtropical karst forest, southwestern China. J. Plant Ecol. 2013, 6, 131–140. [Google Scholar] [CrossRef]
- Chen, H.; Li, D.; Xiao, K.; Wang, K. Soil microbial processes and resource limitation in karst and non-karst forests. Funct. Ecol. 2018, 32, 1400–1409. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Piao, D.; Kim, M.; Choi, G.; Moon, J.; Yu, H.; Lee, W.; Wang, S.; Jeon, S.W.; Son, Y.; Son, Y.; et al. Development of an Integrated DBH Estimation Model Based on Stand and Climatic Conditions. Forests 2018, 9, 155. [Google Scholar] [CrossRef]
- Byun, J.G.; Lee, W.K.; Kim, M.; Kwak, D.A.; Kwak, H.; Park, T.; Byun, W.H.; Son, Y.; Choi, J.K.; Lee, Y.J.; et al. Radial growth response of Pinus densiflora and Quercus spp. to topographic and climatic factors in South Korea. J. Plant Ecol. 2013, 6, 380–392. [Google Scholar] [CrossRef]
- Senf, C.; Müller, J.; Seidl, R. Post-disturbance recovery of forest cover and tree height differ with management in Central Europe. Landscape Ecol. 2019, 34, 2837–2850. [Google Scholar] [CrossRef]
- Luo, H.; Zhang, C. Stand diameter growth model of Pinus yunnanensis in central Yunnan, China. J. Northeast. For. Univ. 2018, 46, 1–6. (In Chinese) [Google Scholar]
- Hunter, I.; Schuck, A. Increasing forest growth in Europe—Possible causes and implications for sustainable forest management. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2002, 136, 133–141. [Google Scholar] [CrossRef]
- Guillemot, J.; Delpierre, N.; Vallet, P.; François, C.; Martin-StPaul, N.K.; Soudani, K.; Nicolas, M.; Badeau, V.; Dufrêne, E. Assessing the effects of management on forest growth across France: Insights from a new functional–structural model. Ann. Bot. Lond. 2014, 114, 779–793. [Google Scholar] [CrossRef]
- Xu, X.; Chen, Z. A preliminary study of the biomass of cultivated Chinese white poplar stands. J. Nanjing For. Univ. Nat. Sci. Ed. 1987, 130–136. (In Chinese) [Google Scholar] [CrossRef]
- Monserud, R.A.; Huang, S.; Yang, Y. Predicting lodgepole pine site index from climatic parameters in Alberta. Forest. Chron. 2006, 82, 562–571. [Google Scholar] [CrossRef]
- Yu, L.; Lei, X.; Wang, Y.; Yang, Y.; Wang, Q. Impact of climate on individual tree radial growth based on generalized additive model. J. Beijing For. Univ. 2014, 36, 22–32. (In Chinese) [Google Scholar]
- Zirlewagen, D.; Raben, G.; Weise, M. Zoning of forest health conditions based on a set of soil, topographic and vegetation parameters. For. Ecol. Manag. 2007, 248, 43–55. [Google Scholar] [CrossRef]
- Nigh, G.D.; Ying, C.C.; Qian, H. Climate and Productivity of Major Conifer Species in the Interior of British Columbia, Canada. For. Sci. 2004, 50, 659–671. [Google Scholar]
- Li, W.; Liu, S. Large scaled cedar DBH growth models including climatic variables. J. Cent. South Univ. For. Technol. 2015, 35, 74–77. (In Chinese) [Google Scholar]
- Fontes, L.; Tomé, M.; Thompson, F.; Yeomans, A.; Luis, J.S.; Savill, P. Modelling the Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) site index from site factors in Portugal. Forestry 2003, 76, 491–507. [Google Scholar] [CrossRef]
- Lu, J.; Li, F.; Zhang, H.; Zhang, S. Simulation of foliage distribution for major broad-leaved species in secondary forest in Mao’er Mountain. Sci. Silvae Sin. 2011, 47, 114–120. (In Chinese) [Google Scholar]
- Lang, R.; Xu, J.; Timm, T.; Yang, X.; Bi, Y. A study of stand growth model for Pinus yunnanensis (Pinaceae) based on plots data: A case study in Yangliu Township, Baoshan, Yunnan Province. Plant Divers. Resour. 2011, 33, 357–363. (In Chinese) [Google Scholar]
- Nothdurft, A.; Kublin, E.; Lappi, J. A non-linear hierarchical mixed model to describe tree height growth. Eur. J. For. Res. 2006, 125, 281–289. [Google Scholar] [CrossRef]
- King, J.E. Site index curves for Douglas-fir in the pacific northwest. In Weyerhaeuser Forestry Paper No.8; Weyerhaeuser Forestry Research Center: Centralia, WA, USA, 1966; Volume 8. [Google Scholar]
- Herman, F.R.; Curtis, R.O.; DeMars, D.J. Height Growth and Site Index Estimates for Noble Fir in High-Elevation Forests of the Oregon-Washington Cascades; U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1978.
- Cochran, P.H. Site Index, Height Growth, Normal Yields, and Stocking Levels for Larch in Oregon and Washington; U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1985.
- Wensel, L.C.; Krumland, B. A Site Index System for Redwood and Douglas-fir in California’s North Coast Forest. Hilgardia 1986, 54, 1–14. [Google Scholar] [CrossRef]
- Yu, G.; Chen, Z.; Piao, S.; Peng, C.; Ciais, P.; Wang, Q.; Li, X.; Zhu, X. High carbon dioxide uptake by subtropical forest ecosystems in the East Asian monsoon region. Proc. Natl. Acad. Sci. USA 2014, 111, 4910–4915. [Google Scholar] [CrossRef] [PubMed]
- Teodoridis, V.; Kovar-Eder, J.; Mazouch, P. Intergrated plant record (IPR) vegetation analysis applied to modern vegetation in south China and Japan. Palaios 2011, 26, 623–638. [Google Scholar] [CrossRef]
- Huang, Q.; Cai, Y. Assessment of karst rocky desertification using the radial basis function network model and GIS technique: A case study of Guizhou Province, China. Environ. Geol. 2006, 49, 1173–1179. [Google Scholar]
- Yu, N.; Li, S. Current situation of rocky desertification and the main rehabilitation measures in Guizhou Province. J. Anhui Agric. Sci. 2014, 42, 8702–8704. (In Chinese) [Google Scholar]
- Guizhou Forestry Bureau. Detailed Rules for the Implementation of Fourth Forest Resources Planning and Design Survey of Guizhou Province; Guizhou Forestry Bureau: Guiyang, China, 2015. (In Chinese)
- Tomppo, E.; Gschwantner, T.; Lawrence, M.; McRoberts, R.E. National Forest Inventories: Pathways for Common Reporting; Springer: Heidelberg, Germany, 2010. [Google Scholar]
- Tang, Y.Z.; Shao, Q.Q. Water conservation capacity of forest ecosystem and its spatial variation in the upper reaches of Wujiang River. J. Geo Inf. Sci. 2016, 18, 987–999. (In Chinese) [Google Scholar]
- Shi, P.; Wu, B.; Cheng, G.; Luo, J. Water retention capacity evaluation of main forest vegetation types in the upper Yangtze basin. J. Nat. Resour. 2004, 19, 351–360. (In Chinese) [Google Scholar]
- Hutchinson, M.F. Interpolating mean rainfall using thin plate smoothing splines. Int. J. Geogr. Inf. Syst. 1995, 9, 385–403. [Google Scholar] [CrossRef]
- Hutchinson, M.F. Interpolation of Rainfall Data with Thin Plate Smoothing Splines: II. Analysis of Topographic Dependence. J. Geogr. Inf. Decis. Anal. 1998, 2, 152–167. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- MathWorks, I. Curve Fitting Toolbox: For Use with MATLAB®: User’s Guide; MathWorks: Natick, MA, USA, 2002. [Google Scholar]
- National Forestry Administration. Technical Regulations for Defining Forest Land Border in Forest Land Planning on Protection and Utilization; National Forestry Administration: Beijing, China, 2011. (In Chinese)
- Wilcoxon, F. Individual comparisons of grouped data by ranking methods. J. Econ. Entomol. 1946, 39, 269–270. [Google Scholar] [CrossRef]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Richards, F.J. A Flexible Growth Function for Empirical Use. J. Exp. Bot. 1959, 10, 290–301. [Google Scholar] [CrossRef]
- Von Bertalanffy, L. Quantitative Laws in Metabolism and Growth. Q. Rev. Biol. 1957, 32, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Osumi, S. Growth Models for Total and Average Basal Area in Even-Aged Pure Stands Based on the Richards Growth Function (I): Derivation of the Models. J. Jpn. For. Soc. 1985, 67, 434–441. [Google Scholar]
- Liu, Z.; Li, F. The generalized Chapman-Richards function and applications to tree and stand growth. J. For. Res. 2003, 14, 19–26. [Google Scholar]
- Shoda, T.; Imanishi, J.; Shibata, S. Growth characteristics and growth equations of the diameter at breast height using tree ring measurements of street trees in Kyoto City, Japan. Urban For. Urban Green. 2020, 49, 126627. [Google Scholar] [CrossRef]
- Wei, X.; Sun, Y.; Ma, W. A height growth model for Cunninghamia Lanceolata based on Richards’ equation. J. Zhejiang A F Univ. 2012, 29, 661–666. (In Chinese) [Google Scholar]
- Duan, A.; Zhang, J.; Tong, S. Application of six growth equations on stands diameter structure of Chinese fir plantations. For. Res. 2003, 16, 423–429. (In Chinese) [Google Scholar]
- Pickett, S.T.A. Space-for-Time Substitution as an Alternative to Long-Term Studies. In Long-Term Studies in Ecology; Likens, G.E., Ed.; Springer: New York, NY, USA, 1989; pp. 110–135. [Google Scholar]
- Ma, J.; Xiao, X.; Bu, R.; Doughty, R.; Hu, Y.; Chen, B.; Li, X.; Zhao, B. Application of the space-for-time substitution method in validating long-term biomass predictions of a forest landscape model. Environ. Model. Softw. 2017, 94, 127–139. [Google Scholar] [CrossRef]
- Blois, J.L.; Williams, J.W.; Fitzpatrick, M.C.; Jackson, S.T.; Ferrier, S. Space can substitute for time in predicting climate-change effects on biodiversity. Proc. Natl. Acad. Sci. USA 2013, 110, 9374–9379. [Google Scholar] [CrossRef]
- Feng, Y. The Research on Carbon Budget of Forest Ecosystem in Pu’er Region of Yunnan Province Based on CBM Model. Master’s Thesis, Chinese Academy of Forestry, Beijing, China, 2014. (In Chinese). [Google Scholar]
- Zeng, W.; Wang, X.; Chen, Z.; Yao, S. Analysis of impacts of forest origin on single tree biomass models. For. Resour. Manag. 2014, 40–45. (In Chinese) [Google Scholar] [CrossRef]
- Steinnocher, K.; De Bono, A.; Chatenoux, B.; Tiede, D.; Wendt, L. Estimating urban population patterns from stereo-satellite imagery. Eur. J. Remote Sens. 2019, 52, 12–25. [Google Scholar] [CrossRef]
- Yao, Z.; Liu, J. Models for biomass estimation of four shrub species planted in urban area of Xi’an City, Northwest China. Chin. J. Appl. Ecol. 2014, 25, 111–116. (In Chinese) [Google Scholar]
- Boudewyn, P.A.; Song, X.; Magnussen, S.; Gillis, M.D. Model-Based, Volume-to-Biomass Conversion for Forested and Vegetated Land in Canada; Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 2007.
- Niemand, C.; Köstner, B.; Prasse, H.; Grünwald, T.; Bernhofer, C. Relating tree phenology with annual carbon fluxes at Tharandt forest. Meteorologische Zeitschrift 2005, 14, 197–202. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Viovy, N.; Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cycles 2007, 21, GB3018. [Google Scholar] [CrossRef]
- Potter, C.; Klooster, S.; Genovese, V. Net primary production of terrestrial ecosystems from 2000 to 2009. Clim. Chang. 2012, 115, 365–378. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, L.; Fensholt, R.; Wang, K.; Vitkovskaya, I.; Tian, F. Climate Contributions to Vegetation Variations in Central Asian Drylands: Pre- and Post-USSR Collapse. Remote Sens. 2015, 7, 2449–2470. [Google Scholar] [CrossRef]
- Piao, S.; Yin, G.; Tan, J.; Cheng, L.; Huang, M.; Li, Y.; Liu, R.; Mao, J.; Myneni, R.B.; Peng, S.; et al. Detection and attribution of vegetation greening trend in China over the last 30 years. Glob. Chang. Biol. 2015, 21, 1601–1609. [Google Scholar] [CrossRef]
- Zhang, Q.; Qi, T.; Li, J.; Singh, V.P.; Wang, Z. Spatiotemporal variations of pan evaporation in China during 1960–2005: Changing patterns and causes. Int. J. Climatol. 2015, 35, 903–912. [Google Scholar] [CrossRef]
- Cui, L.; Wang, L.; Qu, S.; Deng, L.; Wang, Z. Impacts of temperature, precipitation and human activity on vegetation NDVI in Yangtze river basin, China. Earth Sci. 2019, 45, 1–24. (In Chinese) [Google Scholar]
- Chen, Y.; Shi, X.; Zhang, L.; Baskin, J.M.; Baskin, C.C.; Liu, H.; Zhang, D. Effects of increased precipitation on the life history of spring- and autumn-germinated plants of the cold desert annual Erodium oxyrhynchum (Geraniaceae). Aob Plants 2019, 11, plz004. [Google Scholar] [CrossRef]
- Hui, D.; Yu, C.; Deng, Q.; Dzantor, E.K.; Zhou, S.; Dennis, S.; Sauve, R.; Johnson, T.L.; Fay, P.A.; Shen, W.; et al. Effects of precipitation changes on switchgrass photosynthesis, growth, and biomass: A mesocosm experiment. PLoS ONE 2018, 13, e0192555. [Google Scholar] [CrossRef] [PubMed]
- Dube, O.P.; Pickup, G. Effects of rainfall variability and communal and semi-commercial grazing on land cover in southern African rangelands. Clim. Res. 2001, 17, 195–208. [Google Scholar] [CrossRef]
- Cao, S.; Chen, L.; Yu, X. Impact of China’s Grain for Green Project on the landscape of vulnerable arid and semi-arid agricultural regions: A case study in northern Shaanxi Province. J. Appl. Ecol. 2009, 46, 536–543. [Google Scholar] [CrossRef]
- Li, Z.; Chen, Y.; Li, W.; Deng, H.; Fang, G. Potential impacts of climate change on vegetation dynamics in Central Asia. J. Geophys. Res. Atmos. 2015, 120, 12345–12356. [Google Scholar] [CrossRef]
- Mizutani, F.; Yamada, M.; Tomana, T. Differential Water Tolerance and Ethanol Accumulation in Prunus Species under Flooded Conditions. J. Jpn. Soc. Hortic. Sci. 1982, 51, 29–34. [Google Scholar] [CrossRef]
- Crawford, R.M.M. Alcohol Dehydrogenase Activity in Relation to Flooding Tolerance in Roots. J. Exp. Bot. 1967, 18, 458–464. [Google Scholar] [CrossRef]
- Wang, J.; Wang, K.; Zhang, M.; Zhang, C. Impacts of climate change and human activities on vegetation cover in hilly southern China. Ecol. Eng. 2015, 81, 451–461. [Google Scholar] [CrossRef]
- Wimmer, R.; Grabner, M. A comparison of tree-ring features in Picea abies as correlated with climate. IAWA J. 2000, 21, 403–416. [Google Scholar] [CrossRef]
- Rezaei, S.A.; Gilkes, R.J.; Andrews, S.S. A minimum data set for assessing soil quality in rangelands. Geoderma 2006, 136, 229–234. [Google Scholar] [CrossRef]
- Mielke, L.N.; Schepers, J.S. Plant response to topsoil thickness on an eroded loess soil. J. Soil Water Conserv. 1986, 41, 59–63. [Google Scholar]
- Gao, S.; Yuan, X.; Gan, S.; Zhang, X. Study of the relationship between soil thickness and vegetation growth in the karst region in southeast Yunnan. Value Eng. 2013, 32, 178–179. (In Chinese) [Google Scholar]
- Pellissier, L.; Fournier, B.; Guisan, A.; Vittoz, P. Plant traits co-vary with altitude in grasslands and forests in the European Alps. Plant Ecol. 2010, 211, 351–365. [Google Scholar] [CrossRef]
- Hodkinson, I.D. Terrestrial insects along elevation gradients: Species and community responses to altitude. Biol. Rev. 2005, 80, 489–513. [Google Scholar] [CrossRef] [PubMed]
- McCool, D.K.; George, G.O.; Freckleton, M.; Douglas, C.L.; Papendick, R.I. Topographic effect on erosion from cropland in the northwestern wheat region. Trans. ASAE 1993, 36, 1067–1071. [Google Scholar] [CrossRef]
- Liu, B.Y.; Nearing, M.A.; Risse, L.M. Slope gradient effects on soil loss for steep slopes. Trans. ASAE 1994, 37, 1835–1840. [Google Scholar] [CrossRef]
- El-Hassanin, A.S.; Labib, T.M.; Gaber, E.I. Effect of vegetation cover and land slope on runoff and soil losses from the watersheds of Burundi. Agric. Ecosyst. Environ. 1993, 43, 301–308. [Google Scholar] [CrossRef]
- Kuo, T.; Liu, Z.; Li, H.; Wan, N.; Shen, C.; Ku, T. Climate and environmental changes during the past millennium in central western Guizhou, China as recorded by Stalagmite ZJD-21. J. Asian Earth Sci. 2011, 40, 1111–1120. [Google Scholar] [CrossRef]
- Cai, Y.; Meng, J. Ecological reconstruction of degraded land: A social approach. Sci. Geogr. Sin. 1999, 19, 198–204. (In Chinese) [Google Scholar]
- Damgaard, C. A Critique of the Space-for-Time Substitution Practice in Community Ecology. Trends Ecol. Evol. 2019, 34, 416–421. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Unit | Classification | Range/Categories | Notes |
|---|---|---|---|---|
| Temperature | °C | Continuous | 6.95–21.67 | Mean annual temperature in 2016, accurate to 2 decimal places |
| Precipitation | mm | Continuous | 832.11–1994.77 | Sum annual precipitation in 2016, accurate to 2 decimal places |
| Stand origin | -- | Categorical | Natural stand; Plantation | Whether a stand grow up naturally or by cultivated |
| Elevation | m | Continuous | 145–2900 | The height above sea level, accurate to integer |
| Slope gradient | ° | Continuous | 1–90 | The degree of slope, accurate to integer |
| Aspect | -- | Categorical | North; Northeast; East; Southeast; South; Southwest; West; Northwest; Flat | The direction in which the normal of a slope is projected onto a horizontal plane, was divided into 9 classes. |
| Slope position | -- | Categorical | Ridge; Upper slope; Middle slope; Lower slope; Valley; Flat ground; All slope | The geomorphic part located in a slope, was divided into 7 classes. |
| Topsoil thickness | cm | Continuous | 1–115 | Sum of the thickness of A horizon and B horizon, accurate to an integer. |
| Site quality degree | -- | Ordinal | I (removed); II; III; IV; V | The degree from I to V represents the site quality from high to low. As the forestland denoted by degree I in the study area were very rare and cannot provide sufficient samples, we remove it in the following analysis. |
| Rocky desertification type | -- | Categorical | Rocky desertified (RD); Potential rocky desertified (PRD); Nonrocky desertified (NRD) | RD refers to the stands with ≥ 30% bedrock exposed and < 50% vegetation-covered; PRD refers to the stands with ≥ 30% bedrock exposed and ≥ 50% vegetation-covered; NRD refers to the stands with < 30% bedrock exposed. |
| Rocky desertification degree | -- | Categorical | Slight; Moderate; Severe; Extremely severe (removed) | Only those stands denoted by RD and PRD have a value of rocky desertification degree. The land extremely severe desertified could hardly grow any vegetation so we remove it in the following analysis. |
| No | Dominant Tree Species (Groups) | Canopy Density | Stand Age(a) | Stand DBH (cm) | Stand Height (m) | Total Stand Volume (103 m3) | Stand Volume (m3 ha−1) | SD of Stand Volume (m3 ha−1) |
|---|---|---|---|---|---|---|---|---|
| 1 | Fir | 0.59 | 19.96 | 14.37 | 9.10 | 29.28 | 56.44 | 42.44 |
| 2 | Spruce | 0.52 | 19.52 | 12.04 | 8.66 | 29.69 | 47.33 | 37.65 |
| 3 | Chinese yew | 0.12 | 6.03 | 2.35 | 2.18 | 15.03 | 3.89 | 24.28 |
| 4 | Masson pine | 0.55 | 21.89 | 15.78 | 11.72 | 135,847.70 | 80.65 | 58.14 |
| 5 | Huashan pine | 0.59 | 22.15 | 14.27 | 8.85 | 10,356.21 | 65.94 | 53.41 |
| 6 | Yunnan pine | 0.61 | 24.37 | 12.80 | 8.48 | 11,993.18 | 69.08 | 42.14 |
| 7 | Other pine | 0.50 | 12.92 | 10.32 | 7.56 | 740.61 | 47.82 | 51.85 |
| 8 | Keteleeria | 0.55 | 26.80 | 15.01 | 8.33 | 195.64 | 51.89 | 41.87 |
| 9 | Hemlock | 0.61 | 40.89 | 20.87 | 10.93 | 4.09 | 43.25 | 25.55 |
| 10 | Cypress | 0.48 | 20.67 | 10.35 | 8.10 | 12,000.31 | 37.06 | 31.62 |
| 11 | Other cypress | 0.39 | 14.53 | 6.92 | 5.91 | 931.57 | 23.18 | 34.20 |
| 12 | China fir | 0.48 | 15.20 | 10.99 | 8.49 | 13,8317.35 | 81.59 | 70.49 |
| 13 | Cryptomeria | 0.56 | 13.72 | 11.04 | 8.32 | 11,964.14 | 70.83 | 66.02 |
| 14 | Metasequoia | 0.62 | 15.06 | 11.58 | 9.11 | 84.02 | 48.37 | 51.90 |
| 15 | Other fir | 0.53 | 23.20 | 13.70 | 8.54 | 69.34 | 51.77 | 52.01 |
| 16 | OCTS 1 | 0.55 | 22.26 | 13.93 | 9.21 | 86.76 | 43.73 | 65.58 |
| 17 | Poplar/aspen | 0.48 | 14.68 | 12.65 | 10.39 | 2698.11 | 33.68 | 32.32 |
| 18 | Willow | 0.36 | 9.90 | 8.01 | 6.16 | 18.20 | 22.12 | 40.68 |
| 19 | Eucalyptus | 0.53 | 6.74 | 9.05 | 10.20 | 2747.77 | 61.06 | 46.50 |
| 20 | Camphor | 0.49 | 15.35 | 10.20 | 7.26 | 313.11 | 32.55 | 35.38 |
| 21 | Phoebe | 0.45 | 22.79 | 12.15 | 8.70 | 187.71 | 60.59 | 40.67 |
| 22 | Oak | 0.56 | 18.62 | 9.97 | 7.69 | 7990.99 | 36.24 | 29.96 |
| 23 | Cyclobalanopsis | 0.51 | 12.73 | 8.90 | 8.42 | 9732.52 | 45.83 | 27.63 |
| 24 | Beech | 0.54 | 16.21 | 12.31 | 9.01 | 180.95 | 62.54 | 27.54 |
| 25 | Birch | 0.50 | 14.75 | 11.02 | 8.95 | 5786.67 | 32.51 | 25.81 |
| 26 | Basswood | 0.65 | 33.55 | 16.84 | 11.69 | 6.96 | 32.65 | 44.47 |
| 27 | Locust | 0.48 | 22.16 | 12.86 | 9.03 | 702.96 | 37.43 | 35.65 |
| 28 | Katus | 0.50 | 17.53 | 15.41 | 10.48 | 933.59 | 53.83 | 29.10 |
| 29 | Maple | 0.46 | 18.03 | 16.09 | 11.98 | 127.46 | 75.47 | 30.09 |
| 30 | Melia | 0.57 | 20.28 | 11.94 | 9.30 | 122.99 | 38.34 | 34.35 |
| 31 | Chinese toon | 0.63 | 31.74 | 16.23 | 11.39 | 814.76 | 38.31 | 38.94 |
| 32 | Elm | 0.55 | 21.63 | 14.57 | 10.43 | 255.87 | 29.41 | 23.73 |
| 33 | Ebony | 0.55 | 22.18 | 12.02 | 9.49 | 56.82 | 72.72 | 30.75 |
| 34 | Firmiana | 0.46 | 20.20 | 19.67 | 12.46 | 206.55 | 38.56 | 35.03 |
| 35 | OBTS 2 | 0.52 | 19.01 | 11.87 | 9.47 | 69,078.98 | 41.45 | 35.46 |
| 36 | BMTS 3 | 0.55 | 21.19 | 11.89 | 9.22 | 27,841.36 | 58.71 | 38.15 |
| Environmental Factors | Dominant Tree Species (Groups) | Slope | Spearman’s ρ | p-Value |
|---|---|---|---|---|
| Temperature | China fir | 12.76 | 0.2838 | <0.0001 ** |
| Masson pine | 0.29 | 0.0051 | 0.3366 | |
| Cypress | −0.65 | −0.0173 | 0.5520 | |
| Oak | −1.59 | −0.1023 | <0.0001 ** | |
| Cyclobalanopsis | 2.03 | 0.1543 | <0.0001 ** | |
| Birch | 5.38 | 0.2569 | <0.0001 ** | |
| Precipitation | China fir | 0.13 | 0.2833 | <0.0001 ** |
| Masson pine | −0.01 | −0.0305 | 0.5409 | |
| Cypress | 0.01 | 0.0510 | 0.0373 * | |
| Oak | −0.01 | −0.0921 | 0.0036 ** | |
| Cyclobalanopsis | 0.02 | 0.0729 | 0.0214 * | |
| Birch | 0.07 | 0.1742 | <0.0001 ** | |
| Elevation | China fir | −0.05 | −0.2453 | <0.0001 ** |
| Masson pine | −0.005 | −0.0101 | 0.2422 | |
| Cypress | 0.01 | 0.0621 | 0.0863 | |
| Oak | 0.01 | 0.1309 | <0.0001 ** | |
| Cyclobalanopsis | −0.01 | −0.0716 | 0.0236 * | |
| Birch | −0.02 | −0.1702 | <0.0001 ** | |
| Slope gradient | China fir | −0.51 | −0.0545 | 0.0267 * |
| Masson pine | −0.42 | −0.0515 | 0.0448 * | |
| Cypress | −0.15 | −0.0430 | 0.1745 | |
| Oak | −0.60 | −0.2921 | <0.0001 ** | |
| Cyclobalanopsis | −0.38 | −0.1129 | 0.0097 ** | |
| Birch | −0.43 | −0.1235 | <0.0001 ** | |
| Topsoil thickness | China fir | 0.84 | 0.2652 | <0.0001 ** |
| Masson pine | 0.58 | 0.1958 | <0.0001 ** | |
| Cypress | 0.25 | 0.1391 | 0.0024 ** | |
| Oak | 0.22 | 0.1668 | <0.0001 ** | |
| Cyclobalanopsis | 0.37 | 0.2202 | <0.0001 ** | |
| Birch | 0.28 | 0.1444 | 0.0002 ** |
| Dominant Tree Species (Groups) | Natural Stand | Plantation | Wilcoxon Test p-Value | ||||
|---|---|---|---|---|---|---|---|
| Mean | SD | CV | Mean | SD | CV | ||
| China fir | 141.9 | 71.34 | 50.28 | -- | -- | -- | -- |
| Masson pine | 95.28 | 57.06 | 59.88 | 112.9 | 58.7 | 51.99 | 0.0007 ** |
| Cypress | 55.6 | 25.64 | 46.12 | 51.61 | 32.06 | 62.11 | 0.0009 ** |
| Oak | 42.9 | 30.48 | 71.04 | 58.99 | 32.11 | 54.43 | 0.0030 ** |
| Cyclobalanopsis | 64.75 | 37.08 | 57.27 | 63.86 | 34.79 | 54.48 | 0.8700 |
| Birch | 51.49 | 34.38 | 66.76 | 62.21 | 39.92 | 64.18 | 0.0040 ** |
| Environmental Factors | China Fir | Masson Pine | Cypress | Oak | Cyclobalanopsis | Birch | Sum | Rank |
|---|---|---|---|---|---|---|---|---|
| Temperature | ** | ** | ** | ** | 4 | 6 | ||
| Precipitation | ** | * | ** | * | ** | 5 | 3 | |
| Stand origin | -- | ** | ** | ** | ** | 4 | 5 | |
| Elevation | ** | ** | * | ** | 4 | 8 | ||
| Slope gradient | * | * | ** | ** | ** | 5 | 3 | |
| Aspect | ** | * | 2 | 10 | ||||
| Slope position | ** | 1 | 11 | |||||
| Topsoil thickness | ** | ** | ** | ** | ** | ** | 6 | 1 |
| Site quality degree | ** | ** | * | ** | ** | ** | 6 | 2 |
| Rocky desertification type | ** | ** | ** | ** | 4 | 6 | ||
| Rocky desertification degree | * | * | ** | 3 | 9 | |||
| Sum | 8 | 6 | 5 | 10 | 7 | 9 | -- | -- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Shao, Q.; Shi, T.; Wu, G. Developing Growth Models of Stand Volume for Subtropical Forests in Karst Areas: A Case Study in the Guizhou Plateau. Forests 2021, 12, 83. https://doi.org/10.3390/f12010083
Tang Y, Shao Q, Shi T, Wu G. Developing Growth Models of Stand Volume for Subtropical Forests in Karst Areas: A Case Study in the Guizhou Plateau. Forests. 2021; 12(1):83. https://doi.org/10.3390/f12010083
Chicago/Turabian StyleTang, Yuzhi, Quanqin Shao, Tiezhu Shi, and Guofeng Wu. 2021. "Developing Growth Models of Stand Volume for Subtropical Forests in Karst Areas: A Case Study in the Guizhou Plateau" Forests 12, no. 1: 83. https://doi.org/10.3390/f12010083
APA StyleTang, Y., Shao, Q., Shi, T., & Wu, G. (2021). Developing Growth Models of Stand Volume for Subtropical Forests in Karst Areas: A Case Study in the Guizhou Plateau. Forests, 12(1), 83. https://doi.org/10.3390/f12010083