Role of Species and Planting Configuration on Transpiration and Microclimate for Urban Trees


 and
and 
Abstract
1. Introduction
2. Materials and Methods
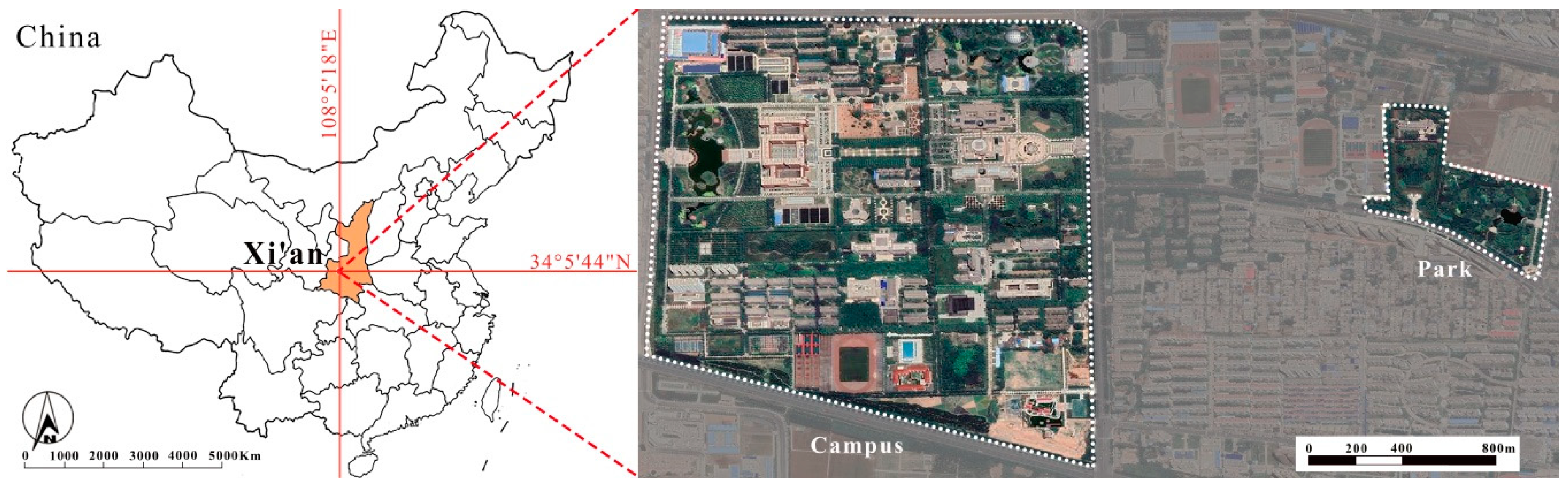
2.1. Study Site and Climatic Conditions
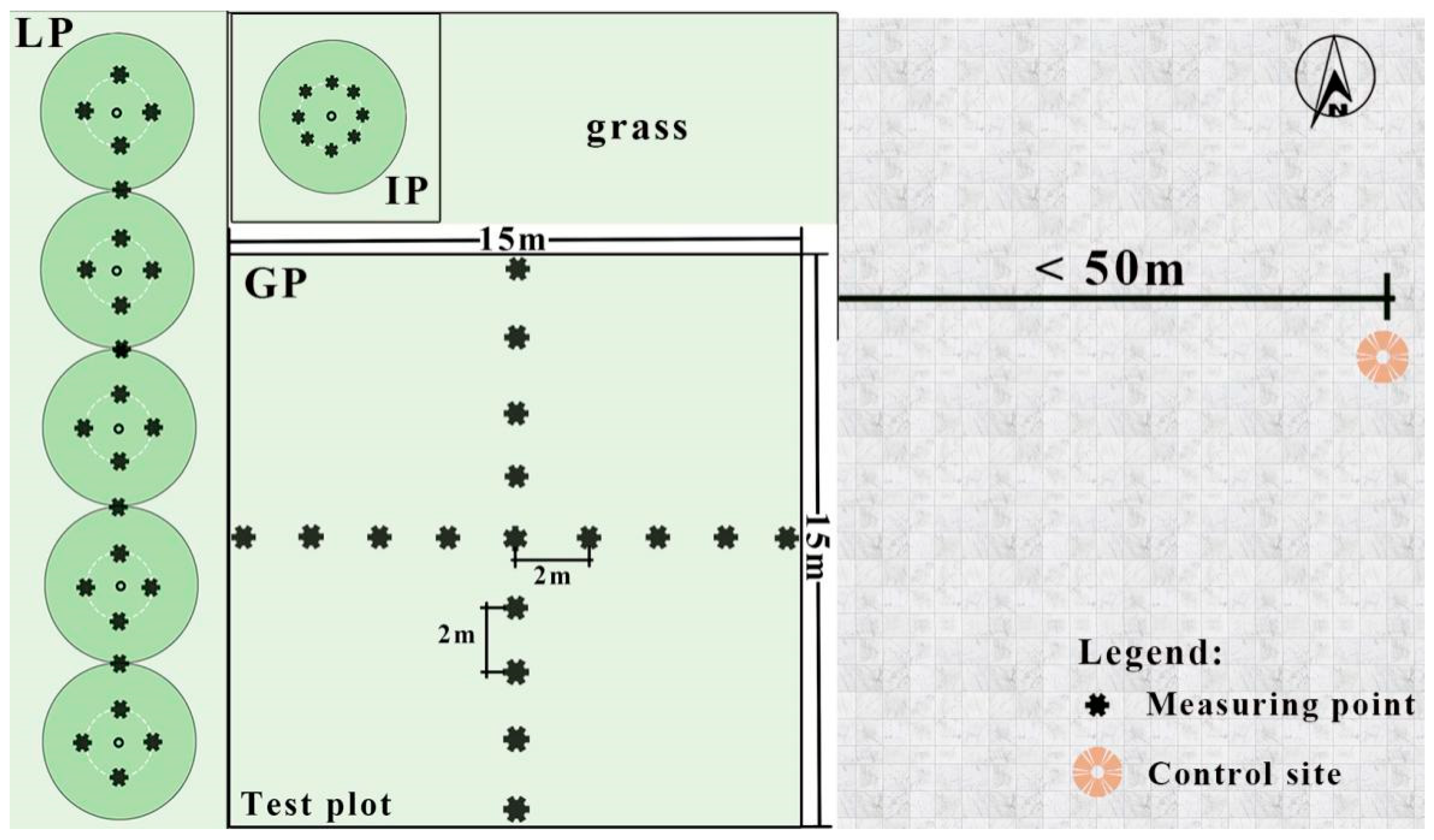
2.2. Experiment Design
2.3. Microclimate Measurements
Measuring Points
2.4. Transpiration Measurements
2.5. Statistical Analyses
3. Results
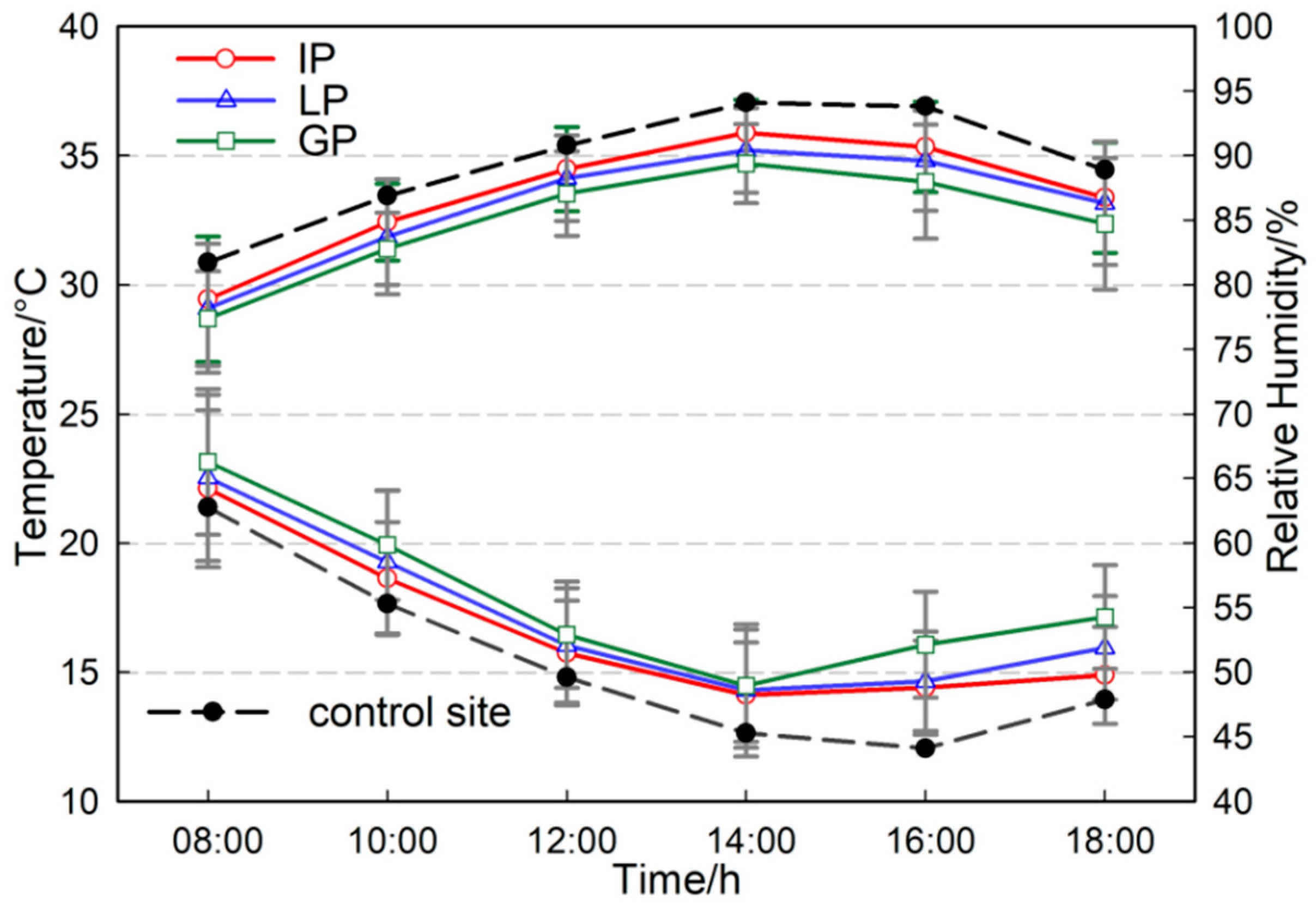
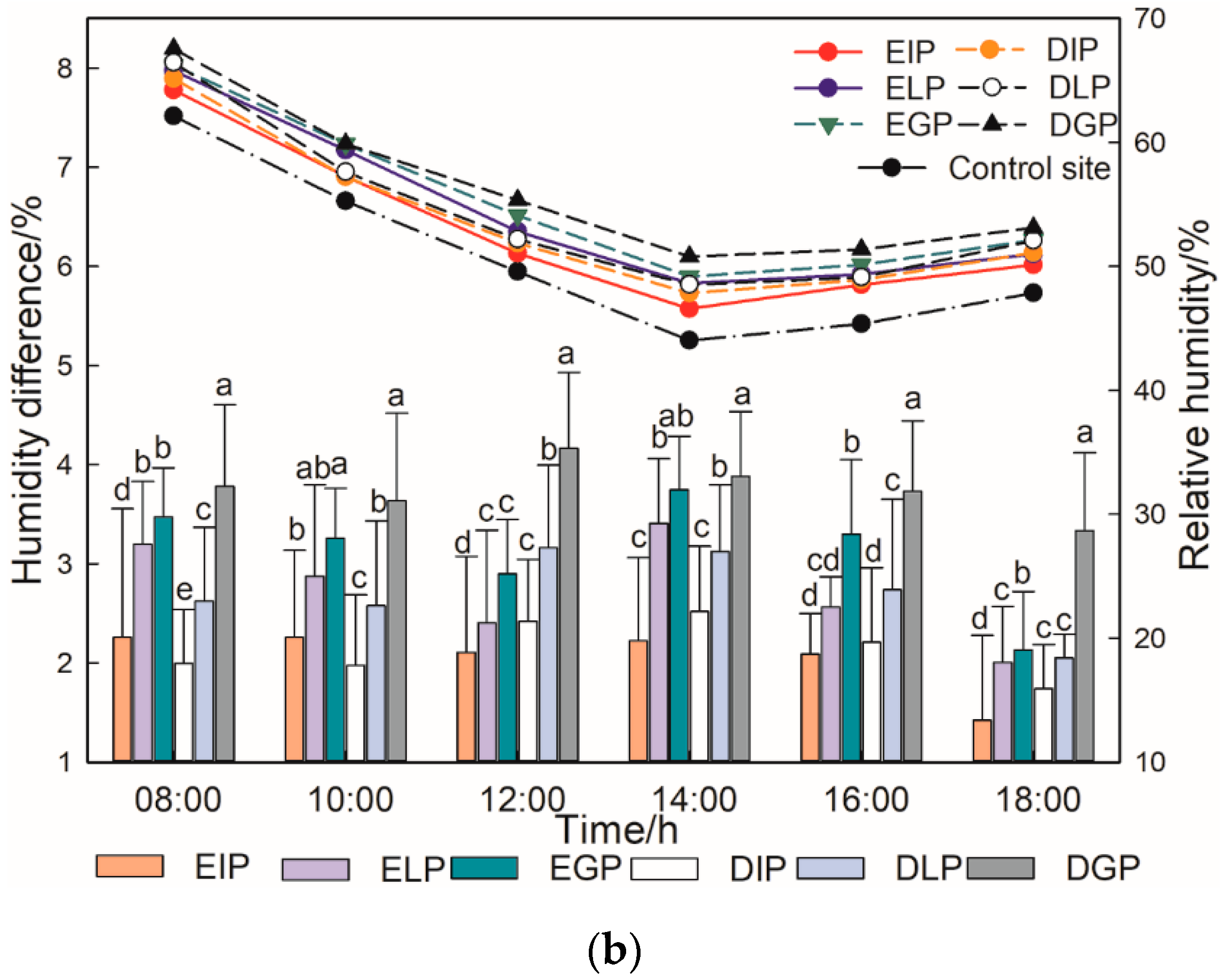
3.1. Diurnal Variations in Air Temperature and Relative Humidity of Urban Trees in Different Configuration Modes
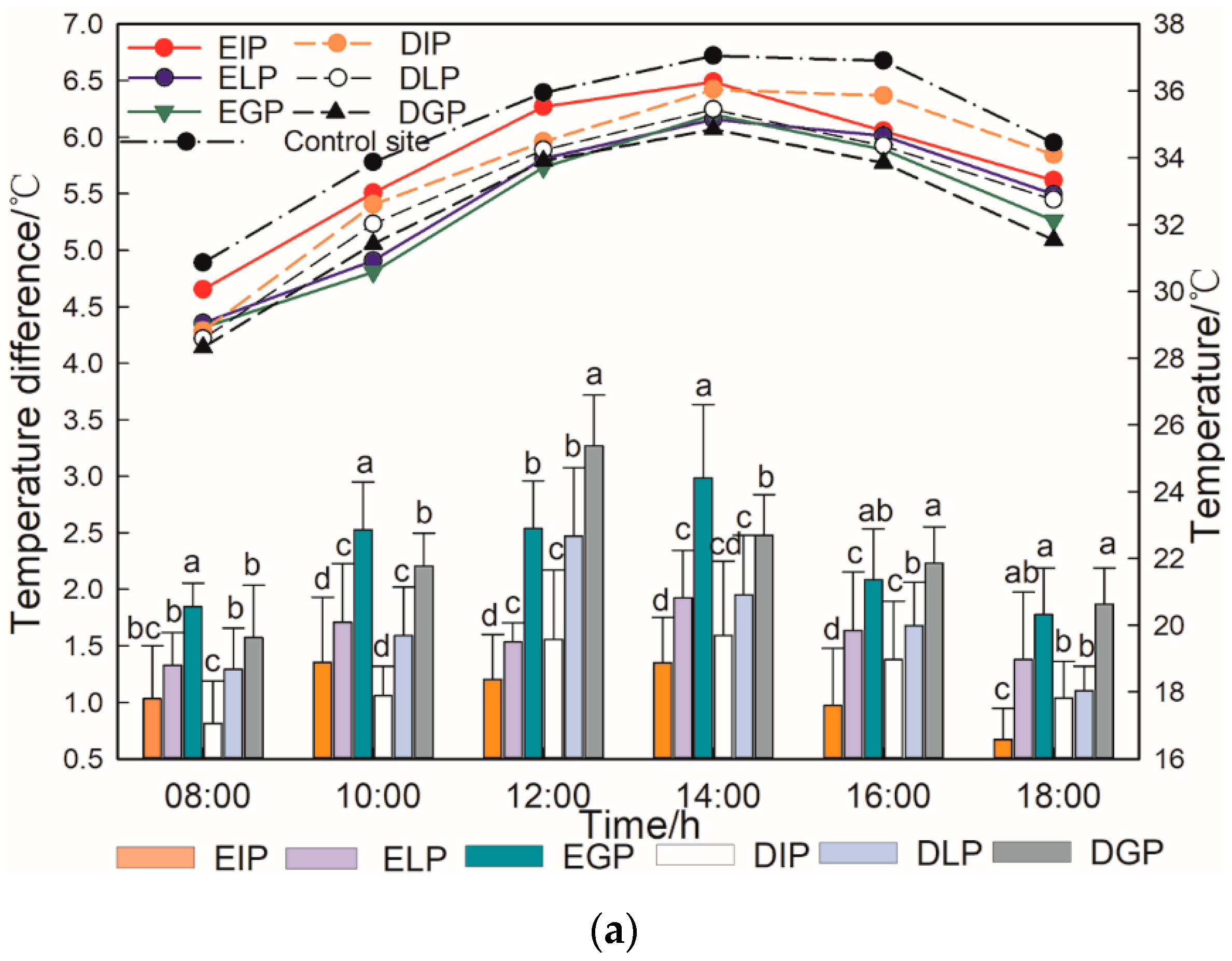
3.2. Microclimate Regulation Capabilities of Urban Trees in Different Types and Different Configuration Modes
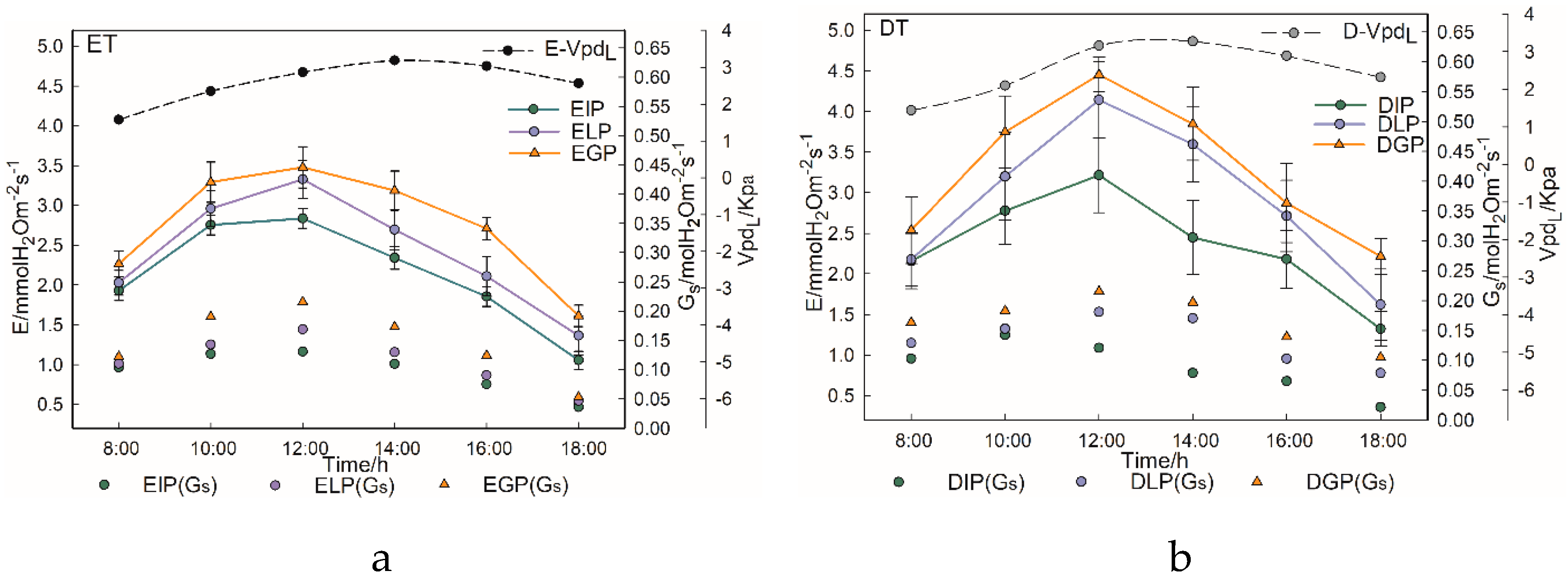
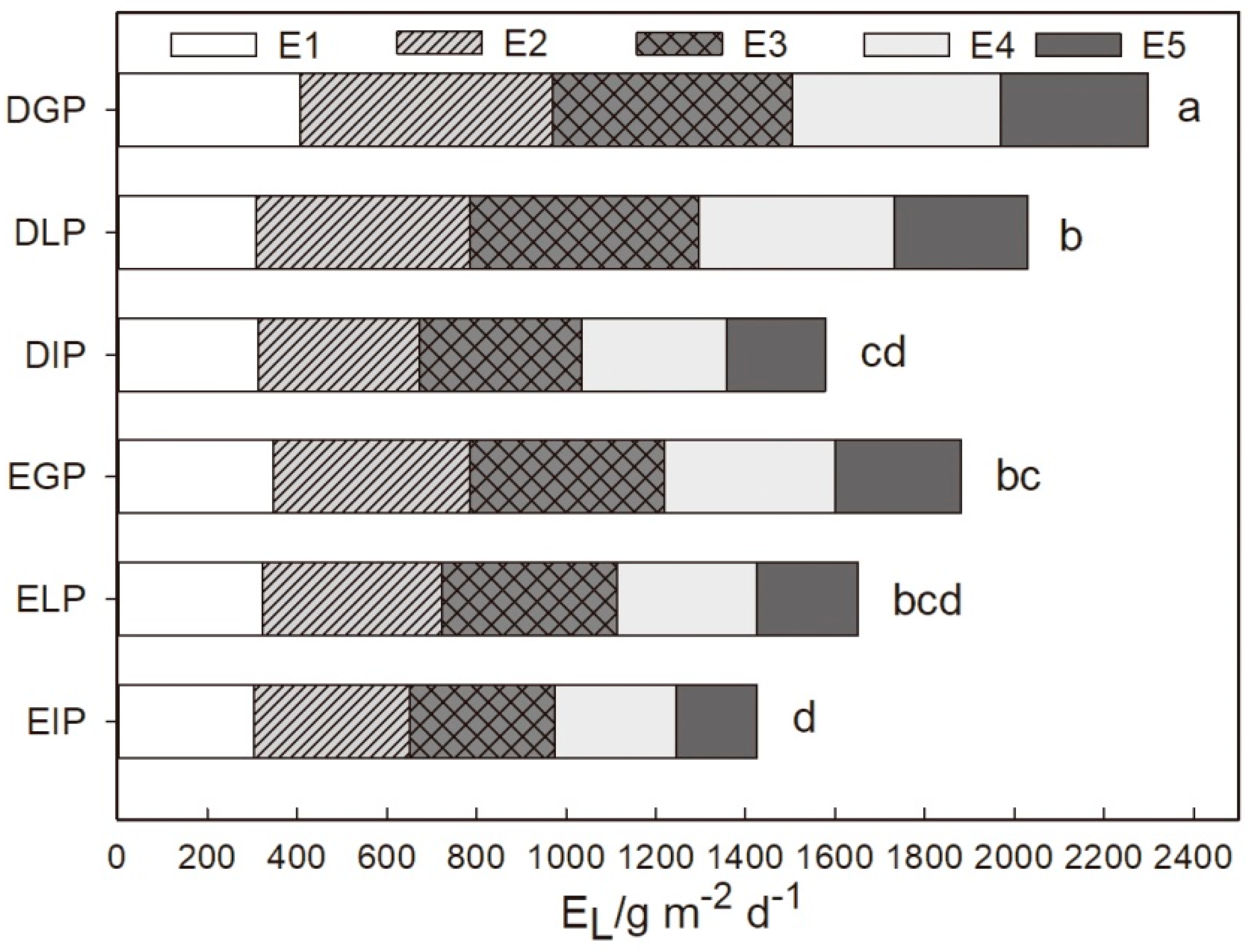
3.3. Transpiration Characteristics of Urban Trees with Different Tree Types and Configuration Modes
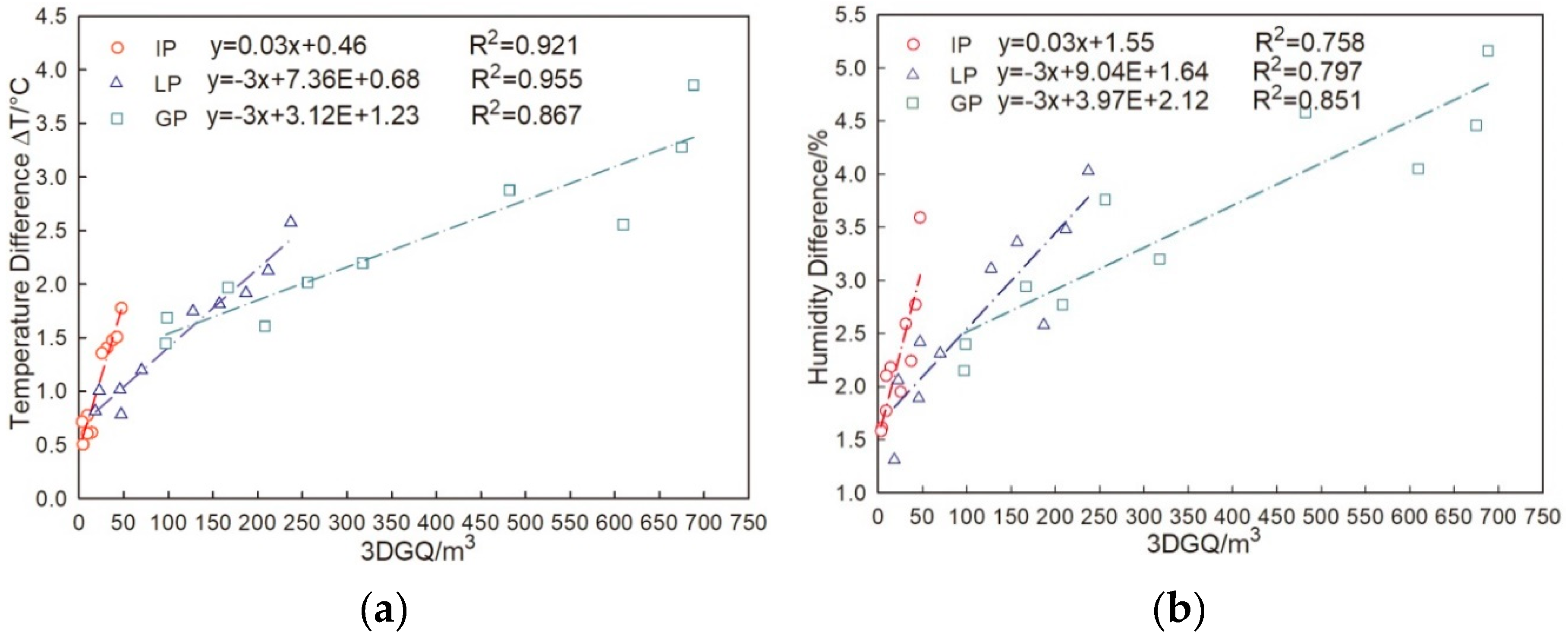
3.4. Correlation between Microclimate Regulation Ability and Influencing Factors of Urban Trees with Different Configuration Modes
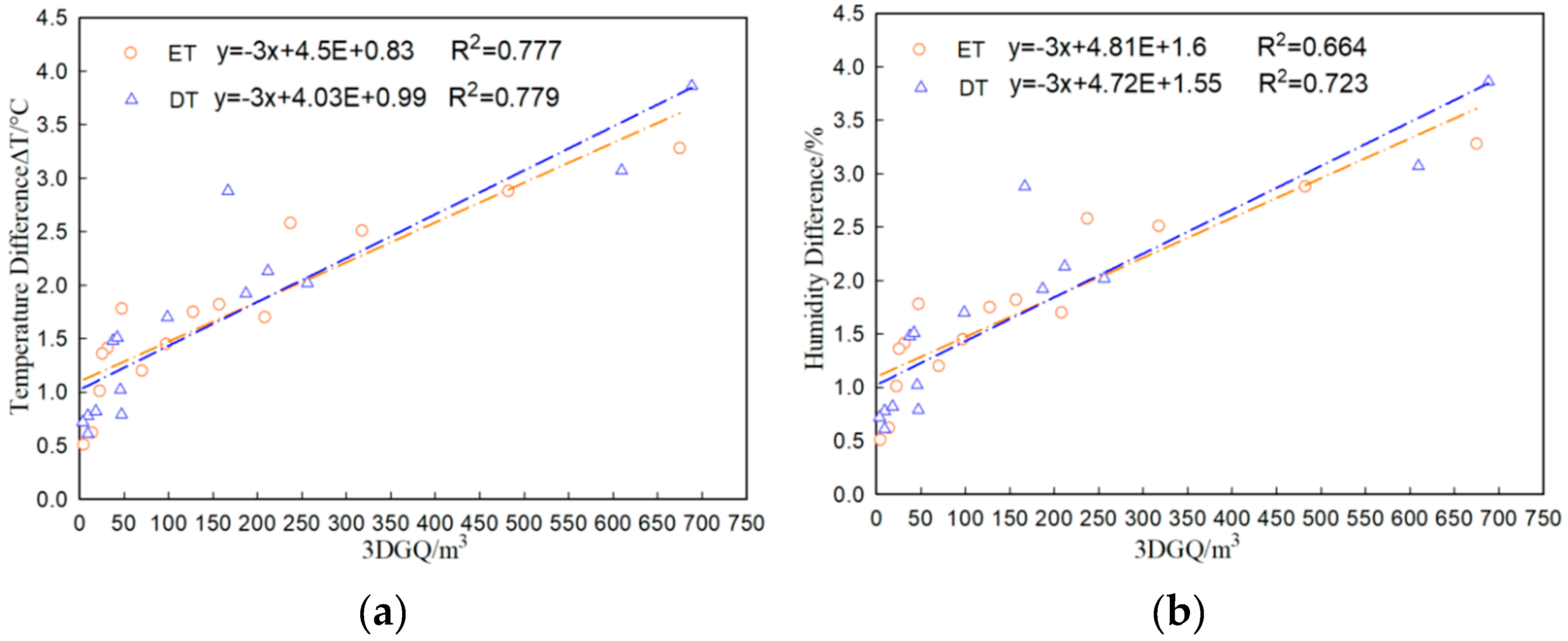
3.5. Correlation between Microclimate Regulation and Influencing Factors of ET and DT
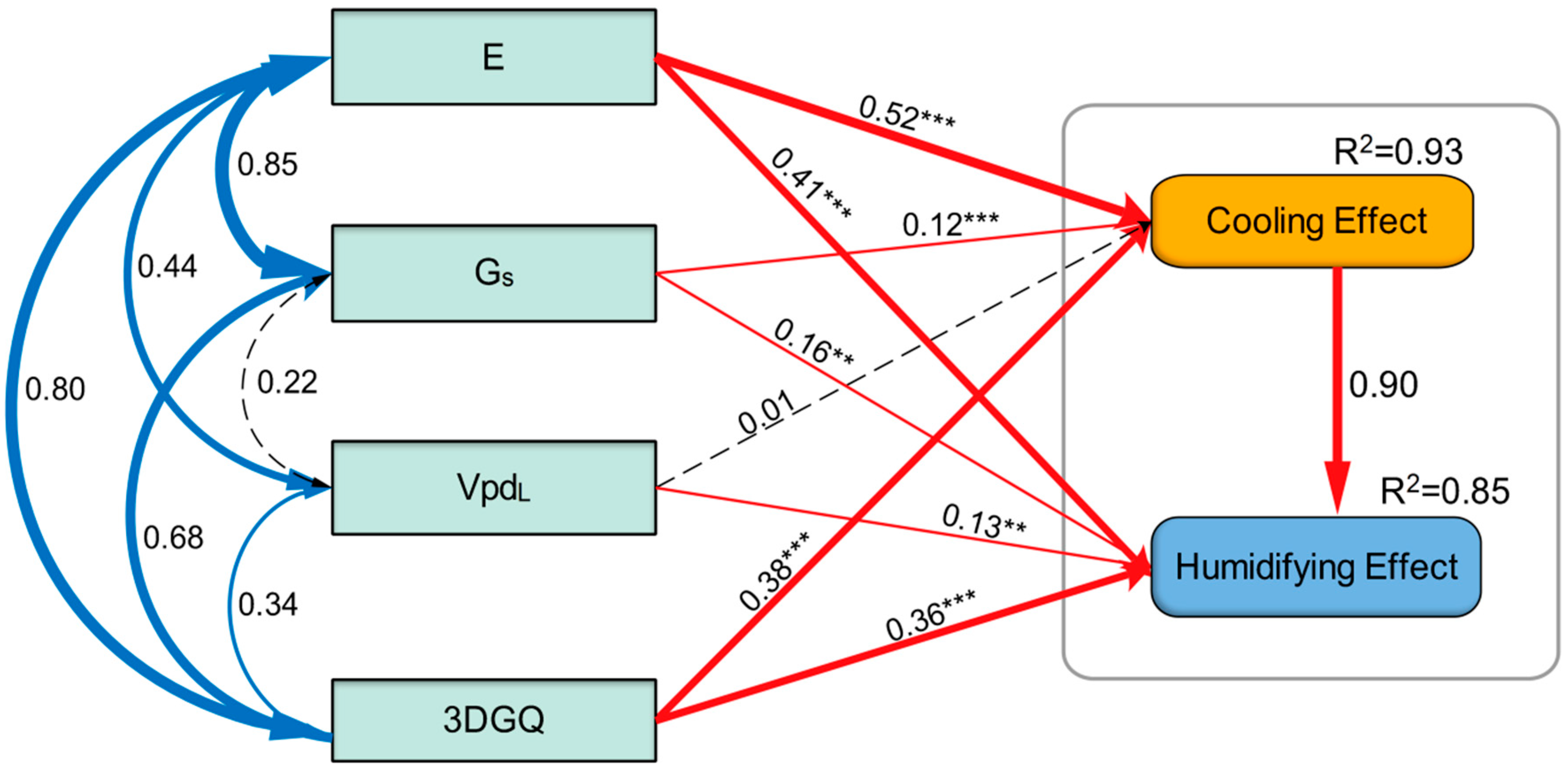
3.6. Structural Equation Model of Microclimate Regulation Ability of Urban Trees
4. Discussion
4.1. Differences of Microclimate Regulation Ability of Urban Trees with Different Tree Species Types and Configuration Modes
4.2. Differences in Transpiration Characteristics of Urban Trees with Different Tree Types and Configuration Modes
4.3. Relationship between Microclimate Effect and Transpiration of Trees
4.4. Relationship between Microclimate Regulation Ability and Tree Structure Factors
4.5. Limitations and Future Research
5. Conclusions
- (1)
- Urban trees with three different configuration modes all have specific improving effects on the surrounding environment, among which GP had the most robust microclimate regulation capability, followed by LP and IP.
- (2)
- DT had better microclimate regulation ability than ET. The microclimate regulation capability of deciduous trees in GP was always more robust than that of LP and IP.
- (3)
- Transpiration characteristics of trees were affected by configuration modes. GP had the best transpiration characteristics, followed by LP and IP.
- (4)
- The microclimate regulation capabilities of urban tree species with different configuration modes were affected by E, Gs, and 3DGQ, and have a weak relationship with VpdL.
- (5)
- The physiological parameters and 3DGQ of urban trees could explain 93% of the cooling effect and 85% of the humidifying effect. The transpiration characteristics and microclimate effects of urban trees depended primarily on the species and configuration mode of the tree. The microclimate regulation capability of urban trees is the result of multiple parameters.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stratópoulos, L.M.F.; Duthweiler, S.; Häberle, K.-H.; Pauleit, S. Effect of native habitat on the cooling ability of six nursery-grown tree species and cultivars for future roadside plantings. Urban For. Urban Green. 2018, 30, 37–45. [Google Scholar] [CrossRef]
- Zhang, Z.; Lv, Y.; Pan, H. Cooling and humidifying effect of plant communities in subtropical urban parks. Urban For. Urban Green. 2013, 12, 323–329. [Google Scholar] [CrossRef]
- Konarska, J.; Lindberg, F.; Larsson, A.; Thorsson, S.; Holmer, B. Transmissivity of solar radiation through crowns of single urban trees—Application for outdoor thermal comfort modelling. Theor. Appl. Climatol. 2013, 117, 363–376. [Google Scholar] [CrossRef]
- Myint, S.W.; Zheng, B.; Talen, E.; Fan, C.; Kaplan, S.; Middel, A.; Smith, M.; Huang, H.-P.; Brazel, A. Does the spatial arrangement of urban landscape matter? examples of urban warming and cooling in phoenix and las vegas. Ecosyst. Health Sustain. 2017, 1, 1–15. [Google Scholar] [CrossRef]
- Fan, C.; Myint, S.W.; Zheng, B. Measuring the spatial arrangement of urban vegetation and its impacts on seasonal surface temperatures. Prog. Phys. Geogr. 2015, 39, 199–219. [Google Scholar] [CrossRef]
- Sodoudi, S.; Zhang, H.; Chi, X.; Müller, F.; Li, H. The influence of spatial configuration of green areas on microclimate and thermal comfort. Urban For. Urban Green. 2018, 34, 85–96. [Google Scholar] [CrossRef]
- Amani-Beni, M.; Zhang, B.; Xie, G.; Xu, J. Impact of urban park’s tree, grass and waterbody on microclimate in hot summer days: A case study of Olympic Park in Beijing, China. Urban For. Urban Green. 2018, 32, 1–6. [Google Scholar] [CrossRef]
- Wang, Y.; Berardi, U.; Akbari, H. Comparing the effects of urban heat island mitigation strategies for Toronto, Canada. Energy Build. 2016, 114, 2–19. [Google Scholar] [CrossRef]
- Hajat, S.; Kosatky, T. Heat-related mortality: A review and exploration of heterogeneity. J. Epidemiol. Community Health 2010, 64, 753–760. [Google Scholar] [CrossRef]
- Battisti, D.S.; Naylor, R.L. Historical Warnings of Future Food Insecurity with Unprecedented Seasonal Heat. Science 2009, 323, 240–244. [Google Scholar] [CrossRef]
- Morakinyo, T.E.; Kong, L.; Lau, K.K.-L.; Yuan, C.; Ng, E. A study on the impact of shadow-cast and tree species on in-canyon and neighborhood’s thermal comfort. Build. Environ. 2017, 115, 1–17. [Google Scholar] [CrossRef]
- Bratman, G.N.; Anderson, C.B.; Berman, M.G.; Cochran, B.; de Vries, S.; Flanders, J.; Folke, C.; Frumkin, H.; Gross, J.J.; Hartig, T.; et al. Nature and mental health: An ecosystem service perspective. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef] [PubMed]
- Duarte, D.H.S.; Shinzato, P.; Gusson, C.d.S.; Alves, C.A. The impact of vegetation on urban microclimate to counterbalance built density in a subtropical changing climate. Urban Clim. 2015, 14, 224–239. [Google Scholar] [CrossRef]
- Zölch, T.; Maderspacher, J.; Wamsler, C.; Pauleit, S. Using green infrastructure for urban climate-proofing: An evaluation of heat mitigation measures at the micro-scale. Urban For. Urban Green. 2016, 20, 305–316. [Google Scholar] [CrossRef]
- El-Bardisy, W.M.; Fahmy, M.; El-Gohary, G.F. Climatic Sensitive Landscape Design: Towards a Better Microclimate through Plantation in Public Schools, Cairo, Egypt. Procedia Soc. Behav. Sci. 2016, 216, 206–216. [Google Scholar] [CrossRef]
- Hami, A.; Abdi, B.; Zarehaghi, D.; Maulan, S.B. Assessing the thermal comfort effects of green spaces: A systematic review of methods, parameters, and plants’ attributes. Sustain. Cities Soc. 2019, 49. [Google Scholar] [CrossRef]
- Zhang, L.; Zhan, Q.; Lan, Y. Effects of the tree distribution and species on outdoor environment conditions in a hot summer and cold winter zone: A case study in Wuhan residential quarters. Build. Environ. 2018, 130, 27–39. [Google Scholar] [CrossRef]
- Wang, Y.; Akbari, H. The effects of street tree planting on Urban Heat Island mitigation in Montreal. Sustain. Cities Soc. 2016, 27, 122–128. [Google Scholar] [CrossRef]
- Connors, J.P.; Galletti, C.S.; Chow, W.T.L. Landscape configuration and urban heat island effects: Assessing the relationship between landscape characteristics and land surface temperature in Phoenix, Arizona. Landsc. Ecol. 2012, 28, 271–283. [Google Scholar] [CrossRef]
- Taha, H. Urban Climates and Heat Islands: Albedo, Evapotranspiration, and Anthropogenic Heat. Energy Build. 1997, 25, 99–103. [Google Scholar] [CrossRef]
- Rahman, M.A.; Moser, A.; Roetzer, T.; Pauleit, S. Within canopy temperature differences and cooling ability of Tilia cordata trees grown in urban conditions. Build. Environ. 2017, 114, 118–128. [Google Scholar] [CrossRef]
- Smithers, R.J.; Doick, K.J.; Burton, A.; Sibille, R.; Steinbach, D.; Harris, R.; Groves, L.; Blicharska, M. Comparing the relative abilities of tree species to cool the urban environment. Urban Ecosyst. 2018, 21, 851–862. [Google Scholar] [CrossRef]
- Jiao, M.; Zhou, W.; Zheng, Z.; Wang, J.; Qian, Y. Patch size of trees affects its cooling effectiveness: A perspective from shading and transpiration processes. Agric. For. Meteorol. 2017, 247, 293–299. [Google Scholar] [CrossRef]
- Ballinas, M.; Barradas, V.L. Transpiration and stomatal conductance as potential mechanisms to mitigate the heat load in Mexico City. Urban For. Urban Green. 2016, 20, 152–159. [Google Scholar] [CrossRef]
- Liang, H.; Li, W.; Zhang, Q.; Zhu, W.; Chen, D.; Liu, J.; Shu, T. Using unmanned aerial vehicle data to assess the three-dimension green quantity of urban green space: A case study in Shanghai, China. Landsc. Urban Plan. 2017, 164, 81–90. [Google Scholar] [CrossRef]
- Cheng, H.; Matteo, C.; Feng, Z.; Zhang, S.; Matteo, C. Using LiDAR Data to Measure the 3D Green Biomass of Beijing Urban Forest in China. PLoS ONE 2013, 8, e75920. [Google Scholar]
- Qiu, G.Y.; Zou, Z.; Li, X.; Li, H.; Guo, Q.; Yan, C.; Tan, S. Experimental studies on the effects of green space and evapotranspiration on urban heat island in a subtropical megacity in China. Habitat Int. 2017, 68, 30–42. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, J.; Cadenasso, M.L. Effects of the spatial configuration of trees on urban heat mitigation: A comparative study. Remote Sens. Environ. 2017, 195, 1–12. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Z.-H.; Wang, C.; Myint, S.W. Environmental cooling provided by urban trees under extreme heat and cold waves in U.S. cities. Remote Sens. Environ. 2019, 227, 28–43. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Yu, Z.; Xu, S.; Zhang, Y.; Jorgensen, G.; Vejre, H. Strong contributions of local background climate to the cooling effect of urban green vegetation. Sci. Rep. 2018, 8, 6798. [Google Scholar] [CrossRef] [PubMed]
- Gaylon, S.; Campbell, J.M.N. An Introduction to Environmental Biophysics; Springer: New York, NY, USA, 1998; pp. 75–80. [Google Scholar] [CrossRef]
- Dimoudi, A.; Nikolopoulou, M. Vegetation in the urban environment: Microclimatic analysis and benefits. Energy Build. 2003, 35, 69–76. [Google Scholar] [CrossRef]
- Zhao, Q.; Sailor, D.J.; Wentz, E.A. Impact of tree locations and arrangements on outdoor microclimates and human thermal comfort in an urban residential environment. Urban For. Urban Green. 2018, 32, 81–91. [Google Scholar] [CrossRef]
- Shashua-Bar, L.; Tsiros, I.X.; Hoffman, M.E. A modeling study for evaluating passive cooling scenarios in urban streets with trees. Case study: Athens, Greece. Build. Environ. 2010, 45, 2798–2807. [Google Scholar] [CrossRef]
- Hsieh, C.-M.; Li, J.-J.; Zhang, L.; Schwegler, B. Effects of tree shading and transpiration on building cooling energy use. Energy Build. 2018, 159, 382–397. [Google Scholar] [CrossRef]
- Tobias, S.; Angela, H.; Thomas, S. Cooling Effects and Regulating Ecosystem Services Provided by Urban Trees—Novel Analysis Approaches Using Urban Tree Cadastre Data. Sustainability 2018, 10, 712. [Google Scholar]
- Wang, Y.; Bakker, F.; de Groot, R.; Wortche, H.; Leemans, R. Effects of urban green infrastructure (UGI) on local outdoor microclimate during the growing season. Environ. Monit. Assess. 2015, 187, 732. [Google Scholar] [CrossRef]
- Dronova, I.; Friedman, M.; McRae, I.; Kong, F.; Yin, H. Spatio-temporal non-uniformity of urban park greenness and thermal characteristics in a semi-arid region. Urban For. Urban Green. 2018, 34, 44–54. [Google Scholar] [CrossRef]
- Hamada, S.; Ohta, T. Seasonal variations in the cooling effect of urban green areas on surrounding urban areas. Urban For. Urban Green. 2010, 9, 15–24. [Google Scholar] [CrossRef]
- Wu, Z.; Chen, L. Optimizing the spatial arrangement of trees in residential neighborhoods for better cooling effects: Integrating modeling with in-situ measurements. Landsc. Urban Plan. 2017, 167, 463–472. [Google Scholar] [CrossRef]
- Rahman, M.A.; Stratopoulos, L.M.F.; Moser-Reischl, A.; Zölch, T.; Häberle, K.-H.; Rötzer, T.; Pretzsch, H.; Pauleit, S. Traits of trees for cooling urban heat islands: A meta-analysis. Build. Environ. 2020, 170. [Google Scholar] [CrossRef]
- Lin, B.S.; Lin, Y.J. Cooling effect of shade trees with different characteristics in a subtropical urban park. Hortscience 2010, 45, 83–86. [Google Scholar] [CrossRef]
- Shashua-Bar, L.; Pearlmutter, D.; Erell, E. The influence of trees and grass on outdoor thermal comfort in a hot-arid environment. Int. J. Climatol. 2011, 31, 1498–1506. [Google Scholar] [CrossRef]
- Manickathan, L.; Defraeye, T.; Allegrini, J.; Derome, D.; Carmeliet, J. Parametric study of the influence of environmental factors and tree properties on the transpirative cooling effect of trees. Agric. For. Meteorol. 2018, 248, 259–274. [Google Scholar] [CrossRef]
- Bowden, J.D.; Bauerle, W.L. Measuring and modeling the variation in species-specific transpiration in temperate deciduous hardwoods. Tree Physiol. 2008, 28, 1675–1683. [Google Scholar] [CrossRef]
- Rahman, M.A.; Hartmann, C.; Moser-Reischl, A.; von Strachwitz, M.F.; Paeth, H.; Pretzsch, H.; Pauleit, S.; Rötzer, T. Tree cooling effects and human thermal comfort under contrasting species and sites. Agric. For. Meteorol. 2020, 287. [Google Scholar] [CrossRef]
- Moss, J.L.; Doick, K.J.; Smith, S.; Shahrestani, M. Influence of evaporative cooling by urban forests on cooling demand in cities. Urban For. Urban Green. 2019, 37, 65–73. [Google Scholar] [CrossRef]
- Rahman, M.A.; Moser, A.; Rötzer, T.; Pauleit, S. Comparing the transpirational and shading effects of two contrasting urban tree species. Urban Ecosyst. 2019, 22, 683–697. [Google Scholar] [CrossRef]
- JL, H. Stomata of trees growing in CO2-enriched air show reduced sensitivity to vapour pressure deficit and drought. Plant Cell Environ. 1998, 21, 1077–1088. [Google Scholar]
- Peters, E.B.; McFadden, J.P.; Montgomery, R.A. Biological and environmental controls on tree transpiration in a suburban landscape. J. Geophys. Res. 2010, 115. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, P.; Hu, Y.; Ouyang, L.; Zhu, L.; Ni, G. Canopy transpiration and its cooling effect of three urban tree species in a subtropical city—Guangzhou, China. Urban For. Urban Green. 2019, 43. [Google Scholar] [CrossRef]
- Moser-Reischl, A.; Rahman, M.A.; Pauleit, S.; Pretzsch, H.; Rötzer, T. Growth patterns and effects of urban micro-climate on two physiologically contrasting urban tree species. Landsc. Urban Plan. 2019, 183, 88–99. [Google Scholar] [CrossRef]
- Lindén, J.; Fonti, P.; Esper, J. Temporal variations in microclimate cooling induced by urban trees in Mainz, Germany. Urban For. Urban Green. 2016, 20, 198–209. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Sun, X.; Berlyn, G.P.; Rehim, A. Effect of pavement and water deficit on biomass allocation and whole-tree transpiration in two contrasting urban tree species. Urban Ecosyst. 2020. [Google Scholar] [CrossRef]
- Eamus, D.; Boulain, N.; Cleverly, J.; Breshears, D.D. Global change-type drought-induced tree mortality: Vapor pressure deficit is more important than temperature per se in causing decline in tree health. Ecol. Evol. 2013, 3, 2711–2729. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Hwang, S.J.; Lee, S.B.; Hwang, H.S.; Sung, H.C. Landscape ecological approach to the relationships of land use patterns in watersheds to water quality characteristics. Landsc. Urban Plan. 2009, 92, 80–89. [Google Scholar] [CrossRef]
- Morakinyo, T.E.; Lam, Y.F. Simulation study on the impact of tree-configuration, planting pattern and wind condition on street-canyon’s micro-climate and thermal comfort. Build. Environ. 2016, 103, 262–275. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Functional Type | Latin | Mean Height (m) | X (m) | DBH (cm) | Y (m) | Three Dimensional Quantity Expressions |
|---|---|---|---|---|---|---|
| Deciduous broadleaf Trees | Cerasus serrulata var. lannesiana | 5 | 4.6 | 20 | 3.6 | πX2Y/6 |
| Malus Halliana | 3.3 | 2.8 | 7.5 | 2.23 | πX2Y/6 | |
| Acer palmatum | 2.6 | 2.5 | 6.3 | 1.4 | π(2Y3−Y2)/3 | |
| Cercis chinensis | 4.1 | 4.8 | — | 3.1 | πX2Y/6 | |
| Hibiscus syriacus | 3.2 | 3 | 10 | 2 | πX2Y/6 | |
| Evergreen broadleaf Trees | Ligustrum lucidum | 7.3 | 4.7 | 24.8 | 4.1 | πX2Y/6 |
| Eriobotrya japonica | 4.9 | 4.2 | 19.3 | 3.4 | πX2Y/6 | |
| Viburnum odoratissinum | 4.1 | 2.2 | — | 3.7 | πX2Y/4 | |
| Photinia × fraseri | 1.6 | 2.4 | — | 1.5 | πX2Y/6 | |
| Michelia maudiae | 8.2 | 4.1 | 13 | 5.8 | πX2Y/12 |
| Variables | Equipment | Measurement Range | Accuracy |
|---|---|---|---|
| Air Temperature | PH-II hand-held weather station | −50–80 °C | ± 0.3 °C |
| Relative humidity | 0–100% | ± 5%RH | |
| Wind Speed | 0–45 | ± (0.3 + 0.03V) m/s | |
| Transpiration rate | LI-6400XT portable photosynthesis measurement system | _ | Maximum error of H2O analvzer: ± 1.0 mmol/mol |
| Stomata conductance | |||
| Tree height/Canopy crown | Rxiry laser tree altimeter | 0–820 m | ± 0.5 m |
| TD | E | Gs | VpdL | 3DGQ | Ta | RH | ||
|---|---|---|---|---|---|---|---|---|
| IP | TD | 1 | 0.910 ** | 0.769 ** | 0.330 * | 0.903 ** | 0.136 | 0.048 |
| HD | 0.855 ** | 0.837 ** | 0.690 ** | 0.352 ** | 0.939 ** | 0.352 * | −0.127 | |
| LP | TD | 1 | 0.928 ** | 0.840 ** | 0.386 ** | 0.915 ** | 0.149 | −0.061 |
| HD | 0.895 ** | 0.943 ** | 0.850 ** | 0.491 ** | 0.879 ** | 0.204 | −0.117 | |
| GP | TD | 1 | 0.910 ** | 0.840 ** | 0.379 ** | 0.896 ** | 0.255 * | −0.005 |
| HD | 0.875 ** | 0.837 ** | 0.779 ** | 0.514 ** | 0.903 ** | 0.319 * | −0.054 |
| TD | E | Gs | VpdL | 3DGQ | ||
|---|---|---|---|---|---|---|
| ET | TD | 1 | 0.892 ** | 0.770 ** | 0.309 ** | 0.929 ** |
| HD | 0.899 ** | 0.854 ** | 0.757 ** | 0.444 ** | 0.843 ** | |
| DT | TD | 1 | 0.908 ** | 0.819 ** | 0.426 ** | 0.925 ** |
| HD | 0.899 ** | 0.884 ** | 0.753 ** | 0.443 ** | 0.914 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, D.; Lei, Q.; Shi, Y.; Wang, M.; Chen, S.; Shah, K.; Ji, W. Role of Species and Planting Configuration on Transpiration and Microclimate for Urban Trees. Forests 2020, 11, 825. https://doi.org/10.3390/f11080825
Zhao D, Lei Q, Shi Y, Wang M, Chen S, Shah K, Ji W. Role of Species and Planting Configuration on Transpiration and Microclimate for Urban Trees. Forests. 2020; 11(8):825. https://doi.org/10.3390/f11080825
Chicago/Turabian StyleZhao, Dan, Quanhuan Lei, Yajie Shi, Mengdi Wang, Sibo Chen, Kamran Shah, and Wenli Ji. 2020. "Role of Species and Planting Configuration on Transpiration and Microclimate for Urban Trees" Forests 11, no. 8: 825. https://doi.org/10.3390/f11080825
APA StyleZhao, D., Lei, Q., Shi, Y., Wang, M., Chen, S., Shah, K., & Ji, W. (2020). Role of Species and Planting Configuration on Transpiration and Microclimate for Urban Trees. Forests, 11(8), 825. https://doi.org/10.3390/f11080825