Quantification of Lichen Cover and Biomass Using Field Data, Airborne Laser Scanning and High Spatial Resolution Optical Data—A Case Study from a Canadian Boreal Pine Forest



Abstract
1. Introduction
2. Materials and Methods
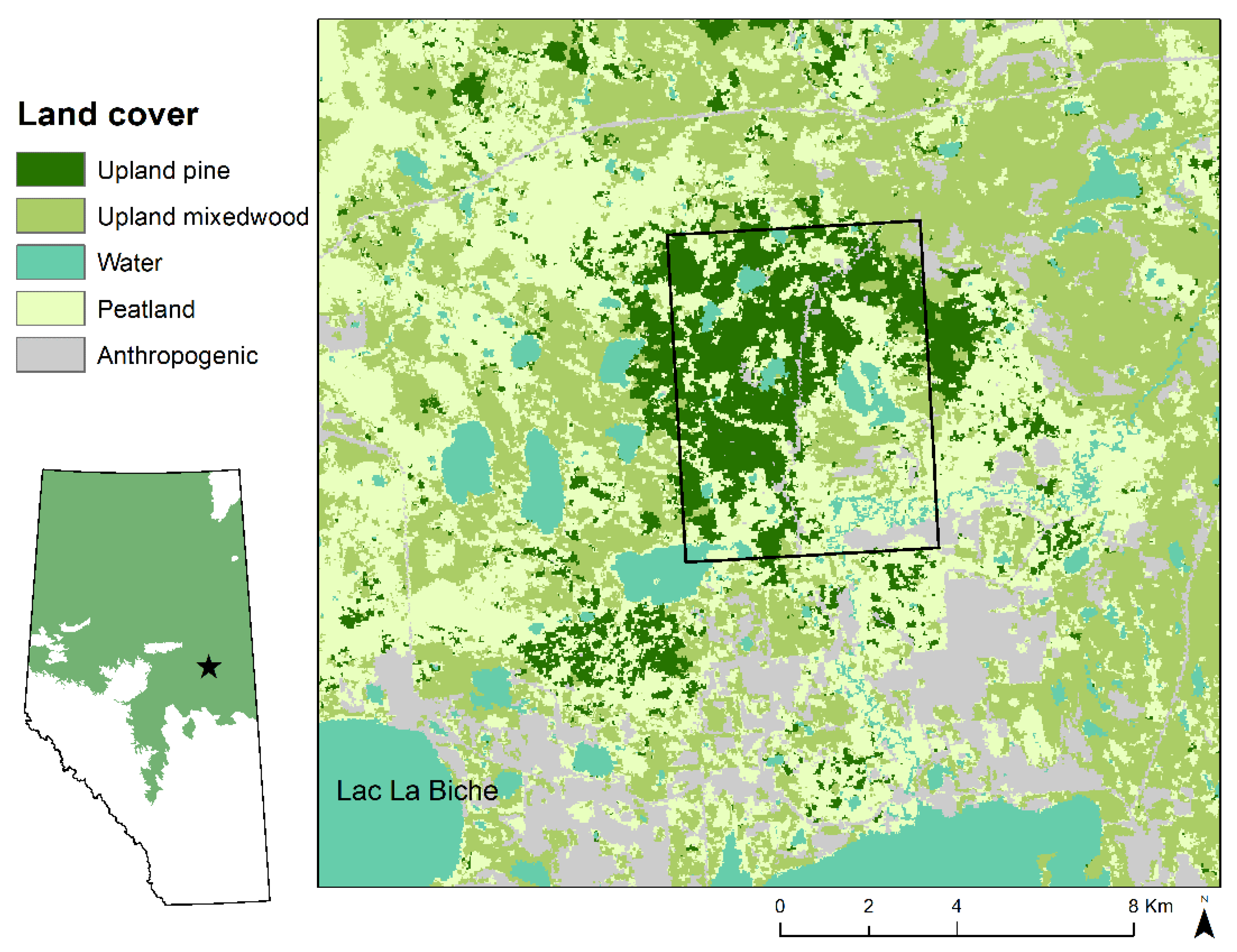
2.1. Study Area
2.2. Remote Sensing Predictor Variables
2.3. Sample Plot Selection
2.4. Field Measurements of Cover and Biomass
2.5. Allometric Model of Lichen Cover and Biomass
2.6. Statistical Modelling
3. Results
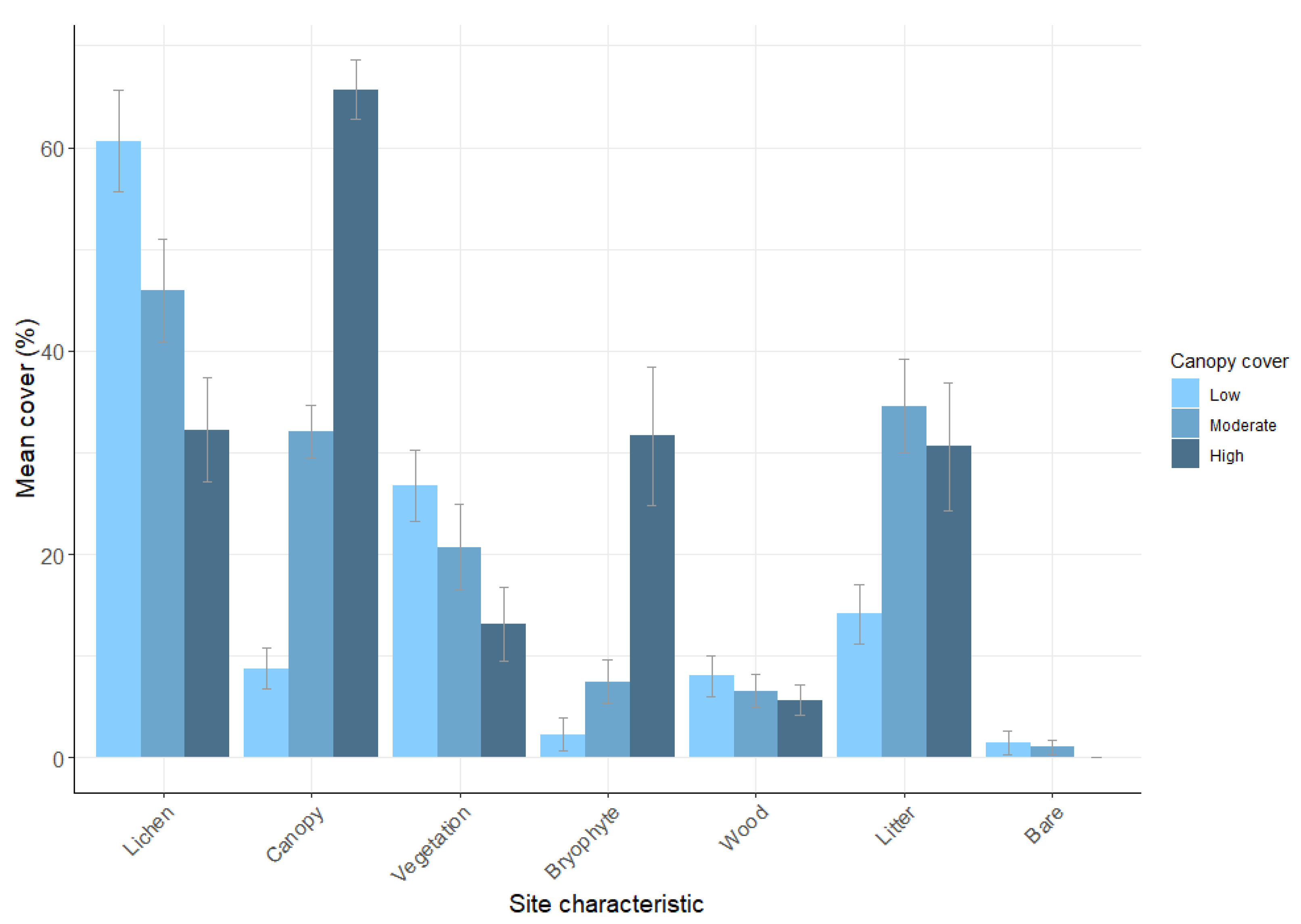
3.1. Plot Characteristics
3.2. Model Selection
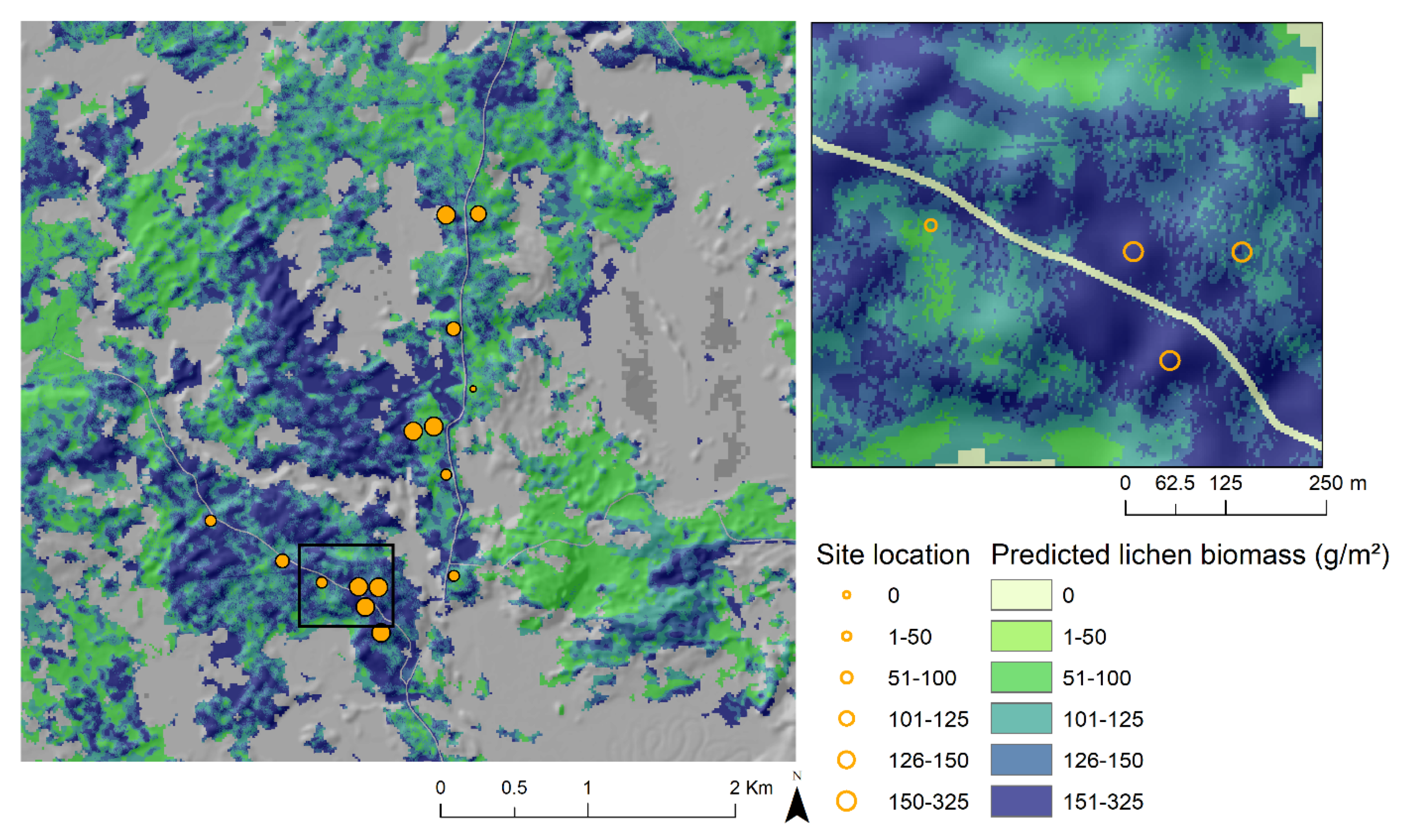
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | Quadrat | Easting | Northing | Lichen Cover (%) | Lichen Biomass (g/m2) | ALS-Derived Canopy Cover (%) | Band 3 (Blue) |
|---|---|---|---|---|---|---|---|
| L1 | L1-C | 435310.41 | 6093434.95 | 84 | 258.03 | 7 | 1853 |
| L1 | L1-NE | 435319.23 | 6093443.17 | 2 | 6.14 | 7 | 2044 |
| L1 | L1-SE | 435319.28 | 6093424.69 | 74 | 227.31 | 7 | 2405 |
| L1 | L1-SW | 435300.41 | 6093425.63 | 62 | 190.45 | 25 | 1977 |
| L1 | L1-NW | 435302.57 | 6093446.64 | 69 | 211.95 | 18 | 1610 |
| L2 | L2-C | 435272.78 | 6093572.72 | 55 | 168.95 | 15 | 1926 |
| L2 | L2-NE | 435280.45 | 6093581.18 | 80 | 245.74 | 15 | 2058 |
| L2 | L2-SE | 435281.30 | 6093560.35 | 87 | 267.25 | 32 | 2019 |
| L2 | L2-SW | 435260.19 | 6093560.88 | 30 | 92.15 | 32 | 2189 |
| L2 | L2-NW | 435263.24 | 6093581.09 | 92 | 282.61 | 15 | 1953 |
| L3 | L3-C | 436011.89 | 6096061.35 | 45 | 138.23 | 39 | 1810 |
| L3 | L3-NE | 436023.26 | 6096068.65 | 52 | 159.73 | 39 | 1848 |
| L3 | L3-SE | 436021.27 | 6096050.76 | 36 | 110.58 | 39 | 1604 |
| L3 | L3-SW | 436000.76 | 6096052.05 | 80 | 245.74 | 39 | 1881 |
| L3 | L3-NW | 436002.63 | 6096070.84 | 34 | 104.44 | 39 | 1511 |
| L4 | L4-C | 435702.91 | 6094608.81 | 87 | 267.25 | 37 | 1527 |
| L4 | L4-NE | 435713.12 | 6094620.01 | 35 | 107.51 | 64 | 2099 |
| L4 | L4-SE | 435714.29 | 6094598.74 | 54 | 165.88 | 64 | 2630 |
| L4 | L4-SW | 435694.39 | 6094603.25 | 25 | 76.80 | 37 | 1446 |
| L4 | L4-NW | 435695.17 | 6094622.39 | 95 | 291.82 | 37 | 1986 |
| L5 | L5-C | 435407.70 | 6093253.91 | 98 | 301.04 | 6 | 2266 |
| L5 | L5-NE | 435416.70 | 6093265.36 | 52 | 159.73 | 24 | 1944 |
| L5 | L5-SE | 435417.40 | 6093247.32 | 46 | 141.30 | 6 | 2303 |
| L5 | L5-SW | 435396.90 | 6093245.72 | 72 | 221.17 | 6 | 2333 |
| L5 | L5-NW | 435397.91 | 6093267.91 | 70 | 215.03 | 24 | 1674 |
| M1 | M1-C | 435407.39 | 6093564.43 | 75 | 230.39 | 53 | 1639 |
| M1 | M1-NE | 435418.35 | 6093574.51 | 82 | 251.89 | 53 | 1537 |
| M1 | M1-SE | 435417.98 | 6093553.59 | 57 | 175.09 | 55 | 1451 |
| M1 | M1-SW | 435397.47 | 6093556.23 | 90 | 276.46 | 56 | 1465 |
| M1 | M1-NW | 435398.64 | 6093575.02 | 72 | 221.17 | 47 | 1644 |
| M2 | M2-C | 434766.11 | 6093777.52 | 36 | 110.58 | 28 | 2158 |
| M2 | M2-NE | 434772.69 | 6093790.00 | 47 | 144.37 | 46 | 1190 |
| M2 | M2-SE | 434773.59 | 6093768.06 | 45 | 138.23 | 28 | 1300 |
| M2 | M2-SW | 434755.49 | 6093773.33 | 20 | 61.44 | 43 | 1755 |
| M2 | M2-NW | 434757.06 | 6093788.89 | 52 | 159.73 | 13 | 1357 |
| M3 | M3-C | 434295.61 | 6094080.33 | 4 | 12.29 | 48 | 1292 |
| M3 | M3-NE | 434307.67 | 6094090.84 | 10 | 30.72 | 48 | 1491 |
| M3 | M3-SE | 434304.34 | 6094073.20 | 41 | 125.94 | 53 | 2176 |
| M3 | M3-SW | 434285.74 | 6094074.92 | 55 | 168.95 | 55 | 1233 |
| M3 | M3-NW | 434286.62 | 6094091.37 | 40 | 122.87 | 45 | 2151 |
| M4 | M4-C | 436015.54 | 6095286.12 | 42 | 129.02 | 59 | 1134 |
| M4 | M4-NE | 436024.67 | 6095291.11 | 7 | 21.50 | 53 | 1403 |
| M4 | M4-SE | 436024.32 | 6095271.09 | 32 | 98.30 | 53 | 1374 |
| M4 | M4-SW | 436002.78 | 6095271.95 | 32 | 98.30 | 59 | 1567 |
| M4 | M4-NW | 436003.72 | 6095293.42 | 75 | 230.39 | 59 | 1822 |
| M5 | M5-C | 436230.31 | 6096055.12 | 87 | 267.25 | 54 | 1261 |
| M5 | M5-NE | 436239.56 | 6096066.23 | 55 | 168.95 | 54 | 1369 |
| M5 | M5-SE | 436239.25 | 6096049.20 | 55 | 168.95 | 54 | 1595 |
| M5 | M5-SW | 436218.95 | 6096047.49 | 30 | 92.15 | 54 | 1053 |
| M5 | M5-NW | 436220.02 | 6096064.28 | 6 | 18.43 | 54 | 1041 |
| H1 | H1-C | 435023.08 | 6093619.85 | 30 | 92.15 | 58 | 1207 |
| H1 | H1-NE | 435032.45 | 6093630.07 | 17 | 52.22 | 58 | 1148 |
| H1 | H1-SE | 435032.31 | 6093607.37 | 50 | 153.59 | 60 | 1125 |
| H1 | H1-SW | 435013.39 | 6093604.19 | 42 | 129.02 | 77 | 1196 |
| H1 | H1-NW | 435014.79 | 6093630.44 | 7 | 21.50 | 67 | 1332 |
| H2 | H2-C | 435922.83 | 6093611.45 | 0 | 0.00 | 60 | 1163 |
| H2 | H2-NE | 435932.76 | 6093616.31 | 17 | 52.22 | 60 | 970 |
| H2 | H2-SE | 435932.05 | 6093598.18 | 8 | 24.57 | 60 | 1359 |
| H2 | H2-SW | 435911.61 | 6093601.26 | 0 | 0.00 | 67 | 1212 |
| H2 | H2-NW | 435912.07 | 6093619.95 | 50 | 153.59 | 67 | 1249 |
| H3 | H3-C | 435911.01 | 6094300.87 | 22 | 67.58 | 74 | 1199 |
| H3 | H3-NE | 435922.42 | 6094306.72 | 17 | 52.22 | 74 | 1262 |
| H3 | H3-SE | 435920.04 | 6094288.17 | 8 | 24.57 | 74 | 1058 |
| H3 | H3-SW | 435899.31 | 6094288.35 | 17 | 52.22 | 57 | 1296 |
| H3 | H3-NW | 435900.35 | 6094307.48 | 67 | 205.81 | 57 | 1203 |
| H4 | H4-C | 436125.92 | 6094872.09 | 27 | 82.94 | 76 | 928 |
| H4 | H4-NE | 436135.71 | 6094881.41 | 22 | 67.58 | 76 | 1016 |
| H4 | H4-SE | 436138.37 | 6094861.67 | 2 | 6.14 | 82 | 1132 |
| H4 | H4-SW | 436117.24 | 6094859.97 | 0 | 0.00 | 82 | 1037 |
| H4 | H5-NW | 436118.28 | 6094879.66 | 24 | 73.72 | 76 | 986 |
| H5 | H5-C | 435844.78 | 6094631.70 | 80 | 245.74 | 68 | 1130 |
| H5 | H5-NE | 435856.22 | 6094639.66 | 42 | 129.02 | 68 | 1145 |
| H5 | H5-SE | 435855.30 | 6094624.65 | 60 | 184.31 | 68 | 1059 |
| H5 | H5-SW | 435838.90 | 6094622.99 | 45 | 138.23 | 68 | 1300 |
| H5 | H5-NW | 435835.54 | 6094638.73 | 87 | 267.25 | 68 | 1001 |
References
- Vitt, D.H.; Finnegan, L.; House, M. Terrestrial bryophyte and lichen responses to canopy opening in pine-moss-lichen forests. Forests 2019, 10, 233. [Google Scholar] [CrossRef]
- Dunford, J.S.; McLoughlin, P.D.; Dalerum, F.; Boutin, S. Lichen abundance in the peatlands of northern Alberta: Implications for boreal caribou. Ecoscience 2007, 13, 469–474. [Google Scholar] [CrossRef]
- Hale, M. The Biology of Lichens, 2nd ed.; Edward Arnold Publishers Ltd.: London, UK, 1974. [Google Scholar]
- Environment Canada. National Recovery Strategy for Woodland Caribou (Rangifer tarandus caribou), Boreal Population, in Canada; Environment Canada: Ottawa, ON, Canada, 2012; ISBN 9781100207698.
- COSEWIC. Assessment and Update Status Report on the Woodland Caribou; Committee on the Status of Endangered Wildlife of Canada: Ottawa, ON, Canada, 2002. [Google Scholar]
- McLoughlin, P.D.; Dzus, E.; Wynes, B.; Boutin, S. Declines in Populations of Woodland Caribou. J. Artic. 2019, 67, 755–761. [Google Scholar] [CrossRef]
- Dickie, M.; Serrouya, R.; McNay, R.S.; Boutin, S. Faster and farther: Wolf movement on linear features and implications for hunting behaviour. J. Appl. Ecol. 2017, 54, 253–263. [Google Scholar] [CrossRef]
- Wotton, B.M.; Flannigan, M.D.; Marshall, G.A. Potential climate change impacts on fire intensity and key wildfire suppression thresholds in Canada. Environ. Res. Lett. 2017, 12, 9. [Google Scholar] [CrossRef]
- Klein, D.R. Fire, Lichens, and Caribou. J. Range Manag. 2007, 35, 390. [Google Scholar] [CrossRef]
- Metsaranta, J.M. Assessing the length of the post-disturbance recovery period for woodland caribou habitat after fire and logging in west-central Manitoba. Rangifer 2007, 27, 103. [Google Scholar] [CrossRef][Green Version]
- Petzold, D.E.; Goward, S.N. Reflectance spectra of subarctic lichens. Remote Sens. Environ. 1988, 24, 481–492. [Google Scholar] [CrossRef]
- Gilichinsky, M.; Sandström, P.; Reese, H.; Kivinen, S.; Moen, J.; Nilsson, M. Mapping ground lichens using forest inventory and optical satellite data. Int. J. Remote Sens. 2011, 32, 455–472. [Google Scholar] [CrossRef]
- Nelson, P.R.; Roland, C.; Macander, M.J.; McCune, B. Detecting continuous lichen abundance for mapping winter caribou forage at landscape spatial scales. Remote Sens. Environ. 2013, 137, 43–54. [Google Scholar] [CrossRef]
- Rosso, A.; Neitlich, P.; Smith, R.J. Non-destructive lichen biomass estimation in northwestern Alaska: A comparison of methods. PLoS ONE 2014, 9, e103739. [Google Scholar] [CrossRef]
- Lim, K.; Treitz, P.; Wulder, M.; St-Ongé, B.; Flood, M. LiDAR remote sensing of forest structure. Prog. Phys. Geogr. 2003, 27, 88–106. [Google Scholar] [CrossRef]
- Falldorf, T.; Strand, O.; Panzacchi, M.; Tømmervik, H. Estimating lichen volume and reindeer winter pasture quality from Landsat imagery. Remote Sens. Environ. 2014, 140, 573–579. [Google Scholar] [CrossRef]
- Kennedy, B.; Pouliot, D.; Manseau, M.; Fraser, R.; Duffe, J.; Pasher, J.; Chen, W.; Olthof, I. Assessment of Landsat-based terricolous macrolichen cover retrieval and change analysis over caribou ranges in northern Canada and Alaska. Remote Sens. Environ. 2020, 240, 111694. [Google Scholar] [CrossRef]
- Konkolics, S.M. A Burning Question: The Spatial Response of Woodland Caribou to Wildfire in Northeastern Alberta. Master’s Thesis, Department of Biological Sciences, University of Alberta, Edmonton, AB, Canada, 2019. [Google Scholar]
- Downing, D.J.; Pettapiece, W.W. Natural Regions and Subregions of Alberta; Government of Albeta: Stony Plain, AB, Canada, 2006; ISBN 0778545725.
- Alberta Wildfire. Historical Wildfire Perimeter Data: 1931–2017. Available online: http://wildfire.alberta.ca/resources/historical-data/spatial-wildfire-data.aspx (accessed on 1 April 2017).
- Ducks Unlimited. Enhanced Wetland Classifcation Layer; Ducks Unlimited: Edmonton, AB, Canada, 2011. [Google Scholar]
- Guo, X.; Coops, N.C.; Tompalski, P.; Nielsen, S.E.; Bater, C.W.; John Stadt, J. Regional mapping of vegetation structure for biodiversity monitoring using airborne lidar data. Ecol. Inform. 2017, 38, 50–61. [Google Scholar] [CrossRef]
- Alberta Environment and Sustainable Resource Develeopment. General Specifications for Acquisition of LiDAR Data; Environment and Sustainable Resource Development: Edmonton, AB, Canada, 2013. [Google Scholar]
- McGaughey, R.J. FUSION/LDV: Software for LIDAR Data Analysis and Visualization; US Department of Agriculture Forest Service Pacific Northwest Research Station: Seattle, WA, USA, 2014.
- Alberta Geological Survey. Surficial Geology of Alberta, Generalized Digital Mosaic (DIG 2013-0002). Available online: https://geology-ags-aer.opendata.arcgis.com/datasets/surficial-geology-of-alberta-generalized-digital-mosaic-dig-2013-0002 (accessed on 1 April 2017).
- Alberta Agriculture and Forestry. Wet Areas Mapping. Available online: https://geodiscover.alberta.ca/geoportal/rest/metadata/item/2ef5a9d1b2154067bc1af0160499ef3c/html (accessed on 1 April 2017).
- Korean Aerospace Research Institute. KOMPSAT-3 Data; Korean Aerospace Research Institute: Daejeon, Korea, 2016.
- French Space Agency. Pleidates 1 Satellite Data; French Space Agency: Paris, French, 2016.
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2011. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Barton, K. Mu-MIn: Multi-model Inference. 2009. Available online: http://R-Forge.R-project.org/projects/mumin/ (accessed on 1 September 2017).
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved-A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- Majasalmi, T.; Rautiainen, M. The impact of tree canopy structure on understory variation in a boreal forest. For. Ecol. Manag. 2020, 466, 118100. [Google Scholar] [CrossRef]
- Haughian, S.R.; Haughian, S.R. Microhabitat associations of lichens, feathermosses, and vascular plants in a caribou winter range, and their implications for understory development. Botany 2015, 93, 221–231. [Google Scholar] [CrossRef]
- Peltoniemi, J.I.; Kaasalainen, S.; Näränen, J.; Rautiainen, M.; Stenberg, P.; Smolander, H.; Smolander, S.; Voipio, P. BRDF measurement of understory vegetation in pine forests: Dwarf shrubs, lichen, and moss. Remote Sens. Environ. 2005, 94, 343–354. [Google Scholar] [CrossRef]
- Scotter, G.W. Growth Rates of Cladonia Alpestris, C. Mitis, and C. Rangiferina in the Taltson River Region, N.W.T. Can. J. Bot. 1963, 41, 1199–1202. [Google Scholar] [CrossRef]





| Predictor Variable | Minimum | Median | Maximum | Mean (SE) |
|---|---|---|---|---|
| Canopy cover < 25% | ||||
| ALS-derived canopy cover (%) | 6 | 25 | 64 | 26.92 (3.31) |
| KOMPSAT-3 blue band (reflectance value) | 0.30 | 0.60 | 1.00 | 0.60 (0.03) |
| DWT (m) | 2 | 5 | 9 | 5.08 (0.40) |
| Canopy cover 25–50% | ||||
| ALS-derived canopy cover (%) | 13 | 53 | 59 | 48.96 (2.14) |
| KOMPSAT-3 blue band (reflectance value) | 0.07 | 0.31 | 0.73 | 0.34 (0.04) |
| DWT (m) | 0 | 3 | 10 | 3.92 (0.62) |
| Canopy cover > 50% | ||||
| ALS-derived canopy cover (%) | 57 | 68 | 82 | 68.08 (1.54) |
| KOMPSAT-3 blue band (reflectance value) | 0.00 | 0.13 | 0.25 | 0.13 (0.01) |
| DWT (m) | 0 | 1 | 4 | 1.68 (0.26) |
| Model Name | Model Structure |
|---|---|
| Blue spectrum | Biomass ~ band 3 |
| Canopy cover | Biomass ~ canopy |
| Depth-to-water (DWT) | biomass ~ logDWT |
| Blue + Canopy cover | biomass ~ band 3 + canopy |
| Blue + DWT | biomass ~ band 3 + log DWT |
| Blue × Canopy cover | biomass ~ band 3 + canopy + band3 × canopy |
| Blue × DWT Null | biomass ~ band 3 + logDWT + band3 × logDWT biomass ~ 1 + plot random effect |
| Model | K | AIC | ∆AIC | wi |
|---|---|---|---|---|
| Canopy cover | 2 | 867.3 | 0.0 | 0.433 |
| Blue + Canopy cover | 3 | 868.2 | 0.9 | 0.276 |
| Blue + DWT | 3 | 869.9 | 2.6 | 0.118 |
| Blue spectrum | 2 | 870.6 | 3.3 | 0.083 |
| Blue × Canopy cover | 4 | 872.0 | 4.7 | 0.041 |
| Blue × DWT | 4 | 873.5 | 6.2 | 0.020 |
| Null | 2 | 873.5 | 6.2 | 0.020 |
| DWT | 2 | 875.1 | 7.8 | 0.009 |
| Variable | β Value | Beta | Std. Error | z | p |
|---|---|---|---|---|---|
| Intercept | 156.70 | 67.53 | 2.32 | 0.020 | |
| band 3 (blue) | 0.03 | 0.15 | 0.03 | 1.06 | 0.290 |
| Lidar (canopy cover) | −1.34 | −0.32 | 0.64 | −2.1 | 0.035 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hillman, A.C.; Nielsen, S.E. Quantification of Lichen Cover and Biomass Using Field Data, Airborne Laser Scanning and High Spatial Resolution Optical Data—A Case Study from a Canadian Boreal Pine Forest. Forests 2020, 11, 682. https://doi.org/10.3390/f11060682
Hillman AC, Nielsen SE. Quantification of Lichen Cover and Biomass Using Field Data, Airborne Laser Scanning and High Spatial Resolution Optical Data—A Case Study from a Canadian Boreal Pine Forest. Forests. 2020; 11(6):682. https://doi.org/10.3390/f11060682
Chicago/Turabian StyleHillman, Ashley C., and Scott E. Nielsen. 2020. "Quantification of Lichen Cover and Biomass Using Field Data, Airborne Laser Scanning and High Spatial Resolution Optical Data—A Case Study from a Canadian Boreal Pine Forest" Forests 11, no. 6: 682. https://doi.org/10.3390/f11060682
APA StyleHillman, A. C., & Nielsen, S. E. (2020). Quantification of Lichen Cover and Biomass Using Field Data, Airborne Laser Scanning and High Spatial Resolution Optical Data—A Case Study from a Canadian Boreal Pine Forest. Forests, 11(6), 682. https://doi.org/10.3390/f11060682