Radial Growth Adaptability to Drought in Different Age Groups of Picea schrenkiana Fisch. & C.A. Mey in the Tianshan Mountains of Northwestern China


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Sampling
2.3. Meteorology Data
2.4. Data Processing
2.4.1. Relationship between Radial Growth and Climatic Factors
2.4.2. Pointer Year Selection
2.4.3. Resistance Indexes Calculation and Difference Comparison
3. Results
3.1. Statistical Characteristics of Tree-Ring Chronologies in the Three Age Groups of Schrenk Spruce
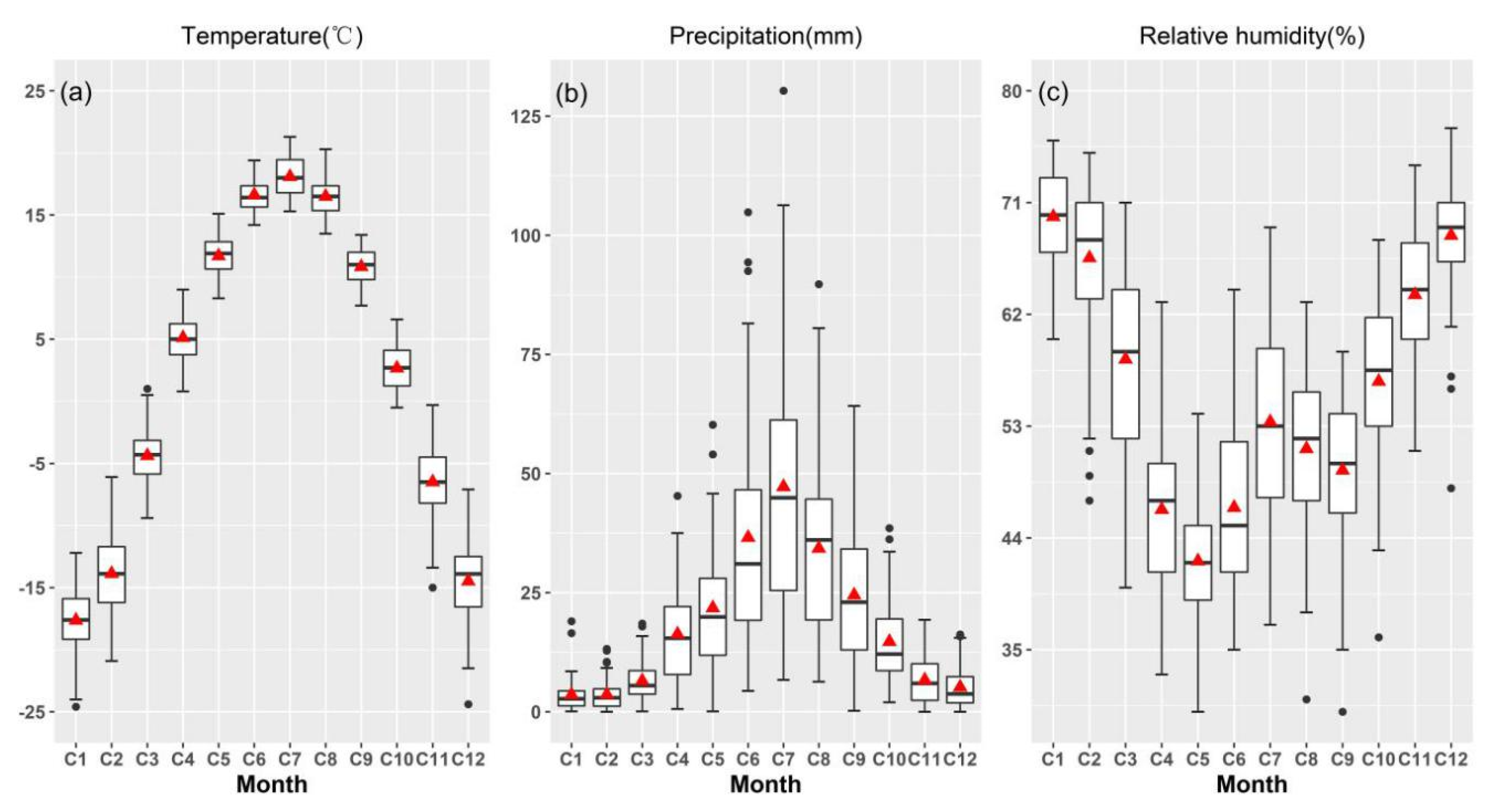
3.2. Inter-Annual Variability of Climate in the Study Regions
3.3. Relationship between Radial Growth and Climatic Factors in Three Age Groups
3.3.1. Relationship between Radial Growth and Temperature in Three Age Groups
3.3.2. Relationship between Radial Growth and Moisture Conditions (Precipitation and Relative Humidity) in Three Age Groups
3.3.3. Relationship between Radial Growth and SPEI in Three Age Groups
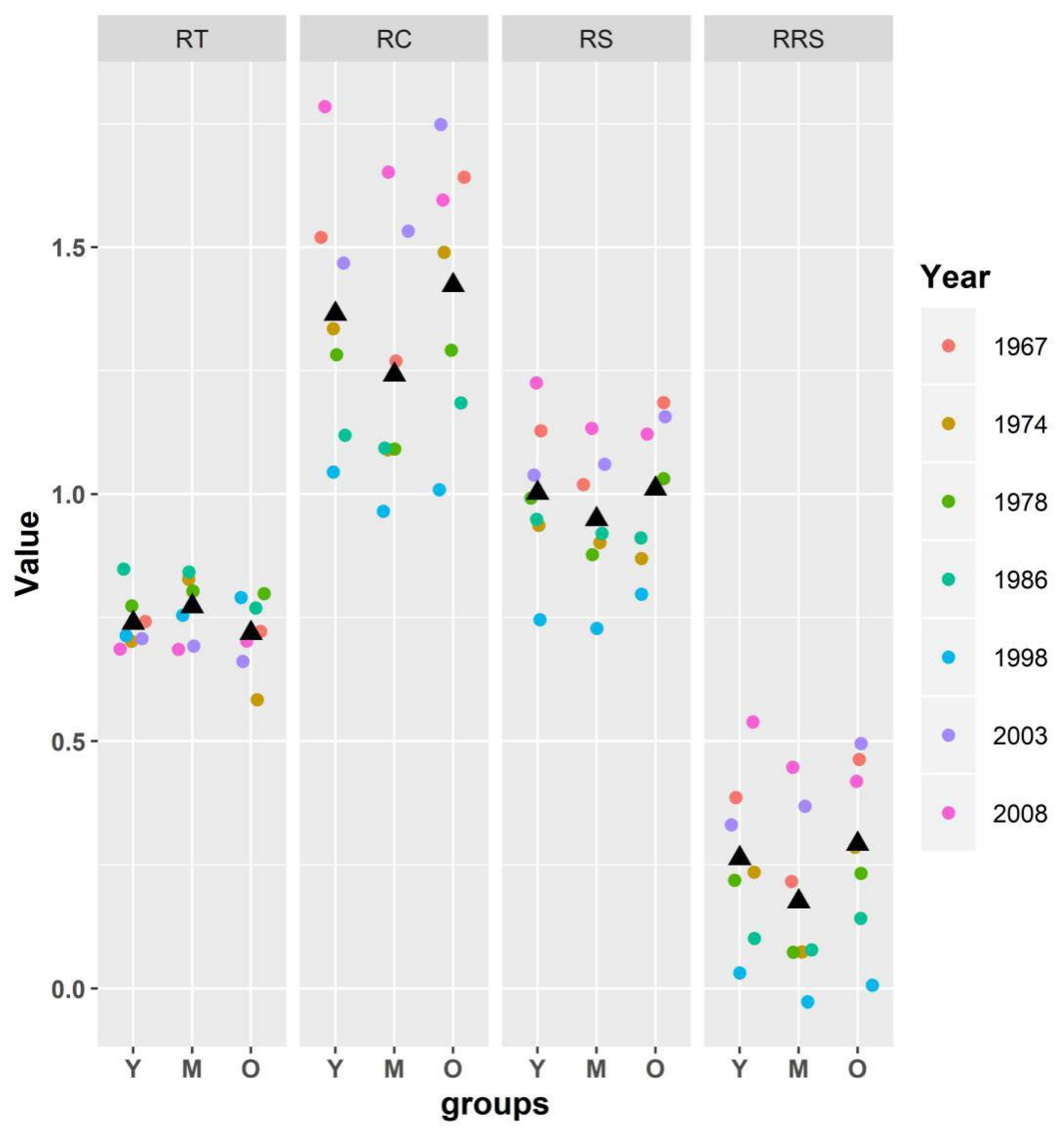
3.4. Resistance Variation of Radial Growth to Drought in Three Age Groups
4. Discussion
4.1. Response Discrepancy of Radial Growth to Drought in Three Age Groups of Schrenk Spruce
4.2. Resistance Discrepancy of Radial Growth to Drought in Three Age Groups of Schrenk Spruce
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zwiers, F.W.; Alexander, L.V.; Hegerl, G.C.; Knutson, T.R.; Kossin, J.P.; Naveau, P.; Nicholls, N.; Schär, C.; Seneviratne, S.; Zhang, X. Climate extremes: Challenges in estimating and understanding recent changes in the frequency and intensity of extreme climate and weather events. In Climate Science for Serving Society; Springer: Dordrecht, The Netherlands, 2013; pp. 339–389. [Google Scholar] [CrossRef]
- Hartmann, D.L.; Tank, A.; Rusticucci, M. IPCC fifth assessment report, climate change 2013: The physical science basis. IPCC Ar5 2013, 5, 31–39. [Google Scholar]
- Sun, W.; Mu, X.; Song, X.; Wu, D.; Cheng, A.; Qiu, B. Changes in extreme temperature and precipitation events in the Loess Plateau (China) during 1960–2013 under global warming. Atmos. Res. 2016, 168, 33–48. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate change and forest disturbances: Climate change can affect forests by altering the frequency, intensity, duration, and timing of fire, drought, introduced species, insect and pathogen outbreaks, hurricanes, windstorms, ice storms, or landslides. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Shi, C.; Masson-Delmotte, V.; Daux, V.; Li, Z.; Zhang, Q.B. An unstable tree-growth response to climate in two 500 year chronologies, North Eastern Qinghai-Tibetan Plateau. Dendrochronologia 2010, 28, 225–237. [Google Scholar] [CrossRef]
- Liu, H.; Williams, A.P.; Allen, C.D.; Guo, D.; Wu, X.; Anenkhonov, O.A.; Liang, E.; Sandanov, D.V.; Yin, Y.; Qi, Z.; et al. Rapid warming accelerates tree growth decline in semi-arid forests of Inner Asia. Glob. Chang. Biol. 2013, 19, 2500–2510. [Google Scholar] [CrossRef]
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Cook, E.R.; Johnson, A.H. Climate change and forest decline: A review of the red spruce case. Water Air Soil Pollut. 1989, 48, 127–140. [Google Scholar] [CrossRef]
- Mei, S.X. Progress of tree-ring chronology. Quat. Sci. 1997, 17, 265–271. [Google Scholar]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef]
- Phillips, O.L.; Van Der Heijden, G.; Lewis, S.L.; López-González, G.; Aragão, L.E.; Lloyd, J.; Malhi, Y.; Monteagudo, A.; Almeida, S.; Dávila, E.A.; et al. Drought–mortality relationships for tropical forests. New Phytol. 2010, 187, 631–646. [Google Scholar] [CrossRef]
- Clark, J.S.; Iverson, L.; Woodall, C.W.; Allen, C.D.; Bell, D.M.; Bragg, D.C.; D’Amato, A.W.; Davis, F.W.; Hersh, M.H.; Ibanez, I.; et al. The impacts of increasing drought on forest dynamics, structure, and biodiversity in the United States. Glob. Chang. Biol. 2016, 22, 2329–2352. [Google Scholar] [CrossRef] [PubMed]
- Madrigal-González, J.; Herrero, A.; Ruiz-Benito, P.; Zavala, M.A. Resilience to drought in a dry forest: Insights from demographic rates. For. Ecol. Manag. 2017, 389, 167–175. [Google Scholar] [CrossRef]
- Aakala, T.; Kuuluvainen, T. Summer droughts depress radial growth of Picea abies in pristine taiga of the Arkhangelsk province, northwestern Russia. Dendrochronologia 2011, 29, 67–75. [Google Scholar] [CrossRef]
- Peng, C.H.; Ma, Z.H.; Lei, X.D.; Zhu, Q.; Chen, H.; Wang, W.F.; Liu, S.R.; Li, W.Z.; Fang, X.Q.; Zhou, X.L. A drought-induced pervasive increase in tree mortality across Canada’s boreal forests. Nat. Clim. Chang. 2011, 1, 467–471. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Camarero, J.J.; Manzanedo, R.D.; Sanchez-Salguero, R.; Navarro-Cerrillo, R.M. Growth response to climate and drought change along an aridity gradient in the southernmost Pinus nigra relict forests. Ann. For. Sci. 2013, 70, 769–780. [Google Scholar] [CrossRef]
- D’Amato, A.W.; Bradford, J.B.; Fraver, S.; Palik, B.J. Effects of thinning on drought vulnerability and climate response in north temperate forest ecosystems. Ecol. Appl. 2013, 23, 1735–1742. [Google Scholar] [CrossRef]
- Bottero, A.; D’Amato, A.W.; Palik, B.J.; Bradford, J.B.; Fraver, S.; Battaglia, M.A.; Asherin, L.A. Density-dependent vulnerability of forest ecosystems to drought. J. Appl. Ecol. 2017, 54, 1605–1614. [Google Scholar] [CrossRef]
- Cook, E.R. A Time Series Analysis Approach to Tree Ring Standardization. Ph.D. Thesis, University of Arizona, Tucson, AZ, USA, 1985. [Google Scholar]
- Ding, W.X. Tree Rings and Climate Change; Meteorological Press: Beijing, China, 1990; pp. 63–70. [Google Scholar]
- Voelker, S.L. Age-dependent changes in environmental influences on tree growth and their implications for forest responses to climate change. In Size- and Age-Related Changes in Tree Structure and Function; Springer: Dordrecht, The Netherlands, 2011; pp. 455–479. [Google Scholar] [CrossRef]
- Kramer, P. Physiology of Woody Plants; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Wang, X.; Zhang, Y.; McRae, D.J. Spatial and age-dependent tree-ring growth responses of Larix gmelinii to climate in northeastern China. Trees 2009, 23, 875–885. [Google Scholar] [CrossRef]
- Wu, G.; Xu, G.; Chen, T.; Liu, X.; Zhang, Y.; An, W.; Wang, W.; Fang, Z.; Yu, S. Age-dependent tree-ring growth responses of Schrenk spruce (Picea schrenkiana) to climate—A case study in the Tianshan Mountain, China. Dendrochronologia 2013, 31, 318–326. [Google Scholar] [CrossRef]
- Madrigal-González, J.; Zavala, M.A. Competition and tree age modulated last century pine growth responses to high frequency of dry years in a water limited forest ecosystem. Agric. For. Meteorol. 2014, 192, 18–26. [Google Scholar] [CrossRef]
- Primicia, I.; Camarero, J.J.; Janda, P.; Čada, V.; Morrissey, R.C.; Trotsiuk, V.; Bače, R.; Teodosiu, M.; Svoboda, M. Age, competition, disturbance and elevation effects on tree and stand growth response of primary Picea abies forest to climate. For. Ecol. Manag. 2015, 354, 77–86. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; López, B.C.; Loepfe, L.; Lloret, F. Stand-and tree-level determinants of the drought response of Scots pine radial growth. Oecologia 2012, 168, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Lei, P.; Kwon, S.; Shu, L.Y.; Hui, Y.X.; Jie, S.Z. The radial growth response of natural pinus camphora in sandy land to drought. J. Beijing For. Univ. 2018, 7, 27–35. [Google Scholar] [CrossRef]
- Gao, L.; Guo, X.; Deng, Y.; Yang, M.; Huo, Y.; Chen, Q. The advance of dendroclimatology in arid area of Northwest China. Mar. Geol. Quat. Geol. 2013, 33, 25. [Google Scholar] [CrossRef]
- Feng, L. Climate of Xinjiang; Meteorology Press: Beijing, China, 1991; pp. 97–104. [Google Scholar]
- Jing, Y.X.; Yong, Z. Simulation of the influence of topography on summer precipitation in Tianshan Mountains. J. Desert Reach 2016, 36, 1133–1143. [Google Scholar] [CrossRef]
- Chen, F.H.; Chen, J.H.; Holmes, J.; Boomer, I.; Austin, P.; Gates, J.B.; Wang, N.L.; Brooks, S.J.; Zhang, J.W. Moisture changes over the last millennium in arid central Asia: A review, synthesis and comparison with monsoon region. Quat. Sci. Rev. 2010, 29, 1055–1068. [Google Scholar] [CrossRef]
- Jiao, L.; Jiang, Y.; Wang, M.; Xin, K.; Wen, Z.; Ling, Z.; Shou, Z. Responses to climate change in radial growth of Picea schrenkiana along elevations of the eastern Tianshan Mountains, northwest China. Dendrochronologia 2016, 40, 117–127. [Google Scholar] [CrossRef]
- Zhang, R.; Yuan, Y.; Gou, X.; Zhang, T.; Zou, C.; Ji, C.; Fan, Z.; Qin, L.; Shang, H.; Li, X. Intra-annual radial growth of Schrenk spruce (Picea schrenkiana Fisch. et Mey) and its response to climate on the northern slopes of the Tianshan Mountains. Dendrochronologia 2016, 40, 36–42. [Google Scholar] [CrossRef]
- Duff, G.H.; Nolan, N.J. Growth and morphogenesis in the Canadian forest species: I. The controls of cambial and apical activity in Pinus resinosa Ait. Can. J. Bot. 1953, 31, 471–513. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Growth and Development of Trees; Academic Press: New York, NY, USA, 1971. [Google Scholar]
- Harcombe, P.A. Tree life tables. Bioscience 1987, 37, 557–568. [Google Scholar] [CrossRef]
- Bond, B.J. Age-related changes in photosynthesis of woody plants. Trends Plant Sci. 2000, 5, 349–353. [Google Scholar] [CrossRef]
- England, J.R.; Attiwill, P.M. Changes in leaf morphology and anatomy with tree age and height in the broadleaved evergreen species, Eucalyptus regnans F. Muell. Trees 2006, 20. [Google Scholar] [CrossRef]
- Vieira, J.; Campelo, F.; Nabais, C. Age-dependent responses of tree-ring growth and intra-annual density fluctuations of Pinus pinaster to Mediterranean climate. Trees 2009, 23, 257–265. [Google Scholar] [CrossRef]
- Ting, W.; Bao, R.; Ping, M. Preliminary study on population dynamics of Schrenk spruce in Tianshan Mountain of central Xinjiang. Ecol. Environ. Sci. 2006, 15, 564–571. [Google Scholar] [CrossRef]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; The University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree Ring Bull. 1983, 43, 69–95. [Google Scholar]
- Szeicz, J.M.; MacDonald, G.M. Age-dependent tree-ring growth responses of subarctic white spruce to climate. Can. J. For. Res. 1994, 24, 120–132. [Google Scholar] [CrossRef]
- Jiao, L.; Jiang, Y.; Wang, M.; Wen, Z.; Yi, Z. Age-effect radial growth responses of Picea schrenkiana to climate change in the eastern Tianshan Mountains, Northwest China. Forests 2017, 8, 294. [Google Scholar] [CrossRef]
- Fritts, H.C.; Shatz, D.J. Selecting and characterizing tree-ring chronologies for dendroclimatic analysis. Tree Ring Bull. 1975, 35, 31–46. [Google Scholar]
- Wigley, T.M.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Jiao, L.; Jiang, Y.; Zhang, W.; Wang, M.; Wang, S.; Liu, X. Assessing the stability of radial growth responses to climate change by two dominant conifer trees species in the Tianshan Mountains, northwest China. For. Ecol. Manag. 2019, 433, 667–677. [Google Scholar] [CrossRef]
- Zhao, N.; Jiao, Y.; Ma, T.; Zhao, M.; Fan, Z.; Yin, X.; Liu, Y.; Yue, T. Estimating the effect of urbanization on extreme climate events in the Beijing-Tianjin-Hebei region, China. Sci. Total. Environ. 2019, 688, 1005–1015. [Google Scholar] [CrossRef]
- Beguería, S.; Vicente-Serrano, S.M.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatol. 2014, 34, 3001–3023. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, Y.; Zhao, S.; Jiao, L.; Wen, Y. Relationships between tree age and climate sensitivity of radial growth in different drought conditions of Qilian Mountains, northwestern China. Forests 2018, 9, 135. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Schweingruber, F.; Eckstein, D.; Serrebachet, F.; Bra¨ker, O.U. Identification, presentation and interpretation of event years and pointer years in dendrochronology. Dendrochronologia 1990, 9, 9–38. [Google Scholar]
- Sánchez-Salguero, R.; Camarero, J.J.; Dobbertin, M.; Fernández-Cancio, Á.; Vilà-Cabrera, A.; Manzanedo, R.D.; Zaval, M.A.; Navarro-Cerrillo, R.M. Contrasting vulnerability and resilience to drought-induced decline of densely planted vs. natural rear-edge Pinus nigra forests. For. Ecol. Manag. 2013, 310, 956–967. [Google Scholar] [CrossRef]
- Gao, S.; Liu, R.; Zhou, T.; Fang, W.; Yi, C.; Lu, R.; Zhao, X.; Luo, H. Dynamic responses of tree-ring growth to multiple dimensions of drought. Glob. Chang. Biol. 2018, 24, 5380–5390. [Google Scholar] [CrossRef]
- Xu, M.; Kang, S.; Wu, H.; Xu, Y. Detection of spatio-temporal variability of air temperature and precipitation based on long-term meteorological station observations over Tianshan Mountains, Central Asia. Atmos. Res. 2018, 203, 141–163. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, M.; Sun, M.; Bao, W.; Xiao, L. Changes in precipitation extremes in alpine areas of the Chinese Tianshan Mountains, central Asia, 1961–2011. Quat. Int. 2013, 311, 97–107. [Google Scholar] [CrossRef]
- Nearing, M.A.; Jetten, V.; Baffaut, C.; Cerdan, O.; Couturier, A.; Hernandez, M.; Le Bissonnais, Y.; Nichols, M.H.; Nunes, J.P.; Renschler, C.S.; et al. Modeling response of soil erosion and runoff to changes in precipitation and cover. Catena 2005, 61, 131–154. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, J.; Su, W.; Zhao, X.; Xu, X. Spring precipitation effects on formation of first row of earlywood vessels in Quercus variabilis at Qinling Mountain (China). Trees 2018, 33, 457–468. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Lachenbruch, B.; Dawson, T.E. Size- and Age-Related Changes in Tree Structure and Function. In Tree physiology; Springer: Dordrecht, The Netherlands, 2011; Volume 4. [Google Scholar] [CrossRef]
- Will, R.E.; Wilson, S.M.; Zou, C.B.; Hennessey, T.C. Increased vapor pressure deficit due to higher temperature leads to greater transpiration and faster mortality during drought for tree seedlings common to the forest–grassland ecotone. New Phytol. 2013, 200, 366–374. [Google Scholar] [CrossRef]
- Trugman, A.T.; Detto, M.; Bartlett, M.K.; Medvigy, D.; Anderegg, W.R.L.; Schwalm, C.; Schaffer, B.; Pacala, S.W. Tree carbon allocation explains forest drought-kill and recovery patterns. Ecol. Lett. 2018, 21, 1552–1560. [Google Scholar] [CrossRef]
- Cailleret, M.; Jansen, S.; Robert, E.M.; Desoto, L.; Aakala, T.; Antos, J.A.; Beikircher, B.; Bigler, C.; Bugmann, H.; Caccianiga, M.; et al. A synthesis of radial growth patterns preceeding tree mortality. Glob. Chang. Biol. 2017, 23, 1675–1690. [Google Scholar] [CrossRef]
- Saatchi, S.; Asefi-Najafabady, S.; Malhi, Y.; Aragão, L.E.; Anderson, L.O.; Myneni, R.B.; Nemani, R. Persistent effects of a severe drought on Amazonian forest canopy. Proc. Natl. Acad. Sci. USA 2013, 110, 565–570. [Google Scholar] [CrossRef]
- Mcdowell, N.G.; Williams, A.P.; Xu, C.; Pockman, W.T.; Dickman, L.T.; Sevanto, S.; Pangle, R.; Limousin, J.; Plaut, J.; Mackay, D.S.; et al. Multi-scale predictions of massive conifer mortality due to chronic temperature rise. Nat. Clim. Chang. 2016, 6, 295–300. [Google Scholar] [CrossRef]
- Rozas, V.; DeSoto, L.; Olano, J.M. Sex-specific, age-dependent sensitivity of tree-ring growth to climate in the dioecious tree Juniperus thurifera. New Phytol 2009, 182, 687–697. [Google Scholar] [CrossRef]
- Pompa-García, M.; Hadad, M.A. Sensitivity of pines in Mexico to temperature varies with age. Atmósfera 2016, 29, 209–219. [Google Scholar] [CrossRef]
- Carrer, M.; Urbinati, C. Age-dependent tree-ring growth responses to climate in Larix decidua and Pinus cembra. Ecology 2004, 85, 730–740. [Google Scholar] [CrossRef]
- Rong, L.; Wen, L.H.; Yang, S.Y.; Li, Y.Y. Structure characteristics and spatial distribution of picea schrenkiana population on the northern slopes of the Tianshan Mountain. Arid Land Resour. Environ. 2013, 27, 91–96. [Google Scholar]
- Pallardy, S.G. Physiology of Woody Plants; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Jiao, S.X.; Li, C.S.; Chen, S.C.; Tao, Z. Changes of stoichiometric characteristics in N, P and K in different organs with growth stages of Schrenk spruce. Chin. J. Ecol. 2018, 3, 3–10. [Google Scholar] [CrossRef]
- Gui, L.J.; Kui, P.S.; De, P.C.; Sheng, Z.L. Population statistics and survival analysis of Schrenk spruce in Tianshan Mountains. J. Beijing For. Univ. 2001, 23. [Google Scholar] [CrossRef]
- Yu, G.; Liu, Y.; Wang, X.; Ma, K. Age-dependent tree-ring growth responses to climate in Qilian juniper (Sabina przewalskii Kom.). Trees 2008, 22, 197–204. [Google Scholar] [CrossRef]
- Martín-Benito, D.; Cherubini, P.; Río, M.; Cañellas, I. Growth response to climate and drought in Pinus nigra Arn. trees of different crown classes. Trees 2008, 22, 363–373. [Google Scholar] [CrossRef]
- Merlin, M.; Perot, T.; Perret, S.; Korboulewsky, N.; Vallet, P. Effects of stand composition and tree size on resistance and resilience to drought in sessile oak and Scots pine. For. Ecol. Manag. 2015, 339, 22–33. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistical Parameters | Young Group | Middle Group | Old Group |
|---|---|---|---|
| Sample depth (core/tree number) | 36/18 | 42/21 | 38/19 |
| Sequence length | 1914–2012 (99) | 1823–2012 (190) | 1735–2012 (278) |
| Mean age | 69 ± 18.34 | 122 ± 18.52 | 236 ± 21.05 |
| Mean tree-ring width | 1.724 | 1.294 | 0.590 |
| Standard deviation (SD) | 0.204 | 0.232 | 0.235 |
| Mean sensitivity (MS) | 0.249 | 0.256 | 0.261 |
| Correlation coefficient (R) | 0.419 | 0.533 | 0.608 |
| Mean correlation among trees (R1) | 0.733 | 0.718 | 0.758 |
| Mean correlation between trees (R2) | 0.367 | 0.525 | 0.596 |
| The first eigenvector percentage (PC1) | 0.496 | 0.559 | 0.639 |
| Signal-to-noise ratio (SNR) | 5.776 | 29.668 | 21.734 |
| Expressed population signal (EPS) | 0.852 | 0.967 | 0.956 |
| Temperature | Precipitation | R95p | R99p | Relative Humidity | |
|---|---|---|---|---|---|
| Temperature | 1 | 0.1235 | 0.3299 * | 0.2845 * | −0.7906 ** |
| Precipitation | 1 | 0.6730 ** | 0.5156 ** | 0.0956 | |
| R95p | 1 | 0.6456 ** | −0.1895 | ||
| R99p | 1 | −0.2136 | |||
| Relative humidity | 1 |
| Resistance (RT) | Recovery (RC) | Resilience (RS) | Relative Resilience (RRS) | |
|---|---|---|---|---|
| Y | 0.804 ± 0.0167 ab | 1.263 ± 0.0461 ab | 1.002 ± 0.0272 ab | 0.198 ± 0.0336 ab |
| M | 0.841 ± 0.0180 a | 1.149 ± 0.0462 b | 0.949 ± 0.0239 b | 0.109 ± 0.0345 b |
| O | 0.775 ± 0.0179 b | 1.316 ± 0.0490 a | 1.005 ± 0.0290 a | 0.230 ± 0.0346 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, L.; Liu, X.; Wang, S.; Chen, K. Radial Growth Adaptability to Drought in Different Age Groups of Picea schrenkiana Fisch. & C.A. Mey in the Tianshan Mountains of Northwestern China. Forests 2020, 11, 455. https://doi.org/10.3390/f11040455
Jiao L, Liu X, Wang S, Chen K. Radial Growth Adaptability to Drought in Different Age Groups of Picea schrenkiana Fisch. & C.A. Mey in the Tianshan Mountains of Northwestern China. Forests. 2020; 11(4):455. https://doi.org/10.3390/f11040455
Chicago/Turabian StyleJiao, Liang, Xiaoping Liu, Shengjie Wang, and Ke Chen. 2020. "Radial Growth Adaptability to Drought in Different Age Groups of Picea schrenkiana Fisch. & C.A. Mey in the Tianshan Mountains of Northwestern China" Forests 11, no. 4: 455. https://doi.org/10.3390/f11040455
APA StyleJiao, L., Liu, X., Wang, S., & Chen, K. (2020). Radial Growth Adaptability to Drought in Different Age Groups of Picea schrenkiana Fisch. & C.A. Mey in the Tianshan Mountains of Northwestern China. Forests, 11(4), 455. https://doi.org/10.3390/f11040455