Abstract
Hardwood-dominated forests in south-central Chile have shade-tolerant and mid-tolerant tree species capable of regenerating and growing well in partial shade. To test the potential for using an uneven-aged silviculture in these forests, we established single-tree selection treatments at two mid-elevation sites within the Evergreen forest type in the Coastal range (Llancahue and Los Riscos, 40–42° S Lat). They had an average initial basal area of 70–80 m2 ha−1. In each stand, we established four 2000 m2 plots with a residual basal area of ~40 m2 ha−1, and four with a residual basal area of ~60 m2 ha−1. We planned for a maximum residual diameter of 80 cm, but needed to leave 20%–25% of the residual basal area in larger trees due to their great abundance in these old-growth forests. We re-measured these plots 5–6 years after the cuttings. We used mixed-effects models to evaluate the periodic annual increment (pai) in diameter and the abundance of tree regeneration, and linear models to evaluate ingrowth and changes in the basal area and volume. At Llancahue, the diameter pai of individual trees was significantly greater in the treatment with lower residual densities, especially for mid-tolerant species in lower diameter classes (5–20 cm). At both sites, the pai in the stand basal area and volume was greater in the more heavily stocked treatment, but differences were significant only at Llancahue. Regeneration was dominated by shade-tolerant species at both sites but was more abundant and more diverse at Llancahue. Taller tree regeneration (50–<200 cm) significantly increased after the cuttings at both sites, while small regeneration (5–<50 cm) overall remained at pre-cut levels. This pattern was similar for mid- and shade-tolerant species. However, we found no differences in regeneration responses between the lower and higher levels of the residual basal area. Sapling densities did not differ at both sites for shade-tolerant species, but for mid-tolerant species these were more abundant at Los Riscos. While both sites had many similar trends after implementing selection cuts (a greater individual growth in the treatment with lower basal areas but a higher stand-level growth in the treatment with a high basal area, more abundant regeneration of shade-tolerant species, etc.), they illustrate a differential potential for implementing uneven-aged silviculture, especially due to site-species interactions. These results are a first step towards evaluating the prospects for selection cuttings in these experiments and elsewhere in Valdivian temperate rainforests.
1. Introduction
Foresters across the world have expressed interest in methods for uneven-aged and continuous cover forestry (CCF) [1,2,3]. These include a range of silvicultural treatments, which largely fall into selection systems and irregular shelterwood systems. With selection systems, harvesting and regeneration take place simultaneously and continuously, and tree sizes and ages are inter-mixed singly or in groups [4]. While single-tree selection systems have traditionally been most effective in stands dominated by the more shade-tolerant tree species, group selection systems can provide some flexibility for managing mid-tolerant species [5], as may single-tree selection regimes that use a low-density residual [4,6,7], also when managing stands with mid-tolerant species that have a strong root sprouting capacity [8].
Valdivian temperate rainforests [9] have species with characteristics favorable to selection systems, and cover an extensive geographical area in Chile (33–45° S). Many different forest types develop in this region due to differences in climate along the latitudinal gradient and associated with the Coastal and Andean ranges. The Evergreen forest type is most prevalent, covering nearly 4 million ha. It is dominated by shade-tolerant or mid-tolerant evergreen hardwood species [10,11,12,13,14], although mid-tolerant ones (e.g., Eucryphia cordifolia Cav., one of the species with the highest basal area in these forests) are scarce in the regeneration layer and among the smaller tree size classes in forests in advanced successional stages of development [13,15,16,17]. Mature and old-growth stands are uneven-aged, with negative exponential diameter distributions and basal areas as high as 80–100 m2 per hectare ([11,18]; see also Ponce et al. [19], and references therein, for a definition of old-growth forests). The structure and composition of these forests is the result of recurring small-scale disturbances [20]. The dynamics of mature and old-growth forests are mostly driven by the gap regeneration (e.g., for mid-tolerant E. cordifolia and Drymis winteri J.R. et G. Forster; sensu [21]) and the continuous regeneration modes (e.g., for shade-tolerant Laureliopsis philippiana (Looser) Schodde, Aextoxicon punctatum R. et Pav., and Persea lingue (Ruiz & Pav.) Nees [22]).
Selection silviculture is uncommon in Chile, and we know only one field-based study that examined single-tree selection cutting in Valdivian rainforests [15], in one of the two experiments that we evaluate here. They documented changes in the stand structure and mature tree species composition, and concluded that single-tree selection can maintain many old-growth attributes of these forests. However, they did not analyze tree growth or tree regeneration responses.
Evaluating these forest responses is warranted before recommending widespread use of this forest management regime. To that end, we evaluated 5–6 year responses after single-tree selection cuttings at two low-elevation sites in the Coastal range of south-central Chile, where old-growth forests have traditionally been partially cut in the past [15]. We hypothesized that selection system cutting to a low residual basal area (LRBA) would result in better residual tree growth and would enable the regeneration of mid-tolerant tree species, while shade-tolerant ones would dominate the treatment with a higher residual basal area (HRBA). Our objectives were to: (a) Develop a mid-term evaluation of regeneration, growth, and production following selection cutting at two sites; and (b) Evaluate the effect of two levels of residual basal areas on the development of mid-tolerant and shade-tolerant species. We did not select unharvested stands for evaluation at these sites (control), since our aim is to provide and discuss new knowledge for forest matrices that are mostly harvested in these landscapes, although most times without silviculture [23]. Although we acknowledge this limitation in this study, we provide several citations on untreated old-growth forests in the region as a reference.
2. Methods
2.1. Study Area
We selected stands at Llancahue (LL; 300 m a.s.l., 39° 50′ 20″ S and 73° 07′ 18″ W) and Los Riscos (LR; 250 m a.s.l., 40° 53′ 07″ S and 73° 31′ 30″ W) (Figure 1). Both sites had been partially harvested in the past through illegal selective cuttings, leaving average basal areas in the range of 70 to 80 m2 ha−1. These mid-elevation forests correspond to the Evergreen forest type, and specifically to the subtype dominated by shade-tolerant species and having a few emergent Nothofagus trees [13]. The main species at each site are listed for Llancahue (Table 1) and Los Riscos (Table 2) and are identified by different functional groups; shade-tolerant, mid-tolerant, and intolerant of shade. In the Evergreen forest type, the shade-tolerant species have a good regeneration potential [16]. The mid-tolerant ones usually occur as medium- and large-sized trees but are scarce in the regeneration layer and among the smaller tree size classes [13,15,16,17]. Shade-intolerant species, if present, are usually constrained to large-sized trees, and their regeneration in canopy gaps is <1% [20].
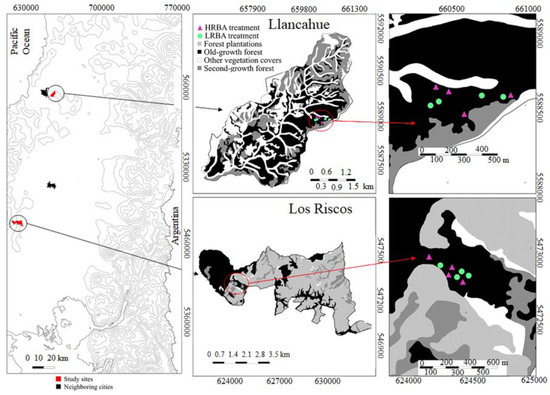
Figure 1.
Location of the study sites and the position of the high residual basal area (HRBA, n = 8) and low residual basal area (LRBA, n = 8) plots at the two sites.

Table 1.
Mean of the regeneration density (per ha values) of seedlings and saplings according to species and treatment (HRBA and LRBA) in Llancahue (LL) following the cuts (2013) and five years after the cuts (2018). Table S7 in the Supplementary Materials provides both the mean and standard deviation.
Table 2.
Mean and standard deviation of the regeneration density (per ha values) of seedlings and saplings according to species and treatment (HRBA and LRBA) in Los Riscos (LR) following the cuts (2013) and six years after the cuts (2019). Table S8 in the Supplementary Materials provides both the mean and standard deviation.
Both study sites have a temperate, rainy climate with a Mediterranean influence [24]. At Llancahue, the annual rainfall is 2300 mm and the average annual temperature is 12.2 °C [25], while at the Purranque weather station (a city near Los Riscos, but closer to the central valley with less rainfall) annual rainfall reaches 1550 mm and the average annual temperature is 10.9 °C [26].
Llancahue lies between 50 and 410 m a.s.l. [27], and soils are moderately deep with a clay texture, originating from ancient volcanic ashes deposited in the metamorphic complex of the Coastal range [28]. Llancahue has mostly old-growth forests with different degrees of past harvesting, generally classed as being only minor disturbances. They are dominated by the shade-tolerants A. punctatum and L. philippiana, and the mid-tolerants E. cordifolia and D. winteri.
Los Riscos is located in a transition between the Intermediate Depression and the Coastal range, with a strongly undulating topography that varies between 220 and 310 m a.s.l. [29]. Soils are moderately deep and originated from Holocene volcanic ash [28]. The old-growth forest of Los Riscos is dominated by the same species as those at Llancahue (A. punctatum, L. philippiana, and E. cordifolia), but D. winteri is nearly absent and P. lingue is common.
2.2. Experimental Design
At each site, eight plots (50 m × 40 m; 2000 m2) were located in old-growth forests that had partial cuts in the past and reverse-J diameter distributions (e.g., [15]), and the locations were selected to ensure that the initial starting conditions for the experiment were as similar as possible using visual criteria. Two treatments (described later) were then randomly allocated to the eight plots so that each treatment had four plots at each site. We marked all the trees at breast height and measured ones with a diameter at breast height (dbh, at 1.3 m) ≥ 5 cm, recording: diameter, tree species according to shade tolerance (shade-tolerant; mid-tolerant; intolerant), and the crown position (emergent, higher and lower canopy, higher and lower reserve, or subcanopy) before the selection cuttings. For a random subsample stratified by the broad diameter classes of trees per plot, we also measured the total height (h) with an ultrasound VERTEX III hypsometer (Haglof Inc., Madison, Mississippi, USA) to develop height-diameter functions to use in estimating heights for all trees. We measured 320 trees in Llancahue and 399 trees in Los Riscos (Tables S1 and S2 in the Supplementary Materials).
The two selection cutting treatments used in the experiment were specified using the BDq approach described by Guldin [30]. The two treatments differed in terms of their residual basal area (B): one was 60 m2 ha−1 (high-HRBA) and the other was 42 m2 ha−1 (low-LRBA; Table 3). Both treatments used a maximum diameter (D) of 80 cm and a q factor of 1.3. However, since these forests have many trees >80 cm [11], for this first selection cut we left 10–13 m2 per ha of these large trees (20% or 25% of the total residual basal area with HRBA and LRBA, respectively), and that basal area was included in B. To implement the selection cutting we followed procedures provided by Nyland [4], and constructed a marking guide that compared the target structure with the actual diameter distribution in each stand. We merged on average four 5-cm diameter classes to deliver a marking guide that included one prescription for each broad range of diameters (e.g., cut one of two trees, cut one of four trees, etc.). The selection criteria used in the cuttings was to leave the best possible quality trees (i.e., quality 1 trees—those with straight boles, well-developed crowns, and no apparent health problems), within the limitations of the residual basal areas imposed in the experimental design. Harvesting was conducted with oxen and closely supervised. As a result, Bacardit [31] determined that damage to the residual trees was marginal in Llancahue, and we estimate that it was similar in Los Riscos. The selection cuttings were conducted during spring-summer of 2012–2013. The harvesting intensities across plots ranged from 25% to 35% of the initial basal area, the latter representing the maximum harvest allowed by the Chilean law for this silvicultural system (Table 3). Therefore, the value chosen for the low basal area is a result of both legal constraints and a conservative estimate in these forests where Chusquea quila Kunth represents a potential hazard for regeneration when too much light is available (which could be the case with lower basal areas; Donoso and Nyland [16], and references therein). In addition, residual basal areas of 60 to 70 percent of the original have been used in regions with decades of selection silviculture [4,32]. The high basal area value was selected since it represents close to two-thirds of the maximum basal area in these forests (see above), e.g., similar to the harvesting intensities elsewhere in hardwood forests subjected to selection cutting [33]. We re-measured these plots after the fifth (LL) and sixth (LR) growing seasons, including ingrowth to the lower diameter class and trees that died. Then, we assessed the tree growth and regeneration, with a special focus on functional groups according to shade tolerances based on information from Donoso [14,22] and Gutiérrez et al. [34].
Table 3.
Numbers of trees and basal area in all plots selected for the study before and after the selection cuttings. LRBA: Low residual basal area; HRBA: high residual basal area. Notice that original values were lower in the LRBA plots.
2.3. Estimation of Stand Variables
With the diameter from each plot, we calculated the basal area (G) and estimated the height (h) for each tree. The height functions dependent on the measured diameters were adjusted following the Stage’s model ([35]; Equation S1 in the Supplementary Materials). We also fit the volume (V) equations for each individual species (Table S3 in the Supplementary Materials). With this information, we determined the stand-level (ha) values for the tree density (N, number of trees), basal area, ingrowth (trees entering the minimum 5 cm diameter during the measured period (last five or six years)), mortality (trees that died during the measured period), and volume post-treatment (2013) and at 5–6 years after treatment (2018–2019). By these means, we calculated the periodic annual increment (pai; value in t1—value in t0 divided by the number of years) in tree diameter, stand basal area, and volume, where the latter two variables included ingrowth and mortality.
Regeneration was sampled by systematically establishing 27 circular subplots of 4 m2 at a distance of 5 m in three parallel transects in each plot. Seedling regeneration was counted by height class (5–<50 cm, 50–<100 cm, 100–<200 cm) and species on three occasions: pre-treatment (dormant season 2012), post-treatment (after growing season 2012–2013), and after 5–6 years. Saplings (trees ≥200 cm in height and <5 cm in d) were measured only at 5–6 years after the cuts.
2.4. Statistical Analyses
For individual trees, we analyzed the growth response of the diameter by functional group (all trees, mid-tolerant species, and shade-tolerant species), using linear mixed models (LMM), following the framework of Zuur et al. [36]. In these, the log of pai in diameter (cm year−1) was used as the response variable. The log(d) and diameter class (5–≤20 cm, 20–≤50 cm, 50–80 cm, >80 cm) in 2013, site (LL or LR), and treatment (HRBA or LRBA) were fixed effects, and significant interactions were included as fixed effects. The nested plots within the sites were identified as a random effect to represent the nested data structure (see Table S4 in the Supplementary Materials).
For the stand-level data, a generalized linear model was conducted for ingrowth (n ha−1) and a linear model for pai in the basal area (m2 ha−1 year−1) and volume (m3 ha−1 year−1), where the treatment (HRBA, LRBA) and site (LL, LR) were independent variables (see Table S6 in the Supplementary Materials). Mortality was nominal in these experiments (from 0.53 to 1.18 m2 ha−1 for the entire period, in trees that averaged from 11 to 21 cm in dbh).
For tree regeneration, we made pairwise comparisons between the two sites for all seedlings and seedlings by functional group. We also analyzed the tree regeneration density for each functional group (shade-intolerant, mid-tolerant, and shade-tolerant species) with generalized linear mixed models. In these models, the response was the regeneration density, the probability density function followed a Poisson distribution (n ha−1), while the year (pre-treatment, post treatment, or 5–6 years later), height class (5–<50, 50–<100, and 100–<200 cm), site (LL and LR), and treatment (HRBA and LRBA), as well as significant interactions, were fixed. The nested subplots within the plot and the sites were identified as random effects to account for the hierarchical structure of the data (see Table S5 in the Supplementary Materials).
We used the Tukey HSD post-hoc test with the emmeans package of R to compare differences between levels with significant variables and interactions. Comparisons were conducted according to treatment (low and high residual basal areas) and site (Llancahue and Los Riscos) for the periodic annual increment (pai) in diameter growth, by classes, and for the regeneration density, by years of measurement and height classes.
All statistical analyses were performed in R [37] using the nlme package for the LMMs [38] and a significance level of α = 0.05. The random and fixed effect structure was determined via using the restricted maximum likelihood (REML) and maximum likelihood estimation, respectively. The final model results are based on the REML estimation.
3. Results
3.1. Diameter Growth
The periodic annual increment (pai) in diameter was significantly higher for mid-tolerant species in the LRBA plots compared to the HRBA plots at Llancahue for the lower diameter classes (<20 cm) and the highest diameter classes (>80 cm) (Figure 2). At Los Riscos, there were no significant differences for any combination of functional group and diameter class (Figure 2).
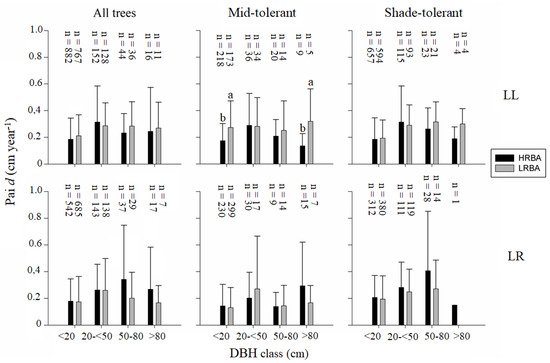
Figure 2.
Mean (bars) and standard deviation (whiskers) in pai in d (cm year−1) according to treatment (HRBA, LRBA), site (LL, LR), DBH CLASS (<20 cm; 20–50 cm; 50–80 cm; >80 cm), and functional group. Different letters between bars of the same DBH class show significant differences (value of p < 0.05).
3.2. Ingrowth
Ingrowth (Figure 3), as measured by trees reaching or surpassing the threshold minimum diameter of 5 cm during the post-cutting period, was significantly higher in the LRBA treatment at Llancahue for all tree species together. At Los Riscos, the HRBA treatment showed more ingrowth, but this was not significantly different from the LRBA plots. We did not statistically compare ingrowth between shade- and mid-tolerant species, but at Llancahue the values were greater for the former, while at Los Riscos they were similar. Furthermore, there was a very dynamic change in density across most diameter classes in all treatment/site combinations (Figure S1 in the Supplementary Materials).
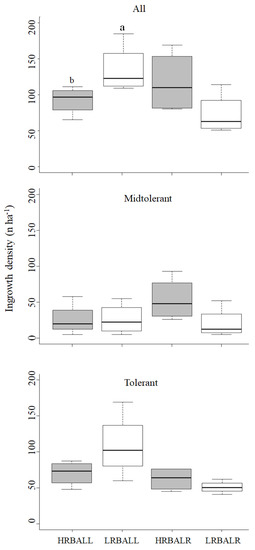
Figure 3.
Ingrowth according to functional group by site (LL or LR) and treatment (HRBA or LRBA). Different letters between the same site show significant differences (p-value < 0.05). Boxes represent the interquartile range (IQR, Q3 minus Q1), including the median of the data, and whiskers include the remaining data.
3.3. Growth in Basal Area and Volume
At Llancahue and Los Riscos, the pai in both the stand basal area and volume were greater in the HRBA plots (a median of 0.7–1.0 vs. 0.4–0.6 m2 ha−1 year−1 in the basal area, and 5–7 vs. 4–5 m3 ha−1 year−1), but these differences were only significant at Llancahue. For equivalent residual basal area classes, Llancahue had a higher pai than Los Riscos (Figure 4).
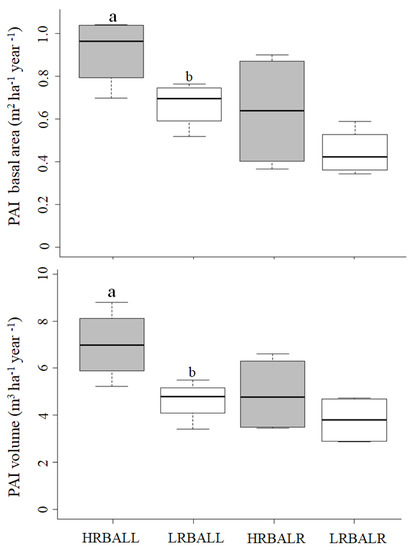
Figure 4.
Pai for the net basal area and volume according to treatment (HRBA, LRBA) and site (LL, LR). Different letters between the same site show significant differences (p-value < 0.05). Boxes represent the interquartile range (IQR, Q3 minus Q1), including the median of the data, and whiskers include the remaining data.
3.4. Regeneration
First, we compared tree regeneration throughout the periods of evaluation (Figure 5). This was dominated by shade-tolerant species at both sites, before and after the cuts (Figure 5). Initial conditions (before the cuts, 2012) showed high tree regeneration densities at both sites (20,000–40,000 seedlings per ha; Figure 5). After the cuts, tree regeneration was more abundant and richer at Llancahue, where it included 24 tree species, while Los Riscos had 17 tree species (Figure 5; Table 1 and Table 2).
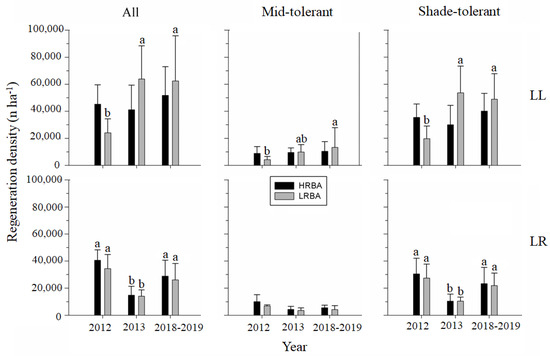
Figure 5.
Mean (bars) and standard deviation (whiskers) in the density of all seedlings (n ha−1) by functional group according to year, treatment, and site. The different letters between bars of the same color show significant differences between the years (value of p < 0.05).
Tree regeneration had significant differences before and after the cuts, and along the post-cut measured period at both sites (Figure 5). At Llancahue, the tree regeneration density was higher after harvesting for the LRBA treatment, and five years after the cuts it did not differ significantly from the post-cut values for all species pooled, nor for the shade tolerance group. The tree regeneration density for the HRBA treatment did not differ significantly across the years. In contrast, at Los Riscos, for both treatments the tree regeneration density was higher before the cuts in both residual density levels for all species pooled and for shade-tolerant species, but six years after harvest it increased to numbers not significantly different from the pre-harvest levels (Figure 5).
The tree regeneration density of mid-tolerant (Figure 6) and shade-tolerant species (Figure 7) showed significant differences throughout measurements for every single height class at both sites. With regards to mid-tolerant species, at Llancahue regeneration behaved differently according to height classes. For the HRBA treatment, the lower regeneration height class (5–<50 cm) had a significant increase after the cuts, but it declined significantly to pre-cut levels five years after. For the LRBA treatment, numbers significantly increased after the cut and remained similar during the evaluation period. Regeneration had similar responses in the intermediate and taller height classes (50–<100 cm and 100–<200 cm) for both treatments, basically with significant increases at the end of the evaluation period. At Los Riscos, both the HRBA and the LRBA treatments showed very similar results throughout the evaluation period. Tree regeneration in the lower height class did not significantly differ between the pre- and post-cut conditions, but it significantly declined six years after the cut. The intermediate and taller height classes showed similar trends, with significant reductions in the tree density after the cuts and with significant increases five years after, meaning a recovery to pre-cut densities.
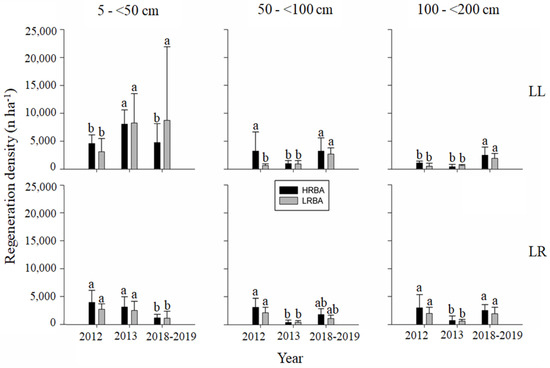
Figure 6.
Mean (bars) and standard deviation (whiskers) in the density of seedlings (n ha−1) for mid-tolerant species according to height classes (5–<50 cm; 50–<100 cm; 100–200 cm), year, treatment, and site. Different letters between bars of the same color show significant differences between measurement years (p-value < 0.05).
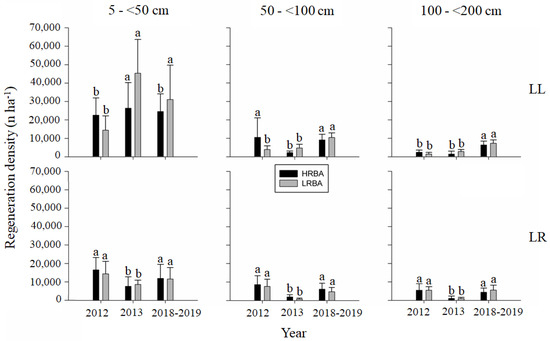
Figure 7.
Mean (bars) and standard deviation (whiskers) in the density of seedlings (n ha−1) for shade-tolerant species according to height classes (5–<50 cm; 50–<100 cm; 100–200 cm), year, treatment and site. Different letters between bars of the same color show significant differences between measurement years (p-value < 0.05).
For shade-tolerant species, at Llancahue tree regeneration in the HRBA treatment significantly increased (5–<50 cm), declined (50–<100 cm), or remained unchanged (100–<200 cm) when comparing pre-cut and post-cut conditions. Five years after the cut, tree regeneration significantly declined to pre-cut levels in the case of the lower height class, and reached significantly greater densities in the intermediate and taller height classes. For the LRBA treatment, regeneration densities significantly increased following the cut in the case of the lower height class, and remained unchanged in the intermediate and taller height classes; and five years after the cut the lower height class did not show any further significant change in regeneration density, but the intermediate and taller height classes had significant increases in comparison with the post-cut densities.
At Los Riscos (Figure 7), the behavior of the tree regeneration of shade-tolerant species was the same for each height class and residual basal area treatment. Tree regeneration significantly declined following the cut, and from there significantly increased six years after the cut to densities similar to those in the pre-cut conditions.
4. Discussion
4.1. General Response Patterns Following Selection Cuttings in Valdivian Rainforests
Privately-owned old-growth or late-successional uneven-aged forests are usually harvested, but they tend to be high-graded, since, beyond theory (e.g., [12,23]), no management prescriptions or options have been tested for them. We evaluated the first trials in the temperate rainforest of Chile (specifically in the Evergreen forest type), aimed at converting unmanaged uneven-aged forests (actually forests with unplanned partial cuts in the past) into selection forests. We implemented the same experiments and protocols at two study sites, based on the facts that these were at similar elevations, were dominated by similar species, and had similar original basal areas before being harvested to the LRBA (lower pre-cut basal areas) or to the HRBA (higher pre-cut basal areas) treatments. Harvesting intensities had to be constrained to legal requirements (not cutting more than 35% of the basal area). To reach the target basal area thresholds (expected residual basal areas), these cuts left appreciable numbers of old and senescent trees larger than the preferred residual maximum diameter (80 cm), as well as some poor quality trees among the immature classes. Even so, 56% and 55% of the trees in the high residual basal area treatments (HRBA) at Llancahue and Los Riscos, and 65% and 62% in the low residual basal area (LRBA) treatments, were of the quality class 1 at the time of the last measurement (data not shown). Overall, these experiments allowed robust comparisons between two levels of residual basal areas in stands with similar pre-cut and post-cut characteristics. However, minor differences in the climate and species composition may reflect site differences that might explain some of the differential results observed at the two sites.
Evergreen forests in south-central Chile are usually dominated by one mid-tolerant species especially dominating the basal area (E. cordifolia) and two shade-tolerant canopy species dominating the tree densities (L. philippiana and A. punctatum) [12,16,18]. This was also the case in Llancahue and Los Riscos, but in Llancahue D. winteri was the fourth most important tree species, while in Los Riscos P. lingue was the fourth most important (measured by their relative importance values). While D. winteri is well adapted to soils with high moisture throughout the year [22,39], P. lingue is well adapted to dryer conditions, especially during the summer months [22]. Earlier, Donoso [18] reported Llancahue as a better site for Evergreen forests than ones with dryer conditions or shallower soils. Based on these evidences, and the differential rainfall at the two sites, differences that we observed in the current research suggest that Llancahue is a relatively better site than Los Riscos.
General responses to selection cuttings were similar between the two sites. Mid-tolerant tree species tended to have greater diameter growth rates in the lower residual basal area than in the higher residual basal area treatment (differences were significant in some diameter classes in Llancahue), and seemed very similar for shade-tolerant species in treatments with low and high residual basal areas. Growth in basal area and volume was greater in the treatment with a high residual basal area. Tree regeneration increased during the 5/6 years that followed the selection cuttings, especially for the shade-tolerant species and the taller regeneration classes (from 50 to <200 cm). Sapling densities were similar for shade-tolerant species at both sites and basal area densities.
In spite of many common trends at both sites, Llancahue had greater stand-level growth rates (basal area and volume) and greater regeneration densities than Los Riscos, except for saplings for mid-tolerant species, where densities were greater at Los Riscos (see discussion below).
4.2. Growth in Stand Variables: Comparisons and Expectations
Growth rates in the basal area at both sites ranged (median) between 0.4 and 0.7 m2 ha−1 year−1 in the LRBA treatment, and between 0.6 and 1.0 m2 ha−1 year−1 in the treatment with HRBA. However, the basal area growth for these research sites was lower than the 1.4 m2 ha−1 year−1 simulated by Donoso [12] and Schütz et al. [2] for selection forests on relatively good sites (e.g., in the Intermediate Depression or low elevations of the Coastal range; [18]). Basal area growth for these hardwood-dominated research sites can be compared with that compiled by Forget et al. [33] for northern hardwoods in North America, who reported a range from 0.18 to 0.42 m2 ha−1 year−1 in Quebec (Canada), and from 0.38 to 0.75 m2 ha−1 year in Northeastern USA, for stands with residual basal areas between 11 and 27 m2 ha−1. Nyland [4] estimated that northern hardwoods of the USA having a mid-cycle stocking of about 20.6 m2 ha would have a basal area growth of 0.67 m2 ha−1 year−1. Thus, overall the basal area growths reported for these Chilean forests are comparable, considering that: (a) the experimental sites are in an early transformation into selection forests, and their growth will likely increase with an increase of stocking of more vigorous and higher quality trees; and (b) the first five years in a cutting cycle likely represent growth rates lower than the mean growth throughout an entire cutting cycle [4,33]. Thus, we must wait for future re-measurements to see if growth rates eventually reach that level.
Ultimately, the post-cutting growth and production will determine when stands with different residual basal areas have a sufficient added volume for a minimum operable cut, and will determine the cutting cycle length [6]. Initially, we projected waiting until the basal area reaches 55 m2 ha−1 before the next cuts for the LRBA treatment, and 70 m2 ha−1 for the HRBA treatment. That should provide an adequate volume for an operable cut at an acceptable time. With the growth rates reported here, the cutting cycles would be 25 and 17 years, respectively. However, if growth rates in these stands reach 1.0 m2 ha−1 year−1, i.e., intermediate values between the reported values in this study and those estimated by Donoso [12], cutting cycles would shorten to 15 and 10 years for the LRBA and HRBA conditions. Future evaluations of these stands will allow us to better determine appropriate cutting cycles. However, the current data indicate that the 5-year cutting cycle coupled with 35% harvesting of the basal area, as allowed in Chilean law, is unsustainable [23].
4.3. Does a Lower Residual Basal Area Promote Mid-Tolerant Species?
This question has been of interest from the onset of the experiment, and was central to our working hypothesis. Overall, during the initial 5–6 years following selection cuttings, treatment with lower residual basal areas have shown a tendency for better diameter growth and better regeneration of both the mid-tolerant and shade-tolerant species.
Our data show a greater diversity of mid-tolerant species at Llancahue than at Los Riscos, with the former site having three long-lived species in this category (E. cordifolia, D. winteri, and the conifer (Podocarpaceae) Podocarpus saligna D. Don). At Los Riscos, only E. cordifolia is a canopy species (C. paniculata (Cav.) D. Don and Gevuina avellana Molina are small-sized mid-tolerant tree species [22]). The post-cutting diameter growth for these species at both Llancahue and Los Riscos seemed greater (0.2–0.4 cm, Figure 2) than that reported for old-growth Valdivian forests (<0.2 cm; [12]), suggesting an improvement in diameter growth due to the selection cuts. However, only at Llancahue did the lower level of the residual basal area significantly improve the diameter increment for these mid-tolerant species, but only for trees in the smallest and largest diameter classes. These results are consistent with previous ones for northern hardwoods in North America. There, selection cutting resulted in better growth across all diameter classes, the highest rates for the trees with mid-range diameters, and better growth for lower residual basal areas ([40], and references therein).
The data also show favorable results for regeneration. The densities of mid-tolerant species ranged from 9.5 to 13.4 thousand seedlings per ha in Llancahue, and 3.5 to 5.5 thousand in Los Riscos. That is much higher than the <2 thousand per ha reported by Donoso and Nyland (2005) [16] for these species in unmanaged Coastal old-growth forests. Regeneration in general increased after harvesting for both shade-tolerant and mid-tolerant species, and especially for mid-tolerant species in the intermediate and taller height classes. This suggests a growth response for this group, plus recruitment from the smaller regeneration class. The data also indicate the additions of new seedlings to ensure the stocking of advance regeneration for the future. The fact that regeneration responses did not differ between residual basal area treatments suggests that the canopy still had unclosed gaps and an enhanced understory brightness through the 5–6 year post-harvest period.
Sapling densities nearly doubled those reported by Donoso and Nyland [16] for better sites in the unmanaged old-growth forests in the Coastal range. As with seedlings of intermediate and tall height classes, these increasing sapling densities reflect recruitment from taller regeneration classes, triggered by an improved growth of advance regeneration released by the selection cutting [41]. Mid-tolerant E. cordifolia ranked first in sapling density at Los Riscos, but more moderately so at Llancahue. The fact that E. cordifolia occurred at a much lower sapling density at Llancahue compared to Los Riscos may indicate the negative effect of browsing by cows and oxen from neighboring ownerships, especially upon this highly palatable species [42]. In addition to expecting an increased regeneration and growth of mid-tolerant species (e.g., E. cordifolia) in lower residual basal areas, saplings’ vigor and tolerance to browsing (measured as plant fitness and recovery after browsing) should also be enhanced by selection cutting [43]. The four-fold larger numbers of E. cordifolia sapling in the LRBA treatment compared to the HRBA treatment in Llancahue may signal this.
4.4. Prospects for Uneven-Aged Management in Valdivian Temperate Rainforests
Donoso [12] evaluated the ecological bases to sustain uneven-aged silviculture, arguing that long-term success depended on the uneven-agedness of the forests, a reverse-J or rotated-S diameter structure, and the dominance of shade-tolerant tree species. Consistent with this, when Schnabel et al. [15] examined the short-term effects of single-tree selection cutting on stand structure and tree species in the Llancahue experiment, they determined that the cut left a balanced structure with a reverse-J diameter distribution, resulted in continuous forest cover, and left sufficient small-sized trees. That suggests a potential for sustainable management by returning periodically to recreate the desired residual conditions. Furthermore, the selection cutting did not lead to significant changes in tree species richness and diversity compared to old-growth forests, but it did significantly reduce the diameter and height complexity, especially by removing an important number of large-sized trees (dbh 50 cm+, height 23 m+) [15]. Considering that we followed the same protocols at Los Riscos, we believe that these results also occurred at this site. Thus, while improving the quality and vigor of the residual stand and working toward a balanced diameter distribution are crucial for timber production objectives, they must be reconciled with the need to maintain key structural attributes of ecologic importance, such as ensuring an appropriate diameter and height diversity. Such modifications could be combined with setting aside groups of habitat trees within a landscape of stands managed via selection silviculture [15,44]. With those kinds of adjustments, findings from the current investigation of regeneration and growth responses suggest that selection cutting can adequately accommodate the concerns of Donoso [12], and serve as a useful alternative for managing Valdivian hardwood-dominated forests.
The continued monitoring of the experimental treatment, coupled with findings from other studies in Andean forests, should provide additional information about these combined effects of uneven-aged silviculture in South American temperate forests. For example, at a broader scale, the Evergreen forests in the Andean range are considered to occupy better sites than those in the Coastal range [17,45], so better growth rates could be expected in these forests. Furthermore, while the forest harvesting intensities in this study are in line with the Chilean law and with general recommendations, the monitoring of these experiments and of future experiments with different residual basal areas or harvesting intensities must signal which densities may trigger greater growth rates in more commercially valuable species, while also maintaining high degrees of diversity and complexity. These experiments should also evaluate alternative cutting cycle lengths. Additionally, new experiments should evaluate different residual structures and maximum diameters. Considering all these variables, and the fact that selection silviculture, rather than just unplanned partial cuttings, is just beginning to be practiced by some landowners, there is lots of room to continue studying the prospects for uneven-aged silviculture in these southern hemisphere hardwood-dominated temperate forests.
5. Conclusions
Prospects for the implementation of uneven-aged silviculture in hardwood-dominated Evergreen forests in Chile seem optimistic. We evaluated responses to selection cuts in two forests within the lower elevations of the Coastal range, where site productivity is considered lower than in the Andes. Since this was the first entry to these forests, we had to leave an unusually high amount of overmature trees, beyond the expected diameter for maturity. Still, the results indicated positive responses in tree regeneration and growth, although with some differences between the two sites. For example, basal area growths were similar to those found in hardwood-dominated forests in North America, where uneven-aged management is common. Further evaluations of the present experiments, plus the implementation of new studies or operational cuts with selection silviculture (hopefully at some stage with truly balanced diameter structures), will eventually illustrate the potential of this silvicultural approach for the South American temperate rainforests.
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4907/11/4/412/s1, Figure S1: Diameter distribution after harvest and 5/6 years later, according to site and treatment: a) Low residual basal area Llancahue; b) High residual basal area Llancahue; c) Low residual basal area Los Riscos; d) High residual basal area Los Riscos, Table S1: Parameters and measures of goodness of fit and prediction of height-diameter functions in Llancahue (LL). n: number of samples, Table S2: Parameters and measures of goodness of fit and prediction of height-diameter functions in Llancahue (LL). n: number of samples, Table S3: Total stem volume functions used in the study, Table S4: Mixed-effect models for the effect of site and treatment on the diameter increment, Table S5: Mixed-effect models for the effect of year, height class, site, and treatment on the regeneration density, Table S6: Effect of site and treatment in the generalized linear model (ingrowth) and the linear model (Pai in basal area and volume), Table S7: Mean and standard deviation of the regeneration density (per ha values) of seedlings and saplings according to species and treatment (HRBA and LRBA) in Llancahue (LL), Table S8: Mean and standard deviation of the regeneration density (per ha values) of seedlings and saplings according to species and treatment (HRBA and LRBA) in Los Riscos (LR).
Author Contributions
Conceptualization, P.J.D.; methodology, P.J.D.; formal analysis, P.J.D., P.F.O., and F.S.; investigation, P.J.D.; resources, P.J.D.; data curation, P.J.D., P.F.O., and F.S.; writing—original draft preparation, P.J.D., and P.F.O.; writing—review and editing, P.J.D., and R.D.N.; visualization, P.J.D.; supervision, P.J.D.; project administration, P.J.D.; funding acquisition, P.J.D. All authors have read and agreed to the published version of the manuscript.
Funding
Pablo J. Donoso thanks project FIBN-CONAF 034/2011 that provided the financial support to establish this experiment. Florian Schnabel acknowledges funding by the International Research Training Group TreeDì funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Fundation)-319936945/GRK2324.
Acknowledgments
Pablo J. Donoso and Patricio F. Ojeda thank Forestal Anchile Ltda. that harvested the experiment at Los Riscos, and provided lodging support for the final measurement.
Conflicts of Interest
The authors declare no conflict of interest.
References
- O’Hara, K. Multiaged Silviculture: Managing for Complex Forest Stands Structures; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Schütz, J.P.; Pukkala, T.; Donoso, P.; von Gadow, K. Historical Emergence and Current Application of CCF. In Continuous Cover Forestry. Book Series Managing Forest Ecosystems; Pukkala, T., Gadow, K.V., Eds.; Springer Science+Business Media B.V.: Berlin/Heidelberg, Germany, 2011; Volume 24, pp. 1–28. [Google Scholar]
- Pommerening, A.; Murphy, S.T. A review of the history, definitions and methods of continuous cover forestry with special attention to afforestation and restocking. Forestry 2004, 77, 27–44. [Google Scholar] [CrossRef]
- Nyland, R.D. Silviculture Concepts and Applications, 3rd ed.; Waveland Press, Inc.: Long Grove, IL, USA, 2016. [Google Scholar]
- Knapp, S.P.; Webster, C.R.; Kern, C.C. Can group selection with legacy retention change compositional trajectories in conventionally managed hardwoods? For. Ecol. Manag. 2019, 448, 174–186. [Google Scholar] [CrossRef]
- Baker, J.B.; Cain, M.D.; Guldin, J.M.; Murphy, P.A.; Shelton, M.G. Uneven-Aged silviculture for the loblolly and shortleaf pine forests cover types. US For. Serv. Gen. Tech. Rep. 1996. [Google Scholar] [CrossRef]
- Prévost, M.; Charette, L. Selection cutting in a yellow birch–Conifer stand, in Quebec, Canada: Comparing the single-Tree and two hybrid methods using different sizes of canopy opening. For. Ecol. Manag. 2015, 357, 195–205. [Google Scholar] [CrossRef]
- Escandón, A.B.; Paula, S.; Rojas, R.; Corcuera, L.J.; Coopman, R.E. Sprouting extends the regeneration niche in temperate rain forests: The case of the long-Lived tree Eucryphia cordifolia. For. Ecol. Manag. 2013, 310, 321–326. [Google Scholar] [CrossRef]
- Tecklin, D.; DellaSala, D.A.; Luebert, F.; Pliscoff, P. Valdivian temperate forests of Chile and Argentina. In Temperate and Boreal Rainforests of the World: Ecology and Conservation; Della, S., Ed.; Island Press: Washington, DC, USA, 2011; pp. 132–153. [Google Scholar]
- Donoso, P.J.; Ponce, D.B.; Pinto, J.B.; Triviño, I.L. Cobertura y regeneración arbórea en bosques siempreverdes en diferentes estados sucesionales en el sitio experimental de Llancahue, cordillera de la costa de Valdivia, centro sur de Chile. Gayana Bot. 2018, 75, 657–662. [Google Scholar] [CrossRef]
- Ponce, D.B.; Donoso, P.J.; Salas-Eljatib, C. Differentiating Structural and Compositional Attributes across Successional Stages in Chilean Temperate Rainforests. Forests 2017, 8, 329. [Google Scholar] [CrossRef]
- Donoso, P.J. Structure and Growth in Coastal Evergreen Forests as the Bases for Uneven-Aged Silviculture in Chile. Ph.D. Thesis, College of Environmental Science and Forestry, State University of New York (SUNY-ESF), Syracuse, NY, USA, 2002; p. 178. [Google Scholar]
- Donoso, C.; Donoso, P.J.; González, M.; Sandoval, V. Los Bosques Siempreverdes; Donoso, C., Lara, A., Eds.; Silvicultura de los Bosques Nativos de Chile. Ed. Universitaria: Santiago, Chile, 1998; pp. 297–339. [Google Scholar]
- Donoso, C. Antecedentes básicos para la silvicultura del tipo forestal siempreverde. Bosque 1989, 10, 37–53. [Google Scholar] [CrossRef]
- Schnabel, F.; Donoso, P.J.; Winter, C. Short-Term effects of single-Tree selection cutting on stand structure and tree species composition in Valdivian rainforests of Chile. N. Z. J. For. Sc. 2017, 47, 21. [Google Scholar] [CrossRef]
- Donoso, P.J.; Nyland, R.D. Seeding density according to structure, dominance and understory cover in old-Growth forest stands of the evergreen forest type in the coastal range of Chile. Rev. Chil. de Hist. Nat. 2005, 78, 51–63. [Google Scholar] [CrossRef]
- Donoso, C. Regeneración y crecimiento en el tipo forestal Siempreverde costero y andino tras distintos tratamientos silviculturales. Bosque 1989, 10, 69–83. [Google Scholar] [CrossRef]
- Donoso, P.J. Crown Index: A canopy balance indicator to assess growth and regeneration in uneven-Aged forest stands of the Coastal Range of Chile. Forestry 2005, 78, 337–351. [Google Scholar] [CrossRef]
- Ponce, D.B.; Donoso, P.J.; Salas-Eljatib, C. Índice de bosque adulto: Una herramienta para evaluar estados de desarrollo de bosques nativos de tierras bajas del centro-sur de Chile. Bosque 2019, 40, 235–240. [Google Scholar] [CrossRef]
- Veblen, T.T. Forest development in tree-Fall gaps in the temperate rain forests of Chile. Natl. Geogr. Res. 1985, 1, 162–183. [Google Scholar]
- Veblen, T.T. Regeneration Dynamics. In Plant Succession: Theory and Prediction; Glenn-Lewin, C., Peet, R.K., Veblen, T.T., Eds.; Chapman and Hall: London, UK, 1992; pp. 153–187. [Google Scholar]
- Donoso, C. Las especies arbóreas de los bosques templados de Chile y Argentina; Autoecología. Marisa Cuneo ediciones: Valdivia, Chile, 2006; p. 678. [Google Scholar]
- Donoso, P.J. Necesidades, opciones y futuro del manejo multietáneo en Chile. In Silvicultura en Bosques Nativos. Avances en la investigación en Chile, Argentina y Nueva Zelandia. Estudios en Silvicultura de Bosques Nativos; Donoso, P.J., Promis, A., Eds.; Marisa Cuneo: Valdivia, Chile, 2013; Volume 1, pp. 55–85. [Google Scholar]
- Di Castri, F.; Hajek, E. Bioclimatología de Chile; Editorial Universidad Católica de Chile: Santiago, Chile, 1976; pp. 11–12. Available online: https://sites.google.com/site/alvaropromis/Home/libro-silvicultura-bosques-nativos (accessed on 14 June 2019).
- Oyarzún, C.; Nahuelhual, L.; Núñez, D. Los servicios ecosistémicos del bosque templado lluvioso: Producción de agua y su valoración económica. Rev. Ambiente Desarro. 2005, 20, 88–95. [Google Scholar]
- Luebert, F.; Pliscoff, P. Bioclimas de la Cordillera de la Costa del centro-sur de Chile. In Historia, biodiversidad y ecología de los bosques costeros de Chile; Smith-Ramírez, C., Armesto, J.Y., Valdovinos, C., Eds.; Editorial Universitaria: Santiago, Chile, 2005; pp. 60–73. [Google Scholar]
- Donoso, P.J.; Frene, C.; Flores, M.; Moorman, M.; Oyarzún, C.; Zavaleta, J. Balancing water supply and old-Growth forest conservation in the lowlands of south-Central Chile through an adaptive co-Management approach. Landsc. Ecol. 2014, 29, 245–260. [Google Scholar] [CrossRef]
- Centro de Información de Recursos Naturales (CIREN). Estudio Agrológico de la X Región; CIREN: Santiago, Chile, 2003. [Google Scholar]
- Pincheira, J.; Rau, J.; Smith, C. Diversidad de plantas trepadoras y epífitas vasculares en un paisaje agroforestal del sur de Chile: Una Comparación entre fragmentos de bosque nativo. Boletín de la Sociedad Argentina de Botánica 2012, 47, 411–426. [Google Scholar]
- Guldin, J.M. Uneven-Aged BDq regulation of Sierra Nevada mixed conifers. West. J. Appl. For. 1991, 6, 27–32. [Google Scholar] [CrossRef]
- Bacardit, P. Evaluación del daño provocado por cortas de selección sobre los árboles residuales en un bosque del Tipo Forestal Siempreverde en Llancahue, provincia de Valdivia. Bachelor’s Thesis, Universidad Austral de Chile, Valdivia, Chile, 2014; p. 56. [Google Scholar]
- Hansen, G.D.; Nyland, R.D. Effects of diameter distribution on the growth of simulated uneven-Aged sugar maple stands. Can. J. For. Res. 1986, 17, 1–8. [Google Scholar] [CrossRef]
- Forget, E.; Nolet, P.; Doyon, F.; Delagrange, S.; Jardon, Y. Ten-Year response of northern hardwood stands to commercial selection cutting in southern Quebec, Canada. For. Ecol. Manag. 2007, 242, 764–775. [Google Scholar] [CrossRef]
- Gutiérrez, A.G.; Armesto, J.J.; Aravena, J.C.; Carmona, M.; Carrasco, N.V.; Christie, D.A.; Peña, M.P.; Pérez, C.; Huth, A. Structural and environmental characterization of old-Growth temperate rainforests of northern Chiloe Island, Chile: Regional and global relevance. For. Ecol. Manag. 2009, 258, 376–388. [Google Scholar] [CrossRef]
- Stage, A.R. A mathematical approach to polymorphic site index curves for grand fir. For. Sci. 1963, 9, 167–180. [Google Scholar]
- Zuur, A.; Ieno, E.; Walker, N.; Saveliev, A.; Smith, G. Mixed Effects Models and Extensions in Ecology with R. North Carolina, USA; Springer: Berlin/Heidelberg, Germany, 2009; p. 589. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2017. Available online: http://www.R-project.org (accessed on 16 December 2019).
- Pinheiro, J.; Bates, D.; Debroy, S.; Sarkar, D.F.; R Core Team. Nlme: Linear and nonlinear mixed effects models. R Package Version 2017, 3, 1–131. [Google Scholar]
- Donoso, P.J.; Soto, D.P.; Bertín, R.A. Size–Density relationships in Drimys winteri secondary forests of the Chiloe Island, Chile: Effects of physiography and species composition. For. Ecol. Manag. 2007, 239, 120–127. [Google Scholar] [CrossRef]
- Kiernan, D.H.; Bevilacqua, E.; Nyland, R.D. Individual-Tree diameter growth model for sugar maple trees in uneven-Aged northern hardwood stands under selection system. For. Ecol. Manag. 2009, 256, 1579–1586. [Google Scholar] [CrossRef]
- Donoso, P.J.; Nyland, R.D.; Zhang, L. Growth of saplings after selection cutting in Northern Hardwood. North. J. Appl. For. 2000, 17, 149–152. [Google Scholar] [CrossRef]
- Zamorano-Elgueta, C.; Cayuela, L.; Rey-Benayas, J.M.; Donoso, P.J.; Geneletti, D.; Hobbs, R. The differential influences of human-Induced disturbances on tree regeneration community: A landscape approach. Ecosphere 2014. [Google Scholar] [CrossRef]
- Kupferschmid, A.D.; Bugmann, H. Timing, light availability and vigour determine the response of Abies alba saplings to leader shoot browsing. Eur. J. For. Res. 2013, 132, 47–60. [Google Scholar] [CrossRef]
- Gustafsson, L.; Bauhus, J.; Asbeck, T.; Derci Augustynczik, A.L.; Basile, M.; Frey, J.; Gutzat, F.; Hanewinkel, M.; Helbach, J.; Jonker, M.; et al. Retention as an integrated biodiversity conservation approach for continuous-Cover forestry in Europe. Ambio 2019. [Google Scholar] [CrossRef]
- Donoso, P.J.; Soto, D.P. Does site quality affect the additive basal area phenomenon? Results from Chilean old-Growth temperate rainforests. Can. J. For. Res. 2016, 46, 1330–1336. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).