Early Regeneration Dynamics of Pure Black Spruce and Aspen Forests after Wildfire in Boreal Alberta, Canada



Abstract
1. Introduction
2. Materials and Methods
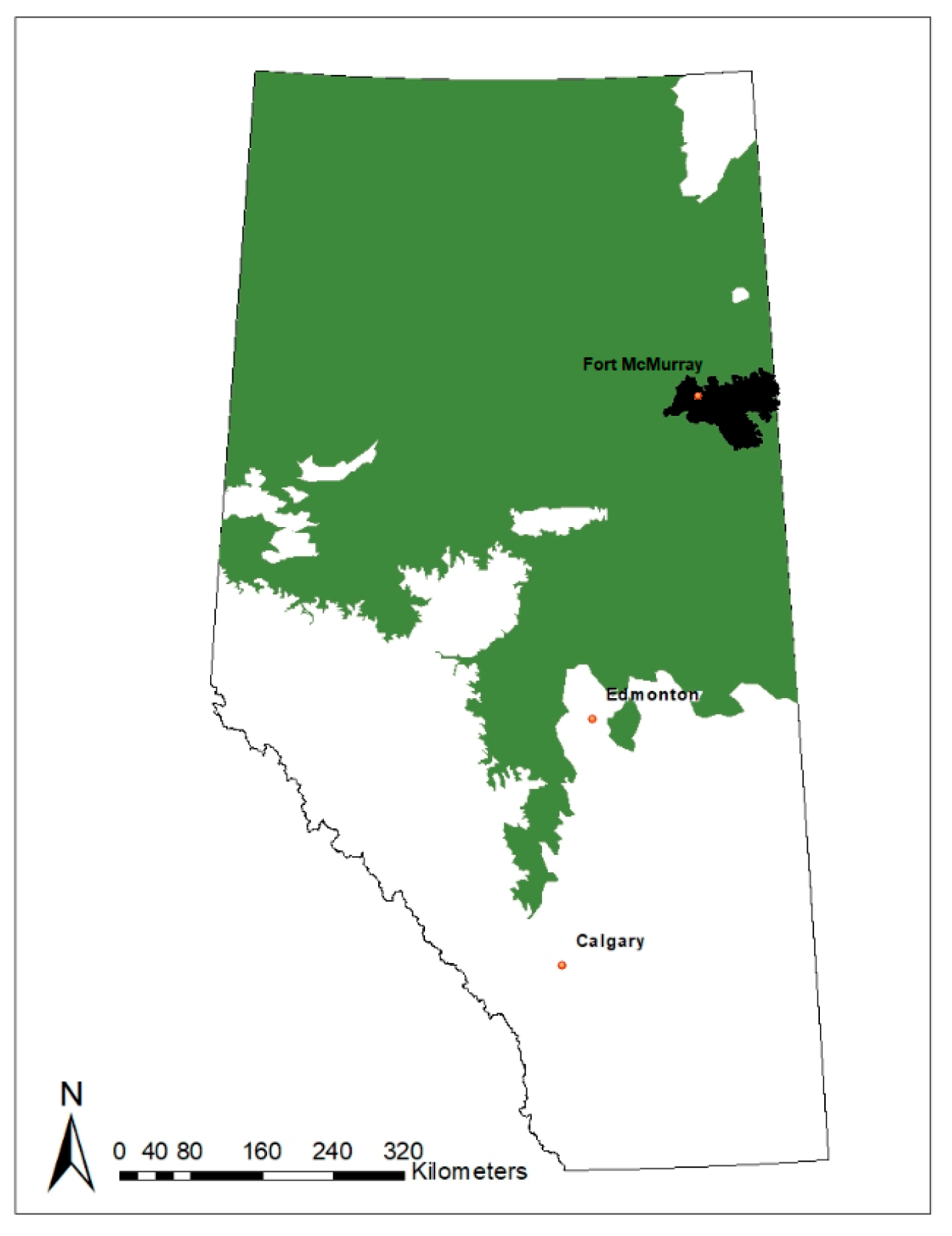
2.1. Study Area
2.2. Field and Lab Methods
2.3. Statistical Analyses
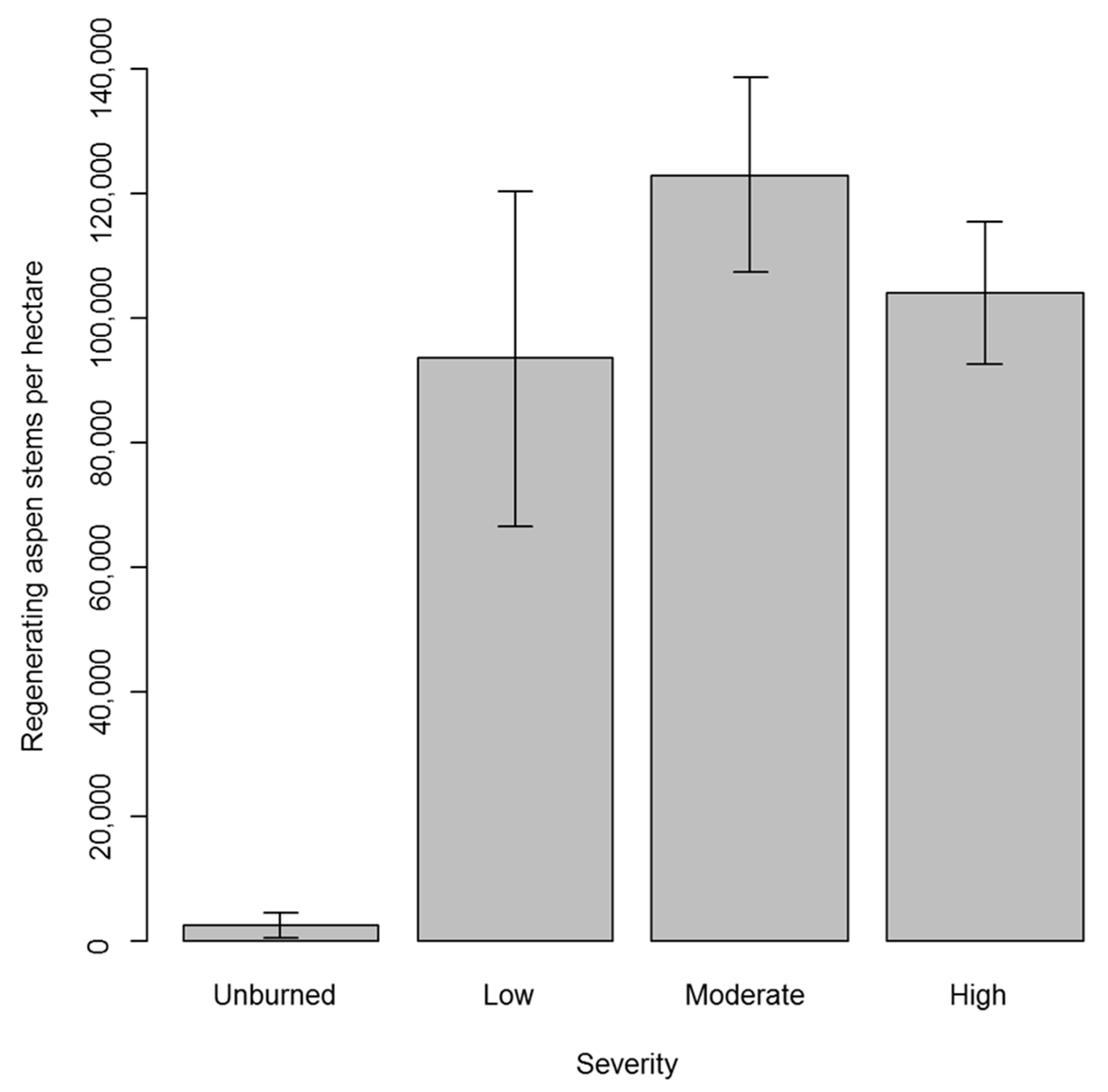
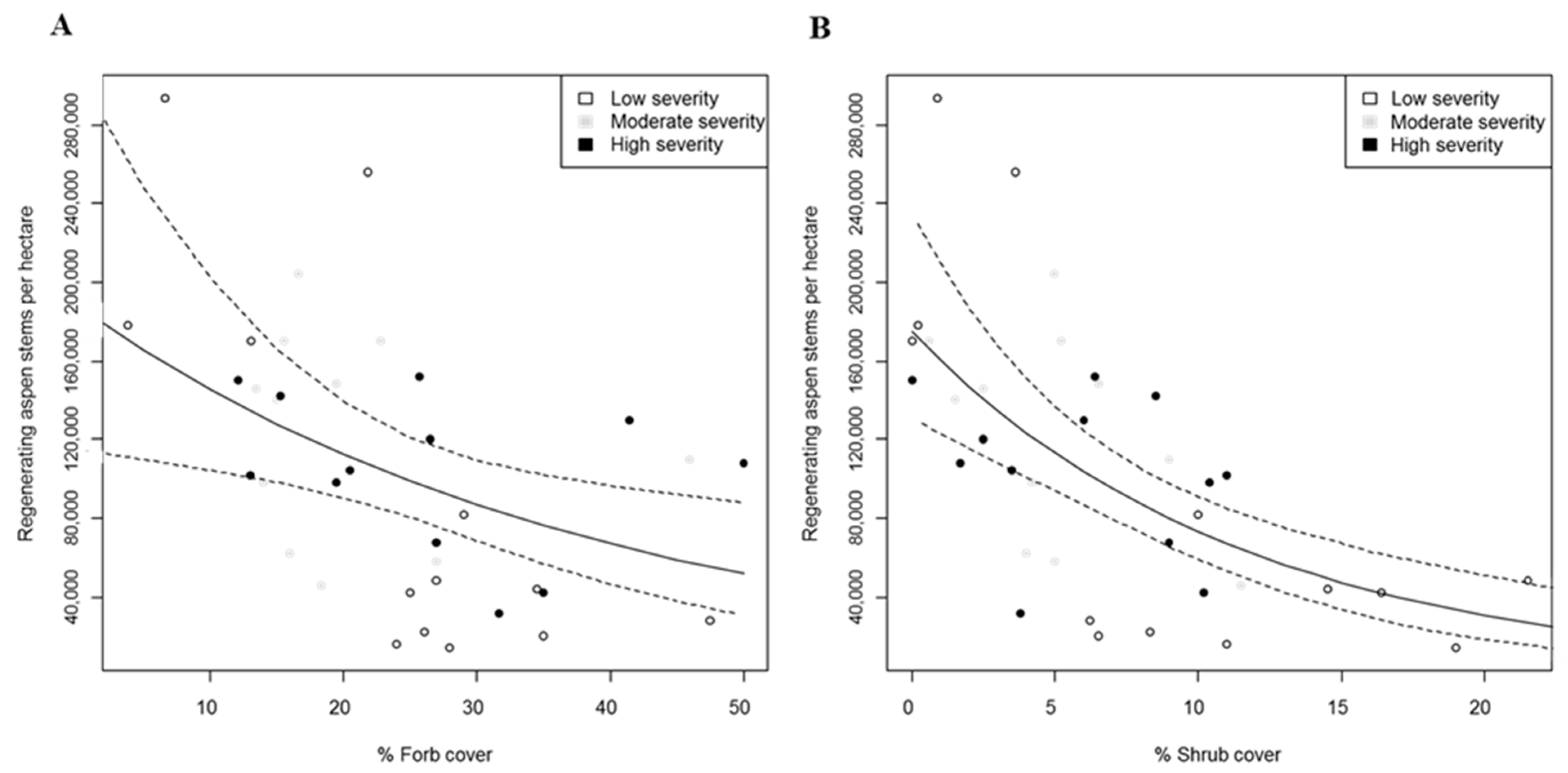
3. Results
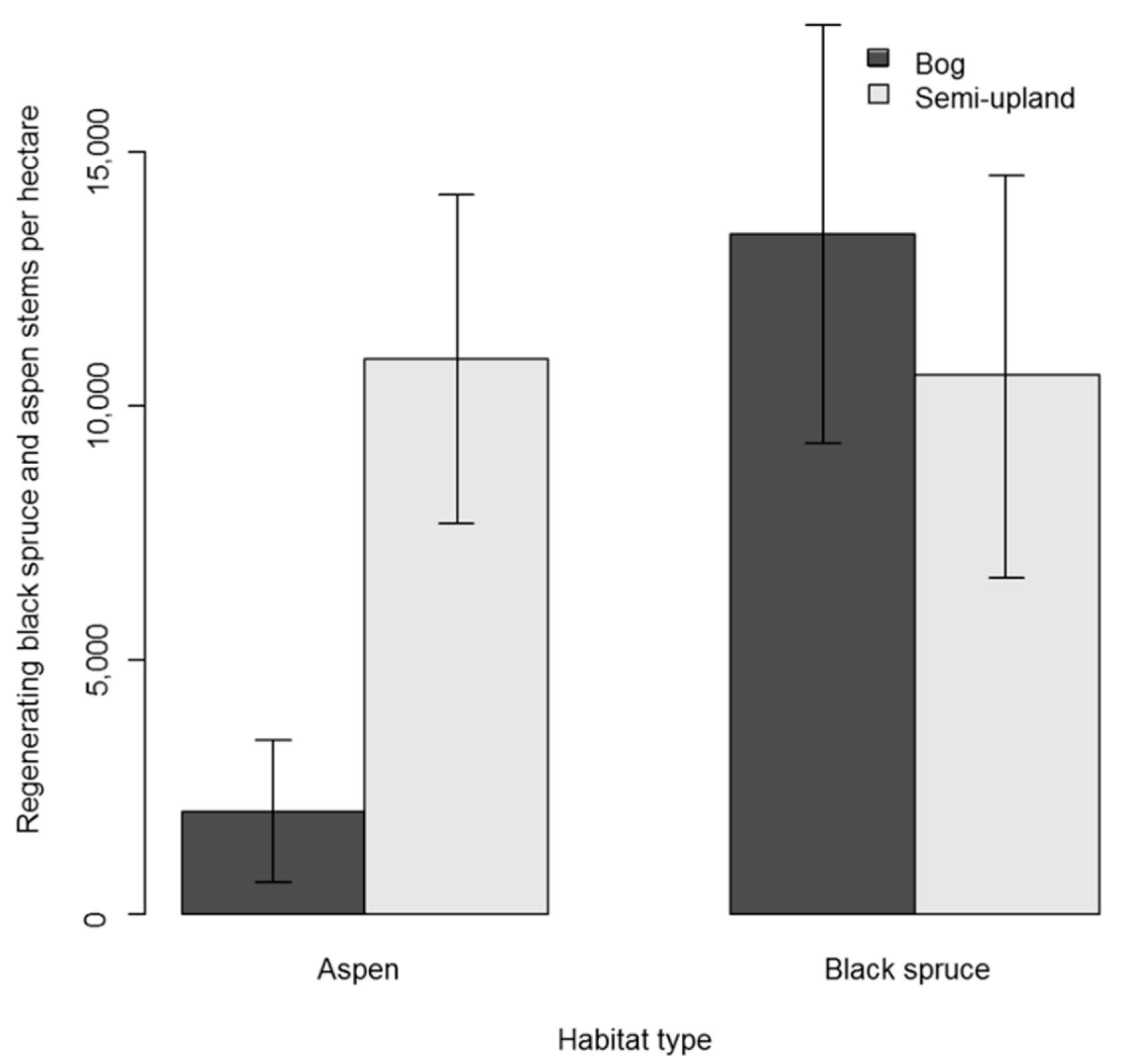
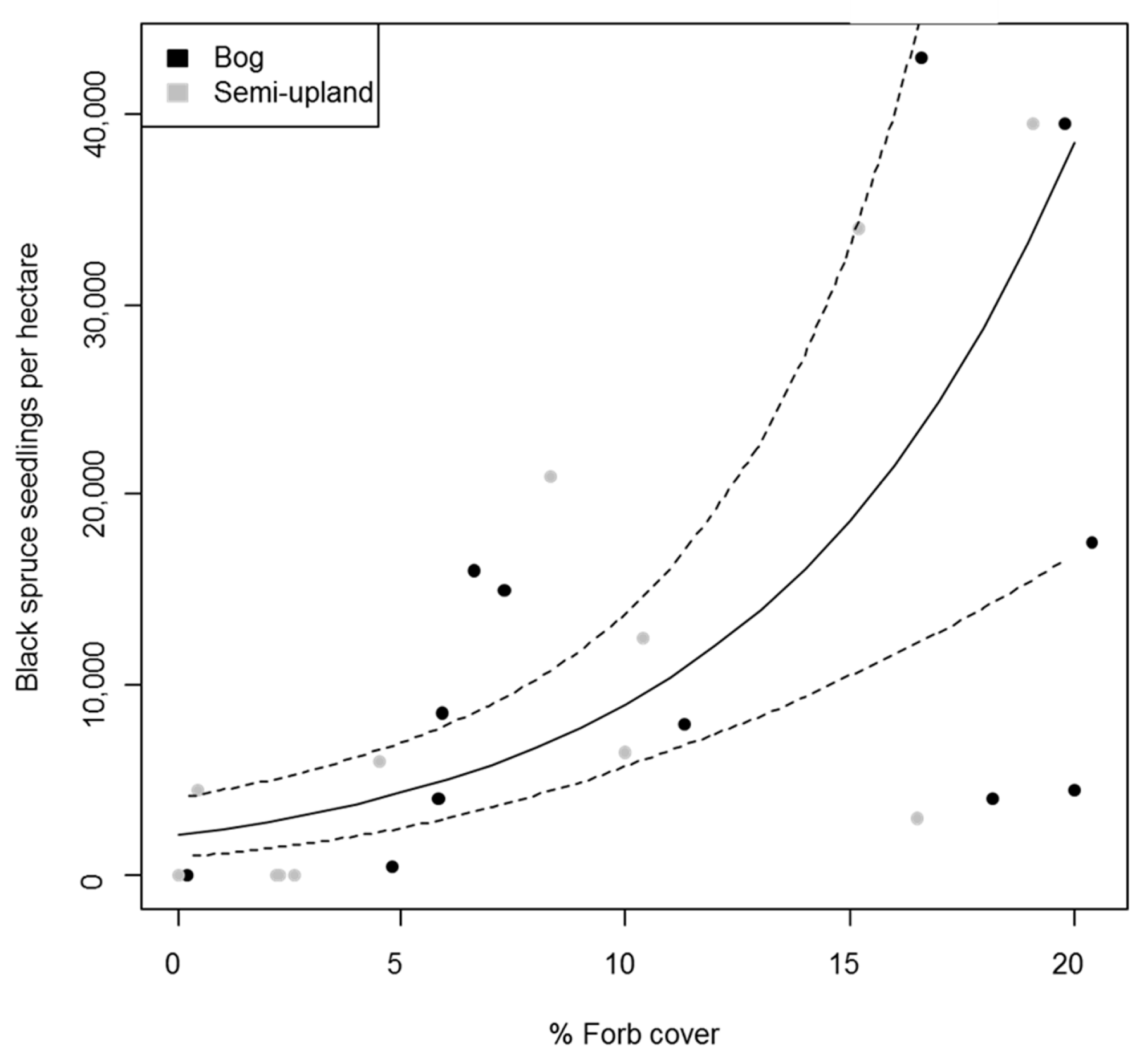
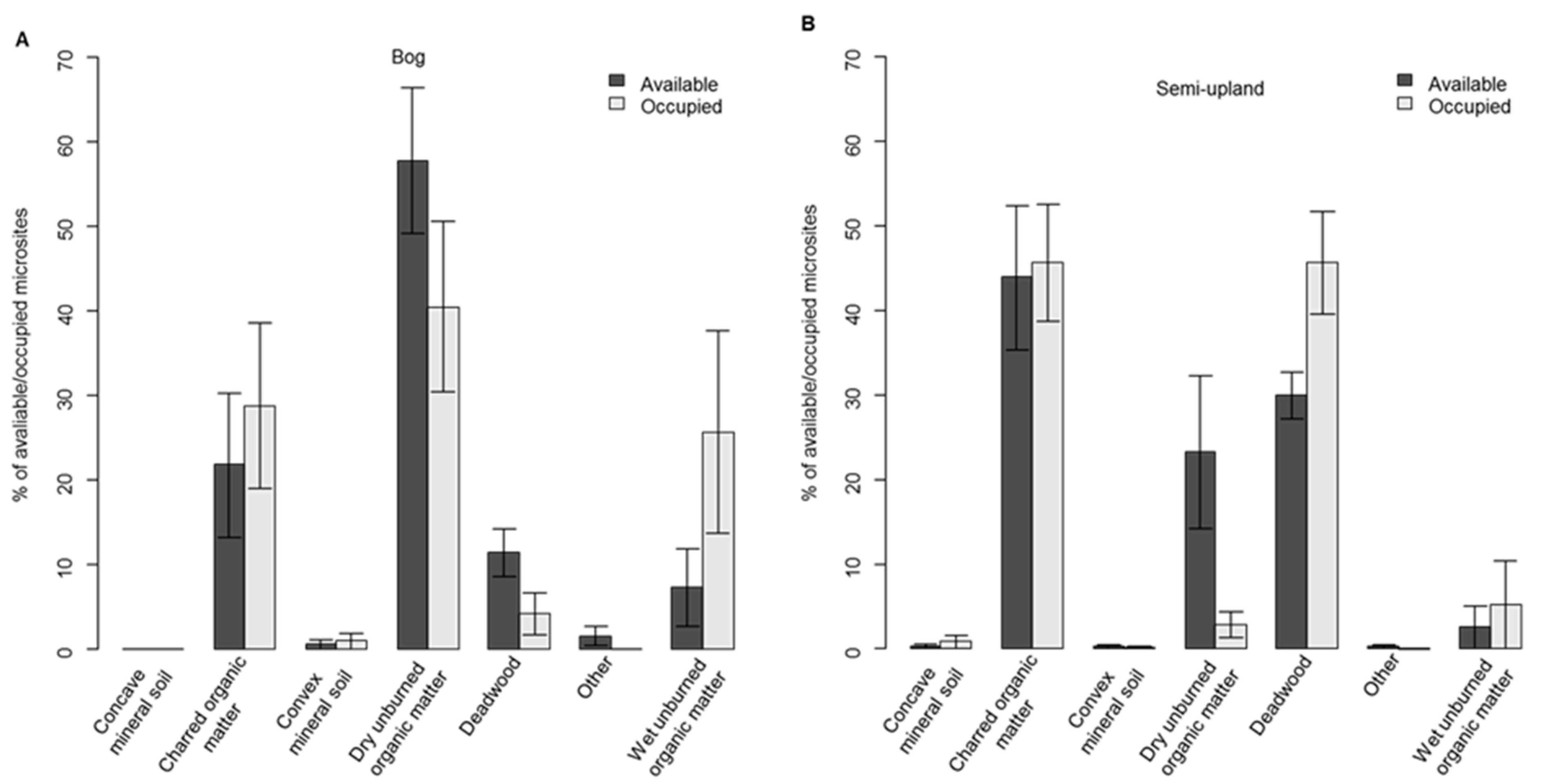
3.1. Regeneration in Black Spruce Stands
3.2. Regeneration in Aspen Stands
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brandt, J.P.; Flannigan, M.D.; Maynard, D.G.; Thompson, I.D.; Volney, W.J.A. An introduction to Canada’s boreal zone: Ecosystem processes, health, sustainability, and environmental issues. Environ. Rev. 2013, 21, 207–226. [Google Scholar] [CrossRef]
- Weber, M.G.; Flannigan, M.D. Canadian boreal forest ecosystem structure and function in a changing climate: Impacts on fire regimes. Environ. Rev. 1997, 5, 145–166. [Google Scholar] [CrossRef]
- Stocks, B.J.; Mason, J.A.; Todd, J.B.; Bosch, E.M.; Wotton, B.M.; Amiro, B.D.; Flannigan, M.D.; Hirsch, K.G.; Logan, K.A.; Martell, D.L.; et al. Large forest fires in Canada, 1959–1997. J. Geophys. Res. 2002, 107, 1–12. [Google Scholar] [CrossRef]
- De Groot, W.J.; Flannigan, M.D.; Cantin, A.S. Climate change impacts on future boreal fire regimes. For. Ecol. Manag. 2012, 294, 35–44. [Google Scholar] [CrossRef]
- Greene, D.F.; Zasada, J.C.; Sirois, L.; Kneeshaw, D.; Morin, H.; Charron, I.; Simard, M.J. A review of the regeneration dynamics of North American boreal forest tree species. Can. J. For. Res. 1999, 29, 824–839. [Google Scholar] [CrossRef]
- Thomas, P.A.; Wein, R.W. Delayed emergence of four conifer species on postfire seedbeds in eastern Canada. Can. J. For. Res. 1985, 15, 727–729. [Google Scholar] [CrossRef]
- Foote, J.M. Classification, Description, and Dynamics of Plant Communities after Fire in the Taiga of Interior Alaska; Research Paper PNW-RP-307; Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1983. [Google Scholar]
- Johnstone, J.F.; Chapin, F.S., III. Fire interval effects on successional trajectory in boreal forests of northwest Canada. Ecosystems 2006, 9, 268–277. [Google Scholar] [CrossRef]
- Horton, K.W.; Lees, J.C. Black Spruce in the Foothills of Alberta; Note 110; Canadian Department of Forestry, Forest Research Branch: Ottawa, ON, Canada, 1961. [Google Scholar]
- Lloyd, A.H.; Fastie, C.L.; Eisen, H. Fire and substrate interact to control the northern range limit of black spruce (Picea mariana) in Alaska. Can. J. For. Res. 2007, 37, 2480–2493. [Google Scholar] [CrossRef]
- Black, R.A.; Bliss, L.C. Reproductive ecology of Picea mariana (Mill.) BSP, at tree line near Inuvik, Northwest Territories, Canada. Ecol. Monogr. 1980, 50, 331–354. [Google Scholar] [CrossRef]
- Auclair, A.N.D. The role of fire in lichen-dominated tundra and forest-tundra. In The Role of Fire in Northern Circumpolar Ecosystems; Wein, R.W., MacLean, D.A., Eds.; John Wiley & Sons: New York, NY, USA, 1983; pp. 235–256. [Google Scholar]
- Kemball, K.J.; Wang, G.G.; Westwood, A.R. Are mineral soils exposed by severe wildfire better seedbeds for conifer regeneration? Can. J. For. Res. 2006, 36, 1943–1950. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Hollingsworth, T.N.; Chapin, F.S., III. A Key for Predicting Postfire Successional Trajectories in Black Spruce Stands of Interior Alaska; General Technical Report PNW-GTR-767; United States Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2008.
- Mallik, A.; Kayes, I. Lichen mated seedbeds inhibit while moss dominated seedbeds facilitate black spruce (Picea mariana) seedling regeneration in post-fire boreal forest. For. Ecol. Manag. 2018, 427, 260–274. [Google Scholar] [CrossRef]
- Wang, G.G. Early regeneration and growth dynamics of Populus tremuloides suckers in relation to fire severity. Can. J. For. Res. 2003, 33, 1998–2006. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Deshaies, D.; Lieffers, V.J. Disturbance facilitates rapid range expansion of aspen into higher elevations of the Rocky Mountains under a warming climate. J. Biogeogr. 2010, 37, 68–76. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Pinno, B.D.; Mock, K.E. Tamm Review: Seedling-based ecology, management, and restoration in aspen (Populus tremuloides). For. Ecol. Manag. 2019, 432, 231–245. [Google Scholar] [CrossRef]
- Yih, K.; Boucher, D.H.; Vandermeer, J.H.; Zamora, N. Recovery of the rain forest of Southeastern Nicaragua after destruction by Hurricane Joan. Biotropica 1991, 23, 106–113. [Google Scholar] [CrossRef]
- Bergeron, Y.; Chen, H.Y.H.; Kenkel, N.C.; Leduc, A.L.; Macdonald, S.E. Boreal mixedwood stand dynamics: Ecological processes underlying multiple pathways. For. Chron. 2014, 90, 202–213. [Google Scholar] [CrossRef]
- Peters, V.S.; Macdonald, S.E.; Dale, M.R.T. The interaction between masting and fire is key to white spruce regeneration. Ecology 2005, 86, 1744–1750. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Hollingsworth, T.N.; Chapin, F.S., III; Mack, M.C. Changes in fire regime break the legacy lock on successional trajectories in Alaskan boreal forests. Glob. Chang. Biol. 2009, 16, 1–15. [Google Scholar] [CrossRef]
- Wan, H.Y.; Olson, A.C.; Muncey, K.D.; Clair, S.B.S. Legacy effects of fire size and severity on forest regeneration, recruitment, and wildlife activity in aspen forests. For. Ecol. Manag. 2014, 329, 59–68. [Google Scholar] [CrossRef]
- Shinneman, D.J.; Krasnow, K.D.; McIlroy, S.K. The Role of Fire in Aspen Ecology and Restoration; WAA Briefs# 3; Utah State University: Logan, UT, USA, 2015. [Google Scholar]
- Natural Resources Canada. The State of Canada’s Forests—Annual Report 2017; Government of Canada: Ottawa, ON, Canada, 2017.
- Kochtubajda, B.; Brimelow, J.; Flannigan, M.; Morrow, B.; Greenhough, M.D. The extreme 2016 wildfire in Fort McMurray, Alberta, Canada. Bull. Am. Meteorol. Soc. 2017, 7, 176–177. [Google Scholar]
- Downing, D.J.; Pettapiece, W.W. Natural Regions and Subregions of Alberta; No. T/852; Government of Alberta Publish: Edmonton, AB, Canada, 2006; 264p.
- Soil Classification Working Group. The Canadian System of Soil Classification, 3rd ed.; Agriculture and Agr-Food Canada, Publication 1646; NRC Research Press: Ottawa, ON, Canada, 1998; 187p. [Google Scholar]
- Key, C.H.; Benson, N.C. Landscape assessment: Ground measure of severity, the composite burn index; and remote sensing of severity, the Normalized Burn Ratio. In FIREMON: Fire Effects Monitoring and Inventory System; Lutes, D.C., Keane, R.E., Caratti, J.F., Key, C.H., Benson, N.C., Sutherland, S., Gangi, L.J., Eds.; RMRS-GTR-164-CD; USDA Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2006; pp. 1–51. [Google Scholar]
- Hendershot, W.H.; Lalande, H.; Duquette, M. Soil reaction and exchangeable acidity. In Soil Sampling and Methods of Analysis, 2nd ed.; Carter, M.R., Gregorich, E.G., Eds.; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Harrell, F.E. Rms: Regression Modelling Strategies; R Package Version 5.1-1; Springer: Heidelberg, Germany, 2017. [Google Scholar]
- Chrosciewicz, Z. Burning for black spruce regeneration on a lowland cutover site in southeastern Manitoba. Can. J. For. Res. 1976, 6, 179–186. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Chapin, F.S., III; Foote, J.; Kemmett, S.; Price, K.; Viereck, L. Decadal observations of tree regeneration following fire in boreal forests. Can. J. For. Res. 2004, 34, 267–273. [Google Scholar] [CrossRef]
- Greene, D.F.; Kneeshaw, D.D.; Messier, C.; Lieffers, V.; Cormier, D.; Doucet, R.; Coats, K.D.; Groot, A.; Grover, G.; Calogeropoulos, C. Modelling silvicultural alternatives for conifer regeneration in boreal mixedwood stands (aspen/white spruce/balsam fir). For. Chron. 2002, 78, 281–295. [Google Scholar] [CrossRef]
- Lacey, M.E.; Dech, J.P. Comparison of black spruce (Picea mariana) radial growth reduction in different soil moisture regimes during a spruce budworm (Choristoneura fumiferana) outbreak. Can. J. For. Res. 2012, 42, 1410–1419. [Google Scholar] [CrossRef]
- Bubier, J.L. Patterns of Picea mariana (black spruce) growth and raised bog development in Victory Basin, Vermont. Bull. Torrey Bot. Club 1991, 118, 399–411. [Google Scholar] [CrossRef]
- Crawford, R.M.M. Root survival in flooded soils. In Mires: Swamp, Bog, Fen, and Moor; Gore, A.J.P., Ed.; Elsevier: Amsterdam, The Netherlands, 1983; Volume 4A. [Google Scholar]
- Lieffers, V.J.; Rothwell, R.L. Rooting of peatland black spruce and tamarack in relation to depth of water table. Can. J. Bot. 1987, 65, 817–821. [Google Scholar] [CrossRef]
- Viglas, J.N.; Brown, C.D.; Johnstone, J.F. Age and size effects on seed productivity of northern black spruce. Can. J. For. Res. 2013, 43, 534–543. [Google Scholar] [CrossRef]
- Wheeler, J.A.; Hermanutz, L.; Marino, P.M. Feathermoss seedbeds facilitate black spruce seedling recruitment in the forest-tundra ecotone (Labrador, Canada). Oikos 2011, 120, 1236–1271. [Google Scholar] [CrossRef]
- Lambers, H.; Atkin, O.K.; Millenaar, F.F. Respiratory patterns in roots in relation to their functioning. In Plant Roots, the Hidden Half, 3rd ed.; Waisel, Y., Eshel, A., Kafkafi, U., Eds.; Marcel Dekker: New York, NY, USA, 2002; pp. 521–552. [Google Scholar]
- Hély, C.; Bergeron, Y.; Flannigan, M.D. Effects of stand composition on fire hazard in mixed-wood Canadian boreal forest. J. Veg. Sci. 2000, 11, 813–824. [Google Scholar] [CrossRef]
- Brown, J.K.; Bebyle, N.V. Fire damage, mortality, and suckering in aspen. Can. J. For. Res. 1987, 17, 1100–1109. [Google Scholar] [CrossRef]
- Hogg, E.H.; Lieffers, V.J. The impact of Calamagrostis canadensis on soil thermal regimes aftere logging in northern Alberta. Can. J. For. Res. 1991, 21, 387–394. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Lieffers, V.J. Growth of Populus tremuloides in association with Calamagrostis canadensis. Can. J. For. Res. 1998, 28, 396–401. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Type | Basal Area | Height | Forb | Shrub | Bryophyte | Clay | Sand | pH | Electrical Conductivity (EC) |
|---|---|---|---|---|---|---|---|---|---|
| (m2 ha−1) | (m) | (%) | (%) | (%) | (%) | (%) | (µS cm−1) | ||
| Black spruce—Bog | 6.7 (1.2) | 11.1 (0.9) | 11 (2) | 8 (1.8) | 3 (1.0) | - | - | 5.0 (0.3) | 140 (30.5) |
| Black spruce—Semi-upland | 13.9 (1.3) | 14.4 (0.7) | 8 (2) | 2 (0.9) | 11 (4.7) | 24.4 (1.9) | 36.8 (4.0) | 5.2 (0.1) | 88 (11.9) |
| Black spruce—Unburned bog | 5.3 (1.7) | 7.7 (2.3) | 14 (4) | 20 (6.5) | 86 (2.5) | - | - | 5.2 (0.2) | 185 (41.2) |
| Black spruce—Unburned semi-upland | 25.1 (5.8) | 15.8 (1.2) | 3 (1) | 10 (3.5) | 85 (5.1) | 14.1 (1.0) | 53.3 (1.6) | 4.4 (0.2) | 165 (46.3) |
| Aspen—Low | 32.7 (3.5) | 22.7 (1.1) | 224 (3) | 9 (2.0) | 2 (0.8) | 31.5 (2.5) | 16.0 (3.2) | 6.0 (0.1) | 124 (14.2) |
| Aspen—Moderate | 40.3 (10.2) | 21.8 (1.0) | 20 (3) | 5 (0.9) | 0 | 27.3 (2.2) | 19.2 (3.6) | 5.9 (0.1) | 84 (5.2) |
| Aspen—High | 27.3 (2.7) | 21.5 (1.0) | 26 (3) | 6 (0.9) | 1 (0.3) | 26.5 (2.2) | 23.0 (3.6) | 5.9 (0.1) | 106 (5.2) |
| Aspen—Unburned | 22.9 (4.5) | 21.9 (1.1) | 20 (3) | 10 (4.2) | 1 (1.3) | 18.1 (6.1) | 56.4 (10.8) | 5.6 (0.3) | 84 (9.7) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jean, S.A.; Pinno, B.D.; Nielsen, S.E. Early Regeneration Dynamics of Pure Black Spruce and Aspen Forests after Wildfire in Boreal Alberta, Canada. Forests 2020, 11, 333. https://doi.org/10.3390/f11030333
Jean SA, Pinno BD, Nielsen SE. Early Regeneration Dynamics of Pure Black Spruce and Aspen Forests after Wildfire in Boreal Alberta, Canada. Forests. 2020; 11(3):333. https://doi.org/10.3390/f11030333
Chicago/Turabian StyleJean, Stephanie A., Bradley D. Pinno, and Scott E. Nielsen. 2020. "Early Regeneration Dynamics of Pure Black Spruce and Aspen Forests after Wildfire in Boreal Alberta, Canada" Forests 11, no. 3: 333. https://doi.org/10.3390/f11030333
APA StyleJean, S. A., Pinno, B. D., & Nielsen, S. E. (2020). Early Regeneration Dynamics of Pure Black Spruce and Aspen Forests after Wildfire in Boreal Alberta, Canada. Forests, 11(3), 333. https://doi.org/10.3390/f11030333