Response of Soil CO2 Efflux to Shelterwood Harvesting in a Mature Temperate Pine Forest


Abstract
1. Introduction
2. Methods
2.1. Study Site
2.2. Shelterwood Harvest Treatment
2.3. Soil CO2 Efflux Measurements
2.4. Data Processing and Quality Control of Soil Flux Data
2.5. Eddy Covariance Flux and Ancillary Data
2.6. Statistical Analysis of Pre- and Post-Harvesting
3. Results
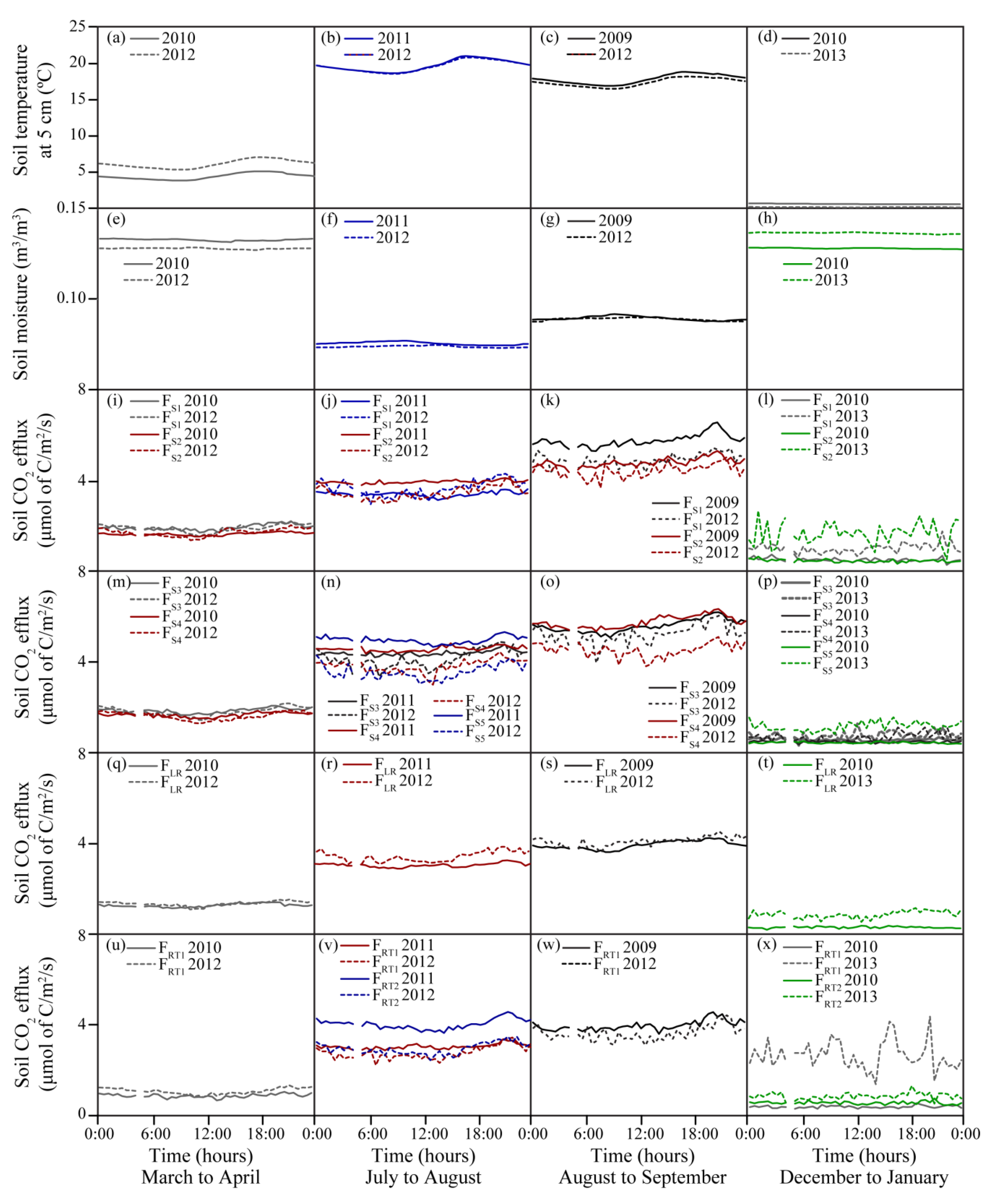
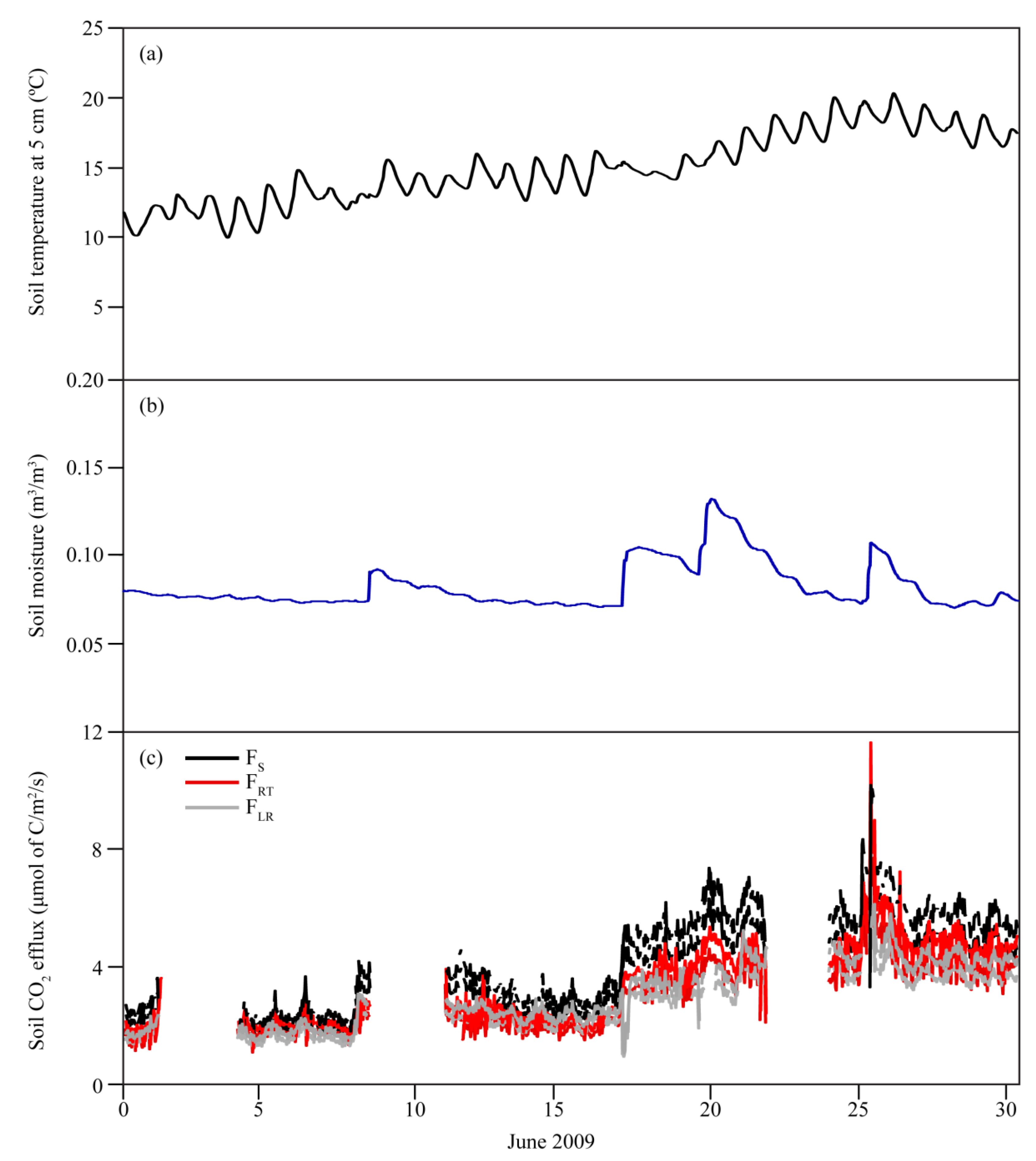
3.1. Seasonal Variations in Soil Temperature and Soil Moisture
3.2. Seasonal Variability of Soil CO2 Efflux
3.3. Comparison of Soil Temperature and Soil Moisture Pre- and Post-Harvesting
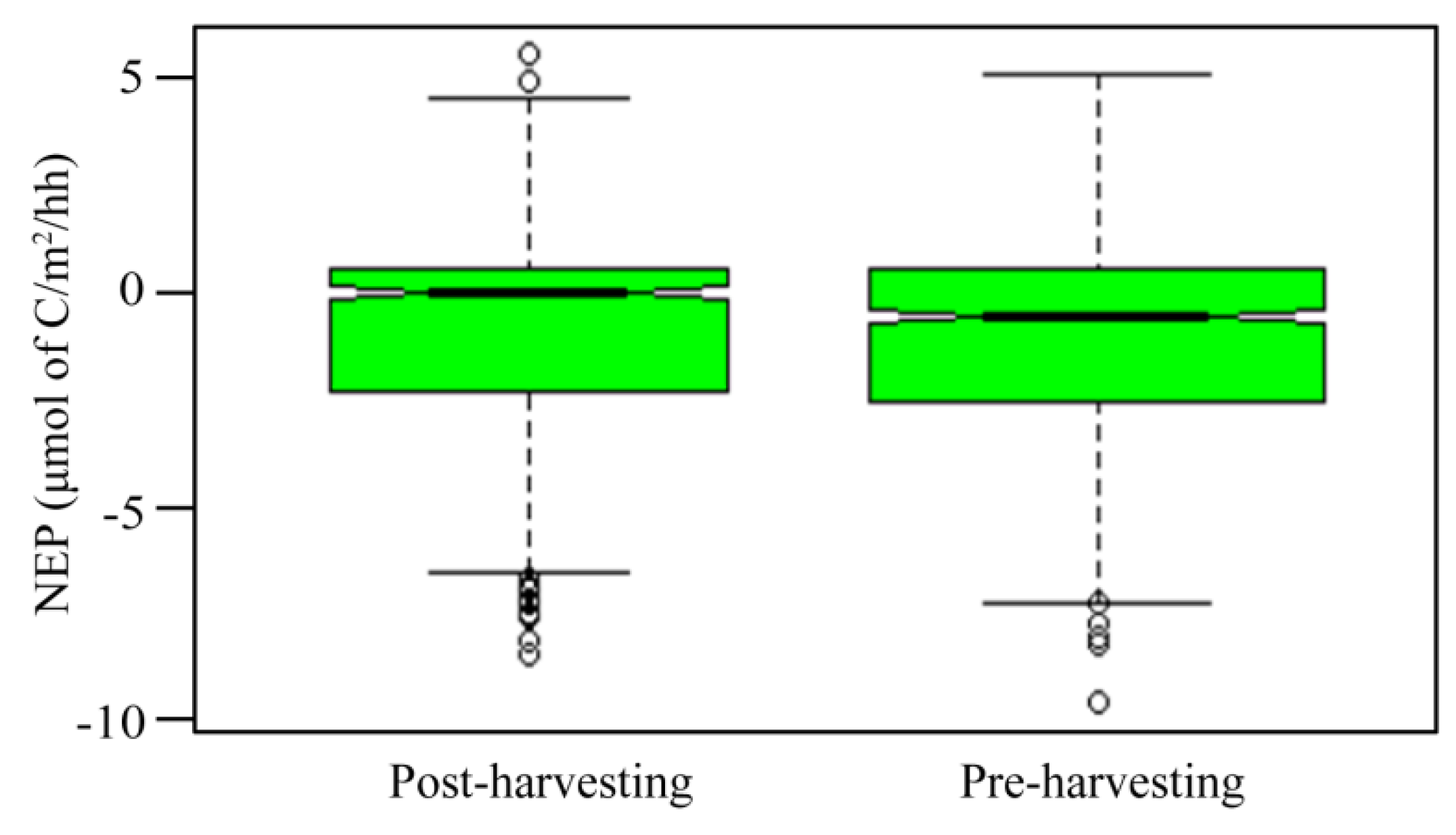
3.4. Comparison of Soil CO2 Effluxes Pre- and Post-Harvesting
3.5. Comparison of Soil CO2 Effluxes to Ecosystem Respiration
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mixed Effects Model fitted (Random effects were: Chamber, DOY) Fixed, as listed below, including interactions: | Sample df | AIC | BIC | 1:1 obs vs pred adj R2 |
|---|---|---|---|---|
| LN(Rs) ~ Ts only | 5 | 3854.785 | 3886.125 | 0.6467 |
| LN(Rs) ~ Ts + SM | 6 | 3597.288 | 3634.895 | 0.7272 |
| LN(Rs) ~ Ts + SM + Thinning | 7 | 3463.574 | 3507.450 | 0.7281 |
| LN(Rs) ~ Ts + SM + Thinning + Variety | 9 | 3459.169 | 3515.581 | 0.7280 |
| LN(Rs) ~ Ts*SM + Thinning + Variety | 10 | 3294.815 | 3357.495 | 0.7537 |
| LN(Rs) ~ Ts*SM*Thinning *Variety | 27 | 2922.077 | 3091.312 | 0.7571 |
| Variable | Pre-Harvesting Estimate | Post-Harvesting Estimate |
|---|---|---|
| Soil temperature | 0.06719 | –0.00243 |
| Soil moisture | 1.8378 | –6.97301 |
| Soil temperature and moisture interaction term | 0.1918 | 0.8676 |
| Intercept | –4.3074 | –3.1823 |
References
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef]
- Heimann, M.; Reichstein, M. Terrestrial ecosystem carbon dynamics and climate feedbacks. Nature 2008, 451, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Ciais, P.; Friedlingstein, P.; Peylin, P.; Reichstein, M.; Luyssaert, S.; Margolis, H.; Fang, J.; Barr, A.; Chen, A.; et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature 2008, 451, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forest. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Khomik, M.; Arain, M.A.; Liaw, K.-L.; McCaughey, J.H. The debut of a flexible model for soil respiration: the Gamma model. J. Geophys. Res.-BIOGEO 2009, 114, GO3004. [Google Scholar] [CrossRef]
- Jassal, R.S.; Black, T.A.; Nesic, Z.; Gaumont-Guay, D. Using automated non-steady-state chamber systems for making continuous long-term measurements of soil CO2 efflux in forest ecosystems. Agric. For. Meteorol. 2012, 161, 57–65. [Google Scholar] [CrossRef]
- Wharton, S.; Falk, M.; Bible, K.; Schroeder, M.; Paw U, K.T. Old-growth CO2 flux measurements reveal high sensitivity to climate anomalies across seasonal, annual and decadal time scales. Agric. For. Meteorol. 2012, 161, 1–14. [Google Scholar] [CrossRef]
- Bastos, A.; Running, S.W.; Gouveia, C.; Trigo, R.M. The global NPP dependence on ENSO: La Niña and the extraordinary year of 2011. J. Geophys. Res.-Atmos. 2013, 118, 1247–1255. [Google Scholar] [CrossRef]
- Hogg, E.H.; Michaelian, M.; Hook, T.I.; Undershultz, M.E. Recent climatic drying leads to age-independent growth reductions of white spruce stands in western Canada. Glob. Chang. Biol. 2017, 23, 5297–5308. [Google Scholar] [CrossRef]
- Vesala, T.; Suni, T.; Rannik, Ü.; Keronen, P.; Markkanen, T.; Sevanto, S.; Grönholm Smolander, S.; Kulmala, M.; Ilvesniemi, H.; Ojansuu, R.; et al. Effect of thinning on surface fluxes in a boreal forest. Glob. Biogeochem. Cycle 2005, 19. [Google Scholar] [CrossRef]
- Peng, Y.; Thomas, S.C.; Tian, D. Forest management and soil respiration: Implications for carbon sequestration. Environ. Rev. 2008, 16, 93–111. [Google Scholar] [CrossRef]
- Powers, M.D.; Webster, C.R.; Pregitzer, K.S.; Palik, B.J. Spatial dynamics of radial growth and growth efficiency in residual Pinus resinosa following aggregated retention harvesting. Can. J. For. Res. 2009, 39, 109–117. [Google Scholar] [CrossRef]
- D’Amato, A.W.; Bradford, J.B.; Fraver, S.; Palik, B.J. Forest management for mitigation and adaptation to climate change: Insights from long-term silviculture experiments. For. Ecol. Manag. 2011, 262, 803–816. [Google Scholar] [CrossRef]
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lohmus, A.; Martinez, P.G.; Messier, C.; et al. Retention forestry to maintain multifunctional forests: A world perspective. BioScience 2012, 62, 633–645. [Google Scholar] [CrossRef]
- Magruder, M.; Chhin, S.; Palik, B.; Bradford, J.B. Thinning increases climatic resilience of red pine. Can. J. For. Res. 2013, 43, 878–889. [Google Scholar] [CrossRef]
- Trant, J. Effects of Thinning on Carbon Dynamics in a Temperate Coniferous Forest. Master’s Thesis, McMaster University, Hamilton, ON, Canada, 2014; p. 64. [Google Scholar]
- OMNRF. A Silvicultural Guide to Managing Southern Ontario Forests; Ontario Ministry of Natural Resources and Forestry: Toronto, ON, Canada, 2000; p. 654.
- OMNRF. A Silvicultural Guide for the Great Lakes-St. Lawrence Conifer Forest in Ontario; Ontario Ministry of Natural Resources and Forestry: Toronto, ON, Canada, 1998; p. 424.
- Marchi, M.; Paletto, A.; Cantiani, P.; Bianchetto, E. Comparing thinning system effects on ecosystem services provision in artificial black pine (Pinus nigra J.F. Arnold) forests. Forests 2018, 9, 188. [Google Scholar] [CrossRef]
- Stoffel, J.I.; Gower, S.T.; Forrester, J.A.; Mladenoff, D.J. Effects of winter selective tree harvest on soil microclimate and surface CO2 efflux of a northern hardwood forest. For. Ecol. Manag. 2010, 259, 257–265. [Google Scholar] [CrossRef]
- Woods, S.; Ahl, R.; Sappington, J.; McCaughey, W. Snow accumulation in thinned lodgepole pine stands, Montana, USA. For. Ecol. Manag. 2006, 235, 202–211. [Google Scholar] [CrossRef]
- Peng, Y.; Thomas, S.C. Soil CO2 efflux in uneven-aged managed forests: Temporal patterns following harvest and effects of edaphic heterogeneity. Plant Soil 2006, 289, 253–264. [Google Scholar] [CrossRef]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- Drewitt, G.B.; Black, T.A.; Nesic, Z.; Humphreys, E.R.; Jork, E.M.; Swanson, R.; Ethier, G.J.; Griffis, T.; Mergenstern, K. Measuring forest floor CO2 fluxes in a Douglas-fir forest. Agric. For. Meteorol. 2002, 110, 299–317. [Google Scholar] [CrossRef]
- Jassal, R.; Black, T.A.; Novak, M.; Morgenstern, K.; Nesic, Z.; Gaumont-Guay, D. Relationship between soil CO2 concentrations and forest-floor CO2 effluxes. Agric. For. Meteorol. 2005, 130, 176–192. [Google Scholar] [CrossRef]
- Khomik, M.; Arain, M.A.; Brodeur, J.J.; Peichl, M.; Restrepo-Coupe, N.; McLaren, J.D. Relative contributions of soil, foliar, and woody tissue respiration to total ecosystem respiration in four pine forests of different ages. J. Geophys. Res. 2010, 115, G03024. [Google Scholar] [CrossRef]
- Peichl, M.; Arain, M.A.; Brodeur, J.J. Age effects on carbon fluxes in temperate pine forests. Agric. For. Meteorol. 2010, 150, 1090–1101. [Google Scholar] [CrossRef]
- Peichl, M.; Brodeur, J.J.; Khomik, M.; Arain, M.A. Biometric and eddy covariance based estimates of carbon fluxes in an age-sequence of temperate pine forests. Agric. For. Meteorol. 2010, 150, 952–965. [Google Scholar] [CrossRef]
- Arain, M.A.; Restrepo-Coupe, N. Net ecosystem production in a temperate pine plantation in southeastern Canada. Agric. For. Meteorol. 2005, 128, 223–241. [Google Scholar] [CrossRef]
- Kula, M. Biometric-Based Carbon Estimates and Environmental Controls within an Age-Sequence of Temperate Forests. Master’s Thesis, McMaster University, Hamilton, ON, Canada, 2014; p. 64. [Google Scholar]
- Nicholas, E. Assessing Seasonal Dynamics of Soil CO2 Efflux Using Continuous Measurements in a Temperate Pine Forest. Master’s Thesis, McMaster University, Hamilton, ON, Canada, 2011. [Google Scholar]
- Gaumont-Guay, D.; Black, T.A.; Barr, A.G.; Jassal, R.S.; Nesic, Z. Biophysical controls on rhizospheric and heterotrophic components of soil respiration in a boreal black spruce stand. Tree Physiol. 2008, 28, 161–171. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Hicks, B.B.; Meyers, T.P. Measuring biosphere-atmosphere exchanges of biologically related gasses with micrometeorological methods. Ecology 1998, 69, 1331–1340. [Google Scholar] [CrossRef]
- Brodeur, J.J. Data-Driven Approaches for Sustainable Operation and Defensible Results in a Long-Term, Multi-Site Ecosystem Flux Measurement Program. Ph.D. Thesis, McMaster University, Hamilton, ON, Canada, 2014; p. 232. [Google Scholar]
- Hungate, B.A.; Jackson, R.B.; Field, C.B.; Chapin III, F.S. Detecting changes in soil carbon in CO2 enrichment experiments. Plant Soil 1996, 187, 135–145. [Google Scholar] [CrossRef]
- Johnson, D.H. The insignificance of statistical significance testing. J. Wildl. Manag. 1999, 63, 763–772. [Google Scholar] [CrossRef]
- Cueva, A.; Bullock, S.H.; Lopez-Reyes, E.; Vargas, R. Potential bias of daily soil CO2 efflux estimates due to sampling time. Sci. Rep. 2017, 7, 11925. [Google Scholar] [CrossRef] [PubMed]
- Heinemeyer, A.; DiBene, C.; Lloyd, R.; Tortorella, D.; Baxter, R.; Huntley, B.; Gelsomino, A.; Ineson, P. Soil respiration: Implications of the plant-soil continuum and respiration chamber collar-insertion depth on measurement and modelling of soil CO2 efflux rates in three ecosystems. Eur. J. Soil Sci. 2011, 62, 82–94. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R.-project.org/ (accessed on 18 February 2020).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hurlbert, S.H. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef]
- Skubel, R.; Arain, M.A.; Peichl, M.; Brodeur, J.J.; Khomik, M.; Thorne, R.; Trant, J.; Kula, M. Age effects on the water-use efficiency and water-use dynamics of temperate pine plantation forests. Hydrol. Process. 2015, 29, 4100–4113. [Google Scholar] [CrossRef]
- Borken, W.; Xu, Y.J.; Davidson, E.A.; Beese, F. Site and temporal variation of soil respiration in European beech, Norway spruce, and Scots pine forests. Glob. Chang. Biol. 2002, 8, 1205–1216. [Google Scholar] [CrossRef]
- Monson, R.K.; Burns, S.P.; Williams, M.W.; Delany, A.C.; Weintraub, M.; Lipson, D.A. The contribution of beneath-snow soil respiration to total ecosystem respiration in a high-elevation, subalpine forest. Glob. Biogeochem. Cycle 2006, 20. [Google Scholar] [CrossRef]
- Ryu, S.-R.; Concilio, A.; Chen, J.; North, M.; Ma, S. Prescribed burning and mechanical thinning effects on belowground conditions and soil respiration in a mixed-conifer forest, California. For. Ecol. Manag. 2009, 257, 1324–1332. [Google Scholar] [CrossRef]
- Winston, G.C.; Sundquist, E.T.; Stephens, B.B.; Trumbore, S.E. Winter CO2 fluxes in a boreal forest. J. Geophys. Res. 1997, 102, 28795–28804. [Google Scholar] [CrossRef]
- DeForest, J.L.; Chen, J.; McNulty, S.G. Leaf litter is an important mediator of soil respiration in an oak-dominated forest. Int. J. Biometeorol. 2009, 53, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Qi, Y.; Xu, M.; Misson, L.; Goldstein, A.H. Forest thinning and soil respiration in a ponderosa pine plantation in the Sierra Nevada. Tree Physiol. 2005, 25, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Shabaga, J.A.; Basiliko, N.; Caspersen, J.P.; Jones, T.A. Seasonal controls on patterns of soil respiration and temperature sensitivity in a northern mixed deciduous forest following partial-harvesting. For. Ecol. Manag. 2015, 348, 208–219. [Google Scholar] [CrossRef]
- Wang, H.; Liu, W.; Wang, W.; Zu, Y. Influence of long-term thinning on the biomass carbon and soil respiration in a larch (Larix gmelinii) forest in Northeastern China. Sci. World J. 2013. [Google Scholar] [CrossRef]
- Sullivan, B.W.; Kolb, T.E.; Hart, S.C.; Kaye, J.P.; Dore, S.; Montes-Helu, M. Thinning reduces soil carbon dioxide but not methane flux from southwestern USA ponderosa pine forests. For. Ecol. Manag. 2008, 255, 4047–4055. [Google Scholar] [CrossRef]
- Selig, M.F.; Seiler, J.R.; Tyree, M.C. Soil carbon and CO2 efflux as influenced by the thinning of loblolly pine (Pinus taeda L.) plantations on the piedmont of Virginia. For. Sci. 2008, 54, 58–66. [Google Scholar]
- Ohashi, M.; Gyokusen, K.; Saito, A. Measurement of carbon dioxide evolution from a Japanese cedar (Crytomeria japonica D. Don) forest floor using an open-flow chamber method. For. Ecol. Manag. 1999, 123, 105–114. [Google Scholar] [CrossRef]
- Concilio, A.; Ma, S.; Li, Q.; LeMoine, J.; Chen, J.; North, M.; Moorhead, D.; Jensen, R. Soil respiration response to prescribed burning and thinning in mixed-conifer and hardwood forests. Can. J. For. Res. 2005, 35, 1581–1591. [Google Scholar] [CrossRef]
- Concilio, A.; Ma, S.; Ryu, S.-R.; North, M.; Chen, J. Soil respiration response to experimental disturbances over 3 years. For. Ecol. Manag. 2006, 228, 82–90. [Google Scholar] [CrossRef]
- Ma, S.; Chen, J.; North, M.; Erickson, H.; Bresee, M.; LeMoine, J. Short-term effects of experimental burning and thinning on soil respiration in an old-growth, mixed-conifer forest. Environ. Manag. 2004, 33, S148–S159. [Google Scholar] [CrossRef]
- Brændholt, A.; Steenberg Larsen, K.; Ibrom, A.; Pilegaard, K. Overestimation of closed-chamber soil CO2 effluxes at low atmospheric turbulence. Biogeosciences 2017, 14, 1603–1616. [Google Scholar] [CrossRef]






| Environmental Variable | Pre-Thinning (no. of observations) | Post-Thinning (no. of observations) | % Difference | t-statistic | p-value |
|---|---|---|---|---|---|
| Soil Temperature | 9.4 ± 7.5 °C (n = 1274) | 9.1 ± 7.5 °C (n = 1095) | −3% | −0.90 | 0.37 |
| Soil Moisture | 0.11 ± 0.03 m3/m3 (n = 1274) | 0.12 ± 0.03 m3/m3 (n = 1095) | 3% | 2.48 | 0.01 |
| Mean Soil CO2 Efflux (μmol of C m2·s−1) | |||||
| FS1 | 6.09 ± 3.66 (n = 805) | 4.01 ± 3.23 (n = 226) | −34% | −4.69* | 0.00 |
| FS2 | 4.64 ± 3.20 (n = 919) | 4.03 ± 3.51 (n = 261) | −13% | −3.48* | 0.00 |
| FS3 | 5.83 ± 3.90 (n = 654) | 5.71 ± 4.03 (n = 253) | −2% | −1.35 | 0.18 |
| FS4 | 5.91 ± 3.78 (n = 578) | 4.90 ± 3.70 (n = 256) | −17% | −4.30* | 0.00 |
| FS5 | 6.73 ± 4.68 (n = 409) | 4.68 ± 4.09 (n = 282) | −30% | −6.02* | 0.00 |
| FLR | 4.06 ± 2.76 (n = 882) | 3.30 ± 2.77 (n = 313) | −19% | 69.2* | 0.00 |
| FRT1 | 3.84 ± 2.80 (n = 627) | 2.58 ± 2.63 (n = 214) | −33% | −7.44* | 0.00 |
| FRT2 | 4.44 ± 2.79 (n = 283) | 3.65 ± 2.84 (n = 191) | −18% | −4.81* | 0.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thorne, R.; Khomik, M.; Hayman, E.; Arain, A. Response of Soil CO2 Efflux to Shelterwood Harvesting in a Mature Temperate Pine Forest. Forests 2020, 11, 304. https://doi.org/10.3390/f11030304
Thorne R, Khomik M, Hayman E, Arain A. Response of Soil CO2 Efflux to Shelterwood Harvesting in a Mature Temperate Pine Forest. Forests. 2020; 11(3):304. https://doi.org/10.3390/f11030304
Chicago/Turabian StyleThorne, Robin, Myroslava Khomik, Emily Hayman, and Altaf Arain. 2020. "Response of Soil CO2 Efflux to Shelterwood Harvesting in a Mature Temperate Pine Forest" Forests 11, no. 3: 304. https://doi.org/10.3390/f11030304
APA StyleThorne, R., Khomik, M., Hayman, E., & Arain, A. (2020). Response of Soil CO2 Efflux to Shelterwood Harvesting in a Mature Temperate Pine Forest. Forests, 11(3), 304. https://doi.org/10.3390/f11030304