Tree Regeneration Structure Following Beech Bark Disease-Motivated Harvests: Factors Associated with Patterns and Management Implications


Abstract
1. Introduction
1.1. Beech Bark Disease, Beech Regeneration Responses, and Implications for Forest Dynamics
1.2. Factors Potentially Influencing BBD Effects on Beech Regneration Patterns
1.3. Objectives
2. Materials and Methods
2.1. Study Area and Stand Selection
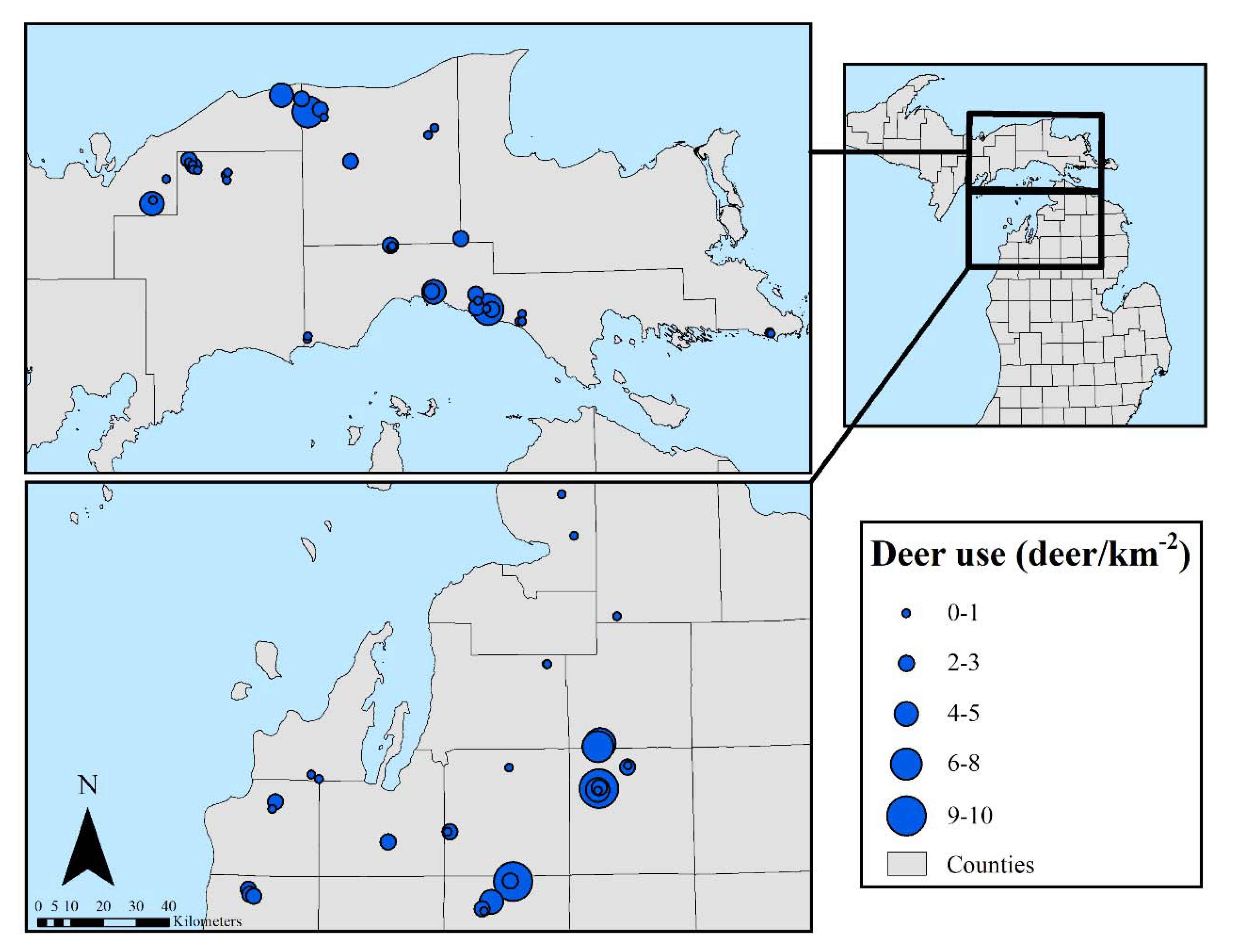
2.2. Measurements
2.3. Statistical Analysis
3. Results
3.1. General Stand Characteristics
3.2. Tree Regeneration
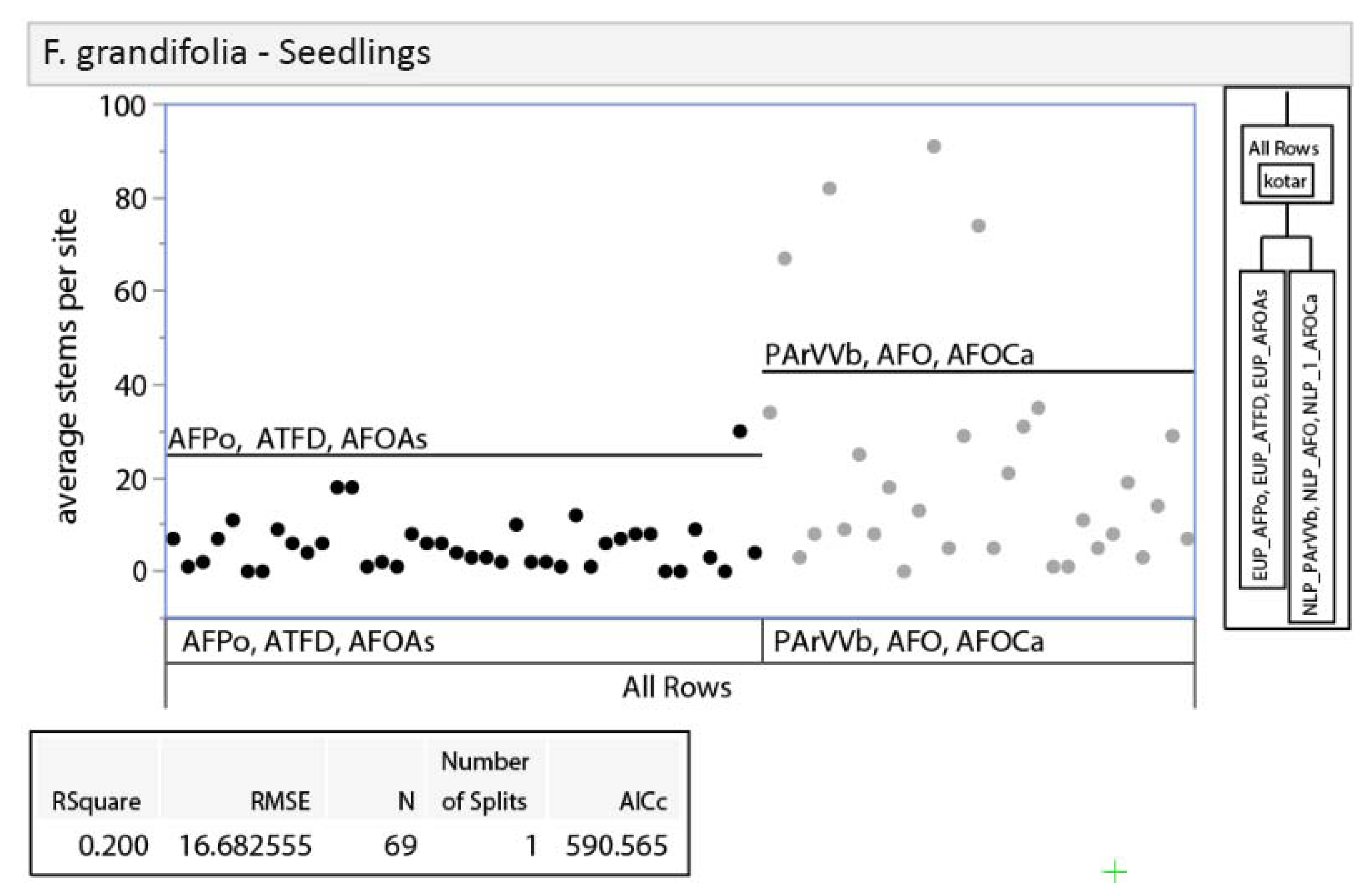
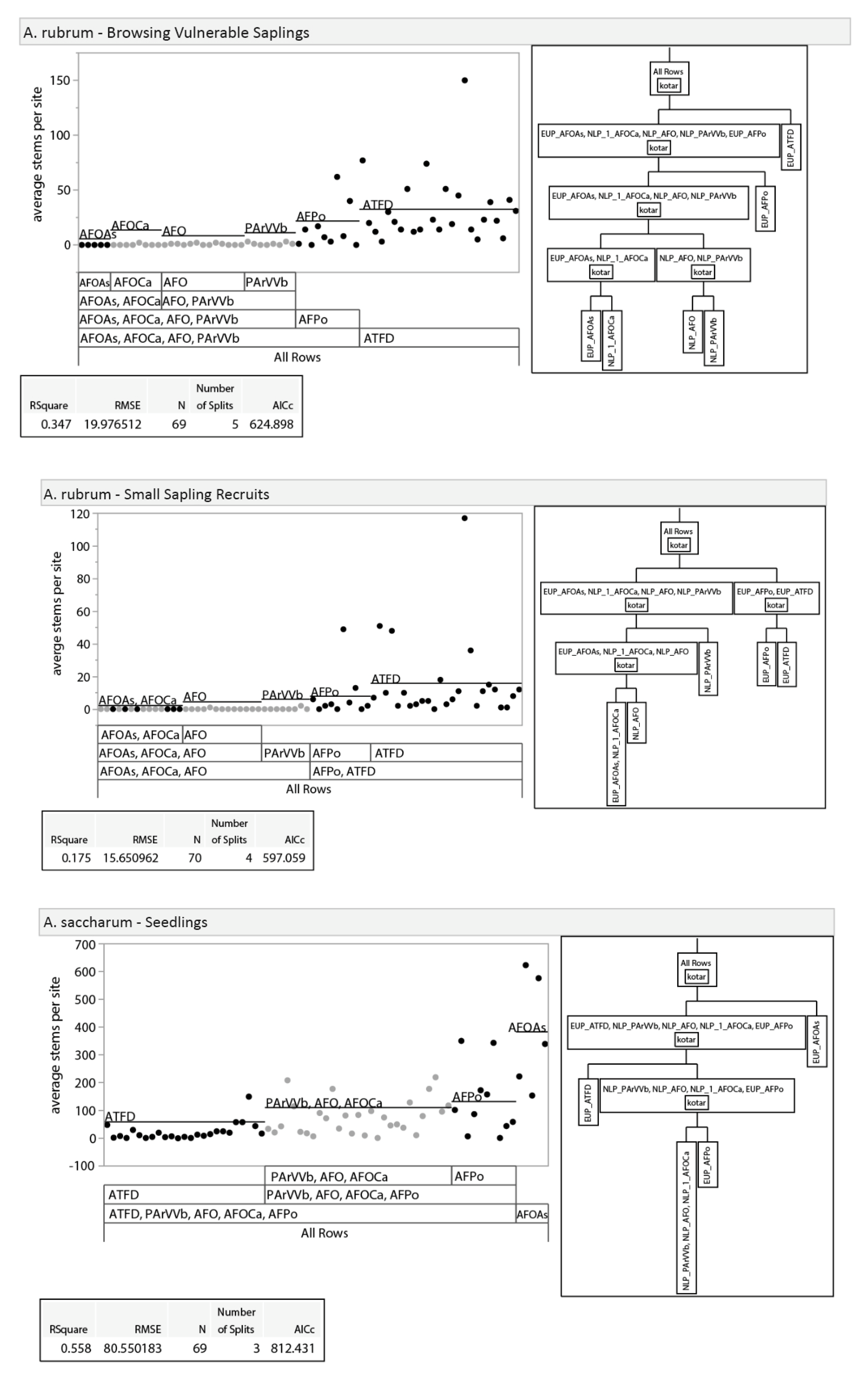
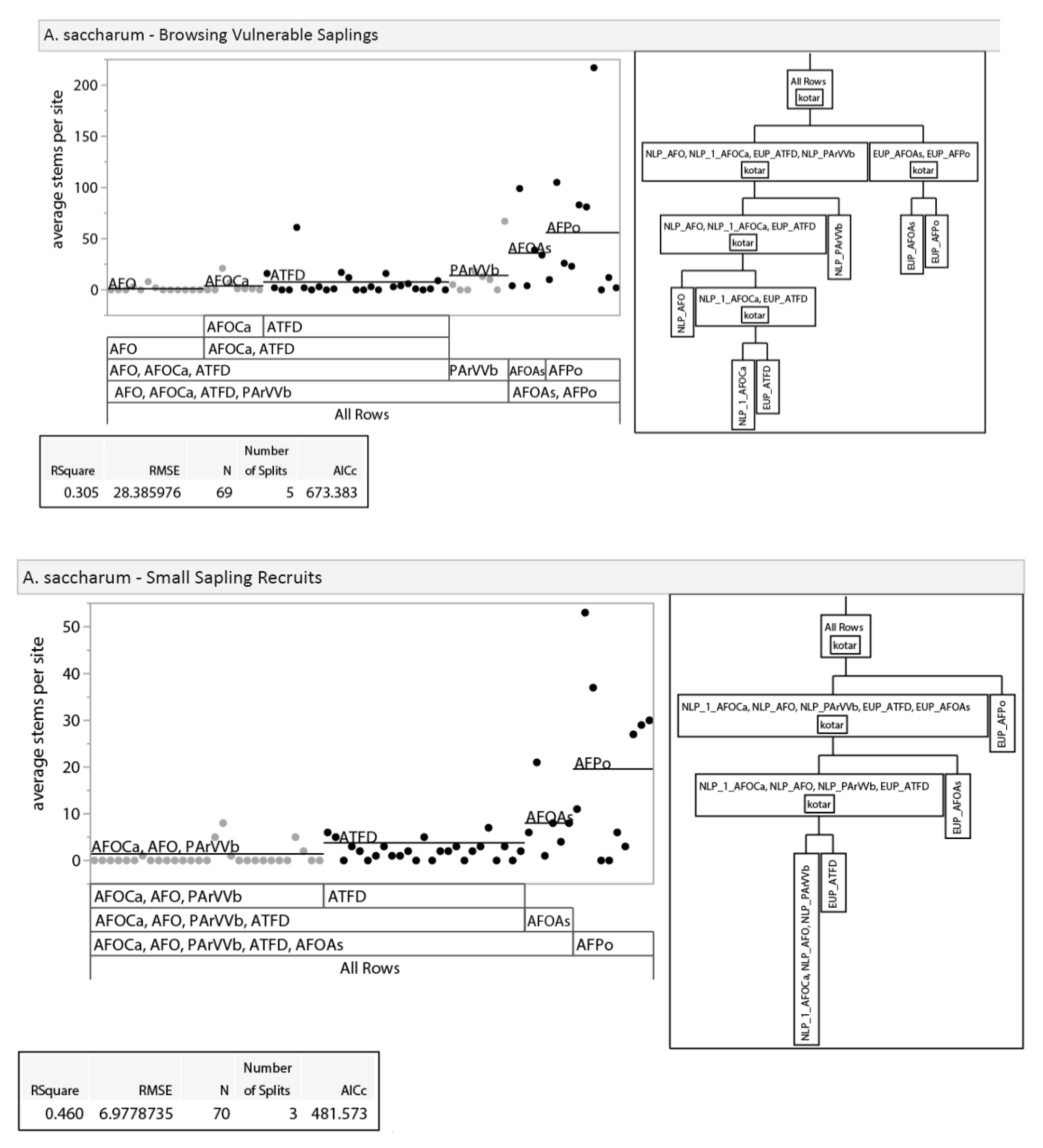
3.2.1. Tree Regeneration—Effects of Habitat Type
3.2.2. Tree Regeneration—Effects of Basal Area
3.2.3. Tree Regeneration–Effects of Deer Use
3.2.4. Tree Regeneration—Effects of Taller Regeneration Strata on Seedling Density
4. Discussion
4.1. Factors Related to Regeneration Size Class Structure
4.2. Management Implications
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Size Class | Habitat Type | Prob > S |
|---|---|---|---|
| F. grandifolia | Seedlings | High | 0.2098 |
| Medium | 0.0006 | ||
| Low | 0.0017 | ||
| Browsing vulnerable saplings | High | 0.2028 | |
| Medium | 0.0002 | ||
| Low | < 0.0001 | ||
| Small sapling recruits | High | 0.4858 | |
| Medium | 0.0038 | ||
| Low | 0.0146 | ||
| A. rubrum | Seedlings | High | 0.0163 |
| Medium | 0.0276 | ||
| Low | 0.0034 | ||
| Browsing vulnerable saplings | High | 0.6154 | |
| Medium | 0.003 | ||
| Low | < 0.0001 | ||
| Small sapling recruits | High | 1.0000 | |
| Medium | 0.0005 | ||
| Low | < 0.0001 | ||
| A. saccharum | Seedlings | High | 0.0031 |
| Medium | 0.1753 | ||
| Low | 0.0007 | ||
| Browsing vulnerable saplings | High | 0.0085 | |
| Medium | < 0.0001 | ||
| Low | 0.1831 | ||
| Small sapling recruits | High | 0.0010 | |
| Medium | 0.0005 | ||
| Low | 0.028 |

| Species | Region | Size Class | AICc | R2 | Prob > ChiSquare |
|---|---|---|---|---|---|
| F. americana | NLP | Seedlings | 148.0 | 0.36 | H = 0.0003 |
| Browsing vulnerable saplings | 156.2 | 0.48 | H < 0.0001, D = 0.0190, H × D = 0.0075 (D-) | ||
| Small sapling recruits | 64.7 | 0.68 | H ≤ 0.0001, (H × D) ≤ 0.0001, (H × BA) ≤ 0.0001 | ||
| Large sapling recruits | - | - | |||
| P. serotina | NLP | Seedlings | 72.5 | 0.00 | |
| Browsing vulnerable saplings | 108.9 | 0.00 | |||
| Small sapling recruits | 59.0 | 0.00 | |||
| Large sapling recruits | - | - | |||
| P. pensylvanica | EUP | Seedlings | 16.8 | 0.00 | |
| Browsing vulnerable saplings | 13.3 | 0.06 | |||
| Small sapling recruits | 110.1 | 0.41 | H ≤ 0.0001, D ≤ 0.0001, BA ≤ 0.0001H × BA = 0.0002, D × BA ≤ 0.0001 (BA-, D-) | ||
| Large sapling recruits | 107.0 | 0.27 | H ≤ 0.0001, D = 0.0072, D × BA = 0.0022 (BA-, D-) | ||
| B. alleghaniensis | EUP | Seedlings | 163.3 | 0.15 | H = 0.0002 |
| Browsing vulnerable saplings | 101.1 | 0.27 | H ≤ 0.0001, BA = 0.0374, H × D = 0.0034 (BA+) | ||
| Small sapling recruits | 55.9 | 0.22 | H ≤ 0.0001, D = 0.0013, BA = 0.0091 (BA+, D-) | ||
| Large sapling recruits | 74.1 | 0.2 | H = 0.0009, D × BA = 0.0233 | ||
| T. canadensis | EUP | Seedlings | 57.9 | 0.24 | H ≤ 0.0001, BA = 0.0104 (BA+) |
| Browsing vulnerable saplings | 34.5 | 0.00 | |||
| Small sapling recruits | 56.4 | 0.2 | H ≤ 0.0001, BA = 0.0357 (BA+) | ||
| Large sapling recruits | 51.0 | 0.00 | |||
| A. balsamea | EUP | Seedlings | 49.5 | 0.00 | |
| Browsing vulnerable saplings | 79.5 | 0.33 | H ≤ 0.0001, BA = 0.0085, H × D = 0.0006 (BA+) | ||
| Small sapling recruits | 101.3 | 0.31 | H ≤ 0.0001, H × D = 0.0034 | ||
| Large sapling recruits | 147.1 | 0.19 | H ≤ 0.0001 | ||
| A. pensylvanicum | EUP | Seedlings | 80.6 | 0.28 | H ≤ 0.0001, D ≤ 0.0001 (D+) |
| Browsing vulnerable saplings | 111.2 | 0.27 | D ≤ 0.0001 (D+) | ||
| Small sapling recruits | 171.7 | 0 | |||
| Large sapling recruits | 102.7 | 0 |
References
- Ehrlich, J. The Beech Bark Disease a Nectria Disease of Fagus, Following Cryptococcus Fagi (Baer.). Can. J. Res. 1934, 10, 593–692. [Google Scholar] [CrossRef]
- Cale, J.A.; Teale, S.A.; Johnston, M.T.; Boyer, G.L.; Perri, K.A.; Castello, J.D. New ecological and physiological dimensions of beech bark disease development in aftermath forests. For. Ecol. Manag. 2015, 336, 99–108. [Google Scholar] [CrossRef]
- Cale, J.A.; McNulty, S.A.; Teale, S.A.; Castello, J.D. The impact of beech thickets on biodiversity. Biol. Invasions 2013, 15, 699–706. [Google Scholar] [CrossRef]
- Houston, D.R. Beech Bark Disease—The Aftermath Forests are Structured for a New Outbreak. J. For. 1975, 73, 660–663. [Google Scholar]
- Shigo, A.L. The Beech Bark Disease Today in the Northeastern, U.S. J. For. 1972, 70, 286–289. [Google Scholar]
- Giencke, L.M.; Dovčiak, M.; Mountrakis, G.; Cale, J.A.; Mitchell, M.J. Beech bark disease: spatial patterns of thicket formation and disease spread in an aftermath forest in the northeastern United States. Can. J. Res. 2014, 44, 1042–1050. [Google Scholar] [CrossRef]
- Nyland, R.D.; Bashant, A.L.; Bohn, K.K.; Verostek, J.M. Interference to Hardwood Regeneration in Northeastern North America: Controlling Effects of American Beech, Striped Maple, and Hobblebush. North. J. Appl. For. 2006, 23, 122–132. [Google Scholar] [CrossRef]
- Hane, E.N. Indirect effects of beech bark disease on sugar maple seedling survival. Can. J. For. Res. 2003, 33, 807–813. [Google Scholar] [CrossRef]
- O’Brien, J.G.; Ostry, M.E.; Mielke, M.E.; Mech, R.; Heyd, R.L.; McCullough, D.G. First Report of Beech Bark Disease in Michigan. Plant Dis. 2001, 85, 921. [Google Scholar] [CrossRef]
- Houston, D.R. Major new tree disease epidemics: beech bark disease. Annu. Rev. Phytopathol. 1994, 32, 75–87. [Google Scholar] [CrossRef]
- Le Guerrier, C.; Marceau, D.J.; Bouchard, A.; Brisson, J. A modelling approach to assess the long-term impact of beech bark disease in northern hardwood forest. Can. J. Forest Res. 2003, 33, 2416–2425. [Google Scholar] [CrossRef][Green Version]
- Kearney, A.; Mccullough, D.G.; Walters, M. Impacts of Beech Bark Disease on Understory Composition in Michigan. In Beech Bark Disease; Michigan State University: East Lansing, MI, USA, 2004; pp. 58–60. [Google Scholar]
- Wieferich, J. Beech Bark Disease in Michigan: Distribution, Impacts and Dynamics. Master’s Thesis, Michigan State University, East Lansing, MI, USA, 2013. [Google Scholar]
- Burger, T.L.; Kotar, J. A Guide to Forest Communities and Habitat Types of Michigan; Department of Forest Ecology and Management, University of Wisonsin-Madison: Madison, WI, USA, 2003. [Google Scholar]
- Rooney, T.P.; Waller, D.M. Direct and indirect effects of white-tailed deer in forest ecosystems. For. Ecol. Manag. 2003, 181, 165–176. [Google Scholar] [CrossRef]
- Randall, J.A.; Walters, M.B. Deer density effects on vegetation in aspen forest understories over site productivity and stand age gradients. For. Ecol. Manag. 2011, 261, 408–415. [Google Scholar] [CrossRef]
- Frigoletto, E.; Wylie, P.; Pasquini, S.C.; Carson, W.P. Excluding deer increases the proportion of beech saplings originating from seed versus those of root sprout origin. J. Torrey Bot. Soc. 2017, 144, 379–384. [Google Scholar] [CrossRef][Green Version]
- Kern, C.; Erdmann, G.; Kenefic, L.; Palik, B.; Strong, T. Development of the Selection System in Northern Hardwood Forests of the Lake States: An 80-Year Silviculture Research Legacy; USDA For. Serv. Exp. For. Ranges, Ed.; Springer: New York, NY, USA, 2014; pp. 201–223. [Google Scholar]
- Eyre, F.H.; Zillgitt, W.M. Partial Cuttings in Northern Hardwoods of the Lake States: Twenty-Year Experimental Results; Technical Bulletin LS-1076; USDA, For. Serv., Lake States Forest Experiment Station: Broomall, PA, USA, 1953; p. 124.
- McShea, W.J.; Underwood, H.B.; Rappole, J.H. The Science of Overabundance; Smithsonian Institution Press: Washington, DC, USA, 1997; ISBN 1560986816. [Google Scholar]
- McClure, J.W.; Lee, T.D. Small-scale disturbance in a northern hardwoods forest: Effects on tree species abundance and distribution. Can. J. For. Res. 1993, 23, 1347–1360. [Google Scholar] [CrossRef]
- Webster, C.R.; Lorimer, C.G. Minimum opening sizes for canopy recruitment of midtolerant tree species: A retrospective approach. Ecol. Appl. 2005, 15, 1245–1262. [Google Scholar] [CrossRef]
- Kneeshaw, D.D.; Prévost, M. Natural canopy gap disturbances and their role in maintaining mixed-species forests of central Quebec, Canada. Can. J. For. Res. 2007, 37, 1534–1544. [Google Scholar] [CrossRef]
- Matonis, M.S.; Walters, M.B.; Millington, J.D.A. Gap-, stand-, and landscape-scale factors contribute to poor sugar maple regeneration after timber harvest. For. Ecol. Manag. 2011, 262, 286–298. [Google Scholar] [CrossRef]
- Handler, S.; Duveneck, M.J.; Iverson, L.; Peters, E.; Scheller, R.M.; Wythers, K.R.; Brandt, L.; Butler, P.; Janowiak, M.; Shannon, P.D.; et al. Michigan Forest Ecosystem Vulnerability Assessment and Synthesis: A Report from the Northwoods Climate Change Response Framework Project; Gen. Tech. Rep. NRS-129; USDA, For. Serv., Northern Research Station: Newtown Square, PA, USA, 2014; p. 229.
- PRISM Climate Group. Oregon State University. 2016. Available online: http//prism.oregonstate.edu (accessed on 10 August 2018).
- Neumann, D. Silvics and Management Guidance Manual; Neumann, D., Ed.; Michigan Department of Natural Resources Forest Resources and Wildlife Division: Lansing, MI, USA, 2015; p. 4111.
- Neff, D.J. The Pellet-Group Count Technique for Big Game Trend, Census, and Distribution: A Review. J. Wildl. Manag. 1968, 32, 597–614. [Google Scholar] [CrossRef]
- Marques, F.F.C.; Buckland, S.T.; Goffin, D.; Dixon, C.E.; Borchers, D.L.; Mayle, B.A.; Peace, A.J. Estimating deer abundance from line transect surveys of dung: Sika deer in southern Scotland. J. Appl. Ecol. 2003, 38, 349–363. [Google Scholar] [CrossRef]
- Millington, J.D.A.; Walters, M.B.; Matonis, M.S.; Liu, J. Effects of local and regional landscape characteristics on wildlife distribution across managed forests. For. Ecol. Manag. 2010, 259, 1102–1110. [Google Scholar] [CrossRef]
- Hill, H.R. The 2001 Deer Pellet Group Surveys; Report No. 3349; Michigan Department of Natural Resources Wildlife Division: Lansing, MI, USA, 2001.
- Henttonen, H.M.; Kangas, A. Optimal plot design in a multipurpose forest inventory. For. Ecosyst. 2015, 2, 31. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture, Forest Service. Forest Iventory and Analysis National Core Field Guide Volume I: Field Data Collection Procedures for Phase 2 Plots. Version 8.0; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2018.
- Walters; Farinosi; Willis. Deer browsing and shrub competition set sapling recruitment height and interact with light to shape recruitment niches for temperate forest tree species. For. Ecol. Man. 2020. submitted. [Google Scholar]
- Mulekar, M.S.; Mauromoustakos, A. More powerful but still easy to use data analysis with JMP© version 5. In Proceedings of the 2002 Annual Meeting of American Statistical Association, New York, NY, USA, 10–13 August 2002. [Google Scholar]
- SAS Institute Inc. JMP® 13 Basic Analysis; SAS Institute Inc.: Cary, NC, USA, 2016. [Google Scholar]
- Crotty, M.; Barker, C. Penalizing Your Models: An Overview of the Generalized Regression Platform; SAS Insitute: Cary, NC, USA, 2014. [Google Scholar]
- OMNR. Ontario Tree Marking Guide, Version 1.1; Ont. Min. Nat. Resour., Queen’s Printer for Ontario: Toronto, ON, Canada, 2004; p. 252. [Google Scholar]
- Walters, M.B.; Farinosi, E.J.; Willis, J.L.; Gottschalk, K.W. Managing for diversity: Harvest gap size drives complex light, vegetation, and deer herbivory impacts on tree seedlings. Ecosphere 2016, 7, 1–29. [Google Scholar] [CrossRef]
- Kern, C.C.; Reich, P.B.; Montgomery, R.A.; Strong, T.F. Do deer and shrubs override canopy gap size effects on growth and survival of yellow birch, northern red oak, eastern white pine, and eastern hemlock seedlings? For. Ecol. Manag. 2012, 267, 134–143. [Google Scholar] [CrossRef]
- McWilliams, W.H.; Westfall, J.A.; Brose, P.H.; Dey, D.C.; D’Amato, A.W.; Dickinson, Y.L.; Fajvan, M.A.; Kenefic, L.S.; Kern, C.C.; Laustsen, K.M.; et al. Subcontinental-Scale Patterns of Large-Ungulate Herbivory and Synoptic Review of Restoration Management Implications for Midwestern and Northeastern Forests; Gen. Tech. Rep. NRS-182; U.S. Dep. Agric. For. Serv. North. Res. Station: Newtown Square, PA, USA, 2018; pp. 1–24. [CrossRef]
- Kern, C.C.; Burton, J.I.; Raymond, P.; D’amato, A.W.; Keeton, W.S.; Royo, A.A.; Walters, M.B.; Webster, C.R.; Willis, J.L. Challenges facing gap-based silviculture and possible solutions for mesic northern forests in North America. For. Int. J. For. Res. For. 2017, 90, 4–17. [Google Scholar] [CrossRef]
- Melillo, J.M.; Aber, J.D.; Muratore, J.F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 1982, 63, 621–626. [Google Scholar] [CrossRef]
- Molofsky, J.; Augspurger, C.K. The effect of leaf litter on early seedling establishment in a tropical forest. Ecology 1992, 73, 68–77. [Google Scholar] [CrossRef]
- Nolet, P.; Bouffard, D.; Doyon, F.; Delagrange, S. Relationship between canopy disturbance history and current sapling density of Fagus grandifolia and Acer saccharum in a northern hardwood landscape. Can. J. For. Res. 2008, 38, 216–225. [Google Scholar] [CrossRef]
- Angers, V.A.; Messier, C.; Beaudet, M.; Leduc, A. Comparing composition and structure in old-growth and harvested (selection and diameter-limit cuts) northern hardwood stands in Quebec. For. Ecol. Manag. 2005, 217, 275–293. [Google Scholar] [CrossRef]
- Leak, W.B. Species composition and structure of a northern hardwood stand after 61 years of group/patch selection. North. J. Appl. For. 1999, 16, 151–153. [Google Scholar] [CrossRef]
- Roy, M.È.; Nolet, P. Early-stage of invasion by beech bark disease does not necessarily trigger American beech root sucker establishment in hardwood stands. Biol. Invasions 2018, 20, 3245–3254. [Google Scholar] [CrossRef]
- Diamond, J. Overview: Laboratory Experiments, Field Experiments, and Natural Experiments. In Community Ecology; Diamond, J., Case, T.J., Eds.; Harper & Row: New York, NY, USA, 1986; pp. 3–22. [Google Scholar]
- Solomon, D.S.; Leak, W.B. Simulated Yields for Managed Northern Hardwood Stands; Res. Pap. NE578; U.S. Dep. Agric. For. Serv. Northeast. For. Exp. Station: Broomall, PA, USA, 1986; p. 24.
- Arbogast, C.J. Marking Guides for Northern Hardwoods under the Selection System; Station Paper LS-56; U.S. Department of Agriculture, Forest Service, Lake States Forest Experiment Station: St. Paul, MN, USA, 1957; p. 20.
- Donoso, P.J.; Nyland, R.D. Interference to hardwood regeneration in northeastern North America: The effects of raspberries (Rubus spp.) following clearcutting and shelterwood methods. North. J. Appl. For. 2006, 23, 288–296. [Google Scholar] [CrossRef]
- Hannah, P.R. Potential of beech and striped maple to dominate regeneration on eastern hardwood sites. In Proceedings of the 6th Central Hardwood Forest Conference; Knoxville, TN, USA, 24–26 Febraury 1987; Hay, R.L., Gottschalk, K.W., Eds.; University of Tennessee: Knoxville, TN, USA, 1987; Abstract Number 526. [Google Scholar]
- Bohn, K.K.; Nyland, R.D. Forecasting development of understory American beech after partial cutting in uneven-aged northern hardwood stands. For. Ecol. Manag. 2003, 180, 453–461. [Google Scholar] [CrossRef]
- Mallik, A.; Wood, K.; Hollstedt, C.; MCLaughlan, M. Cut Stump Herbicide Treatments to Reduce Sprouting and Root Suckering; Note TN-39; Ont. Min. Natur. Resourc., Northwest. Sci. and Technol.: Thunder Bay, ON, Canada, 1997; p. 12. [Google Scholar]
- Mallett, A.L. Management of Understory American Beech by Manual and Chemical Control Methods. Master’s Thesis, SUNY Coll. Environ. Sci. and For., Syracuse, NY, USA, 2002; 152p. [Google Scholar]
- Farrar, A.; Ostrocsky, W.D. Dynamics of American Beech Regeneration 10 Years following Harvesting. North. J. Appl. For. 2006, 23, 192–196. [Google Scholar] [CrossRef][Green Version]
- Swan, F. Post-Fire Response of Four Plant Communities in South-Central New York State Author. Ecology 1970, 51, 1074–1082. [Google Scholar] [CrossRef]
- Johnson, K.S. Composition of Two Oak-Northern Hardwood Stands 18 Years after Springtime Prescribed Burning. Master’s Thesis, SUNY Coll. Environ. Sci. and For., Syracuse, NY, USA, 2000. [Google Scholar]
- Ramirez, J.I.; Jansen, P.A.; Poorter, L. Effects of wild ungulates on the regeneration, structure and functioning of temperate forests: A semi-quantitative review. For. Ecol. Manag. 2018, 424, 406–419. [Google Scholar] [CrossRef]
- Ramirez, J.I.; Jansen, P.A.; den Ouden, J.; Goudzwaard, L.; Poorter, L. Long-term effects of wild ungulates on the structure, composition and succession of temperate forests. For. Ecol. Manag. 2019, 432, 478–488. [Google Scholar] [CrossRef]







| Strata | Height Class | Definition |
|---|---|---|
| Browsing vulnerable | Seedlings | <25 cm tall |
| Browsing vulnerable saplings | >25–137 cm tall | |
| Sapling recruit | Small sapling recruits | >137 cm tall, <5 cm DBH |
| (i.e., browsing immune) | Large sapling recruits | >5–10 cm DBH |
| Overstory | Poletimber | >10–25.4 cm DBH |
| Sawtimber | >25.4 cm DBH |
| Parameter | Mean (Range) | Significance |
|---|---|---|
| NLP (n = 29) | EUP (n = 40) | |
| Current basal area (m2 ha−1) | 17.9 (8.9–25.7) | 16.5 (7.6–25.2) ns |
| Time since timber sale closing (years) | 5 (3–11) | 6 (1–11) ns |
| Preharvest beech basal area (m2 ha−1) | 15.5 (2.0–31.4) | 10.9 (1.0–22.4) ** |
| Live beech basal area (m2 ha−1) | 5.5 (0–16.7) | 2.0 (0–10.5) *** |
| Seedlings (% cover) | 12 (2–41) | 16 (5–35) ns |
| Herbaceous (% cover) | 7 (0–23) | 6 (0–18) ns |
| Non-tree woody (% cover) | 3 (0–17) | 4 (0–16) ns |
| Leaf litter (% cover) | 87 (63–95) | 89 (72–97) ns |
| Species | Region | Size Class | AICc | R2 | Prob > ChiSquare (Sign of Effects) |
|---|---|---|---|---|---|
| A. saccharum | EUP | Seedlings | 396.4 | 0.63 | H < 0.0001, D = 0.0004 (D+) |
| Browsing vulnerable saplings | 280.7 | 0.49 | H < 0.0001, (D × BA) < 0.0001 | ||
| Small sapling recruits | 219.6 | 0.50 | H < 0.0001 | ||
| Large sapling recruits | 209.3 | 0.27 | H = 0.0035, H × BA = 0.0002, H × D = 0.0036 | ||
| NLP | Seedlings | 317.6 | 0.00 | ||
| Browsing vulnerable saplings | 130.6 | 0.32 | BA (0.0216), BA × H (0.0053), (BA-) | ||
| Small sapling recruits | 59.3 | 0.28 | |||
| Large sapling recruits | 155.0 | 0.24 | D = 0.0009 (D-) | ||
| A. rubrum | EUP | Seedlings | 434.5 | 0.71 | H < 0.0001, BA × H = 0.0070 |
| Browsing vulnerable saplings | 309.6 | 0.58 | H < 0.0001, BA = 0.0019 (BA-) | ||
| Small sapling recruits | 245.6 | 0.52 | H < 0.0001, BA = < 0.0001 (BA-) | ||
| Large sapling recruits | 154.6 | 0.36 | H = 0.0001, H × BA = 0.0188 | ||
| NLP | Seedlings | 307.7 | 0.28 | H < 0.0001 | |
| Browsing vulnerable saplings | 68.7 | 0.08 | |||
| Small sapling recruits | 24.3 | 0.04 | |||
| Large sapling recruits | 25.2 | 0.00 | |||
| F. grandifolia | EUP | Seedlings | 222.4 | 0.31 | H = 0.0009, H × D 0.0104 |
| Browsing vulnerable saplings | 333.6 | 0.29 | H = 0.0012, BA = 0.0898 (BA-) | ||
| Small sapling recruits | 380.2 | 0.35 | H = 0.0073, BA < 0.0001 (BA-) | ||
| Large sapling recruits | 278.7 | 0.62 | H < 0.0001, (H × D) < 0.0001, (H × BA) = 0.0513 | ||
| NLP | Seedlings | 244.2 | 0.26 | H × BA = 0.0700 | |
| Browsing vulnerable saplings | 306.0 | 0.00 | |||
| Small sapling recruits | 282.2 | 0.35 | BA< 0.0001, BA × H < 0.0001 (BA-) | ||
| Large sapling recruits | 158.5 | 0.41 | D × BA = 0.0261 | ||
| O. virginiana | EUP | Seedlings | 60.3 | 0.08 | D = 0.0088 (D-) |
| Browsing vulnerable saplings | 79.5 | 0.30 | H < 0.0001, (H × BA) < 0.0001 | ||
| Small sapling recruits | 104.7 | 0.25 | H = 0.0011 | ||
| Large sapling recruits | 30.6 | 0.21 | H = 0.0006, H × D = 0.0134 | ||
| NLP | Seedlings | 156.6 | 0.16 | D = 0.0186 (D-) | |
| Browsing vulnerable saplings | 230.0 | 0.05 | |||
| Small sapling recruits | 221.6 | 0.02 | |||
| Large sapling recruits | 79.2 | 0.00 |
| Region Size Class | AICc | R2 | Model Terms = p > ChiSquare (Sign of Effect) |
|---|---|---|---|
| EUP | |||
| Seedlings | 220.5. | 0.45 | H = 0.0003, H × D = 0.0027 |
| Browsing vulnerable saplings | 324.0 | 0.40 | PBBA < 0.0001 (+) |
| Small sapling recruits | 379.9 | 0.26 | BA < 0.0018 (-) |
| Large sapling recruits | 273.3 | 0.60 | H < 0.0001, H × D < 0.0001, (H × BA) = 0.0037 |
| NLP | |||
| Seedlings | 244.2 | 0.26 | BA × H = 0.0700 |
| Browsing vulnerable saplings | 296.5 | 0.45 | PBBA = 0.0102 (+) |
| Small sapling recruits | 265.3 | 0.75 | H = 0.0004, PBBA = 0.0002 (+), PBBA × H < 0.0001 |
| Large sapling recruits | 159.1 | 0.39 | D = 0.0007 (−) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elenitsky, L.M.; Walters, M.B.; Farinosi, E.J. Tree Regeneration Structure Following Beech Bark Disease-Motivated Harvests: Factors Associated with Patterns and Management Implications. Forests 2020, 11, 180. https://doi.org/10.3390/f11020180
Elenitsky LM, Walters MB, Farinosi EJ. Tree Regeneration Structure Following Beech Bark Disease-Motivated Harvests: Factors Associated with Patterns and Management Implications. Forests. 2020; 11(2):180. https://doi.org/10.3390/f11020180
Chicago/Turabian StyleElenitsky, Lucas M., Michael B. Walters, and Evan J. Farinosi. 2020. "Tree Regeneration Structure Following Beech Bark Disease-Motivated Harvests: Factors Associated with Patterns and Management Implications" Forests 11, no. 2: 180. https://doi.org/10.3390/f11020180
APA StyleElenitsky, L. M., Walters, M. B., & Farinosi, E. J. (2020). Tree Regeneration Structure Following Beech Bark Disease-Motivated Harvests: Factors Associated with Patterns and Management Implications. Forests, 11(2), 180. https://doi.org/10.3390/f11020180