Biomass Yield of 37 Different SRC Poplar Varieties Grown on a Typical Site in North Eastern Germany



Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description, Experimental Design and Plant Material
2.2. Determination of Growth Parameters
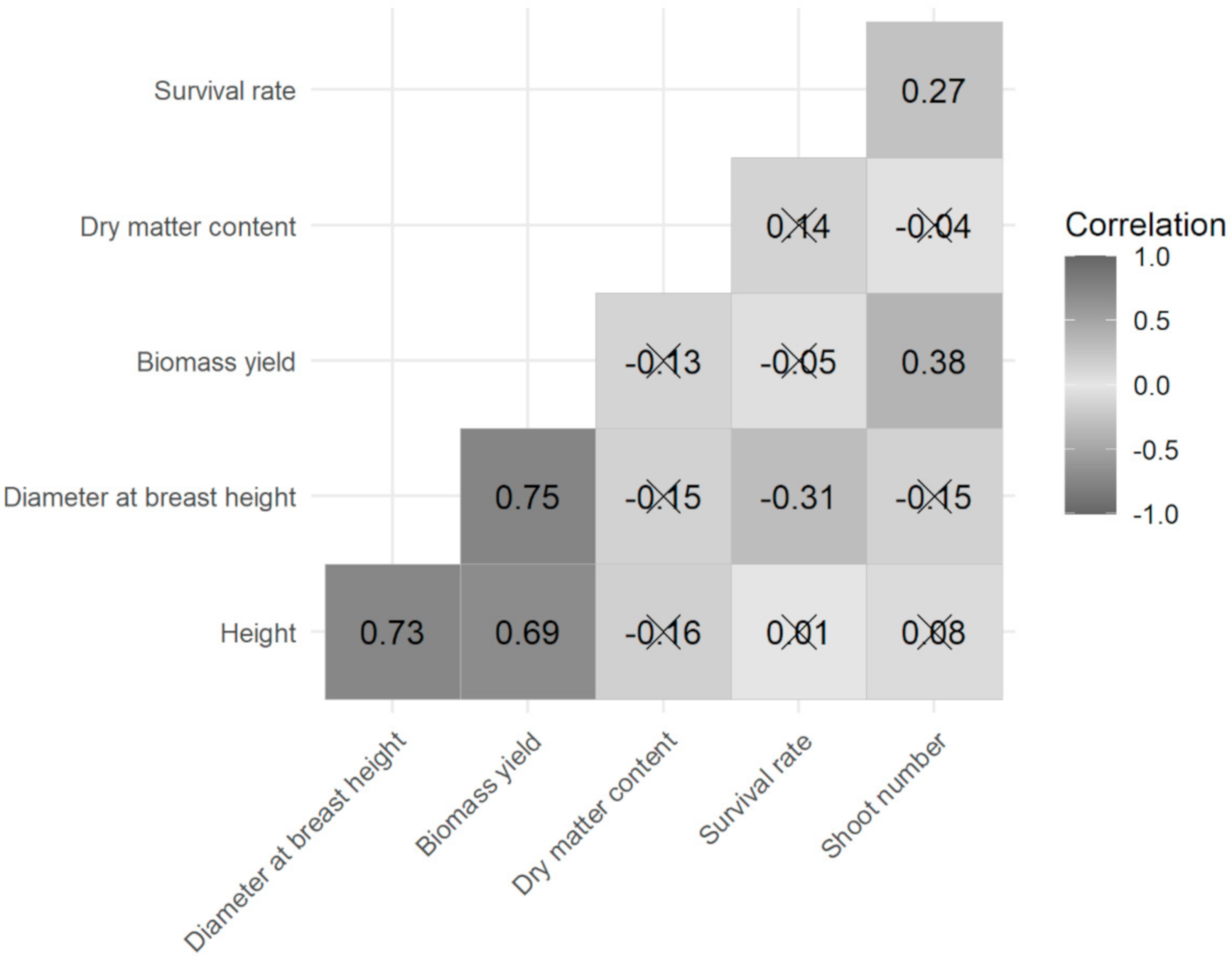
2.3. Data Analysis
3. Results
3.1. Survival Rate
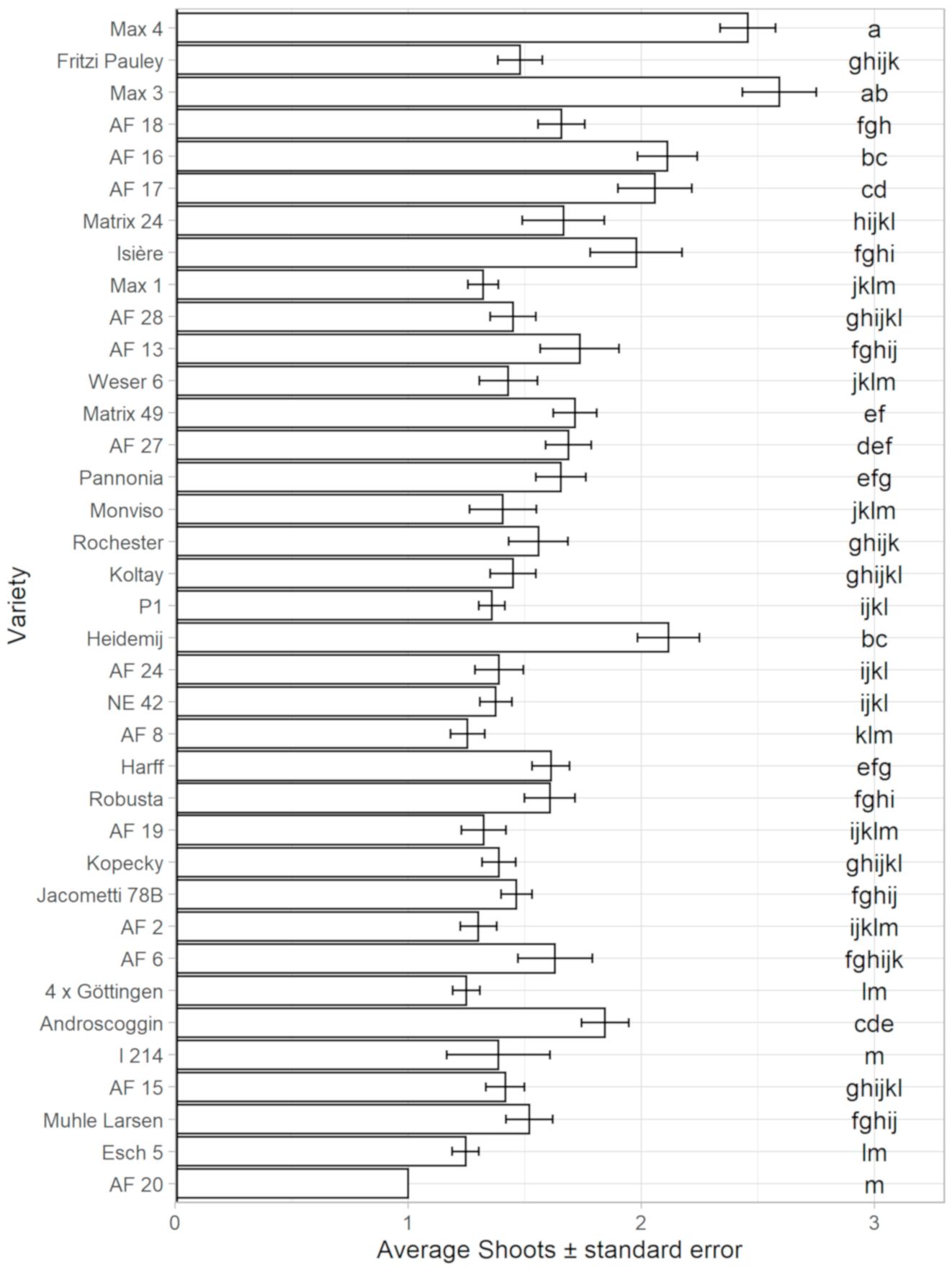
3.2. Resprouting Capacity
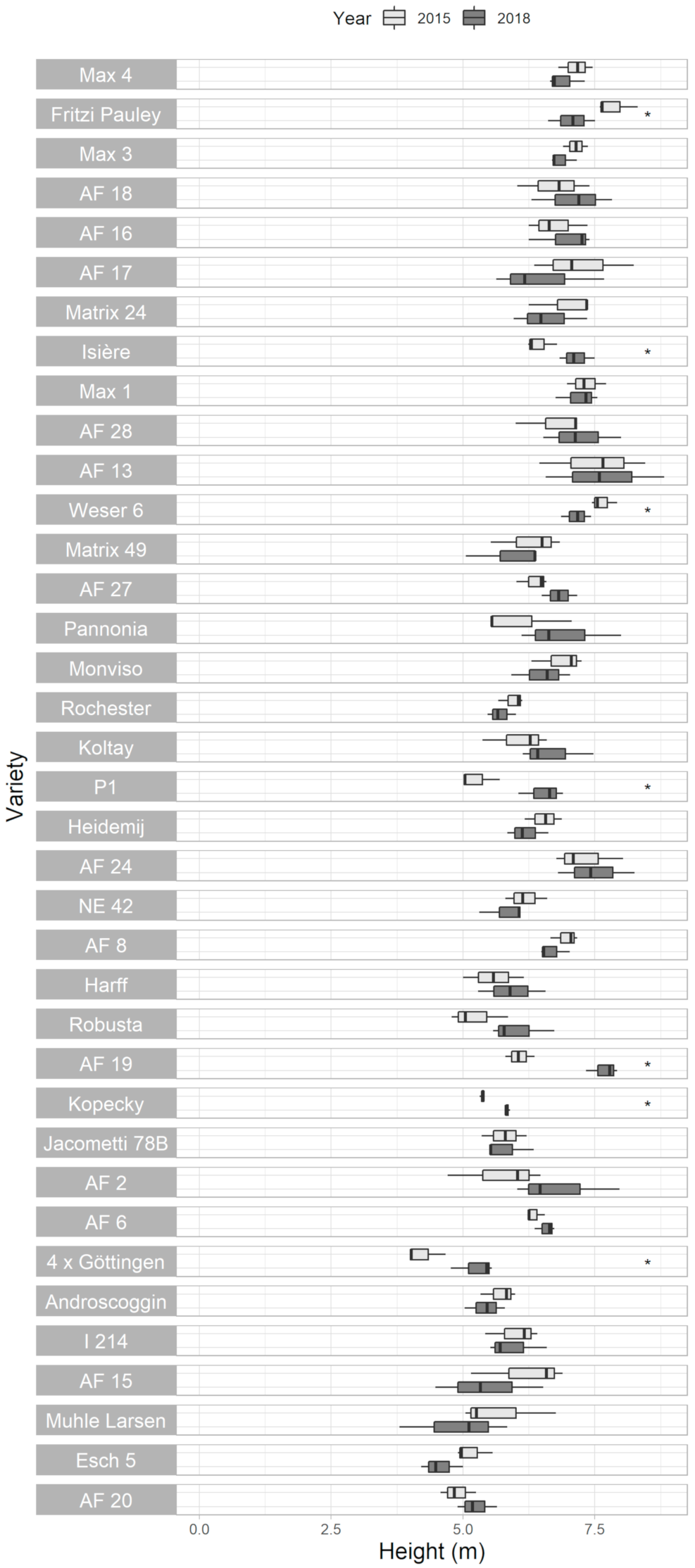
3.3. Plant Height
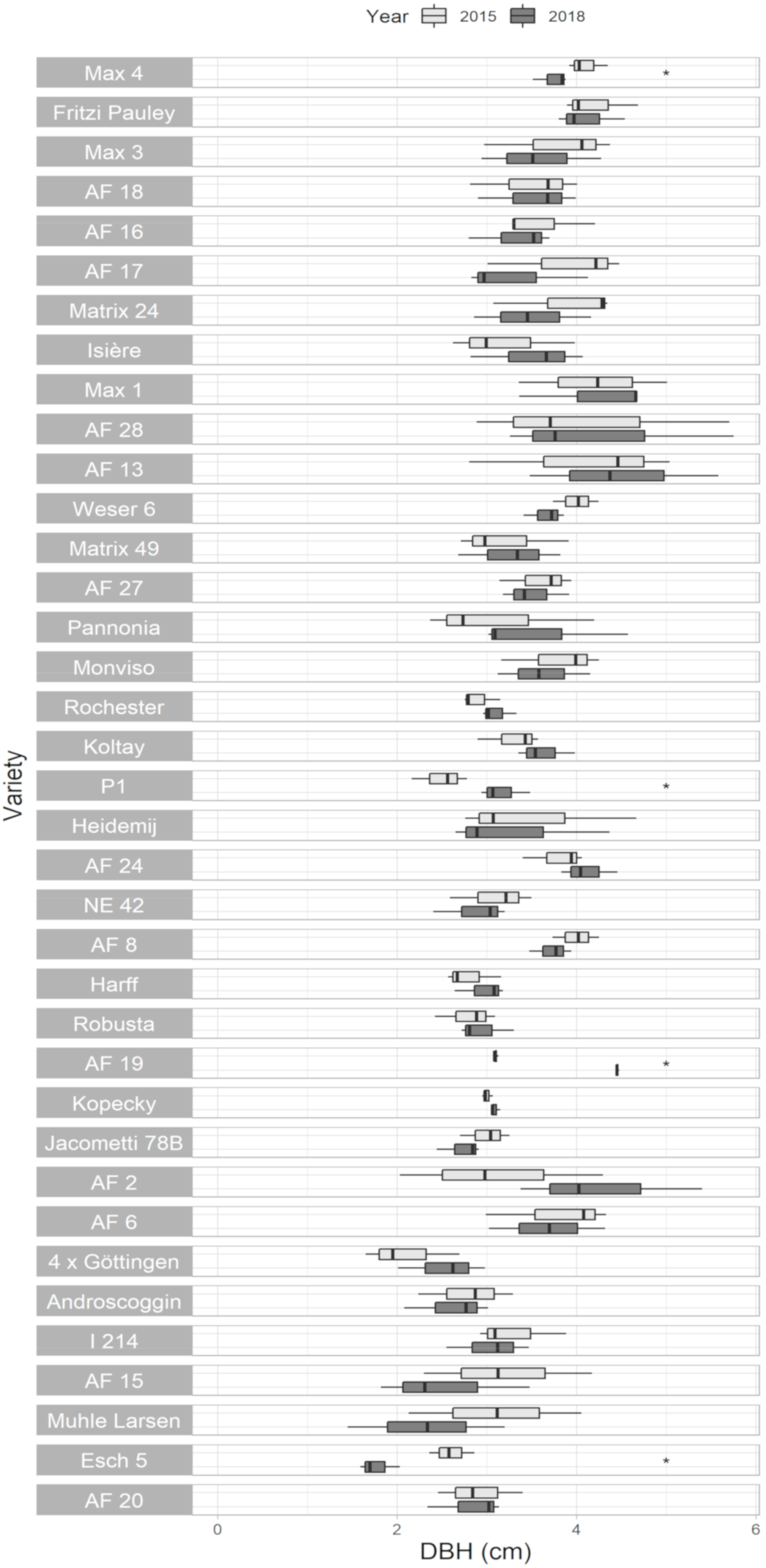
3.4. Diameter at Breast Height (DBH)
3.5. Dry Matter Content (DMC)
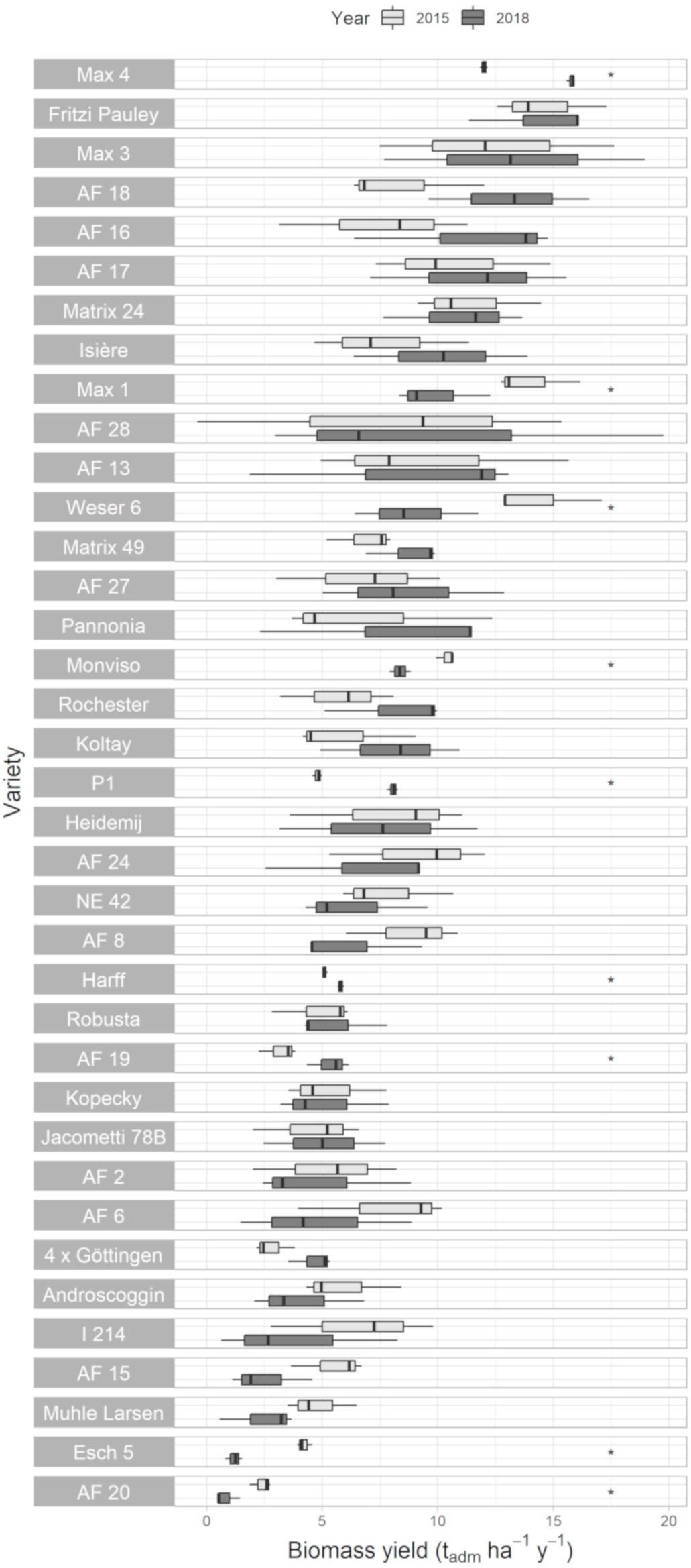
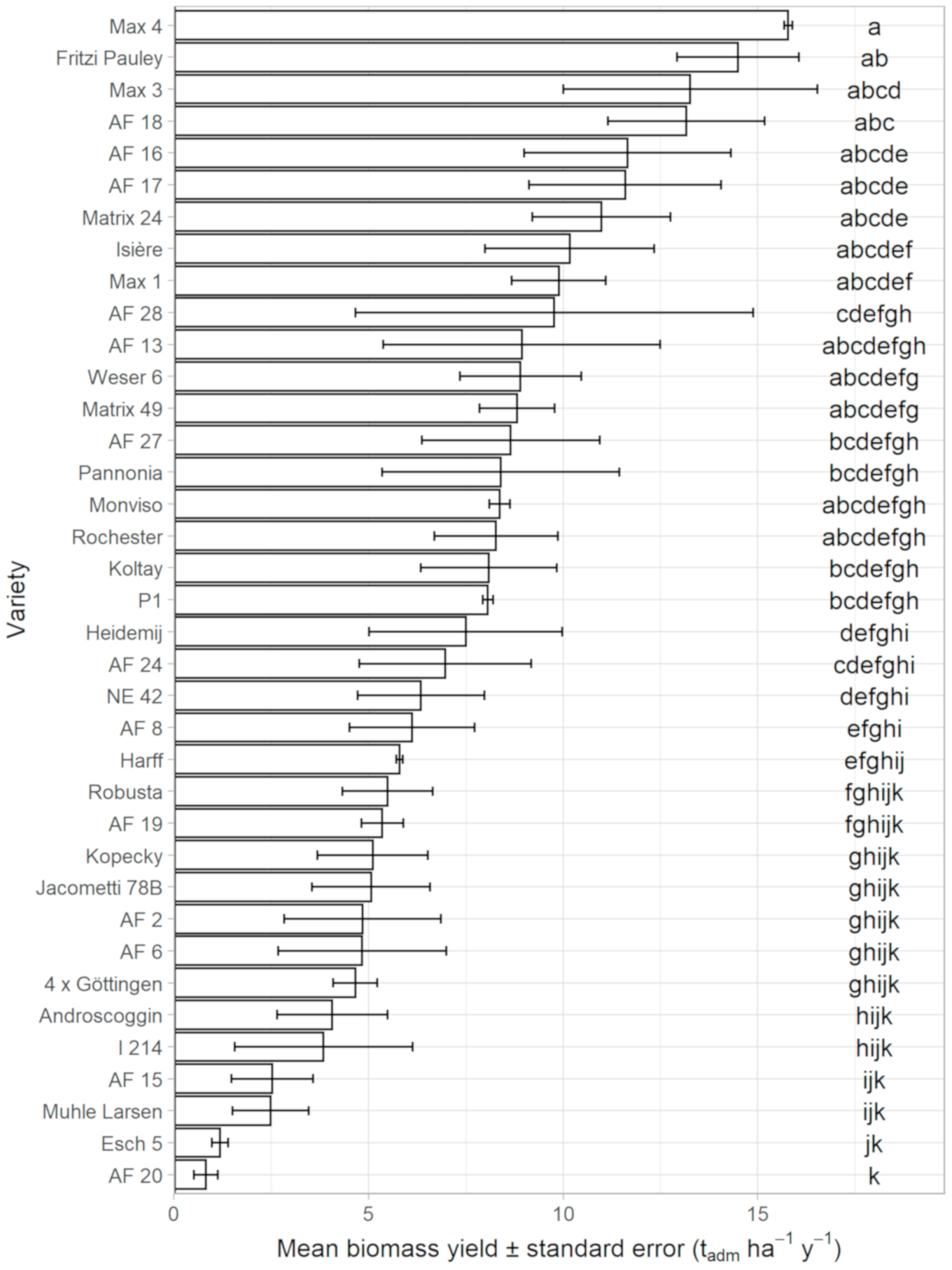
3.6. Dry Matter Biomass Yield (DBY)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Commission. A Policy Framework for Climate and Energy in the Period from 2020 up to 2030. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions Brussels 2014: European Commission. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:52014DC0015&from=EN (accessed on 4 February 2020).
- Niemczyk, M.; Kaliszewski, A.; Jewiarz, M.; Wróbel, M.; Mudryk, K. Productivity and biomass characteristics of selected poplar (Populus spp.) cultivars under the climatic conditions of northern Poland. Biomass Bioenergy 2018, 111, 46–51. [Google Scholar] [CrossRef]
- Tripathi, A.M.; Pohanková, E.; Fischer, M.; Orság, M.; Trnka, M.; Klem, K.; Marek, M.V. The Evaluation of Radiation Use Efficiency and Leaf Area Index Development for the Estimation of Biomass Accumulation in Short Rotation Poplar and Annual Field Crops. Forests 2018, 9, 168. [Google Scholar] [CrossRef]
- Hauk, S.; Wittkopf, S.; Knoke, T. Analysis of commercial short rotation coppices in Bavaria, southern Germany. Biomass Bioenergy 2014, 67, 401–412. [Google Scholar] [CrossRef]
- Hauk, S.; Gandorfer, M.; Wittkopf, S.; Müller, U.K.; Knoke, T. Ecological diversification is risk reducing and economically profitable—The case of biomass production with short rotation woody crops in south German land-use portfolios. Biomass Bioenergy 2017, 98, 142–152. [Google Scholar] [CrossRef]
- Schweier, J.; Arranz, C.; Nock, C.A.; Jaeger, D.; Scherer-Lorenzen, M. Impact of Increased Genotype or Species Diversity in Short Rotation Coppice on Biomass Production and Wood Characteristics. Bioenergy Res. 2019, 12, 497–508. [Google Scholar] [CrossRef]
- Bacenetti, J.; Bergante, S.; Facciotto, G.; Fiala, M. Woody biofuel production from short rotation coppice in Italy: Environmental-impact assessment of different species and crop management. Biomass Bioenergy 2016, 94, 209–219. [Google Scholar] [CrossRef]
- Agostini, F.; Gregory, A.S.; Richter, G.M. Carbon Sequestration by Perennial Energy Crops: Is the Jury Still Out? Bioenergy Res. 2015, 8, 1057–1080. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, D.; Setzer, F. Kurzumtriebsplantagen: Holz vom Acker—So Geht’s; DLG-Verlag: Frankfurt, Germany, 2012; p. 71. ISBN 978-3-7690-2005-2. (In Germany) [Google Scholar]
- Berhongaray, G.; Verlinden, M.S.; Broeckx, L.S.; Janssens, I.A.; Ceulemans, R. Soil carbon and belowground carbon balance of a short-rotation coppice: Assessments from three different approaches. GCB Bioenergy 2016, 9, 299–313. [Google Scholar] [CrossRef]
- Ferrè, C.; Comolli, R. Comparison of soil CO2 emissions between short-rotation coppice poplar stands and arable lands. iForest Biogeosci. For. 2018, 11, 199–205. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation (EC) No 1120/2009 of 29 October 2009 Laying Down Detailed Rules for the Implementation of the Single Payment Scheme Provided for in Title III of Council Regulation (EC) No 73/2009 Establishing Common Rules for Direct Support Schemes for Farmers under the Common Agricultural Policy and Establishing Certain Support Schemes for Farmers. 2009. Available online: https://op.europa.eu/de/publication-detail/-/publication/45b679d0-d9b9-400a-904b-8df1724e00e4/language-en (accessed on 4 February 2020).
- Carl, C.; Biber, P.; Veste, M.; Landgraf, D.; Pretzsch, H. Key drivers of competition and growth partitioning among Robinia pseudoacacia L. trees. For. Ecol. Manag. 2018, 430, 86–93. [Google Scholar] [CrossRef]
- Nicolescu, V.-N.; Buzatu-Goanță, C.; Bakti, B.; Keserű, Z.; Antal, B.; Rédei, K. Black locust (Robinia pseudoacacia L.) as a multi-purpose tree species in Hungary and Romania: A review. J. For. Res. 2018, 29, 1449–1463. [Google Scholar] [CrossRef]
- Vítková, M.; Sádlo, J.; Roleček, J.; Petřík, P.; Sitzia, T.; Müllerová, J.; Pyšek, P. Robinia pseudoacacia-dominated vegetation types of Southern Europe: Species composition, history, distribution and management. Sci. Total Environ. 2020, 707, 134857. [Google Scholar] [CrossRef]
- Larsen, S.U.; Jørgensen, U.; Lærke, P.E. Biomass yield, nutrient concentration and nutrient uptake by SRC willow cultivars grown on different sites in Denmark. Biomass Bioenergy 2018, 116, 161–170. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Olba-Zięty, E.; Rosenqvist, H.; Krzyżaniak, M. Economic efficiency of willow, poplar and black locust production using different soil amendments. Biomass Bioenergy 2017, 106, 74–82. [Google Scholar] [CrossRef]
- Rogers, E.; Zalesny, J.R.S.; Hallett, R.A.; Headlee, W.L.; Wiese, A.H. Relationships among Root–Shoot Ratio, Early Growth, and Health of Hybrid Poplar and Willow Clones Grown in Different Landfill Soils. Forests 2019, 10, 49. [Google Scholar] [CrossRef]
- Rebola-Lichtenberg, J.; Schall, P.; Annighöfer, P.; Ammer, C.; Leinemann, L.; Polle, A.; Euring, D. Mortality of Different Populus Genotypes in Recently Established Mixed Short Rotation Coppice with Robinia pseudoacacia L. Forests 2019, 10, 410. [Google Scholar] [CrossRef]
- Verlinden, M.S.; Broeckx, L.; Bulcke, J.V.D.; Van Acker, J.; Ceulemans, R. Comparative study of biomass determinants of 12 poplar (Populus) genotypes in a high-density short-rotation culture. For. Ecol. Manag. 2013, 307, 101–111. [Google Scholar] [CrossRef]
- González-González, B.D.; Oliveira, N.; González, I.; Cañellas, I.; Sixto, H. Poplar biomass production in short rotation under irrigation: A case study in the Mediterranean. Biomass Bioenergy 2017, 107, 198–206. [Google Scholar] [CrossRef]
- Oliveira, N.; Rodríguez-Soalleiro, R.; Hernández, M.J.; Cañellas, I.; Sixto, H.; Pérez-Cruzado, C. Improving biomass estimation in a Populus short rotation coppice plantation. For. Ecol. Manag. 2017, 391, 194–206. [Google Scholar] [CrossRef]
- Schlepphorst, H.; Hartmann, H.; Murach, D. Yield of fast-growing tree species in northeast Germany: Results of research with experimental plots (2006 to 2015). Appl. Agric. For. Res. 2017, 2, 93–110. [Google Scholar] [CrossRef]
- Oliveira, N.; Rodríguez-Soalleiro, R.; Pérez-Cruzado, C.; Cañellas, I.; Sixto, H.; Ceulemans, R. Above- and below-ground carbon accumulation and biomass allocation in poplar short rotation plantations under Mediterranean conditions. For. Ecol. Manag. 2018, 428, 57–65. [Google Scholar] [CrossRef]
- Goetsch, S.; Peek, R. Vergleichende Feuchtebestimmung an Bohrkernen und geschnittenen Holzproben nach DIN 52 183 Holz als Roh-und Werkstoff. Eur. J. Wood Prod. 1987, 45, 300. [Google Scholar] [CrossRef]
- Hartmann, K.-U. Entwicklung eines Ertragsschätzers für Kurzumtriebsbestände aus Pappel. Ph.D. Thesis, Technische Universität Dresden, Dresden, Germany, 12 August 2010. Available online: https://nbn-resolving.org/urn:nbn:de:bsz:14-qucosa-62853 (accessed on 19 February 2020).
- Ali, M.S.W. Modelling of Biomass Production Potential of Poplar in Short Rotation Plantations on Agricultural Lands of Saxony, Germany. Ph.D. Thesis, Technische Universität Dresden, Dresden, Germany, 3 March 2009. Available online: https://nbn-resolving.org/urn:nbn:de:bsz:14-ds-1237199867841-24821 (accessed on 19 February 2020).
- Kramer, H.; Akça, A. Leitfaden zur Waldmesslehre; J.D. Sauerländer Verlag: Bad Orb, Germany, 2008; ISBN 9783793908807. [Google Scholar]
- R-Core-Team. R: A language and Environment for Statistical Computing. (3.6.3). R Foundation for Statistical Computing. 2020. Available online: https://www.r-project.org/ (accessed on 23 September 2020).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; D’Agostino McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2017, 4, 1686. [Google Scholar] [CrossRef]
- Kassambra, A. Ggcorrplot: Visualization of a Correlation Matrix Using “ggplot2” (0.1.3). 2019. Available online: https://cran.r-project.org/package=ggcorrplot (accessed on 23 September 2020).
- Hartmann, L.; Lamersdorf, N. Site conditions, initial growth and nutrient and litter cycling of newly installed short rotation coppice and agroforestry systems. In Bioenergy from Dendromass for the Sustainable Development of Rural Areas; Butler Manning, D., Bemmann, A., Bredemeier, M., Lamersdorf, N., Ammer, C., Eds.; Wiley-VCH: Weinheim, Germany, 2015; pp. 123–138. [Google Scholar]
- Shifflett, S.D.; Hazel, D.W.; Nichols, E.G. Sub-Soiling and Genotype Selection Improves Populus Productivity Grown on a North Carolina Sandy Soil. Forests 2016, 7, 74. [Google Scholar] [CrossRef]
- Friedrich, E. Anbautechnische Untersuchungen in forstlichen Schnellwuchsplantagen und Demonstration des Leistungsvermögens schnellwachsender Baumarten. In Modellvorhaben “Schnellwachsende Baumarten”; Hofmann, M., Ed.; Zusammenfassender Abschlussbericht; Landwirtschaftsverlag GmbH: Münster, Germany, 1999; pp. 19–150. Available online: https://www.gbv.de/dms/bs/toc/303247398.pdf (accessed on 23 September 2020).
- Röhle, H.; Horn, H.; Müller, M.; Skibbe, K. Site-based yield estimation and biomass calculation in short rotation coppice plantations. In Bioenergy from Dendromass for the Sustainable Development of Rural Areas; Butler Manning, D., Bemmann, A., Bredemeier, M., Lamersdorf, N., Ammer, C., Eds.; Wiley-VCH: Weinheim, Germany, 2015; pp. 173–186. [Google Scholar]
- Paris, P.; Mareschi, L.; Sabatti, M.; Pisanelli, A.; Ecosse, A.; Nardin, F.; Scarascia-Mugnozza, G. Comparing hybrid Populus clones for SRF across northern Italy after two biennial rotations: Survival, growth and yield. Biomass Bioenergy 2011, 35, 1524–1532. [Google Scholar] [CrossRef]
- Larson, P.R.; Isebrands, J.G. The Plastochron Index as Applied to Developmental Studies of Cottonwood. Can. J. For. Res. 1971, 1, 1–11. [Google Scholar] [CrossRef]
- Ceulemans, R.; Deraedt, W. Production physiology and growth potential of poplars under short-rotation forestry culture. For. Ecol. Manag. 1999, 121, 9–23. [Google Scholar] [CrossRef]
- Scarascia-Mugnozza, G.E.; Ceulemans, R.; Heilman, P.E.; Isebrands, J.G.; Stettler, R.F.; Hinckley, T.M. Production physiology and morphology of Populus species and their hybrids grown under short rotation. II. Biomass components and harvest index of hybrid and parental species clones. Can. J. For. Res. 1997, 27, 285–294. [Google Scholar] [CrossRef]
- Marron, N.; Villar, M.; Dreyer, E.; DeLay, D.; Boudouresque, E.; Petit, J.-M.; Delmotte, F.M.; Guehl, J.-M.; Brignolas, F. Diversity of leaf traits related to productivity in 31 Populus deltoides × Populus nigra clones. Tree Physiol. 2005, 25, 425–435. [Google Scholar] [CrossRef]
- Monclus, R.; Dreyer, E.; Delmotte, F.M.; Villar, M.; DeLay, D.; Boudouresque, E.; Petit, J.-M.; Marron, N.; Bréchet, C.; Brignolas, F. Productivity, leaf traits and carbon isotope discrimination in 29 Populus deltoides × P. nigra clones. New Phytol. 2005, 167, 53–62. [Google Scholar] [CrossRef]
- Aylott, M.J.; Casella, E.; Tubby, I.; Street, N.R.; Smith, P.; Taylor, G. Yield and spatial supply of bioenergy poplar and willow short?rotation coppice in the UK. New Phytol. 2008, 178, 358–370. [Google Scholar] [CrossRef]
- Aylott, M.J.; Casella, E.; Tubby, I.; Street, N.R.; Smith, P.; Taylor, G. Corrigendum. New Phytol. 2008, 178, 897. [Google Scholar] [CrossRef]
- Gielen, B.; Ceulemans, R. The likely impact of rising atmospheric CO2 on natural and managed Populus: A literature review. Environ. Pollut. 2001, 115, 335–358. [Google Scholar] [CrossRef]
- Cochard, H.; Casella, E.; Mencuccini, M. Xylem vulnerability to cavitation varies among poplar and willow clones and correlates with yield. Tree Physiol. 2007, 27, 1761–1767. [Google Scholar] [CrossRef] [PubMed]
- Rae, A.M.; Street, N.R.; Robinson, K.M.; Harris, N.; Taylor, G. Five QTL hotspots for yield in short rotation coppice bioenergy poplar: The Poplar Biomass Loci. BMC Plant Biol. 2009, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Dillen, S.; Djomo, S.N.; Al Afas, N.; Vanbeveren, S.P.; Ceulemans, R. Biomass yield and energy balance of a short-rotation poplar coppice with multiple clones on degraded land during 16 years. Biomass Bioenergy 2013, 56, 157–165. [Google Scholar] [CrossRef]
- Landgraf, D.; Bärwolff, M.; Burger, F.; Pecenka, R.; Hering, T.; Schweier, J. Produktivität, Management und Nutzung von Agrarholz, In Agrarholz—Schnellwachsende Bäume in der Landwirtschaft Biologie—Ökologie—Management; Veste, M., Böhm, C., Eds.; Springer Spektrum: Berlin, Germany, 2018; ISBN 978-3-662-49930-6. [Google Scholar] [CrossRef]
- Biertümpfel, A.; Hering, T. Energieholzanbau in Thüringen—Erträge und Praxiserfahrungen. In Erfurter Tagung Schnellwachsender Baumarten; Landgraf, D., Ed.; 2017; ISSN 2567–8922. Available online: https://www.researchgate.net/publication/326774259_1_ERFURTER_TAGUNG_Schnellwachsende_Baumarten_--_Etablierung_Management_und_Verwertung_vom_1611_-_17112017 (accessed on 8 May 2020).
- Nielsen, U.B.; Madsen, P.; Hansen, J.K.; Nord-Larsen, T.; Nielsen, A.T. Production potential of 36 poplar clones grown at medium length rotation in Denmark. Biomass Bioenergy 2014, 64, 99–109. [Google Scholar] [CrossRef]
- Benetka, V.; Novotná, K.; Štochlová, P. Biomass production of Populus nigra L. clones grown in short rotation coppice systems in three different environments over four rotations. iForest Biogeosci. For. 2014, 7, 233–239. [Google Scholar] [CrossRef]
- Verlinden, M.S.; Broeckx, L.; Ceulemans, R. First vs. second rotation of a poplar short rotation coppice: Above-ground biomass productivity and shoot dynamics. Biomass Bioenergy 2015, 73, 174–185. [Google Scholar] [CrossRef]
- Steenackers, V.; Strobl, S.; Steenackers, M. Collection and distribution of poplar species, hybrids and clones. Biomass 1990, 22, 1–20. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Parentage | Sex | Source |
|---|---|---|---|
| Androscoggin | P. maximowiczii × P. trichocarpa | m | P & P |
| Fritzi Pauley | P. trichocarpa | f | |
| Harff | P. × euramericana | f | |
| Heidemij | P. × euramericana | m | |
| I 214 | P. deltoides × P. nigra | f | |
| Isières | P. × euramericana | m | |
| Jacometti 78 B | P. × euramericana | f | |
| Koltay | P. × euramericana | m | |
| Kopecky | P. × euramericana | m | |
| Matrix 24 | P. maximowiczii × P. trichocarpa | ||
| Matrix 49 | P. maximowiczii × P. trichocarpa | ||
| Max 1 | P. nigra × P. maximowiczii | f | |
| Max 3 | P. nigra × P. maximowiczii | f | |
| Max 4 | P. nigra × P. maximowiczii | f | |
| Monviso | P. generosa × P. nigra | f | |
| Muhle Larsen | P. trichocarpa | f | |
| NE 42 * | P. maximowiczii × P. trichocarpa | m | |
| Pannonia | P. × euramericana | f | |
| Rochester | P. maximowicii × P. nigra | f | |
| Robusta | P. × euramericana | m | |
| Weser 6 | P. trichocarpa | ||
| AF 2 | P. × canadensis Moench | m | AFV |
| AF 6 | P. generosa × P. nigra A. Henry | f | |
| AF 8 | P. trichocarpa × P. × generosa | f | |
| AF 13 | P. × canadensis | ||
| AF 15 | P. deltoides × P. nigra | ||
| AF 16 | P. × canadensis | ||
| AF 17 | P. deltoides × P. nigra | ||
| AF 18 | P. deltoides × P. nigra | ||
| AF 19 | P. deltoides × P. nigra | ||
| AF 20 | P. deltoides × P. nigra | ||
| AF 24 | P. deltoides × P. nigra | ||
| AF 27 | P. deltoides × P. nigra | ||
| AF 28 | P. deltoides × P. nigra | ||
| P1 | P. × canescens | TI (UG/INRA 717-1B4) | |
| 4 × Göttingen | P. × canescens (tetraploid) | ||
| Esch 5 | P. tremula × P. tremuloides |
| Survival Rate [%] | |||
|---|---|---|---|
| Variety | 2013 | 2015 | 2018 |
| Max 4 | 98 | 95 | 95 |
| Fritzi Pauley | 84 | 80 | 80 |
| Max 3 | 95 | 83 | 83 |
| AF 18 | 96 | 84 | 84 |
| AF 16 | 77 | 72 | 72 |
| AF 17 | 95 | 86 | 86 |
| Matrix 24 | 91 | 72 | 67 |
| Isières | 98 | 97 | 97 |
| Max 1 | 97 | 95 | 95 |
| AF 28 | 81 | 52 | 52 |
| AF 13 | 90 | 54 | 54 |
| Weser 6 | 96 | 87 | 87 |
| Matrix 49 | 89 | 81 | 81 |
| AF 27 | 95 | 62 | 62 |
| Pannonia | 93 | 64 | 63 |
| Monviso | 93 | 61 | 61 |
| Rochester | 95 | 82 | 82 |
| Koltay | 91 | 56 | 52 |
| P1 | 100 | 96 | 96 |
| Heidemij | 94 | 61 | 61 |
| AF 24 | 94 | 45 | 45 |
| NE 42 | 96 | 87 | 87 |
| AF 8 | 83 | 52 | 52 |
| Harff | 94 | 80 | 76 |
| Robusta | 90 | 75 | 75 |
| AF 19 | 57 | 37 | 37 |
| Kopecky | 96 | 61 | 60 |
| Jacometti 78 B | 88 | 72 | 72 |
| AF 2 | 96 | 43 | 43 |
| AF 6 | 85 | 46 | 46 |
| 4 × Göttingen | 98 | 85 | 85 |
| Androscoggin | 83 | 59 | 59 |
| I 214 | 96 | 62 | 62 |
| AF 15 | 91 | 49 | 48 |
| Muhle Larsen | 73 | 52 | 52 |
| Esch 5 | 93 | 74 | 74 |
| AF 20 | 56 | 17 | 17 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Landgraf, D.; Carl, C.; Neupert, M. Biomass Yield of 37 Different SRC Poplar Varieties Grown on a Typical Site in North Eastern Germany. Forests 2020, 11, 1048. https://doi.org/10.3390/f11101048
Landgraf D, Carl C, Neupert M. Biomass Yield of 37 Different SRC Poplar Varieties Grown on a Typical Site in North Eastern Germany. Forests. 2020; 11(10):1048. https://doi.org/10.3390/f11101048
Chicago/Turabian StyleLandgraf, Dirk, Christin Carl, and Markus Neupert. 2020. "Biomass Yield of 37 Different SRC Poplar Varieties Grown on a Typical Site in North Eastern Germany" Forests 11, no. 10: 1048. https://doi.org/10.3390/f11101048
APA StyleLandgraf, D., Carl, C., & Neupert, M. (2020). Biomass Yield of 37 Different SRC Poplar Varieties Grown on a Typical Site in North Eastern Germany. Forests, 11(10), 1048. https://doi.org/10.3390/f11101048