Response of Photosynthesis and Chlorophyll Fluorescence Parameters of Castanopsis kawakamii Seedlings to Forest Gaps

 ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Study Site and Species
2.2. Experimental Design
2.2.1. Leaf Chlorophyll Contents
2.2.2. Leaf Net Photosynthesis Rate
2.2.3. Leaf Chlorophyll Fluorescence
2.3. Data Analysis
3. Results
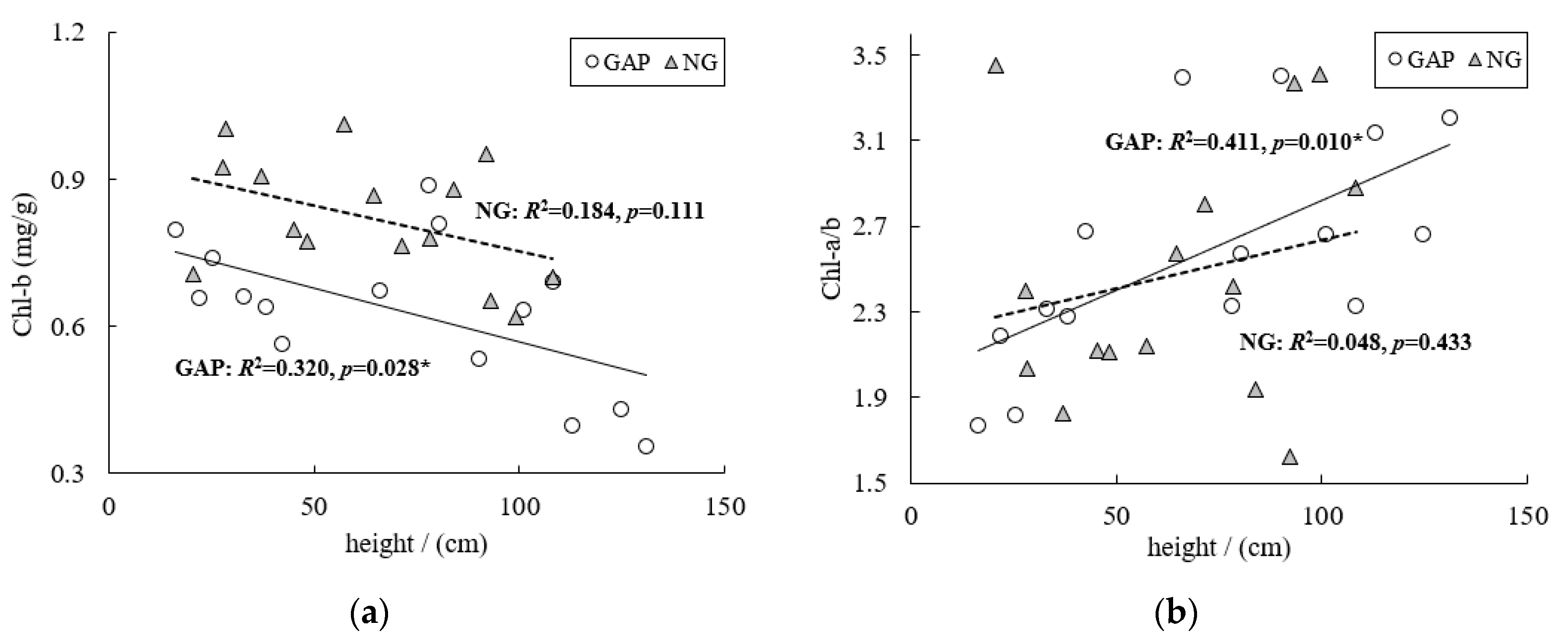
3.1. Leaf Chlorophyll Contents of C. kawakamii Seedlings
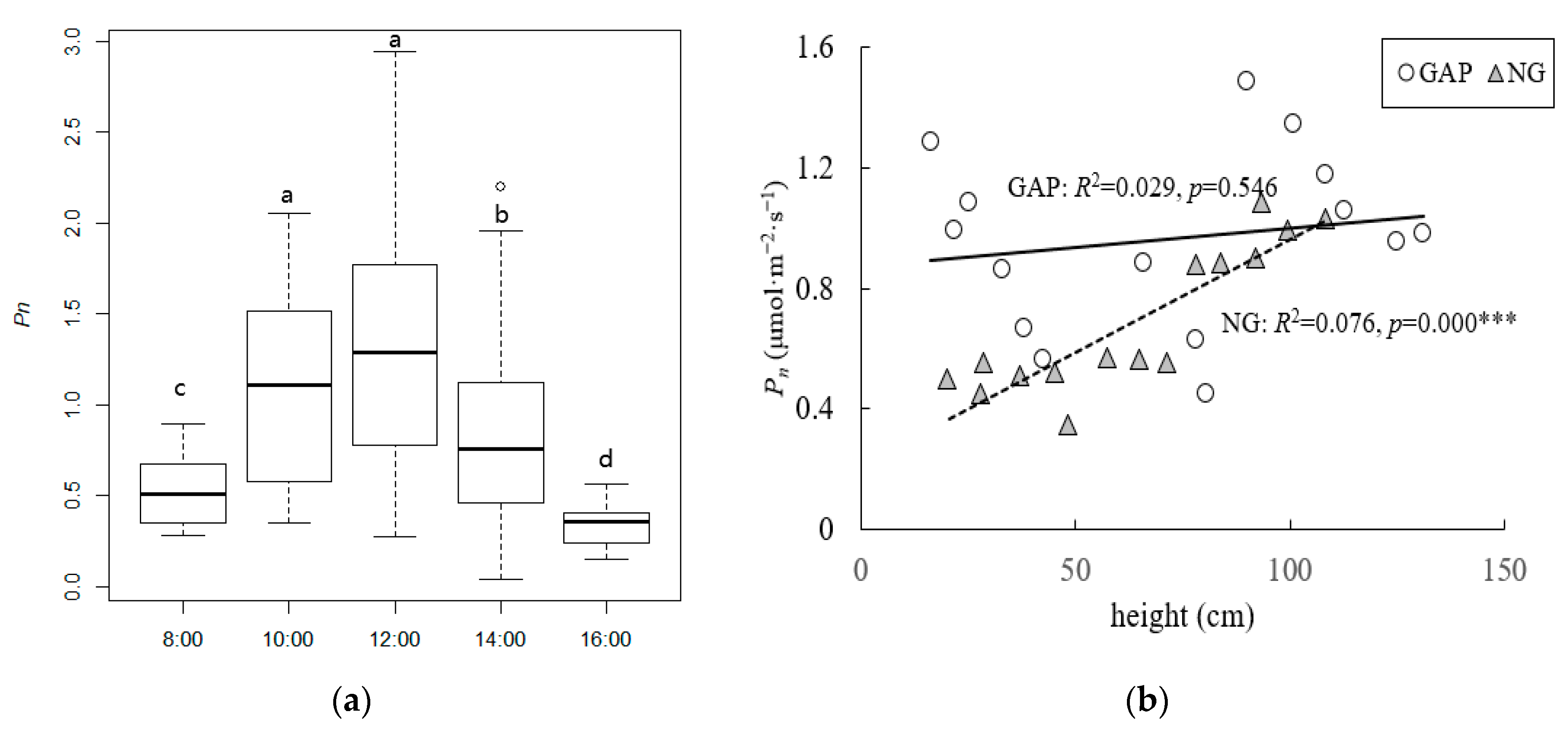
3.2. Pn of C. kawakamii Seedlings
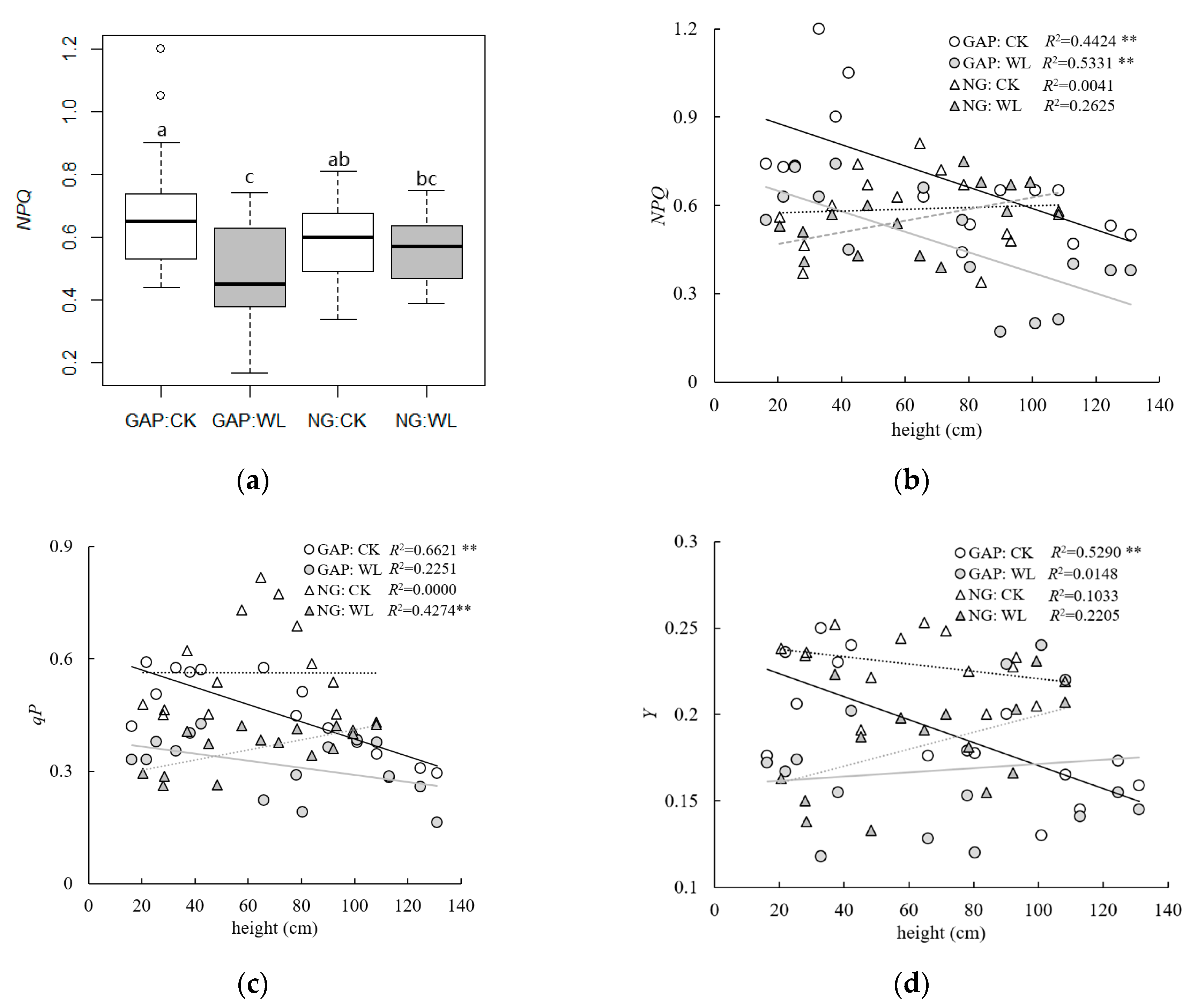
3.3. Chlorophyll Fluorescence Parameters of C. kawakamii Seedlings
4. Discussion
4.1. Leaf Chlorophyll Contents of C. kawakamii Seedlings
4.2. Net Photosynthetic Rate of C. kawakamii Seedlings in Forest Gaps and Non-Gaps
4.3. Chlorophyll Fluorescence Parameters of C. kawakamii Seedlings
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nicotra, A.B.; Chazdon, R.L.; Iriarte, S.V.B. Spatial heterogeneity of light and woody seedling regeneration in tropical wet forests. Ecology 1999, 80, 1908–1926. [Google Scholar] [CrossRef]
- Li, Y.; Tian, X.M.; Wei, M.; Shi, Q.H.; Yang, F.J.; Wang, X.F. Mechanisms of tolerance differences in cucumber seedlings grafted on rootstocks with different tolerance to low temperature and weak light stresses. Turk. J. Bot. 2015, 39, 606–614. [Google Scholar] [CrossRef]
- Qian, C.J.; Zhang, W.; Zhong, X.M.; Li, F.H.; Shi, Z.S. Comparative studies on the photosynthetic characteristics of two maize (Zea mays L.) near-isogenic lines differing in their susceptibility to low light intensity. Emir. J. Food Agric. 2017, 29, 300–311. [Google Scholar] [CrossRef]
- Sui, X.L.; Mao, S.L.; Wang, L.H.; Zhang, B.X.; Zhang, Z.X. Effect of low light on the characteristics of photosynthesis and chlorophyll a fluorescence during leaf development of sweet pepper. J. Integr. Agric. 2012, 11, 1633–1643. [Google Scholar] [CrossRef]
- Park, S.G.; Matsumoto, M. A Study on the effects of light conditions on the longevity and characteristics of Daphniphyllum macropodum leaves. J. Fac. Agric. Kyushu Univ. 2018, 63, 15–19. [Google Scholar]
- Ding, X.T.; Jiang, Y.P.; Wang, H.; Jin, H.J.; Zhang, H.M.; Chen, C.H.; Yu, J.Z. Effects of cytokinin on photosynthetic gas exchange, chlorophyll fluorescence parameters, antioxidative system and carbohydrate accumulation in cucumber (Cucumis sativus L.) under low light. Acta Physiol. Plant. 2013, 35, 1427–1438. [Google Scholar] [CrossRef]
- Wang, J.; Huang, H.J.; Jia, S.; Zhong, X.M.; Li, F.H.; Zhang, K.Y.; Shi, Z.S. Photosynthesis and chlorophyll fluorescence reaction to different shade stresses of weak light sensitive maize. Pak. J. Bot. 2017, 49, 1681–1688. [Google Scholar]
- Wang, M.; Jiang, W.J.; Yu, H.J. Effects of exogenous epibrassinolide on photosynthetic characteristics in tomato (Lycopersicon esculentum Mill) seedlings under weak light stress. J. Agric. Food Chem. 2010, 58, 3642–3645. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Yuan, C.H.; Han, W.; Li, Y.X.; Xiao, F. Effects of low irradiation on photosynthesis and antioxidant enzyme activities in cucumber during ripening stage. Photosynthetica 2016, 54, 251–258. [Google Scholar] [CrossRef]
- Liang, F.; Zheng, C.S.; Sun, X.Z.; Wang, W.L. Effects of low temperature and weak light stress and its recovery on the photosynthesis and chlorophyll fluorescence parameters of cut flower chrysanthemum. Chin. J. Appl. Ecol. 2010, 21, 29–35. [Google Scholar] [CrossRef]
- Bi, H.G.; Liu, P.P.; Jiang, Z.S.; Ai, X.Z. Overexpression of the rubisco activase gene improves growth and low temperature and weak light tolerance in Cucumis sativus. Physiol. Plant. 2017, 161, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.J.; Shen, Z.G.; Liu, Y.; Wang, L.L.; Hannaway, D.; Lu, H.F. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg. Environ. Exp. Bot. 2009, 65, 177–182. [Google Scholar] [CrossRef]
- Liu, J.F.; He, Z.S.; Hong, W.; Zheng, S.Q.; Wang, Z.J. Conservation ecology of endangered plant Castanopsis kawakamii. J. Beijing For. Univ. 2011, 33, 136–143. [Google Scholar] [CrossRef]
- He, Z.S.; Liu, J.F.; Wu, C.T.; Zheng, S.Q.; Hong, W.; Su, S.J.; Wu, C.Z. Effects of forest gaps on some microclimate variables in Castanopsis kawakamii natural forest. J. Mt. Sci. 2012, 9, 706–714. [Google Scholar] [CrossRef]
- Buajan, S.; Liu, J.F.; He, Z.S.; Feng, X.P.; Muhammad, A. Effects of gap size and locations on the regeneration of Castanopsis kawakamii in a subtropical natural forest, China. J. Trop. For. Sci. 2018, 30, 39–48. [Google Scholar] [CrossRef]
- He, Z.S.; Liu, J.F.; Zheng, S.Q.; Su, S.J.; Hong, W.; Wu, Z.Y.; Xu, D.W.; Wu, C.Z. Studies on the seeds dispersal and seedlings regeneration in gaps and understory of Castanopsis kawakamii natural forest. J. Trop. Subtrop. Bot. 2012, 20, 506–512. [Google Scholar]
- He, Z.S.; Wang, L.J.; Jiang, L.; Wang, Z.; Liu, J.F.; Xu, D.W.; Hong, W. Effect of microenvironment on species distribution patterns in the regeneration layer of forest gaps and non-gaps in a subtropical natural forest, China. Forests 2019, 10, 90. [Google Scholar] [CrossRef]
- Buajan, S.; Liu, J.F.; He, Z.S.; Feng, X.P.; Muhammad, A. The effect of light on micro-environment and specific leaf area within the gap, subtropical forest, China. Pak. J. Bot. 2017, 49, 273–282. [Google Scholar]
- Feldmann, E.; Drossler, L.; Hauck, M.; Kucbel, S.; Pichler, V.; Leuschner, C. Canopy gap dynamics and tree understory release in a virgin beech forest, Slovakian Carpathians. For. Ecol. Manag. 2018, 415, 38–46. [Google Scholar] [CrossRef]
- He, Z.S. Study on the Micro-Environment Characteristics and Seedlings Dynamic Regeneration in Castanopsis Kawakamii Natural Forest Gaps; Fujian Agriculture and Forestry University: Fuzhou, China, 2012. [Google Scholar]
- Xue, Y.L. Plant. Physiology Laboratory Manual; Shanghai Science and Technology Press: Shanghai, China, 1985. [Google Scholar]
- He, Z.S.; Liu, J.F.; Zheng, S.Q.; Hong, W.; Wu, C.Z.; Li, J. Diurnal variation of photosynthetic rates of Castanopsis kawakamii seedlings and their relationships with meteorological factors in forest gaps and non-gaps. Pak. J. Bot. 2018, 50, 1361–1368. [Google Scholar]
- Andrews, T.J.; Muller, G.J. Photosynthetic gas exchange of the mangrove, Rhizophora stylosa in its natural environment. Oecologia 1985, 65, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Liu, Y.Q.; Sun, X.Y.; Wang, Y.; Liu, Y. Effects of shades on the photosynthetic characteristics and chlorophyll fluorescence parameters of Urtica dioica. Acta Ecol. Sin. 2007, 27, 3457–3464. [Google Scholar]
- Liu, S.L.; Ma, M.D.; Pan, Y.Z.; Wei, L.L.; He, C.X.; Yang, K.M. Effects of light regime on growth and photosynthetic characteristics of Alnus formosana and A.cremastogyne seedlings. Chin. J. Appl. Ecol. 2013, 24, 351–358. [Google Scholar] [CrossRef]
- Sarijeva, G.; Knapp, M.; Lichtenthater, H.K. Differences in photosynthetic activity, chlorophyll and carotenoid levels, and in chlorophyll fluorescence parameters in green sun and shade leaves of Ginkgo and Fagus. J. Plant Physiol. 2007, 164, 950–955. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.S.; Niu, D.K.; Zhao, Z.H. A Study on photosynthesis and physiological characteristics of Camellia oleifera Abel Clones. Acta Agric. Univ. Jiangxiensis 2007, 29, 219–224. [Google Scholar] [CrossRef]
- Duan, R.Y.; Wei, X.L.; Meng, X.S. Physiological and biochemical responses and growth effects of Ormosia henryi seedlings under different light conditions. J. Cent. South Univ. For. Technol. 2013, 33, 30–34. [Google Scholar] [CrossRef]
- Qi, H.Y.; Hua, L.J.; Zhao, L.; Tang, Y.F. Effect of low night temperature on chlorophyll fluorescence parameters in different genotypes tomato leaves. Acta Agric. Boreali-Sin. 2011, 26, 222–227. [Google Scholar]
- Jiang, Y.; Hao, H.K.; Huang, Z.L.; Shen, W.H.; He, Q.F.; Peng, Y.H.; Huang, X.R. Effects of different light intensity on characteristics of growth and chlorophyll fluorescence of Castanopsis hystrix seedlings. J. Cent. South Univ. For. Technol. 2013, 33, 61–65. [Google Scholar] [CrossRef]
- Wu, F.Y.; Yi, L.T.; Li, X.P.; Yin, X.M.; Liu, M.H.; Yu, S.Q. Effect of different light intensity on intensity chlorophyll content and chlorophyll fluorescence in Lithocarpus glaber. J. Northeast Agric. Univ. 2012, 43, 88–92. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | Chl-a | Chl-b | Chl-T | Chl-a/b | |||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | p | F | p | F | p | F | p | |
| height | 1 | 0.326 | 0.573 | 13.788 | <0.001 *** | 2.832 | 0.104 | 10.042 | 0.004 ** |
| habitat | 1 | 10.663 | 0.003 ** | 15.128 | <0.001 *** | 16.348 | <0.001 *** | 0.07 | 0.793 |
| height × habitat | 1 | 0.01 | 0.919 | 0.082 | 0.776 | 0.000 | 0.996 | 0.401 | 0.532 |
| Habitat | Chl-a | Chl-b | Chl-T | Pn | Log(qP) | Y |
|---|---|---|---|---|---|---|
| GAP | 1.585 ± 0.345 b | 0.632 ± 0.153 b | 2.216 ± 0.455 b | 0.963 ± 0.665 a | −1.003 ± 0.325 b | 0.179 ± 0.038 b |
| NG | 1.976 ± 0.273 a | 0.822 ± 0.124 a | 2.797 ± 0.270 a | 0.691 ± 0.432 b | −0.816 ± 0.296 a | 0.205 ± 0.034 a |
| Between Groups | df | F | p | Within Groups (Time) | df | F | p |
|---|---|---|---|---|---|---|---|
| height | 1 | 5.990 | 0.022 * | time | 4 | 47.216 | <0.001 *** |
| habitat | 1 | 4.976 | 0.035 * | height × time | 4 | 0.572 | 0.684 |
| height × habitat | 1 | 6.627 | 0.016 * | habitat × time | 4 | 1.025 | 0.398 |
| height: habitat: time | 4 | 2.255 | 0.043 * |
| Factors | df | Fv/Fm | Log(qP) | Sin(NPQ) | Y | ||||
|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | ||
| treatment | 1 | 3.878 | 0.007 ** | 55.859 | <0.001 *** | 13.143 | <0.001 *** | 20.908 | <0.001 *** |
| height | 1 | 3.658 | 0.613 | 10.918 | 0.002 ** | 13.319 | <0.001*** | 2.450 | 0.124 |
| habitat | 1 | 0.074 | 0.786 | 10.372 | 0.002 ** | 0.127 | 0.723 | 11.414 | 0.001 ** |
| treatment × height | 1 | 0.030 | 0.864 | 3.257 | 0.077 | 0.012 | 0.914 | 12.554 | <0.001 *** |
| treatment × habitat | 1 | 0.001 | 0.970 | 0.222 | 0.639 | 7.455 | 0.009 ** | 1.679 | 0.201 |
| height × habitat | 1 | 3.395 | 0.071 | 14.725 | <0.001 *** | 18.269 | <0.001 *** | 3.062 | 0.086 |
| treatment × height × habitat | 1 | 1.662 | 0.203 | 0.425 | 0.517 | 1.084 | 0.303 | 0.028 | 0.868 |
| Treatment | Fv/Fm | Log(qP) | Sin(NPQ) | Y |
|---|---|---|---|---|
| WL | 0.681 ± 0.047 a | −1.107 ± 0.245 b | 0.486 ± 0.139 b | 0.175 ± 0.035 b |
| CK | 0.651 ± 0.037 b | −0.713 ± 0.268 a | 0.587 ± 0.135 a | 0.209 ± 0.035 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Z.-s.; Tang, R.; Li, M.-j.; Jin, M.-r.; Xin, C.; Liu, J.-f.; Hong, W. Response of Photosynthesis and Chlorophyll Fluorescence Parameters of Castanopsis kawakamii Seedlings to Forest Gaps. Forests 2020, 11, 21. https://doi.org/10.3390/f11010021
He Z-s, Tang R, Li M-j, Jin M-r, Xin C, Liu J-f, Hong W. Response of Photosynthesis and Chlorophyll Fluorescence Parameters of Castanopsis kawakamii Seedlings to Forest Gaps. Forests. 2020; 11(1):21. https://doi.org/10.3390/f11010021
Chicago/Turabian StyleHe, Zhong-sheng, Rong Tang, Meng-jia Li, Meng-ran Jin, Cong Xin, Jin-fu Liu, and Wei Hong. 2020. "Response of Photosynthesis and Chlorophyll Fluorescence Parameters of Castanopsis kawakamii Seedlings to Forest Gaps" Forests 11, no. 1: 21. https://doi.org/10.3390/f11010021
APA StyleHe, Z.-s., Tang, R., Li, M.-j., Jin, M.-r., Xin, C., Liu, J.-f., & Hong, W. (2020). Response of Photosynthesis and Chlorophyll Fluorescence Parameters of Castanopsis kawakamii Seedlings to Forest Gaps. Forests, 11(1), 21. https://doi.org/10.3390/f11010021