1. Introduction
In the Nordic countries, the planting season for conifer tree species has been extended to cover the entire period of unfrozen soil, from spring to late autumn. In Finland, for example, frozen or open-stored, dormant container seedlings of Norway spruce (
Picea abies (L.) Karst.) are recommended for planting between May and the middle of June. In turn, actively-growing Norway spruce seedlings are suggested to be planted between June and the end of July. From August onwards, it is recommended to use only short-day (SD)-treated seedlings in Norway spruce [
1,
2,
3]. For Scots pine (
Pinus sylvestris L.), it is suggested that container seedlings are only planted in spring or early summer due to their growth rhythm and growing conditions in their typical planting sites in Nordic countries [
3,
4,
5]. Similarly to the Nordic countries, summer plantings in July and August are also performed for different pine and spruce species in North America [
6,
7]. The extension of the planting season is a major prerequisite for enhancing the cost-competitiveness and facilitating the growth of the Nordic mechanized tree planting business [
8].
In Finland, more than half of the seedlings that are intended for spring plantings are overwintered in closed cardboard boxes in freezer storage. The cardboard storage method was originally invented for dormant seedlings that can be stored in closed boxes in the freezer for several months without suffering a significant decrease in vigor [
9]. Seedlings can be safely stored in closed boxes for up to two weeks after freezer storage in cool weather in spring, based on previous studies [
10]. However, in practical forestry in Finland, freezer-stored seedlings are commonly kept in closed cardboard boxes for longer period than that [
11].
Recently, forestry operators have also extended the use of storage of seedlings in closed cardboard boxes to the growing season to improve the efficiency of logistical operations. However, seedlings exist in different physiological stages during spring and summer (active period) when compared to late autumn or winter (dormancy) at the time of their packing for freezer storage. Thus, the forestry operators need better information on the duration limits of field storage in closed cardboard boxes for seedlings at various stages of physiological development, and risks that are related to this storage method.
Recently, there have been some observations of unexplained damage in conifer seedlings that were planted in the non-dormant stage, which may have partially resulted from them being stored for too long in closed boxes [
12]. Based on this, the physiology of non-dormant seedlings might make them unsuitable for long periods of storage in closed boxes in warm weather in summer and autumn [
9]. Overall, the physiology of seedlings changes during the annual cycle and seasonal conditions also affect it [
13]. During late summer and autumn, the shortening photoperiod and decreasing temperature affect the development of frost hardening (FH) [
14]. In late autumn, low temperatures are needed for proper FH [
14,
15,
16]. However, during field storage, the maximum and minimum temperatures can increase inside the closed boxes [
17]. The development of FH may be disturbed in seedlings intended for autumn (August to October) plantings as a result of the limited photoperiod and varying temperature conditions inside the boxes.
In spring, seedlings switch from the dormant stage to the active growing phase when air temperature increases. After flushing, the photosynthetic levels in the seedlings increase. During the flushing and active growing phases, the seedlings are sensitive to environmental stress factors, such as drought and lack of light [
13,
18]. An elevated temperature increases the respiration rate of the seedlings, and thus depletes their carbohydrate reserves [
13,
19,
20]. Therefore, elevated temperature, which restricts photosynthesis and water uptake inside closed boxes, is likely to cause stress and latent or direct damage to the seedlings during storage [
21,
22]. Actively growing seedlings may also be more sensitive to rapid changes in radiation and temperature as compared to seedlings that are dormant [
13].
The growth of new roots is important for rooting and survival of seedlings [
23]. Based on previous studies, root growth in newly planted Norway spruce and Scots pine seedlings slow down from August to October [
5,
24], starting up again in spring, when the soil temperature increases [
25]. However, we do not yet know how field storage in closed boxes affects root growth in late summer, early autumn, and spring, and how these potential changes in root growth affect the field performance of the planted seedlings.
Lately, there has been growing interest in also using small (mini) seedlings (15–40-cm
3 root-plug volume) for forest regeneration [
26,
27], especially in Scots pine. The root-plug volume in the most commonly produced conifer container seedlings is 40–125 cm
3 in Finland [
28]. Mini seedlings are especially used in the northern parts of Sweden and Finland, where the climatic conditions are more humid and the risk of drought is smaller than in the southern parts of these countries [
27,
29]. Smaller seedlings are economically and logistically cheaper to produce and transport. However, they are also more sensitive to environmental stress factors [
30,
31]. Small-volume root plugs, for example, store less water than larger plugs [
27]. Thus, mini seedlings can be very sensitive to prolonged storage and post-planting drought.
The silvicultural operators working in practical forestry should have a better understanding than currently regarding what may happen on field performance of seedlings if storing them for too long duration into closed packages (or kept in dark storehouses) in different planting seasons from spring to summer and autumn, respectively. In the above context, the aim of this study was to determine the safe duration periods for the field storage of non-dormant Norway spruce and Scots pine container seedlings in closed cardboard boxes, at different times (dates) in the planting seasons. We hypothesized that the Norway spruce container seedlings could be stored in closed boxes for some days before planting in summer, for about a week in September, and for 1–2 weeks in October and May, without a significant reduction in the maximum photochemical yield of photosystem II (PSII), root growth, FH, or field performance of seedlings. Further, the Scots pine mini seedlings were assumed to be more sensitive to prolonged storage in closed boxes in spring when compared to the conventional-sized Norway spruce seedlings.
2. Materials and Methods
2.1. Seedling Material
Experiments (Expts) 1 and 2 were conducted while using two-year-old Norway spruce container seedlings. The seeds, which were obtained from seed orchard no. 374 (supplying central Finland), were sown on 7 June 2016. The seedlings were grown in hard plastic Plantek (BCC, Landskrona, Sweden) 81F trays (81 cells per tray, 546 cells m
−2, cell volume 85 cm
3) in a greenhouse at the Suonenjoki Research Nursery of the Natural Resources Institute Finland (Luke; 62°39′ N, 27°03′ E, at an of altitude 142 m a.s.l.) until October 2016. The seedlings were grown according to the regular Finnish nursery practice for one-year-old seedlings, while using standard fertilization and irrigation procedures [
28,
32]. Afterwards, they were transferred to an outdoor area, where they were overwintered and grown during the following growing season. In midwinter, the seedlings were under snow cover. In the second growing season, they were given a commercial fertilizer solution (0.1% Kekkilä Forest Superex with 22N:5P:16K + micronutrients; Kekkilä Co., Tuusula, Finland) 10 times, from 6 June to 4 August 2017, resulting in a total of 94.5 g m
−2 fertilizer solution. In August and September, the seedlings were fertilized four times, which resulted in a total of 114.5 g m
−2 and 124.5 g m
−2 fertilizer solution for the seedlings packed in September and October, respectively. The seedlings were irrigated two to four times per week, depending on the weather conditions. A three-week short-day (SD) treatment (14-hour nights, 10-hour days) of growing seedlings was started on 10 July 2017, while using a blackout curtain in the outdoor growing compound (fertilization was continued during the SD treatment). No fungicides were used to protect the seedlings from storage molds.
One-year-old Scots pine seedlings were used in Expt 3. The seeds from seed orchard no. 323 (supplying central Finland) were sown between 13 and 14 June 2017, in hard plastic Airblock (BCC, Landskrona, Sweden) 196F trays (196 cells per tray, 1322 cells m−2, cell volume 30 cm3) at the Suonenjoki Research Nursery of Luke. The seedlings were fertilized five times, while using a commercial fertilizer solution (0.1% Kekkilä Forest Superex with 22N:5P:16K + micronutrients; Kekkilä Co., Tuusula, Finland), which resulted in total of 30 g m−2 between 27 July and 25 August 2017. Seedlings were overwintered outdoors.
2.2. Storage Treatments
For Expt 1, which was implemented in autumn 2018, a total of 6900 healthy Norway spruce container seedlings were used. Half of the seedlings were lifted and packed into 10 cardboard boxes (70 seedlings in each) in each month (on 7 August, 11 September, and 2 October;
Figure 1;
Appendix A Table A1). For Expts 2 and 3, a total of 375 healthy Norway spruce (Expt 2) and 375 Scots pine (Expt 3) seedlings were used, respectively. On 14 May, these seedlings were lifted and then packed into 10 cardboard boxes, five boxes for each tree species, and 75 seedlings in each box. As a control packing method (i.e., open storage), the other half of the seedlings (25 trays, 81 Norway spruce seedlings in each tray, in Expts 1 and 2, and five trays, 196 Scots pine seedlings in each, in Expt 3) were kept in the nursery in each packing month until planting. On 7 August and 14 May, the root plugs of Norway spruce and Scots pine seedlings, respectively, were loose and lower part of some of the plugs were broken when lifted. In the nursery, the seedlings were irrigated when needed, and fertilized, until they were packed, as described above. A day before packing, all of the seedlings were watered properly. At the time of box opening, the moisture of root plugs was checked, and the seedlings were watered before planting, if needed.
The data loggers for measuring temperature (HOBO Pendant UA-001-64; Onset Computer Corp., Bourne, MA, USA) were placed in each box at the time of packing in September, October, and May. In August, the data loggers were placed into two boxes, one week after the packing. At each packing time, one data logger was also placed between the trays in the nursery field. While using the Hobo logger data, the temperature sums accumulated during each storage periods were calculated as a sum of daily mean temperatures, T ≥ +5 °C). After packing, the boxes were randomized for each storage period (one, three, seven, 14, or 21 days) and then transferred to the shelter (open on one side only, sun exposure in the part of day) in ambient air conditions, where only the handle holes were open. The spacing between the boxes was 10 cm.
The storage periods used in the experiments were one, three, seven, 14, or 21 days (
Table A1). After each of these periods, one (Expts 2 and 3) or two (Expt 1) boxes were randomly selected and opened. At the same time, one (Expt 2 and 3) or two (Exp. 1) trays from the nursery were also randomly selected. Nine randomly selected seedlings from both boxes and trays were sampled for the measurement of chlorophyll fluorescence and seedling morphology at the time of the box opening. The rest of the seedlings were transferred to the planting site.
2.3. Planting of Seedlings
The seedlings were planted in a former nursery field in the Suonenjoki Research Nursery to determine their field performance. The soil in the field was fine sand, containing some organic matter. The field was cultivated before the first planting in August 2017 and May 2018. The seedlings were planted in rows, with 0.5 m between the seedlings within a row and between the rows. There were nine blocks in each experiment. There were three packing months in Expt 1 (August, September, October) and one packing month in Expts 2 and 3 (May) within the blocks. In each packing month, there were five storage periods, and within each storage period, there were two storage methods (open storage or closed box). In each block, five seedlings from both storage methods were planted on each planting date (total of 45 seedlings per storage method per storage period), while using a split-split-plot design (with packing month as the main plot (except for spring), and the storage periods and storage methods as subplots). The target planting depth was 6 cm (the length of stem below ground). However, in September, the seedlings that had been stored for 14 days were planted by mistake at a depth of only 3 cm (see depths,
Table A1). Moreover, the seedlings that were in open storage and the soil in the planting field were both frozen at the time of the last planting (21-day-storage period) in October, and only the seedlings from the closed boxes could be planted. However, on that date it was also not possible to fill the planting holes with soil.
A total of 432 seedlings were planted in separate field experiments in each packing month for the freezing tests in Expt 1. Eight seedlings, which represented storage periods of 0, one, three, seven, 14, and 21 days, were planted in nine blocks on the same dates as for plantings in the field performance experiments in August and September. In October, the seedlings were placed on eight trays and buried in the field. Each tray contained nine seedlings from each storage period, with a total of 54 seedlings in each container. For the root growth assessment, 18 seedlings that were in open storage were planted in the same field as part of Expt 1, two seedlings in each block from each storage period. In Expts 2 and 3, two additional seedlings were planted between the first four seedlings in each row in the field performance test for root growth measurement.
2.4. Measurements
Nine seedlings from each treatment combination were randomly selected for the measurement of chlorophyll fluorescence, height (within an accuracy of 0.1 cm), length of the current-year leader (only in Expts 2 and 3), and the diameter of the stem bases (1 cm above the root collar; accuracy 0.1 mm) at the end of each storage period (one, three, seven, 14, and 21 days). In the morning, between 9 and 12 a.m., chlorophyll fluorescence was measured with a Hansatech Pocket PEA fluorometer (Hansatech Instruments Ltd., King’s Lynn, Norfolk, UK). Seven needles were excised per seedling from two cm below the apical meristem. These were securely mounted on a piece of transparent tape, with the adaxial surface upwards. After dark-adapting the needles for 15 min., their minimum fluorescence (F
0) was determined [
33]. Their maximum fluorescence (F
m) was obtained by exposing the needles to a light pulse (3000 μmol m
−2 s
−1). The maximum photochemical yield of PSII in the dark-adapted state (F
v/F
m) was calculated as (F
m − F
0)/F
m [
34].
In the planting experiments, the vitality of the seedlings (healthy, minor damage, weakened, dead) and the cause of any damage (gray mold, drought, lack of light, other reason) were recorded after planting, first on 6 June 2018 and again in August 2018 and 2019. In 2019, multiple leaders and seedlings with yellowish-colored needles (based on visual observation) were also recorded. The total height (from the ground surface to the top of the terminal buds or growing points) and the current-year height growth were measured after planting (spring experiment) and in August 2018 and 2019 (all seedlings, with an accuracy of 1 mm).
2.5. Freezing Tests
For the freezing tests in the Expt 1, the Norway spruce seedlings packed in August and September were dug up on 9 and 16 October, respectively. From each block, one seedling from each storage period was randomized to each of eight exposure temperatures, with a total of nine seedlings in each test temperature and storage period. On 23 October, the buried eight trays were dug up, each containing the seedlings that were packed in October and represented eight test temperatures.
The seedlings packed in August and September were placed in plastic boxes, while the seedlings packed in October were kept in trays. Cloth bags that were filled with sawdust to prevent the roots from freezing covered both of them. The boxes/trays were placed into polystyrene boxes with 10-cm-thick sides. The seedlings were then exposed to eight temperatures, using four air-cooled chambers (WT600/70, Weiss Umweltechnik GmbH, Reiskirchen-Lidenstruth, Germany) over two consecutive days. The minimum exposure temperatures were 5, −5, −10, −15, −18, −22, −30 and −45 °C for August, 5, −5, −10, −18, −22, −26, −40, and −45 °C for September, and 5, −10, −26, −33, −35, −40, −45 and −50 °C for October. Each chamber was programmed to one target minimum temperature. The rate of cooling and warming was 5 °C h−1, and exposure to the minimum temperature in each chamber lasted for 3 h. The peat plugs were heated using a heating cable (Plug’n Heat, 54 W, Ensto, Finland), which was placed at the bottom of the polystyrene box, beneath the plastic boxes/trays to prevent the root plug temperature from dropping below 0 °C. The heating did not entirely prevent freezing of the peat plugs at the lowest temperatures. Thus, after exposure, the seedlings were kept at +5 °C until the plugs thawed. Subsequently, the seedlings were randomized to nine blocks, one seedling from each exposure temperature and storage period combination per each block, and then transferred to a heated greenhouse (on average, 20 °C by day/18 °C by night), where the natural light was supplemented while using 400-W high-pressure sodium lamps (18 h day−1). In the greenhouse, the seedlings were watered, when needed.
The extent of injury to each seedling was assessed after three weeks. Visual estimation determined injury to the needles. The needles were classed as damaged if there were any damaged needles on a seedling (excluding the part of the seedling that was below ground after planting). The apical bud was classed as being damaged if it did not burst during the four-week period after exposure to freezing. The FH was estimated as DT50; that is, temperatures that caused (needle or bud) damage to 50% of the seedlings.
2.6. Root Growth Assessment
We also used the seedlings stored in boxes and exposed to the two highest temperatures in the freezing tests on each packing date and storage period (18 seedlings per storage period) for root-growth measurements just after the freezing exposures due to the shortage of seedlings. Those seedlings, as well as the seedlings in open storage, were dug up on the same dates as the seedlings for the freezing tests. In Expts 2 and 3, the seedlings planted for the root-growth assessment in the field performance test were dug up after three weeks of growing in the field conditions. The roots growing out of the peat plugs were counted in all of the experiments. Whether the root plug was unbroken or not was also recorded.
2.7. Weather Data
Air temperature was monitored at the nearby weather station in the Suonenjoki Research Nursery of Natural Resources Institute Finland (Luke). Soil temperatures in the test field were measured while using two HOBO data loggers (type H08-008-05, Onset Computer Corp., Bourne, MA, USA), using a total of eight temperature sensors (type HOBO TMC20-HD), from 7 August to 11 November 2017 and from 27 April to 9 November 2018. Temperatures were measured at the depths of 7 and 13 cm (four sensors in each depth) in the test field in Suonenjoki. Volumetric soil water content was measured at the depths of 2 and 5 cm while using Decagon Devices logger (type Em5b) with four sensors (EcH2O EC5 Moisture Sensor; Decagon Devices, Inc., Pullman, WA, USA) from 15 May to 25 October 2018. The temperatures were close to, and precipitation was higher than, the 30-year average during autumn 2017 (
Table 1). The temperatures were above the long-term average during the 2018 growing season. In particular, the extreme temperatures were remarkably higher than the long-term average. The precipitation was clearly higher than the average in June and September 2018, but, in contrast, it was remarkably low in May and July 2018.
2.8. Data Analysis
Among the experimental treatment combinations, the differences in root growth, height, current-year height growth of seedlings in both measuring years, probability of mortality (combining the classes ‘weakened’ and ‘dead’ from the damage classification), and seedlings with multiple leaders and yellowish needles were analyzed, while using IPM SPSS Statistics Version 25.0. A linear mixed model (MIXED) was used to analyze the heights and number of roots. In Expt 1, we analyzed the differences in the average values among the months. After that, we separately analyzed the differences between the storage method and period for each packing month and tree species.
The probabilities for multiple leaders, mortality, or yellowish color were analyzed while using the general linear mixed model (GENLINMIXED). In the analysis, the storage method, storage period, and packing month were considered to be fixed effects, while the block, month within a block, period within a month, and block and method within a period, month and block were the random effects. A normal distribution was used in the MIXED model. We employed a binomial distribution with a logit-link function in the GENLINMIXED models. Multiple comparisons were based on the least significant difference. Chlorophyll fluorescence data, shoot heights, length of current-year leader (in Expts 2 and 3), and diameters of the stem bases of the seedlings after the storage treatments and before the plantings were analyzed while using ANOVA, separately, for each month in each experiment. Differences with a p value p ≤ 0.05 were considered to be significant.
Normal distribution was tested for with the Kolmogorov–Smirnov test, and homogeneity of variance with Levene’s test. In Expt 2, natural log-transformation was used to homogenize the variances in the number of roots growing out of a peat plug. Norway spruce chlorophyll fluorescence and growth data from Expts 2 and 3 were analyzed without any transformations, even though the data were not normally distributed and the variances were not homogenous (transformation did not help in normalizing the distribution or homogenize the variances).
The FH, being based on visually-assessed damage in the buds and needles, was analyzed for each packing month while using a generalized linear mixed model (GLMM) in PROC GLIMMIX in SAS for Windows 9.4 (SAS Institute Inc., Cary, NC, USA). The analysis was performed while using a GLMM with binary data. Maximum Likelihood with Adaptive Quadrature (QUAD) was used as an estimation method in the analysis. The storage period and exposure temperature (continuous), as well as their statistically significant interactions, were used as the fixed effects. The planting block (same blocks used after freezing exposure) was used to provide a random block effect. The model was:
where
yijk is the binary variable describing whether the bud or any of the needles on a seedling were dead/damaged,
i is the block,
j is the seedling, and
pijk is the probability that
yijk = 1. The probability is related to a linear predictor, as follows:
where the link is the logit link function in the GLMM,
xjk is the predictor vector, α is the vector for fixed parameters, and
bi ∼ N(0,
σb2) is a random block effect. The final model was:
where
a is the intercept,
Pk is the effect of storage period
k (1–6),
Tjk the exposure temperature for seedling
j in storage period
k, and
γk is the coefficient of the exposure temperature for the period
k. GLIMMIX gives the significance of different fixed factor levels (e.g., [
35]). We were interested in the temperatures at which the probability of bud or needle damage was 0.5 (DT
50). The statistical significance of the differences between two estimated DT
50 values were calculated while using the delta method and the Wald test statistics that Lappi and Luoranen described [
36].
3. Results
3.1. Environmental Conditions during Storage
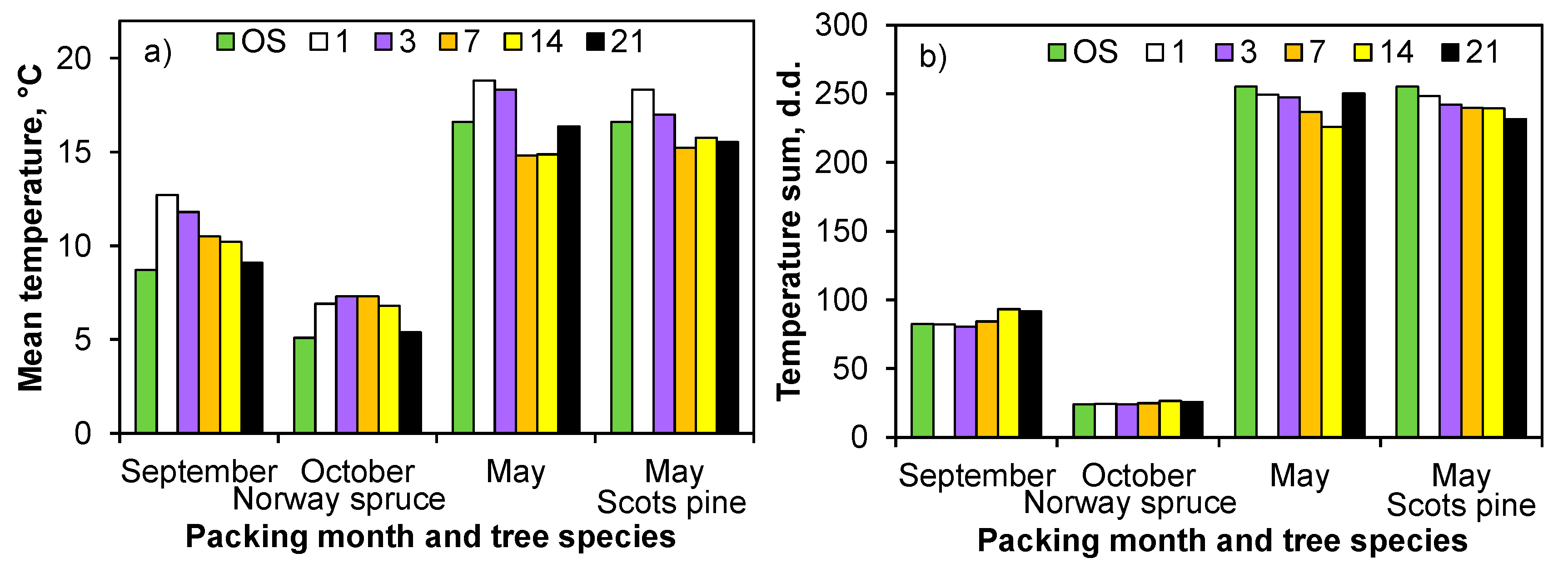
The temperatures inside the boxes did not fluctuate as much as they did in the open storage for the autumn plantings of Norway spruce (Expt 1;
Figure S1). The accumulated temperature sums (T ≥ +5 °C) inside the boxes were 8 d.d. (during the monitoring period), 9 d.d. (three-week storage), and 2 d.d. greater in August, Septembe, and October, respectively, than at the same time in the open storage (
Figure 2b).
In spring, the temperature variation was smaller inside the boxes than in the open storage (Expts 2 and 3). For Scots pine (Expt 3), the temperatures inside the boxes were, on average, 1 °C lower than in the open storage (
Figure 2a;
Figure S1d). The average temperatures and daily maximum temperatures inside the Norway spruce boxes were close to the corresponding averages in the open storage. Nonetheless, the daily minimum temperatures inside the boxes were, on average, 3.5 °C higher when compared to the open storage. For the Scots pine seedlings, the daily minimum temperatures were, on average, 2.4 °C higher, and the maximum temperatures were 2 °C lower inside the boxes as compared with the averages for the open storage. After the 21-day storage, the temperature sum was 19 d.d. lower for the Scots pine seedlings than for the Norway spruce seedlings (
Figure 2b).
3.2. Height Growth, Development of Apical Buds and Vitality of Seedlings During Storage
Height growth ceased in all of the seedlings at the time of packing in autumn (Expt 1). The average heights of the seedlings were 26 ± 0.2 cm, 27 ± 0.3 cm, and 28 ± 0.3 cm, and the average stem base diameters were 3.1 ± 0.04 mm, 3.3 ± 0.05 mm, and 3.8 ± 0.05 mm at the time of packing in August, September, and October, respectively. The seedlings were, on average, 0.3 mm thicker in the open storage than in the boxes (
Table S1). They were also thinner, the longer they were stored in the boxes, especially for the August packing. Seedlings that were stored for 21 days in the boxes were 0.1–0.6 mm thinner than the seedlings that were stored for shorter periods.
In August, the apical buds were developing, but they were not visible in the seedlings for the one-, three-, and seven-day storage periods. At the end of August, 36 and 56% of the seedlings for the 14- and 21-day storage periods in open storage had visible buds. For the same periods, only 11% of the seedlings that were stored in the boxes had visible buds. In September and October, all of the seedlings had visible buds at the time of packing. Based on visual evaluation, most of the seedlings were healthy when the boxes were opened. Some of the seedlings showed signs of gray mold, but its incidence was independent of the storage method or period. In August and September, the visually observed colors of the seedlings stored for seven and 14 days in the boxes were lighter green than those in the open storage.
In the spring, the first visible signs of etiolation were noted in the Norway spruce seedlings stored for seven days in closed boxes in the Expt 2. At that time, 16% of the current-year growth of the seedlings was etiolated, the proportion increasing rapidly as the storage period lengthened (76% of the seedlings at 14 days’ storage). In the seedlings that were stored for 21 days in closed boxes, the whole current-year growth was etiolated. By comparison, only a small percentage of seedlings that were stored for seven or 14 days in open storage were damaged (7% and 2%, respectively). The other seedlings remained in good condition (visually observed to be healthy looking) throughout the storage periods. Almost all of the Scots pine seedlings planted in spring, regardless of the storage method, were classed as being in good condition at the time of box opening in the Expt 3. However, minor signs of etiolation were observed from the 14-day storage period onward in the seedlings stored in boxes.
At the time of box opening in the spring, the average stem base diameter and total height of the Norway spruce seedlings were 4.0 ± 0.06 mm and 29.3 ± 0.5 cm, respectively. The seedlings started to grow during storage, and after the 14- and 21-day storage, lengths of the current year leader were 4.6 ± 0.3 and 8.5 ± 0.7 cm, respectively, without differences between storage methods.
The average stem base diameter and total height of the Scots pine seedlings were 2.2 ± 0.06 mm and 11.6 ± 0.3 cm, respectively, at the time of box opening. The lengths of the current-year leader were 3.8, 4.8, 6.1, 9.1, and 9.5 cm in the one-, three-, seven-, 14- and 21-day storage periods, respectively, without differences between the storage methods.
3.3. Variable/Maximum Fluorescence Ratio
In August, no changes in F
v/F
m were observed the storage periods in the Norway spruce seedlings that were stored in the open storage (Expt 1;
Table S1;
Figure 3a). In the seedlings that were stored in the closed boxes, the F
v/F
m was stable between the one- and three-day-storage periods but decreased afterwards. In September and October, differences in F
v/F
m between the storage periods were smaller than in August. After the three-day storage period in September, the F
v/F
m declined for both storage methods, but was more evident in the seedlings stored in boxes. In October, the F
v/F
m was the highest after one day of storage, declining after that, becoming the lowest after the 21-day storage. On that day, the open-storage seedlings were frozen and their F
v/F
m was lower than in the seedlings that were stored in the boxes.
In the spring, the F
v/F
m in the Norway spruce open-storage seedlings was stable up to 14 days, then decreasing in the Expt 2 (
Figure 3b). In the seedlings stored in closed boxes, the F
v/F
m was stable in the one-, three-, and seven-day storage periods, but then decreased more strongly than in the open-storage seedlings. In the Scots pine seedlings, the F
v/F
m similarly declined between the storage methods as the storage period lengthened in the Expt 3. However data for the one- and three-day storage periods could not be included in the analysis due to technical difficulties in conducting the measurements.
3.4. Root Plugs and Rooting Assessment
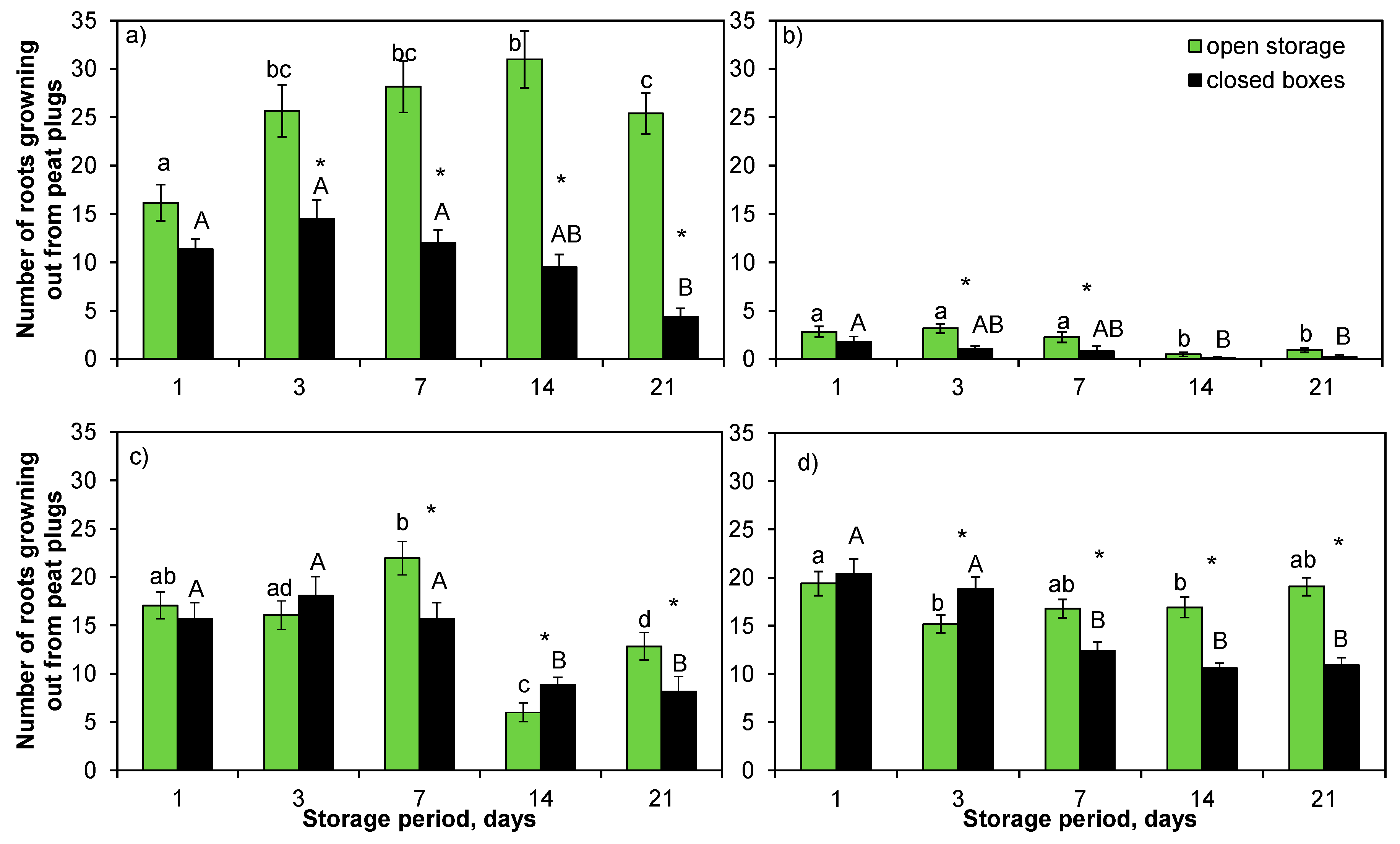
There was a clear effect of storage method and period on the results of Norway spruce seedlings in the Expt 1. When the seedlings were dug up from the soil in October, the lower parts of root plugs of the seedlings that had been packed in August broke. Broken plugs were only found in the one-day-stored (i.e., 33%) seedlings in the open-storage seedlings. However, in the seedlings that were stored in boxes, broken plugs were found in 28%, 28%, 17%, 0% and 17% of the seedlings for the one-, three-, seven-, 14-, and 21-day storage periods, respectively. All of the plugs were unbroken in the other months.
The seedlings used for root-growth assessment were planted in different experimental designs in each packing month and, thus, statistical analysis of root growth among the months was not possible. However, there were clear differences in the number of roots growing out of the plugs among the autumn packing months based on the data that are presented in
Figure 4 and visual observation of the seedlings. Root growth was strong in August, slowed down in September, and entirely ceased in October (no countable white roots). In August, the number of roots increased the longer the open storage period, whereas the root numbers decreased in the seedlings stored in boxes with lengthening storage period (
Table S1). In September, the number of roots decreased with lengthening of the storage period in both storage methods. Fewer roots were observed in the seedlings that were stored in boxes than in open storage in all storage periods in August and September (
Figure 4a,b). In August, root reduction was apparent after only one day of storage, although differences between the storage methods only became statistically significant from the three-day storage period onwards. In September, statistically significant differences in root growth between the storage methods were found for the three- and seven-day storage periods, and they were also almost significant for the one-day storage period.
In spring, prolonged storage reduced root growth in both the Norway spruce (Expt 2) and Scots pine seedlings (Expt 3), being more pronounced in the closed-box than open storage (
Figure 4c,d). In Norway spruce, the difference between storage methods appeared at the seven-day storage point, with decreasing numbers of roots growing out of the peat plugs (
Figure 4c). In Scots pine, the root growth of seedlings in open storage was quite stable across the different storage periods (
Figure 4d).
3.5. Frost Hardiness Development
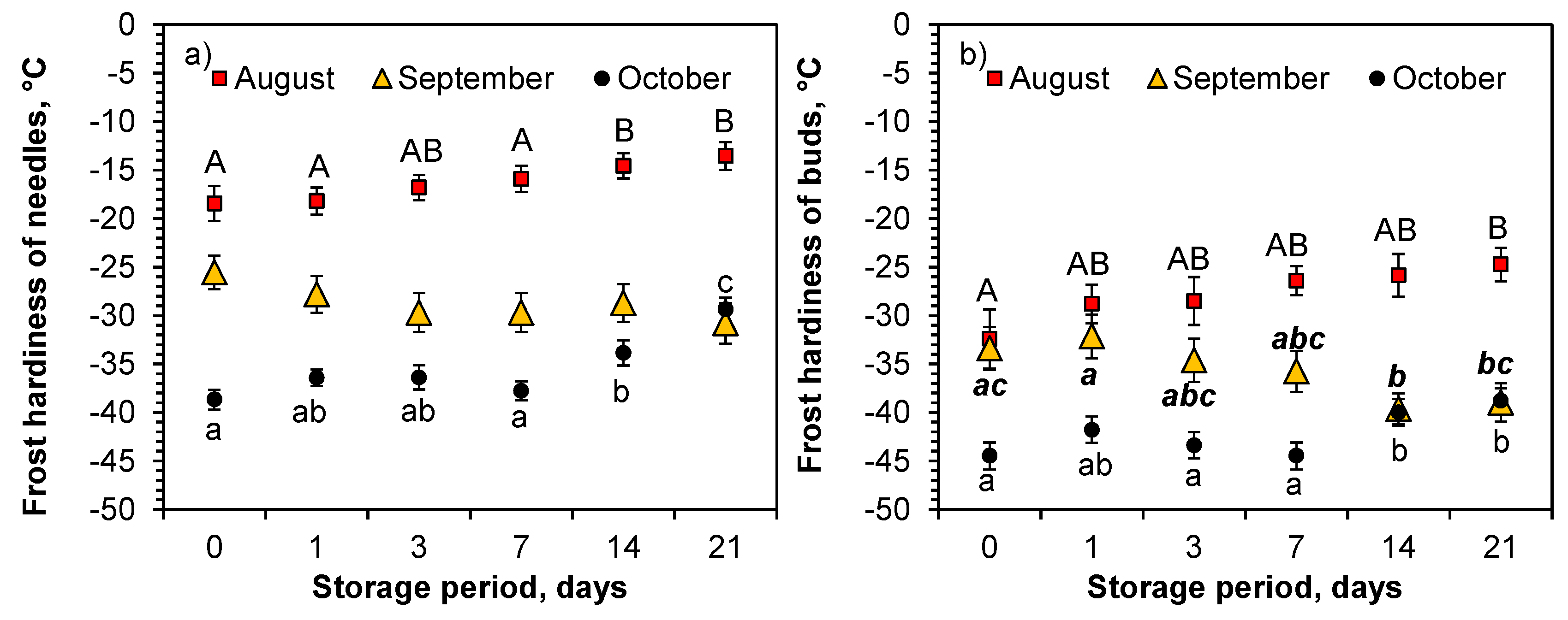
There was a clear trend in the FH (DT
50) of the needles in August and October in the Expt. 1 in the freezing tests. The longer the Norway spruce seedlings were in the closed boxes, the weaker their hardiness was (
Table S1,
Figure 5). The DT
50 estimates of needle damage were 5 °C and 9 °C lower for the 0-day storage than for the 21-day storage periods in August and October, respectively. For the buds, the trend was similar for October (
Figure 5b), but no differences were found in August (
Figure 5b). The trend was the opposite for September, with longer storage periods promoting lower DT
50 estimates, although the differences were only statistically significant for the buds.
3.6. Vitality of Seedlings after Planting
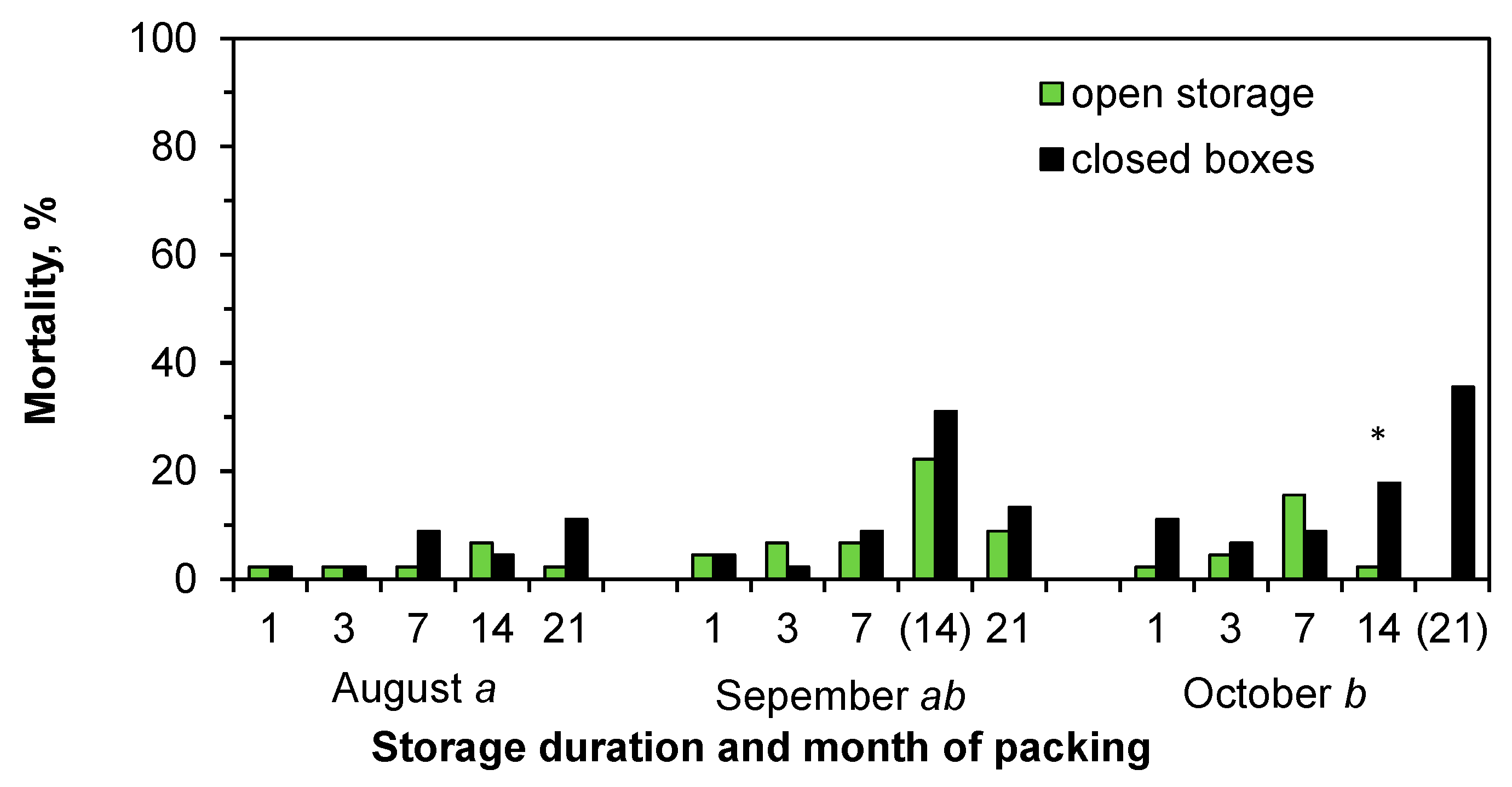
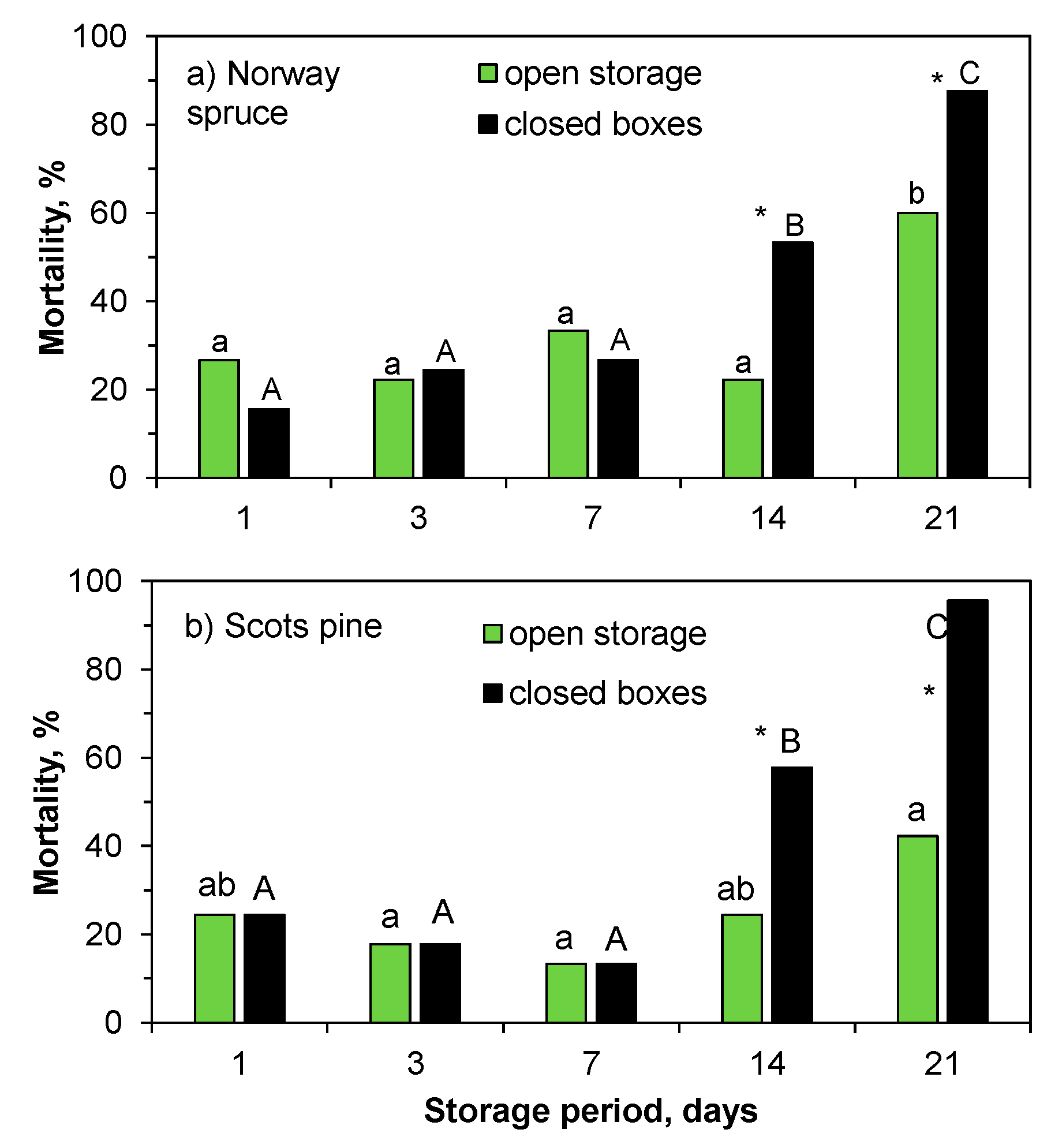
In the field test, mistakes and problems related to the planting (planting depth of 3 cm for the 14-day storage September seedlings, and frozen soil and unfilled planting holes for the 21-day storage October seedlings) clearly increased mortality in the Norway spruce seedlings (
Figure 6), and therefore these two dates were excluded from further analysis in the Expt. 1. On average, 4, 11, and 12% of the Norway spruce seedlings packed in August, September, and October, respectively, were dead or dying at the beginning of June 2018. At that time, the seedlings stored in closed boxes for 14 days had significantly higher mortality than the seedlings in open storage for 14 days (
Figure 6,
Table S1). After the first growing season, the average mortalities were 41%, 58%, and 76% in seedlings that were packed in August, September, and October, respectively. Mortality was caused by an exceptionally warm and dry growing season and it was not judicious to compare the storage methods or storage periods for this reason.
In the spring experiment (Expt. 2), storage from one to seven days increased the mortality of the Norway spruce seedlings by 7 to 24%, respectively, during the first season. After that, mortality drastically increased up to the 21-day storage period, at which point all of the seedlings were either dead or dying (
Figure S2). By comparison, the mortality of the seedlings in open storage was 7%–24% up to the 14-day storage period, increasing to 69% by 21 days of storage. Up to the end of the second season, mortality was similar among storage methods and periods from one to seven days of storage (
Figure 7a). For the longer storage periods, more of the seedlings stored in boxes died as compared to those taken from open storage, with mortality being greatest for the 21-day storage period in both methods. Increasing amounts of yellowish needles were in relation to length of storage period in the previous year (1, 4, 2, 14, and 33% for the one-, three-, seven-, 14-, and 21-day storage periods, respectively). Yellowish needles were found, especially in the seedlings that were stored in closed boxes (11%, on average). Only 5% of the seedlings in open storage had yellowish needles. There were also more seedlings with multiple leaders in the 21-day storage period than in the other periods, for both storage methods (3, 4, 2, 12 and 37% for one-, three-, seven-, 14-, and 21-day storage), with more in the seedlings stored in boxes (11%) than those in open storage.
At the end of the first growing season, 41% of the Scots pine seedlings that were stored in boxes in May and 25% of the seedlings in open storage were dead or dying in the Expt 3. The mortality was 27, 18, and 13% for the one-, three- and seven-day storage periods, respectively, for both methods. The mortality was 29 and 40% for the seedlings in open storage for 14 and 21 days, and 49 and 98% for the seedlings stored in boxes for the same periods (
Figure S3). During the second season, a few more seedlings died, and the trends in mortality were similar to those of the first year. From one to seven days of storage, no differences were found among the storage periods or methods, but mortality increased with longer storage, being greater in the seedlings that were stored in boxes (
Figure 7b). In the second season, more seedlings with yellowish needles were found in the boxes (10%) than in open storage (1%), and more were found in the longer-stored seedlings (3, 0, 3, 13, and 13% from one- to 21-day storage). On average, 4% (varying between 0%–11%) of the seedlings stored in boxes had multiple leaders. All of the seedlings in open storage before planting grew only one leader, and there were no differences among the storage methods or storage periods.
3.7. Height Growth after Planting
The average height growth of the Norway spruce seedlings was only 4 cm for all packing months in autumn in the Expt 1 during the first growing season in the field. The differences in height growth between storage methods or periods were not compared due to the high mortality (caused by exceptionally warm and dry growing season).
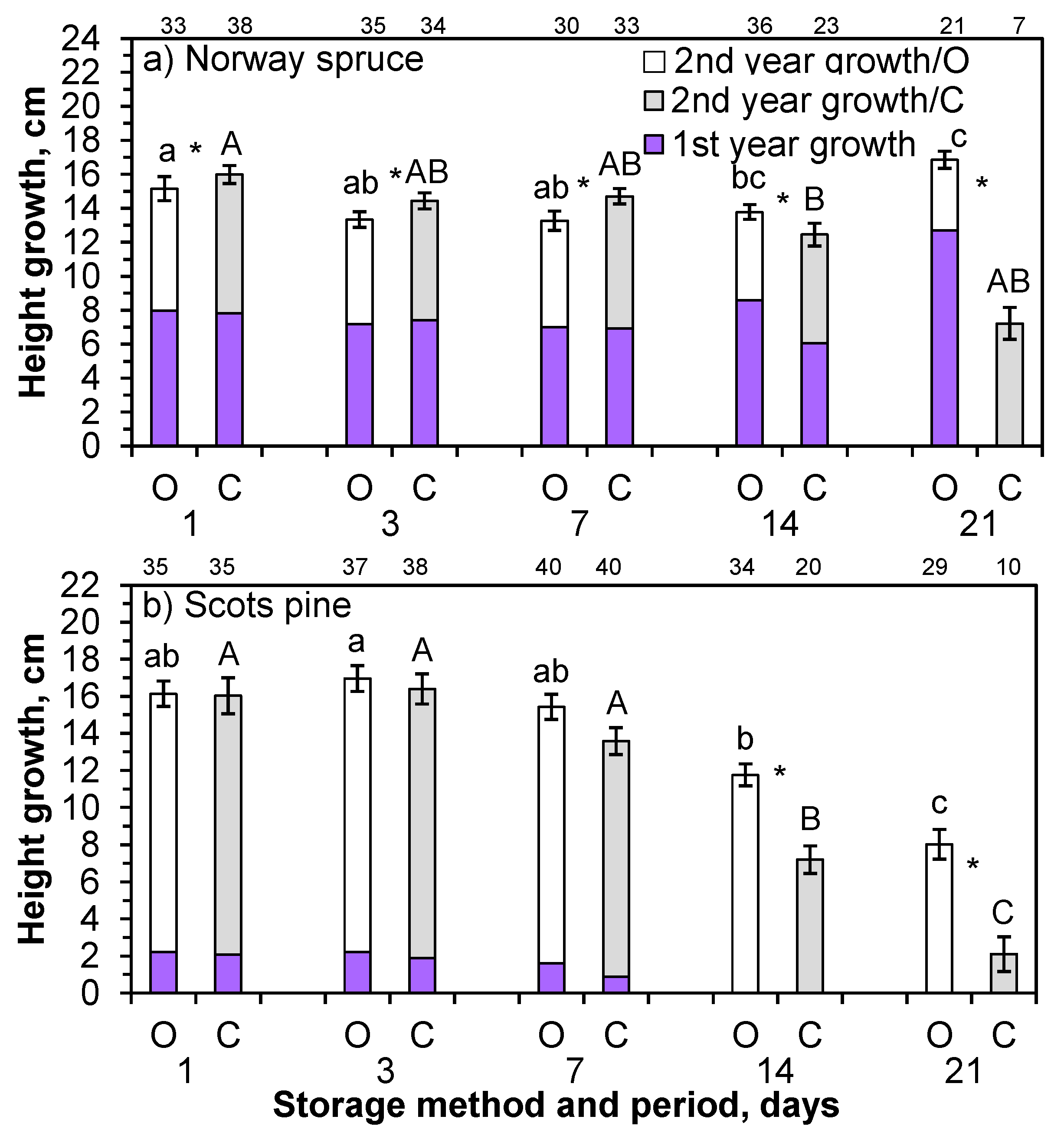
During the first growing season, the height growth in the Norway spruce seedlings in open storage in May slightly decreased between the one- and seven-day storage periods, but increased afterward this in the Expt 2. The height growth of the seedlings stored in boxes steadily decreased with duration of storage (
Figure 8a). All the seedlings stored in closed boxes for 21 days died. During the second growing season, the height of seedlings stored in boxes grew, on average, 1 cm more than the seedlings in open storage, although the height growth reduced after the 14- and 21-day storage periods in both methods (
Figure 8a).
During the first growing season, the average height growth of the Scots pine seedlings (only measured on living seedlings) was similar between storage methods in seedlings stored for one and three days, but decreased in the longer storage periods (Expt 3;
Figure 8b). In the second growing season, the height growth did not differ between the one- and seven-day storage periods for either storage method in Scots pine. However, the Scots pine seedlings that were stored for 14 or 21 days grew less, and the growth reduction was greater in the seedlings that were stored in boxes than in those in open storage.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







