Variability in Mixed Conifer Spatial Structure Changes Understory Light Environments

 ,
,  ,
,
Abstract
1. Introduction
2. Methods
2.1. Modeling Light Availability
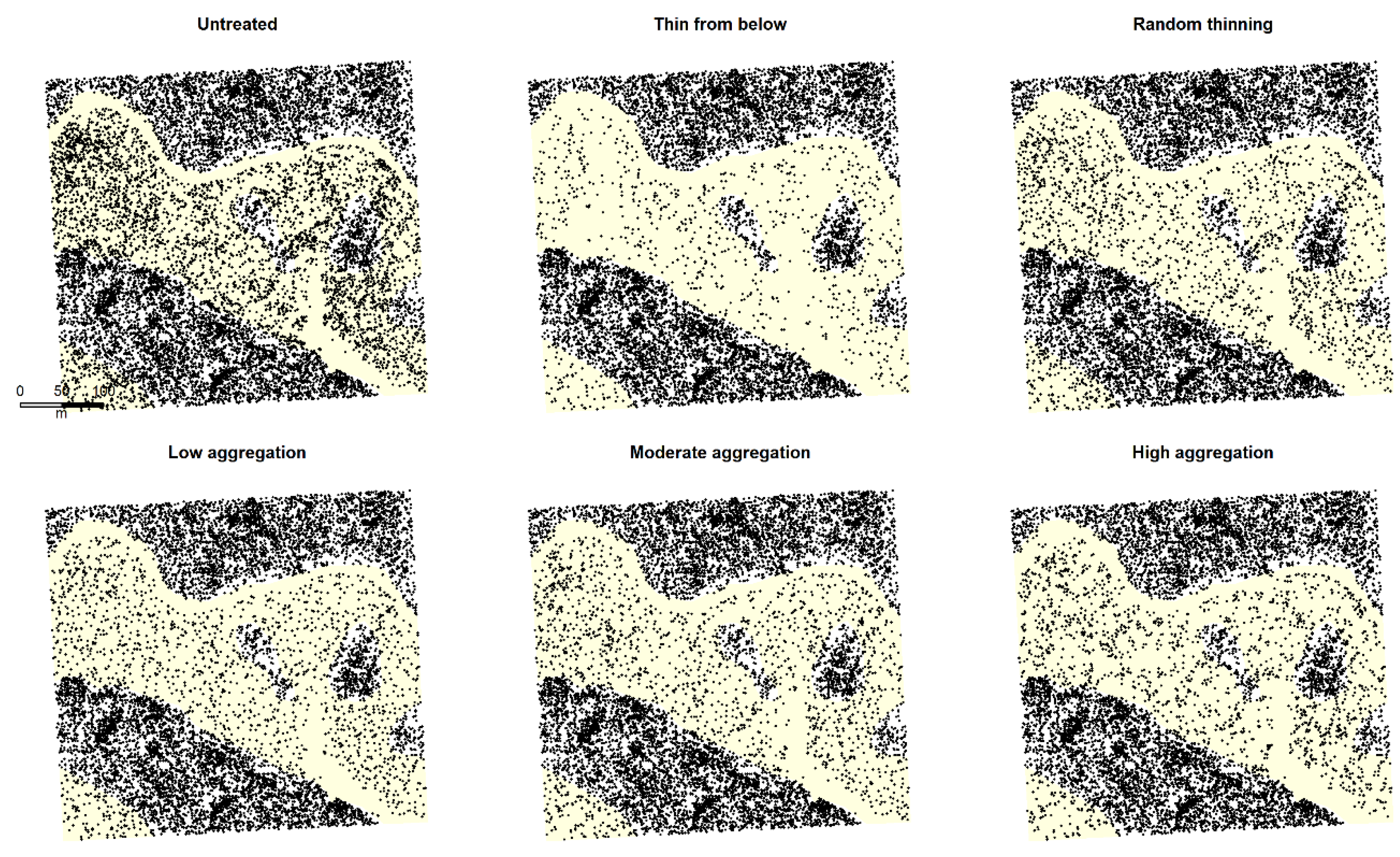
2.2. Simulating Restoration Treatments
2.3. Assessment of Structural Complexity and Light Availability
3. Results
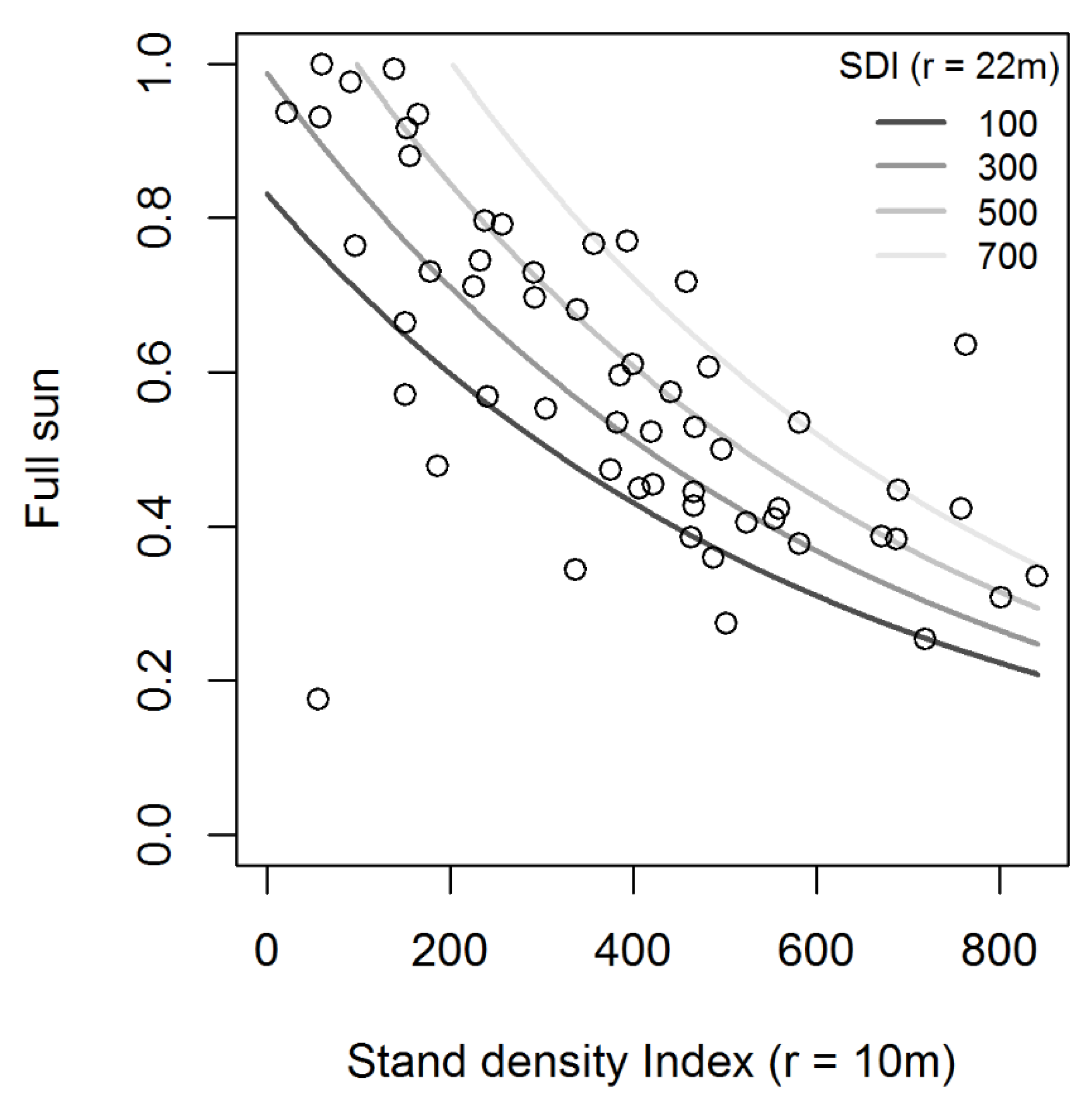
3.1. Modeling Overstory Structural Effects on Light Availability
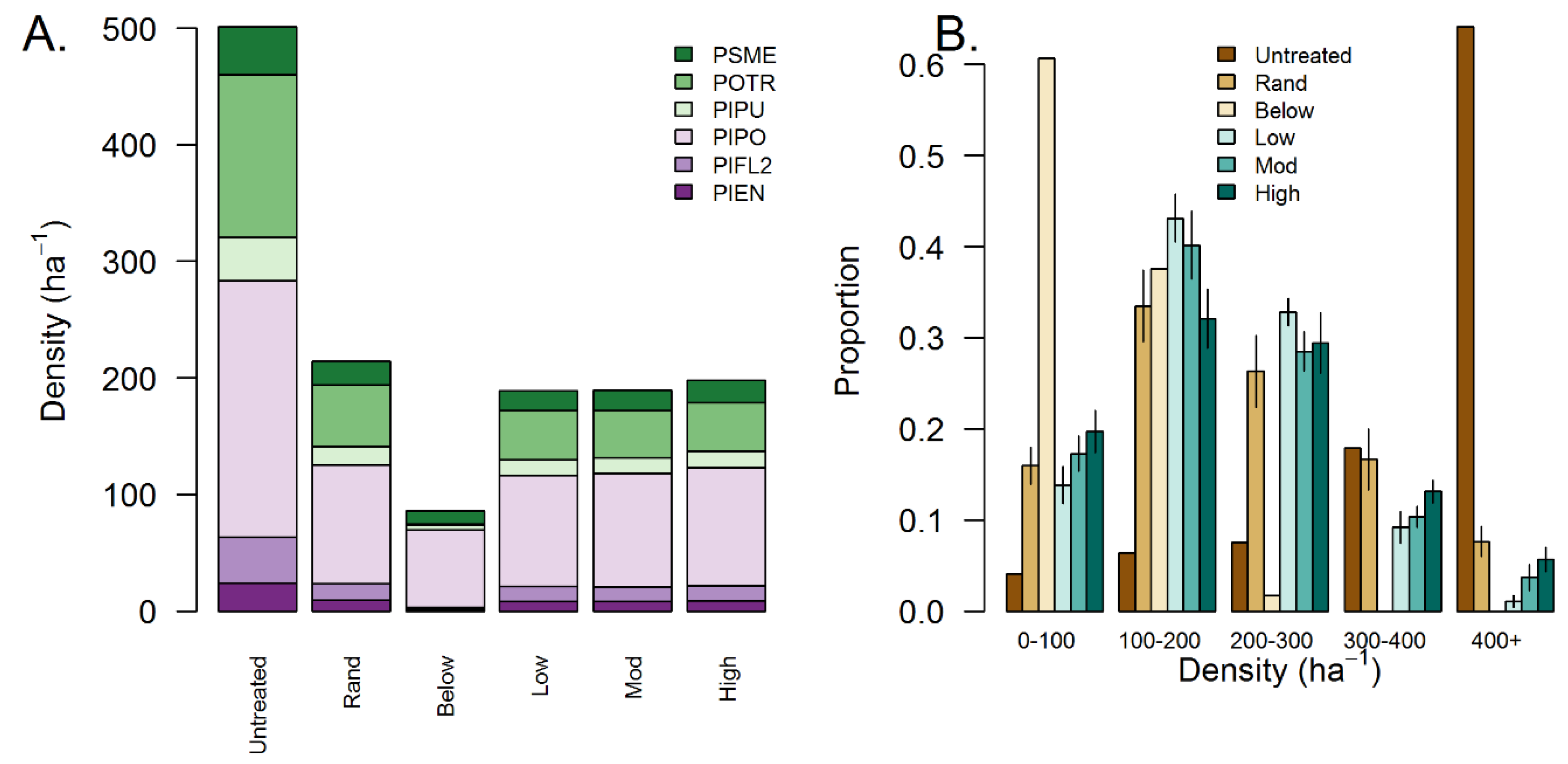
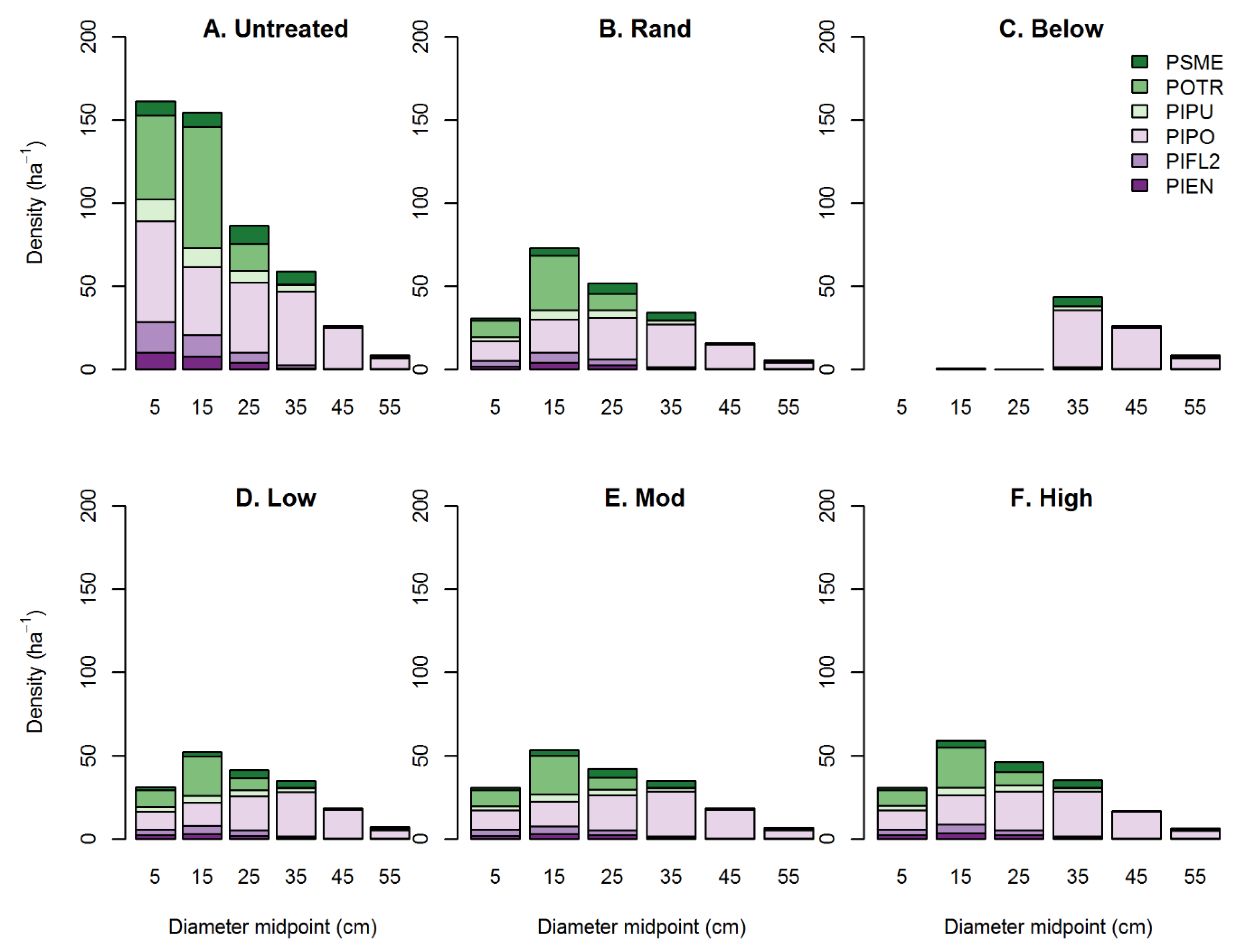
3.2. Effects of Treatment Simulations on Forest Structure and Complexity
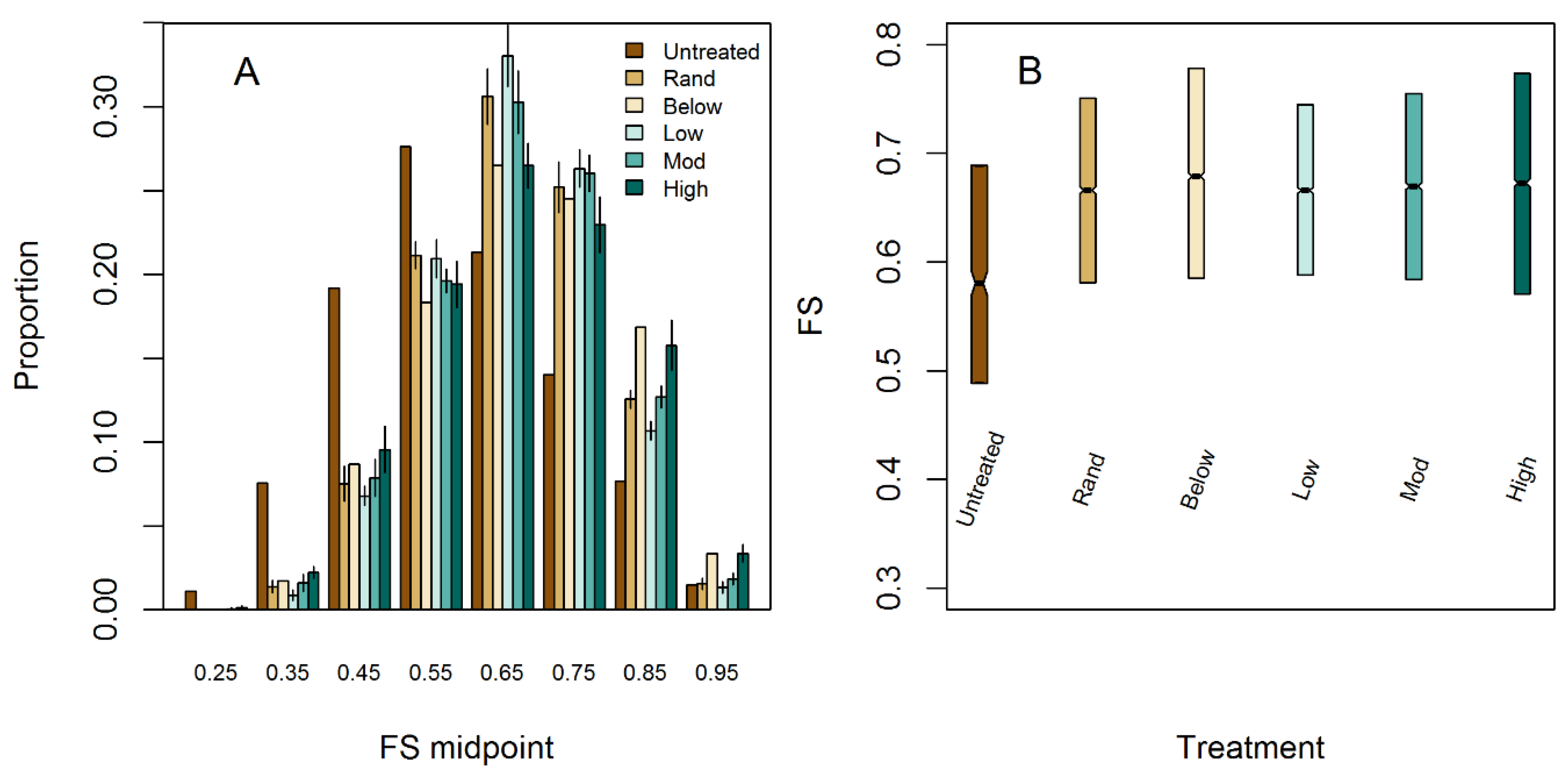
3.3. Effects of Treatment Simulations on Light Availability and Variability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Allen, C.D.; Savage, M.; Falk, D.A.; Suckling, K.F.; Thomas, W.; Schulke, T.; Stacey, P.B.; Morgan, P.; Hoffman, M.; Klingel, J.T. Ecological restoration of southwestern Ponderosa pine ecosystems: A broad perspective. Ecol. Appl. 2002, 12, 1418–1433. [Google Scholar] [CrossRef]
- Battaglia, M.A.; Gannon, B.; Brown, P.M.; Fornwalt, P.J.; Cheng, A.S.; Huckaby, L.S. Changes in forest structure since 1860 in ponderosa pine dominated forests in the Colorado and Wyoming Front Range, USA. For. Ecol. Manag. 2018, 422, 147–160. [Google Scholar] [CrossRef]
- Brown, P.M.; Battaglia, M.A.; Fornwalt, P.J.; Gannon, B.; Huckaby, L.S.; Julian, C.; Cheng, A.S. Historical (1860) forest structure in ponderosa pine forests of the northern Front Range, Colorado. Can. J. For. Res. 2015, 45, 1462–1473. [Google Scholar] [CrossRef]
- Fornwalt, P.J.; Huckaby, L.S.; Alton, S.K.; Kaufmann, M.R.; Brown, P.M.; Cheng, A.S. Did the 2002 hayman fire, Colorado, USA, burn with uncharacteristic severity? Fire Ecol. 2016, 12, 117–132. [Google Scholar] [CrossRef]
- Sherriff, R.L.; Platt, R.V.; Veblen, T.T.; Schoennagel, T.L.; Gartner, M.H. Historical, observed, and modeled wildfire severity in montane forests of the colorado front range. PLoS ONE 2014, 9, e106971. [Google Scholar] [CrossRef]
- Chambers, M.E.; Fornwalt, P.J.; Malone, S.L.; Battaglia, M.A. Patterns of conifer regeneration following high severity wildfire in ponderosa pine—dominated forests of the Colorado Front Range. For. Ecol. Manag. 2016, 378, 57–67. [Google Scholar] [CrossRef]
- Griffis, K.L.; Crawford, J.A.; Wagner, M.R.; Moir, W.H. Understory response to management treatments in northern Arizona ponderosa pine forests. For. Ecol. Manag. 2001, 146, 239–245. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologica 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Moody, J.A.; Martin, D.A. Hydrologic and Sedimentologic Response of Two Burned Watersheds in CO; US Geological Survey Water Resources Investigation Report 01-4122; US Geological Survey: Denver, CO, USA.
- Rhoades, C.C.; Entwistle, D.; Butler, D. The influence of wildfire extent and severity on streamwater chemistry, sediment and temperature following the Hayman Fire, Colorado. Int. J. Wildl. Fire 2011, 20, 430–442. [Google Scholar] [CrossRef]
- Agee, J.K.; Skinner, C.N. Basic principles of forest fuel reduction treatments. For. Ecol. Manag. 2005, 211, 83–96. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Crouse, J.E.; Roccaforte, J.P.; Kalies, E.L. Do thinning and/or burning treatments in western USA ponderosa or Jeffrey pine-dominated forests help restore natural fire behavior? For. Ecol. Manag. 2012, 269, 68–81. [Google Scholar] [CrossRef]
- Ziegler, J.P.; Hoffman, C.; Battaglia, M.; Mell, W. Spatially explicit measurements of forest structure and fire behavior following restoration treatments in dry forests. For. Ecol. Manag. 2017, 386, 1–12. [Google Scholar] [CrossRef]
- Larson, A.J.; Churchill, D. Tree spatial patterns in fire-frequent forests of western North America, including mechanisms of pattern formation and implications for designing fuel reduction and restoration treatments. For. Ecol. Manag. 2012, 267, 74–92. [Google Scholar] [CrossRef]
- Hessburg, P.F.; Agee, J.K.; Franklin, J.F. Dry forests and wildland fires of the inland Northwest USA: Contrasting the landscape ecology of the pre-settlement and modern eras. For. Ecol. Manag. 2005, 211, 117–139. [Google Scholar] [CrossRef]
- Reynolds, R.T.; Sánchez Meador, A.J.; Youtz, J.A.; Nicolet, T.; Matonis, M.S.; Jackson, P.L.; Delorenzo, D.G.; Graves, A.D.; Richard, T.; Meador, S.; et al. Restoring Composition and Structure in Southwestern Frequent-Fire Forests: A science-based framework for improving ecosystem resiliency. Gen. Tech. Rep. PSW-GTR-220 2013, 86, 310. [Google Scholar]
- Dodson, E.K.; Peterson, D.W. Dry coniferous forest restoration and understory plant diversity: The importance of community heterogeneity and the scale of observation. For. Ecol. Manag. 2010, 260, 1702–1707. [Google Scholar] [CrossRef]
- Matonis, M.S.; Binkley, D. Not just about the trees: Key role of mosaic-meadows in restoration of ponderosa pine ecosystems. For. Ecol. Manag. 2018, 411, 120–131. [Google Scholar] [CrossRef]
- Addington, R.N.; Aplet, G.H.; Battaglia, M.A.; Briggs, J.S.; Brown, P.M.; Cheng, A.S.; Dickinson, Y.; Feinstein, J.A.; Fornwalt, P.J.; Gannon, B.; et al. Principles and Practices for the Restoration of Ponderosa Pine and Dry Mixed Conifer Forests of the Colorado Front Range. Gen. Tech. Rep. RMRS-GTR-373; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2018.
- Cannon, J.B.; Barrett, K.J.; Gannon, B.M.; Addington, R.N.; Battaglia, M.A.; Fornwalt, P.J.; Aplet, G.H.; Cheng, A.S.; Underhill, J.L.; Briggs, J.S.; et al. Collaborative restoration treatments on forest structure in ponderosa pine forests of Colorado. For. Ecol. Manag. 2018, 424, 191–204. [Google Scholar] [CrossRef]
- Churchill, D.J.; Carnwath, G.C.; Larson, A.J.; Jeronimo, S.A. Historical Forest Structure , Composition, and Spatial Pattern in Dry Conifer Forests of the Western Blue Mountains , Oregon; General Technical Report (GTR) PNW-GTR-956; Pacific Northwest Research Station: Portland, OR, USA, 2017; p. 93. [Google Scholar]
- Dickinson, Y. Landscape restoration of a forest with a historically mixed-severity fire regime: What was the historical landscape pattern of forest and openings? For. Ecol. Manag. 2014, 331, 264–271. [Google Scholar] [CrossRef]
- Larson, A.J.; Stover, K.C.; Keyes, C.R. Effects of restoration thinning on spatial heterogeneity in mixed-conifer forest. Can. J. For. Res. 2012, 42, 1505–1517. [Google Scholar] [CrossRef]
- Tuten, M.C.; Sánchez, A.; Fulé, P.Z. Forest Ecology and Management Ecological restoration and fine-scale forest structure regulation in southwestern ponderosa pine forests. For. Ecol. Manag. 2015, 348, 57–67. [Google Scholar] [CrossRef]
- Underhill, J.L.; Dickinson, Y.; Rudney, A.; Thinnes, J. Silviculture of the Colorado Front Range Landscape Restoration Initiative. J. For. 2014, 112, 484–493. [Google Scholar] [CrossRef]
- Churchill, D.J.; Larson, A.J.; Dahlgreen, M.C.; Franklin, J.F.; Hessburg, P.F.; Lutz, J.A. Restoring forest resilience: From reference spatial patterns to silvicultural prescriptions and monitoring. For. Ecol. Manag. 2013, 291, 442–457. [Google Scholar] [CrossRef]
- Dickinson, Y.; Pelz, K.; Giles, E.; Howie, J. Have we been successful? Monitoring horizontal forest complexity for forest restoration projects. Restor. Ecol. 2016, 24, 8–17. [Google Scholar] [CrossRef]
- Dickinson, Y.L.; Cadry, J.D. An evaluation of tree marking methods for implementing spatially heterogeneous restoration. J. Sustain. For. 2017, 36, 47–64. [Google Scholar] [CrossRef]
- Tinkham, W.T.; Dickinson, Y.; Hoffman, C.M.; Battaglia, M.A.; Ex, S.; Underhill, J.; Service, F. Visualization of Heterogeneous Forest Structures Following Treatment in the Southern Rocky Mountains Forest Structure Forest Structure Comparison of Treatment Effects; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2017.
- Briggs, J.S.; Fornwalt, P.J.; Feinstein, J.A. Short-term ecological consequences of collaborative restoration treatments in ponderosa pine forests of Colorado. For. Ecol. Manag. 2017, 395, 69–80. [Google Scholar] [CrossRef]
- Battaglia, M.A.; Mou, P.; Palik, B.; Mitchell, R.J. The effect of spatially variable overstory on the understory light environment of an open-canopied longleaf pine forest. Can. J. For. Res. 2002, 32, 1984–1991. [Google Scholar] [CrossRef]
- Boyden, S.; Montgomery, R.; Reich, P.B.; Palik, B. Seeing the forest for the heterogeneous trees: Stand-scale resource distributions emerge from tree-scale structure. Ecol. Appl. 2012, 22, 1578–1588. [Google Scholar] [CrossRef]
- Boyden, S.; Binkley, D.; Shepperd, W. Spatial and temporal patterns in structure, regeneration, and mortality of an old-growth ponderosa pine forest in the Colorado Front Range. For. Ecol. Manag. 2005, 219, 43–55. [Google Scholar] [CrossRef]
- Boyden, S.; Binkley, D. The effects of soil fertility and scale on competition in ponderosa pine. Eur. J. For. Res. 2015, 135, 1–8. [Google Scholar] [CrossRef]
- Palik, B.; Mitchell, R.J.; Pecot, S.; Battaglia, M.; Pu, M. Spatial distribution of overstory retention influences resources and growth of longleaf pine seedlings. Ecol. Appl. 2003, 13, 674–686. [Google Scholar] [CrossRef]
- Bigelow, S.W.; North, M.P.; Salk, C.F. Using light to predict fuels-reduction and group- selection effects on succession in Sierran mixed- conifer forest. Can. J. For. Res. 2011, 2063, 2051–2063. [Google Scholar] [CrossRef]
- Chen, H.Y.H. Interspecific responses of planted seedlings to light availability in interior British Columbia: Survival, growth, allometric patterns, and specific leaf area. Can. J. For. Res. 1997, 27, 1383–1393. [Google Scholar] [CrossRef]
- Vyse, A.; Ferguson, C.; Simard, S.W.; Kano, T.; Puttonen, P. Growth of Douglas-fir, lodgepole pine, and ponderosa pine seedlings underplanted in a partially-cut, dry Douglas-fir stand in south-central British Columbia. For. Chron. 2006, 82, 723–732. [Google Scholar] [CrossRef]
- Ma, S.; Concilio, A.; Oakley, B.; North, M.; Chen, J. Spatial variability in microclimate in a mixed-conifer forest before and after thinning and burning treatments. For. Ecol. Manag. 2010, 259, 904–915. [Google Scholar] [CrossRef]
- Parsons, R.A.; Linn, R.R.; Pimont, F.; Hoffman, C.; Sauer, J.; Winterkamp, J.; Sieg, C.H.; Jolly, W.M. Numerical Investigation of Aggregated Fuel Spatial Pattern Impacts on Fire Behavior. Land 2017, 6, 43. [Google Scholar] [CrossRef]
- Tinkham, W.T.; Hoffman, C.M.; Ex, S.A.; Battaglia, M.A.; Saralecos, J.D. Ponderosa pine forest restoration treatment longevity: Implications of regeneration on fire hazard. Forests 2016, 7, 137. [Google Scholar] [CrossRef]
- Anderson-Teixeira, K.J.; Davies, S.J.; Bennett, A.C.; Gonzalez-Akre, E.B.; Muller-Landau, H.C.; Joseph Wright, S.; Abu Salim, K.; Almeyda Zambrano, A.M.; Alonso, A.; Baltzer, J.L.; et al. CTFS-ForestGEO: A worldwide network monitoring forests in an era of global change. Glob. Chang. Biol. 2015, 21, 528–549. [Google Scholar] [CrossRef]
- Dickinson, Y.L.; SHSFRR. Desirable Forest Structures for a Restored Front Range, CFRI-TB-1402; Front Range Roundtable and Colorado Forest Restoration Institute: Fort Collins, CO, USA, 2014. [Google Scholar]
- Matonis, M.S.; Binkley, D.; Tuten, M.; Cheng, T. The Forests They Are A-Changin’: Ponderosa pine and Mixed Conifer Forests on the Uncompahgre Plateau in 1875 and 2010-13; Colorado Forest Restoration Institute: Forest Collins, CO, USA, 2010. [Google Scholar]
- Frazer, G.W.; Canham, C.D.; Lertzman, K.P. Gap Light Analyzer (GLA): Imaging software to extract canopy structure and gap light tranmision indices from true-colour fisheye photographs, users manual and program documentation. In User Manual and Program Documentation; Simon Fraser University, Burnaby, British Coloumbia, Canada and the Institute of Ecosysteme Studies: Millbrook, NY, USA, 1999. [Google Scholar]
- Thekaekara, M.P.; Drummon, A.J. Standard values for the solar constant and its spectral components. Nat. Phys. Sci. 1971, 229, 6–9. [Google Scholar] [CrossRef]
- Knapp, C.L.; Stoffel, T.L.; Whitaker, S.D. Insolation Data Manual: Long-Term Monthly Averages of Solar Radiation, Temperature, Degree-Days and Global KT for 248 National Weather Service Stations; Solar Energy Information Data Bank, Solar Energy Research Institute: Golden, CO, USA, 1980. [Google Scholar]
- Grant, R.H.; Heisler, G.M. Obscured overcast sky radiance distributions for ultraviolet and photosynthetically active. J. Appl. Meteorol. 1997, 36, 1336. [Google Scholar] [CrossRef]
- Canham, C.D.; Denslow, J.S.; Platt, W.J.; Runkle, J.R.; Spies, T.A.; White, P.S. Light regimes beneath closed canopies and tree-fall gaps in temperate and tropical forests. Can. J. For. Res. 1990, 20, 620–631. [Google Scholar] [CrossRef]
- Shaw, J.D. Application of stand density index to irregularly structured stands. West. J. Appl. For. 2000, 15, 40–42. [Google Scholar] [CrossRef]
- Canham, C.D.; Lepage, P.T.; Coates, K.D. A neighborhood analysis of canopy tree competition: Effects of shading versus crowding. Can. J. For. Res. 2004, 787, 778–787. [Google Scholar] [CrossRef]
- Monsi, M.; Saeki, T. Über den Lichtfaktor in den Pflanzengesellschaften und seine Bedeutung für die Stoffproduktion. Jpn. J. Bot. 1952, 1952 95, 549–567. [Google Scholar]
- Larcher, W. Physiological Plant Ecology. Ecophysiology and Stress Physiology of the Functional Groups; Springer Science & Business Media: Berlin, Germany, 2003. [Google Scholar]
- Cribari-Neto, F.; Zeileis, A. Beta regression in R. J. Stat. Softw. 2010, 34, 1–24. [Google Scholar] [CrossRef]
- Sakamota, Y.; Masato, I.; Katagawa, G. Akaike Information Criterion Statistics; Springer: Dordrecht, The Netherlands, 1986. [Google Scholar]
- Abella, S.R.; Denton, C.W. Spatial variation in reference conditions: Historical tree density and pattern on a Pinus ponderosa landscape. Can. J. For. Res. 2009, 39, 2391–2403. [Google Scholar] [CrossRef]
- Sánchez Meador, A.J.; Parysow, P.F.; Moore, M.M. A New Method for Delineating Tree Patches and Assessing Spatial Reference Conditions of Ponderosa Pine Forests in Northern Arizona. Restor. Ecol. 2011, 19, 490–499. [Google Scholar] [CrossRef]
- North, M.P.; Kane, J.T.; Kane, V.R.; Asner, G.P.; Berigan, W.; Churchill, D.J.; Conway, S.; Gutiérrez, R.J.; Jeronimo, S.; Keane, J.; et al. Cover of tall trees best predicts California spotted owl habitat. For. Ecol. Manag. 2017, 405, 166–178. [Google Scholar] [CrossRef]
- North, M.; Sherlock, J. Chapter 9. Marking and assessing forest heterogeneity. In Managing Sierra Nevada Forests. PSW-GTR-220; Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2012. [Google Scholar]
- Valbuena, R.; Packalén, P.; Martín-Fernández, S.; Maltamo, M. Diversity and equitability ordering profiles applied to study forest structure. For. Ecol. Manag. 2012, 276, 185–195. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- Reineke, L.H. Perfecting a stand-density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Ducey, M.J.; Larson, B.C. Is there a correct stand density index? An alternate interpretation. West. J. Appl. For. 2003, 18, 179–184. [Google Scholar] [CrossRef]
- Shaw, J.D.; Long, J.N. A density management diagram for even-aged ponderosa pine stands. West. J. Appl. For. 2005, 205–215. [Google Scholar]
- Perot, T.; Mårell, A.; Korboulewsky, N.; Seigner, V.; Balandier, P. Modeling and predicting solar radiation transmittance in mixed forests at a within-stand scale from tree species basal area. For. Ecol. Manag. 2017, 390, 127–136. [Google Scholar] [CrossRef]
- Martens, S.N.; Breshears, D.D.; Meyer, C.W. Spatial distributions of understory light along the grassland/forest continuum: Effects of cover, height, and spatial pattern of tree canopies. Ecol. Modell. 2000, 126, 79–93. [Google Scholar] [CrossRef]
- McDonald, P.M.; Abbott, C.S. Seedfall, Regeneration, and Seedling Development in Group-Selection Openings; Pacific Southwest Research Station, Forest Service, US Department of Agriculture: Albany, CA, USA, 1994.
- Shepperd, W.D.; Edminster, C.B.; Mata, S.A. Long-term seedfall, establishment, survival, and growth of natural and planted ponderosa pine in the Colorado Front Range. West. J. Appl. For. 2006, 21, 19–26. [Google Scholar] [CrossRef]
- McDonald, P.M. Forest Regeneration and Seedling Growth from Five Major Cutting Methods in North-Central California; Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1976.
- Jameson, D.A. The Relationship of Tree Overstory and Herbaceous Understory Vegetation. J. Range Manag. 1967, 20, 247. [Google Scholar] [CrossRef]
- North, M.; Oakley, B.; Chen, J.; Erickson, H.; Gray, A.; Izzo, A.; Johnson, D.; Ma, S.; Marra, J.; Meyer, M.; et al. Vegetation and Ecological Characteristics of Mixed Conifer and Red Fir Forests at The Teakettle Experimental Forest; Gen. Tech. Rep. PSW-GTR-186; Pacific Southwest Research Station: Albany, CA, USA, 2002; p. 52.
- Korb, J.E.; Daniels, M.L.; Laughlin, D.C.; Fulé, P.Z. Understory communities of warm-dry, mxied-conifer forests in southwestern Colorado. Southwest. Nat. 2007, 52, 493–503. [Google Scholar] [CrossRef]
- Brown, P.M.; Wu, R. Climate and disturbance forcing of episodic tree recruitment in a southwestern ponderosa pine landscape. Ecology 2005, 86, 3030–3038. [Google Scholar] [CrossRef]
- Petrie, M.D.; Bradford, J.B.; Hubbard, R.M.; Lauenroth, W.K.; Andrews, C.M.; Schlaepfer, D.R. Climate change may restrict dryland forest regeneration in the 21st century. Ecology 2017, 98, 1548–1559. [Google Scholar] [CrossRef]
- Puhlick, J.J.; Laughlin, D.C.; Moore, M.M. Factors influencing ponderosa pine regeneration in the southwestern USA. For. Ecol. Manag. 2012, 264, 10–19. [Google Scholar] [CrossRef]
- Rother, M.T.; Veblen, T.T.; Furman, L.G. A field experiment informs expected patterns of conifer regeneration after disturbance under changing climate conditions. Can. J. For. Res. 2015, 45, 1607–1616. [Google Scholar] [CrossRef]
- Rother, M.T.; Veblen, T.T. Limited conifer regeneration following wildfires in dry ponderosa pine forests of the Colorado Front Range. Ecosphere 2016, 7. [Google Scholar] [CrossRef]
- Savage, M.; Mast, J.N.; Feddema, J.J. Double whammy: High-severity fire and drought in ponderosa pine forests of the Southwest. Can. J. For. Res. 2013, 43, 570–583. [Google Scholar] [CrossRef]
- Bartels, S.F.; Chen, H.Y.H. Is Understory Plant Species Diversity Driven by Resource Quantity or Resource Heterogeneity? Ecology 2010, 91, 1931–1938. [Google Scholar] [CrossRef] [PubMed]
- Getzin, S.; Wiegand, T.; Wiegand, K.; He, F. Heterogeneity influences spatial patterns and demographics. J. Ecol. 2008, 96, 807–820. [Google Scholar] [CrossRef]
- Dobrowski, S.Z.; Swanson, A.K.; Abatzoglou, J.T.; Holden, Z.A.; Safford, H.D.; Schwartz, M.K.; Gavin, D.G. Forest structure and species traits mediate projected recruitment declines in western US tree species. Glob. Ecol. Biogeogr. 2015, 24, 917–927. [Google Scholar] [CrossRef]
- Ffolliott, P.F.; Gottfried, G.J. Mixed conifer and aspen regeneration in small clearcuts within a partially harvested Arizona mixed conifer forest. Aspen Bibliography 1991. Paper 2764. [Google Scholar]
- Platt, W.J.; Rathbun, S.L. Dynamics of an old-growth longleaf pine population. In Proceedings of the Tall TImbers Fire Ecology Conference, Tallahassee FL, USA, 30 May–2 June 1993; p. 275. [Google Scholar]
- Gagnon, J.L.; Jokela, E.J.; Moser, W.K.; Huber, D.A. Dynamics of artificial regeneration in gaps within a longleaf pine flatwoods ecosystem. For. Ecol. Manag. 2003, 172, 133–144. [Google Scholar] [CrossRef]
- McGuire, J.P.; Mitchell, R.J.; Barry Moser, E.; Pecot, S.D.; Gjerstad, D.H.; Hedman, C.W. Gaps in a gappy forest: Plant resources, longleaf pine regeneration, and understory response to tree removal in longleaf pine savannas. Can. J. For. Res. 2001, 31, 765–778. [Google Scholar] [CrossRef]
- Peck, J.E.; Zenner, E.K.; Palik, B. Variation in microclimate and early growth of planted pines under dispersed and aggregated overstory retention in mature managed red pine in Minnesota. Can. J. For. Res. 2012, 42, 279–290. [Google Scholar] [CrossRef]
- Powers, M.D.; Pregitzer, K.S.; Palik, B.J. Physiological performance of three pine species provides evidence for gap partitioning. For. Ecol. Manag. 2008, 256, 2127–2135. [Google Scholar] [CrossRef]
- Barbeito, I.; Fortin, M.J.; Montes, F.; Cañellas, I. Response of pine natural regeneration to small-scale spatial variation in a managed Mediterranean mountain forest. Appl. Veg. Sci. 2009, 12, 488–503. [Google Scholar] [CrossRef]
- Cannon, J.B.; Gannon, B.M.; Feinstein, J.A.; Wolk, B.H. An effects assessment framework for dry forest conservation. Rangelands 2019, in press. [Google Scholar] [CrossRef]
- Ex, S.A.; Ziegler, J.P.; Tinkham, W.T.; Hoffman, C.M. Long-term impacts of fuel treatment placement with respect to forest cover type on potential fire behavior across a mountainous landscape. Forests 2019, 10, 438. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Simulation | Basal Area Target (m2 ha−1) | Small Trees (< 12.7 cm) | Overstory Retention Pattern |
|---|---|---|---|
| Thin from below | 11.5 | Thinned randomly to 40 ha−1 | Smallest trees removed preferentially to stand target |
| Random | 11.5 | Thinned randomly to 40 ha−1 | Trees removed randomly to stand target |
| Low aggregation | 11.5 | Thinned randomly to 40 ha−1 | Single trees (50%); 2–4 trees (40%); 5–9 tree (10%) |
| Mod. aggregation | 11.5 | Thinned randomly to 40 ha−1 | Single trees (35%); 2–4 trees (30%); 5–9 tree (20%); 10–15 trees (10%); 16+ trees (5%) |
| High aggregation | 11.5 | Thinned randomly to 40 ha−1 | Single trees (10%); 2–4 trees (30%); 5–9 tree (35%); 10–15 trees (15%); 16+ trees (10%) |
| Metric | s | s′ | Pseudo-r2 | AIC | ΔAIC |
|---|---|---|---|---|---|
| SDI | 10 | 22 | 0.4217 | −69.42 | 0.00 |
| BA | 10 | 22 | 0.4152 | −69.05 | 0.37 |
| SDI | 10 | 24 | 0.4199 | −67.95 | 1.47 |
| SDI | 10 | 26 | 0.4158 | −66.97 | 2.45 |
| BA | 10 | 24 | 0.4127 | −66.95 | 2.47 |
| SDI | 10 | 28 | 0.4068 | −65.80 | 3.62 |
| SDI | 10 | 20 | 0.4123 | −65.68 | 3.74 |
| SDI | 10 | 30 | 0.3977 | −65.52 | 3.90 |
| NI | 10 | 22 | 0.3512 | −65.49 | 3.93 |
| BA | 10 | 26 | 0.4069 | −65.31 | 4.11 |
| Coefficient | Estimate | Standard Error | p |
|---|---|---|---|
| Intercept | −0.27069 | 0.08399 | 0.001 |
| SDI (r = 10 m) | −0.00164 | 0.00012 | < 0.001 |
| SDI (r = 22 m) | 0.00086 | 0.00021 | < 0.001 |
| Phi | 9.480 | 1.724 | < 0.001 |
| Metric | Units | Untreated | Random | Thin Below | Low Aggregation | Mod. Aggregation | High Aggregation |
|---|---|---|---|---|---|---|---|
| Basal area | m2 ha−1 | 20 | 11.5 (0) | 11.5 (0) | 11.6 (0) | 11.6 (0) | 11.6 (0) |
| Stem density | ha−1 | 501 | 214 (3) | 86 (0) | 189 (1) | 190 (2) | 198 (3) |
| Dq | cm | 22.5 | 26.2 (0.2) | 41.4 (0) | 27.9 (0.1) | 27.9 (0.2) | 27.3 (0.2) |
| Single trees | proportion BA | 0.027 | 0.13 (0.01) | 0.36 (0) | 0.27 (0.01) | 0.24 (0.01) | 0.08 (0.01) |
| 2–4 trees | proportion BA | 0.059 | 0.24 (0.03) | 0.48 (0) | 0.34 (0.02) | 0.27 (0.02) | 0.23 (0.01) |
| 5–9 trees | proportion BA | 0.051 | 0.18 (0.02) | 0.13 (0) | 0.23 (0.03) | 0.22 (0.01) | 0.29 (0.03) |
| 10–15 trees | proportion BA | 0.031 | 0.11 (0.02) | 0.04 (0) | 0.1 (0.02) | 0.12 (0.02) | 0.15 (0.03) |
| 16+ trees | proportion BA | 0.832 | 0.34 (0.02) | 0 (0) | 0.06 (0.01) | 0.15 (0.02) | 0.24 (0.02) |
| Horizontal complexity | proportion BA | 0.68 | 0.93 (0.01) | 0.46 (0) | 0.78 (0.02) | 0.86 (0.03) | 0.91 (0.02) |
| Tree size complexity | proportion BA | 0.11 | 0.07 (0) | 0.01 (0) | 0.08 (0) | 0.07 (0) | 0.07 (0) |
| Treatment | Light Availability (FS) | FS Coefficient of Variation | 75th Percentile FS |
|---|---|---|---|
| Untreated | 0.592 | 0.244 | 0.689 |
| Random | 0.665 (0.002) | 0.181 (0.004) | 0.751 (0.004) |
| Thin below | 0.676 (0.000) | 0.196 (0.000) | 0.778 (0.000) |
| Low agg. | 0.664 (0.001) | 0.17 (0.003) | 0.745 (0.003) |
| Mod. agg | 0.666 (0.001) | 0.184 (0.005) | 0.755 (0.006) |
| High agg. | 0.669 (0.002) | 0.205 (0.003) | 0.773 (0.004) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cannon, J.B.; Tinkham, W.T.; DeAngelis, R.K.; Hill, E.M.; Battaglia, M.A. Variability in Mixed Conifer Spatial Structure Changes Understory Light Environments. Forests 2019, 10, 1015. https://doi.org/10.3390/f10111015
Cannon JB, Tinkham WT, DeAngelis RK, Hill EM, Battaglia MA. Variability in Mixed Conifer Spatial Structure Changes Understory Light Environments. Forests. 2019; 10(11):1015. https://doi.org/10.3390/f10111015
Chicago/Turabian StyleCannon, Jeffery B., Wade T. Tinkham, Ryan K. DeAngelis, Edward M. Hill, and Mike A. Battaglia. 2019. "Variability in Mixed Conifer Spatial Structure Changes Understory Light Environments" Forests 10, no. 11: 1015. https://doi.org/10.3390/f10111015
APA StyleCannon, J. B., Tinkham, W. T., DeAngelis, R. K., Hill, E. M., & Battaglia, M. A. (2019). Variability in Mixed Conifer Spatial Structure Changes Understory Light Environments. Forests, 10(11), 1015. https://doi.org/10.3390/f10111015