
Pre-Carpels from the Middle Triassic of Spain

1
Departamento de Xeociencias Mariñas e Ordenaciόn do Territorio, Universidade de Vigo, 36200 Vigo, Spain
2
State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology and CAS Center for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, Nanjing 210008, China
*
Author to whom correspondence should be addressed.
Plants 2022, 11(21), 2833; https://doi.org/10.3390/plants11212833
Submission received: 30 September 2022
/
Revised: 20 October 2022
/
Accepted: 22 October 2022
/
Published: 25 October 2022
(This article belongs to the Special Issue Plant Morphology and Phylogenetic Evolution)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:In stark contrast to the multitude of hypotheses on carpel evolution, there is little fossil evidence testing these hypotheses. The recent discovery of angiosperms from the Early Jurassic makes the search for precursors of angiosperm carpels in the Triassic more promising. Our light microscopic and SEM observations on Combina gen. nov., a cone-like organ from the Middle Triassic of Spain, indicate that its lateral unit includes an axillary anatropous ovule and a subtending bract, and the latter almost fully encloses the former. Such an observation not only favors one of the theoretical predictions but also makes some Mesozoic gymnosperms (especially conifers and Combina) comparable to some angiosperms. Combina gen. nov. appears to be an important chimeric fossil plant that may complete the evidence chain of the origin of carpels in geological history, partially narrowing the gap between angiosperms and gymnosperms.
1. Introduction
The origin of angiosperms and their relationship with other seed plants have been the foci of botanical debates for a long time [1,2]. Carpels (the basic units of gynoecia in angiosperms) are idiosyncratic to angiosperms [3]. According to the traditional theory, a carpel results from the longitudinal folding and enrolling of a megasporophyll that bears ovules along its margins [4,5,6,7]. This hypothesis sounded rational, especially when Goethe’s dictum “Alles ist Blatt” was taken into consideration [4]. However, it has been discarded since the APG system came into existence [8], and the APG system cannot give a plausible morphological interpretation for carpel homology [9]. Thus, plant systematics has entered a dead end: no widely accepted interpretation for the origin and homology of carpels is given, leaving many botanical questions unanswered. Therefore, using a fossil reproductive organ morphologically intermediate between angiosperms and gymnosperms to sift one hypothesis out of many is of crucial importance to move plant systematics beyond this debate. Although challenged by three groups of authors [10,11,12], Nanjinganthus, based on over 200 specimens of flowers, remains robust as an angiosperm from the Early Jurassic, since these challengers could not reach a consensus on the definition of angiosperms among themselves [13]. The Early Jurassic age of Nanjinganthus [13,14,15] suggests that the Triassic is a promising period for the search for a carpel precursor. Here, we report a new cone-like reproductive organ, Combina gen. nov. (Figure 1 and Figure 2), from the Anisian (the lower Middle Triassic, >242 Ma) of Spain. In contrast to the seed–scale–bract–complex (SSBC) frequently seen in conifer cones, each lateral unit in Combina gen. nov. comprises an anatropous ovule in the axil of a bract that folds longitudinally and almost fully encloses the ovule. Such a configuration demonstrates a certain resemblance to both SSBCs in some Mesozoic “conifers” and carpels in some extant basal angiosperms (e.g., Illicium, Michelia) [16,17], suggesting a possible common ancestor shared by some angiosperms and Mesozoic “conifers”. Its unique morphology and Triassic age make Combina gen. nov. one of the plausible ancestor candidates for angiosperm carpels, which otherwise were thought to emerge out of nowhere.
2. Results
2.1. Systematic Palaeontology
2.1.1. Combina gen. nov.
Diagnosis: Reproductive organ, cone-like and cylindrical, with helically arranged lateral units. Each lateral unit including an axillary ovule and a subtending bract. Ovule anatropous, attached to the organ axis. Bract longitudinally folded, with a ventral longitudinal suture.
Etymology:Combina from the Latin word “combinare”, meaning “unite two things together”, since the fossil combines the characteristics of angiosperms and conifers.
Type species:Combina triassica gen. et sp. nov.
2.1.2. Combina triassica gen. et sp. nov.
(Figure 1)
Diagnosis: In addition to the genus diagnosis: organ with at least 13 lateral units helically arranged. Lateral units decreasing distally in size. Bracts elliptical to ovate in shape, up to 10 mm long. Ovules attached to the organ axis, up to 8 mm long, with a smooth outline and an asymmetrical base.
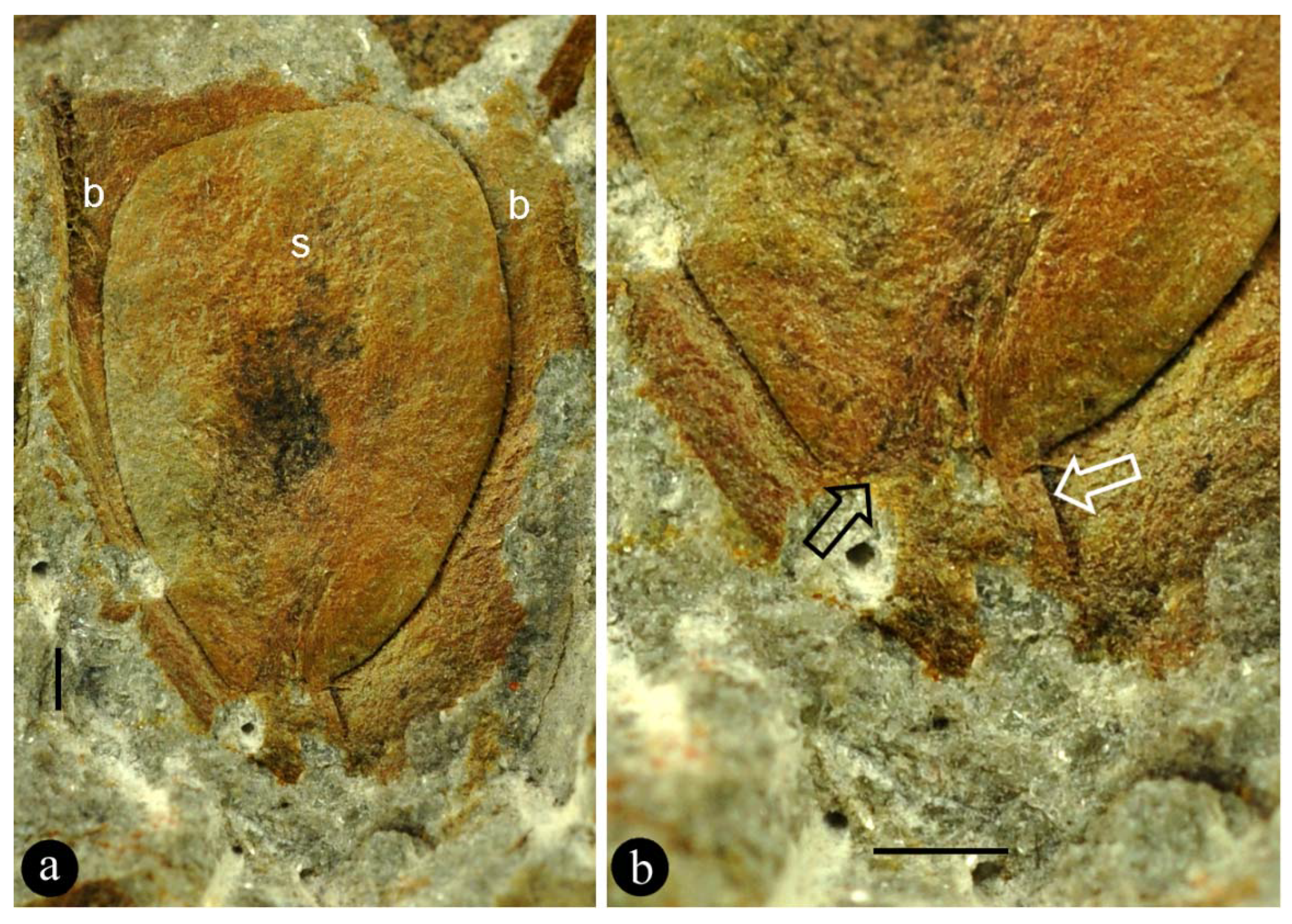
Description: The organ is cylindrical in form, 38 mm long and 16 mm wide, including more than 13 lateral units (Figure 1a). The lateral units are helically arranged along the organ axis, decreasing distally in size (Figure 1a). The lateral units diverge from the organ axis at angles between 27° and 66° (Figure 1a). Each lateral unit comprises a subtending bract and an axillary ovule (Figure 1b,c,g–j). The bract is 7 to 10 mm long, elliptical to ovate in shape, almost fully enclosing the ovule from bottom and laterals, leaving a ventral gaping suture (Figure 1a–j). Each ovule is attached to the organ axis by a short funiculus (Figure 1a–f,i,j). The ovule is anatropous, 7–8 mm long, and 1–2.3 mm thick, with a funiculus about 1 mm long and 0.3–1 mm in diameter (Figure 1b,c,g–j). The ovule is smooth-outlined and basally asymmetrical (Figure 1b,c,g–j; Figure 3).
Associated with the holotype of Combina is a part (Figure 2a,b) that resembles the lateral units in situ and thus provides an otherwise unavailable perspective on the lateral unit of Combina. The ovule and its funiculus in the holotype of Combina are not fully smooth and asymmetrical (Figure 1i,j): the funiculi appear to skew to one side of the ovule, suggestive of an anatropous ovule. This observation and inference of the holotype are further confirmed by an adaxial view of the isolated lateral unit (Figure 2a,b).
Holotype: MPZ2009-425.
Further specimen: R4/40/38/5.
Etymology:triassica for the Triassic, the age of the fossil.
Locality: Rodanas outcrop, Aragonian Branch of Iberian Range, Spain.
Horizon and Age: The Calcena Formation, Anisian, lower Middle Triassic [18].
3. Discussion
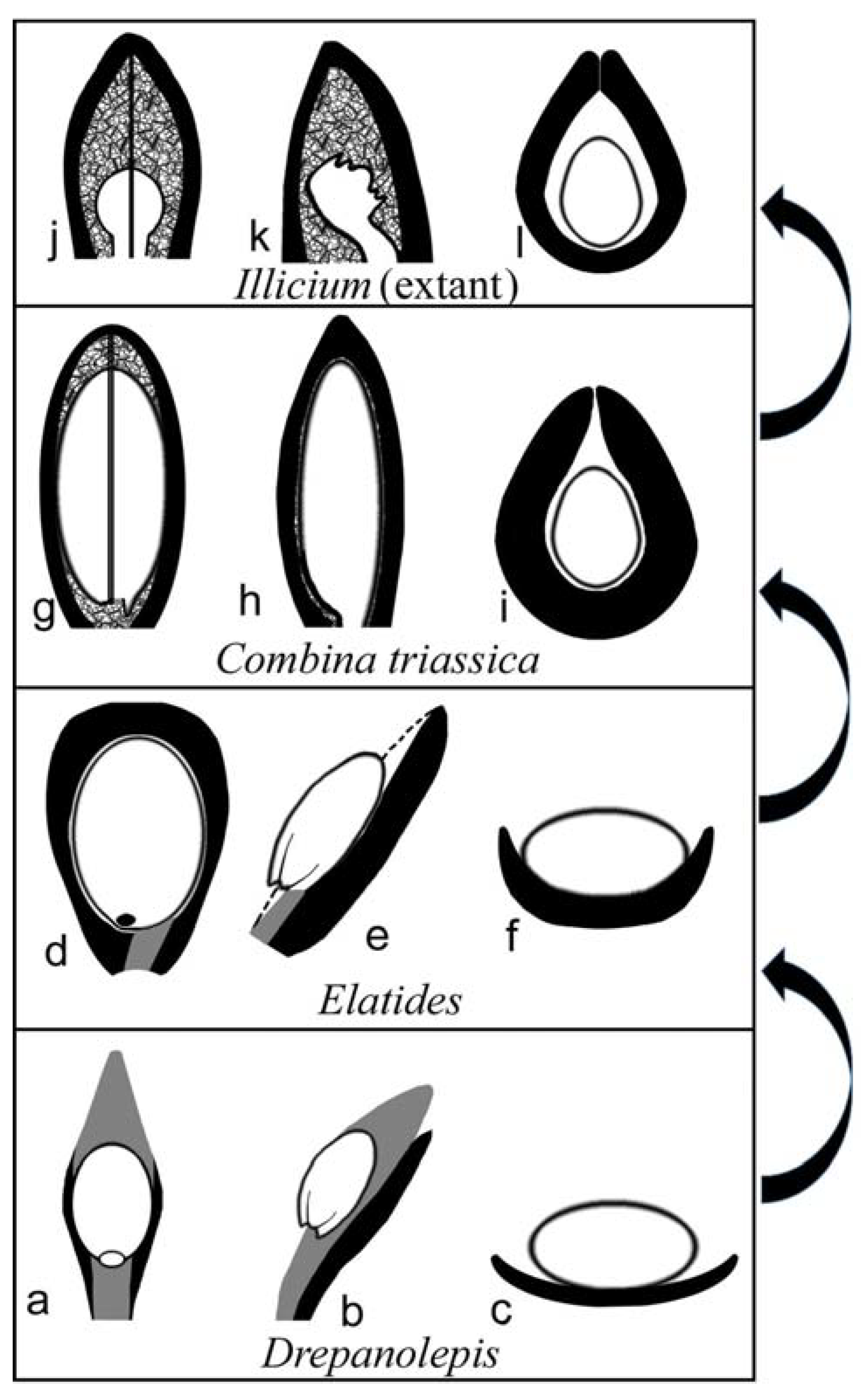
The general morphology of Combina appears to be that of a coniferous cone, in which a lateral unit is composed of an axillary scale and a subtending bract. According to Schweitzer and Kirchner [19], there are only two Mesozoic genera with unilobate one-seeded lateral units, one is Drepanolepis and the other Ontheodendron. Since Ontheodendron has been recognized as a fossil stem [20,21], there is only one fossil genus left for comparison, Drepanolepis. Although both Combina and Drepanolepis appear similar to conifer cones in inverted ovules/seeds, Drepanolepis figured by Schweitzer and Kirchner is much more elongated and slenderer than Combina; most importantly, the bract encloses the axillary ovule to an unprecedented extent and forms an adaxial suture in Combina (Figure 1a,d,e), which is one of the implementations of the universal evolution trend of plant and organism reproduction [22]. Hitherto, the latter has not been seen in any known conifer cones (including Drepanolepis). This comparison justifies Combina as a new genus. Despite these differences between Combina and Drepanolepis, the comparison between them is meaningful for the homology of carpel and plant systematics (see Figure 4).
Currently, there are two competing hypotheses in botany on the homology and origin of angiosperm carpels, the Traditional Theory [4,6,23] and the Unifying Theory [24,25,26]. According to the former, a carpel is derived from a megasporophyll bearing ovules along its margins through longitudinal folding and enrolling [4,6,23]. According to the latter [24,25,26], a carpel is a composite organ comprising two parts of different nature, an axillary placenta (ovule-bearing branch) and a subtending ovarian wall (leaf). Although the Unifying Theory is in line with the results of function gene studies [27,28,29,30] as well as anatomical and morphological studies [16,17,31], it requires further independent observations (especially of fossil plants) for confirmation.
It is evident that the ovules in Combina gen. nov. are borne directly on the organ axis (Figure 1b–j), not on the margins of any foliar part (bract), thus at odds with the widely-accepted Traditional Theory, which expects ovules on the margins of a leaf. It is noteworthy that Combina is not the only evidence against the Traditional Theory, as (1) the carpels in the previously assumed ancestral angiosperm, Michelia (Magnoliaceae) and Illicum (Schisandraceae), have been shown to be composed of a foliar part and an axillary ovuliferous branch [17] or an ovule directly borne on the floral axis [16]; (2) in addition, another Triassic fossil reproductive organ putatively related to angiosperm, Nubilora, has its ovules directly borne on the floral axis [25]; (3) the carpel precursor assumed by the Traditional Theory (namely, megasporophyll) has not yet been found in either fossil or extant plants despite a century-long intensive painstaking search [32,33].
Several attempts have been made to raise hypotheses other than the traditional theory to account for the origin of carpels [3,24,25,31,34,35]. Regrettably, no plausible new morphological interpretation of carpel origin was given under the framework of the APG system, which was mainly based on molecular data. Although Sauquet et al. [1] discussed the arrangement of carpels in ancestral angiosperm flowers, they did not touch on the problem of the origin of carpels; their conclusion became controversial immediately and triggered criticism and heated debates among botanists [36,37,38]. Recent studies of extant basal angiosperms (e.g., Illicium [16] and Michelia [17]) reinforce that a carpel comprises axillary ovule(s) and a subtending foliar part, a conclusion that has long been suggested by gene function studies [29,30]. Integrating the outcomes of previous independent studies, Wang [24,25] proposed that a carpel is a composite organ derived through synorganizing an ovuliferous branch and a subtending leaf, a pattern that is workable for Amborella [25] (the currently assumed basalmost angiosperm [8,39]) as well as Magnolia (the traditionally assumed basalmost angiosperm [4,6]). The Late Triassic Nubilora [25] favors this generalization, as its ovules are attached directly to the cone axis and enwrapped by foliar parts. However, this single piece of fossil evidence appears weak against the overwhelming thinking inertia in botany. Now, with its axillary ovule directly attached to the organ axis and almost fully enclosed by its subtending bract, Combina gen. nov. seems to fit into the scenario depicted in figure 6.3e of Taylor and Kirchner [3] and the cases 23–25 in figure 8.40 of Wang [25]. The difference between Combina and the anticipated precursor of carpel [3] is restricted to its anatropous configuration of its ovule, which may be interpreted as a highly reduced and metamorphosed version of a former ovuliferous branch. It is noteworthy that a single axillary ovule is enclosed by a subtending leaf in Illicium from bottom and laterals [16], especially similar to our observation of Combina. Thus, Combina appears to be a piece of evidence supporting the Unifying Theory [25].
The most intriguing fact about Combina gen. nov. is that the bracts of Combina gen. nov. have started enclosing their axillary ovules, although not fully (Figure 1b–j). Therefore, although we are not sure whether the ovules of Combina gen. nov. are fully enclosed before pollination (if so, then Combina is a bona fide angiosperm) for the time being, Combina gen. nov. is apparently knocking on the door of angiosperms. It is noteworthy that Endress [40] recently took a carpel as the result of synorganization between a foliar part and ovule(s). This point of view is in full agreement with the implications given by Combina gen. nov. The possibility of the bract forming an outer integument in Combina can be easily excluded by the following facts: (1) the bract is directly attached to the organ axis rather than to the funiculus (ovule base) in Combina; (2) the bract and ovules have distinct and separated contours in Combina (Figure 2a,b).
At the right time and with the right morphology, Combina gen. nov. appears to be an ideal precursor for angiosperm carpels, as it seems to have completed the evidence chain for carpel origination from a gymnospermous ancestor. The resemblance between Combina gen. nov. and the Palaeozoic conifers and Cordaitales [41]) (Figure 4a–l) seems to suggest that at least some angiosperms may share a common ancestor with some “conifers” and Cordaitales, if the previous axillary ovuliferous branches (as in Cordaitales [41], Palissya [42,43,44] , Metridiostrobus [45], and Stachyotaxus [44,46]) are reduced into a single ovule. Although we admit that relating conifers to angiosperms is at odds with most systematists (who cannot offer a plausible solution for carpel origin, however), our current proposal appears optimal, at least in terms of carpel homology, in the current academic context. Although we cannot determine, for the time being, that Combina is an angiosperm or an ancestral angiosperm, and the ovules in Combina are not fully enclosed as in angiosperms, it is noteworthy that, among all fossil plants, Combina demonstrates an unprecedented way of ovule-enclosing similar to that in some angiosperms (Illicium). This information is helpful for botanists trying to piece together the picture of plant evolution.
4. Materials and Methods
The plants associated with Combina triassica gen. et sp. nov. included Anomopteris mougeotii, Endolepis sp., Equisetites sp., Neocalamites sp., N. cf. carrerei, Pelourdea vogesiaca, Voltzia heterophylla, V. walchiaeformis, V. sp., and Willsiostrobus rhomboidalis, and the palynological assemblage was characterized by the occurrence of index taxa such as Hexasaccites muelleri, Alisporites grauvogeli, Voltziaceasporites heteromorpha, and several forms of Triadispora (T. aurea, T. crassa, T. epigona, T. falcata, and T. staplinii) [18]. A previous study indicated that Combina gen. nov. was from the Calcena Formation, Anisian, lower Middle Triassic [18].
Photographs were taken using a Nikon D-90 camera with an AF-S Micro Nikkor 60-mm macro lens (Canon Europa N.V., Bovenkerkerweg 59 1185 XB Amstelveen, Netherland). Cross-polarized illumination was used following the technique of image acquisition described by Kerp and Bomfleur [47]. Photographs from Scanning Electron Microscope (SEM) were taken with a JEOL JSM6010LA at CACTI (Centro de Apoio Científico-Tecnolóxico á Investigación, University of Vigo, Ourense, Spain). The pictures were organized for publication using Photoshop 7.0.
Author Contributions
A.A.S. collected the specimen. X.W. initiated the study. X.W. and A.A.S. drafted the manuscript, modified, and finalized the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Strategic Priority Research Program (B) of the Chinese Academy of Sciences (Grant No. XDB26000000) and the National Natural Science Foundation of China (42288201, 41688103, 91514302). A.A.S. is awarded with a fellowship from the Galician Government (Department of Culture, Education and University Planning) supported by the European Social Fund (Ref.: ED481A-2019/243).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank José Bienvenido Diez Ferrer and the staff of the Natural Sciences Museum of the University of Zaragoza for their help in accessing the fossil specimen.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Sauquet, H.; von Balthazar, M.; Magallón, S.; Doyle, J.A.; Endress, P.K.; Bailes, E.J.; Barroso de Morais, E.; Bull-Hereñu, K.; Carrive, L.; Chartier, M.; et al. The ancestral flower of angiosperms and its early diversification. Nat. Commun. 2017, 8, 16047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.-T.; Yi, T.-S.; Gao, L.-M.; Ma, P.-F.; Zhang, T.; Yang, J.-B.; Gitzendanner, M.A.; Fritsch, P.W.; Cai, J.; Luo, Y.; et al. Origin of angiosperms and the puzzle of the Jurassic gap. Nat. Plants 2019, 5, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.W.; Kirchner, G. The origin and evolution of the angiosperm carpel. In Flowering Plant Origin, Evolution and Phylogeny; Taylor, D.W., Hickey, L.J., Eds.; Chapman AND Hall: New York, NY, USA, 1996; pp. 116–140. [Google Scholar]
- Arber, E.A.N.; Parkin, J. On the origin of angiosperms. J. Linn. Soc. Lond. Bot. 1907, 38, 29–80. [Google Scholar] [CrossRef] [Green Version]
- Eames, A.J. Morphology of the Angiosperms; McGraw-Hill Book Company, Inc.: New York, NY, USA, 1961; p. 518. [Google Scholar]
- Cronquist, A. The Evolution and Classification of Flowering Plants, 2nd ed.; New York Botanical Garden: New York, NY, USA, 1988; p. 555. [Google Scholar]
- Takhtajan, A. Flowering Plants, Origin and Dispersal; Oliver AND Boyd Ltd.: Edinburgh, UK, 1969; p. 310. [Google Scholar]
- Qiu, Y.-L.; Lee, J.; Bernasconi-Quadroni, F.; Soltis, D.E.; Soltis, P.S.; Zanis, M.; Zimmer, E.A.; Chen, Z.; Savolainen, V.; Chase, M.W. The earliest angiosperms: Evidence from mitochondrial, plastid and nuclear genomes. Nature 1999, 402, 404–407. [Google Scholar] [CrossRef]
- Endress, P.K.; Doyle, J.A. Reconstructing the ancestral angiosperm flower and its initial specializations. Am. J. Bot. 2009, 96, 22–66. [Google Scholar] [CrossRef]
- Sokoloff, D.D.; Remizowa, M.V.; El, E.S.; Rudall, P.J.; Bateman, R.M. Supposed Jurassic angiosperms lack pentamery, an important angiosperm-specific feature. New Phytol. 2020, 228, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Coiro, M.; Doyle, J.A.; Hilton, J. How deep is the conflict between molecular and fossil evidence on the age of angiosperms? New Phytol. 2019, 223, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Bateman, R.M. Hunting the snark: The flawed search for mythical Jurassic angiosperms. J. Exp. Bot. 2020, 71, 22–35. [Google Scholar] [CrossRef]
- Fu, Q.; Diez, J.B.; Pole, M.; García-Ávila, M.; Wang, X. Nanjinganthus is an angiosperm, isn’t it? China Geol. 2020, 3, 359–361. [Google Scholar] [CrossRef]
- Fu, Q.; Diez, J.B.; Pole, M.; Garcia-Avila, M.; Liu, Z.-J.; Chu, H.; Hou, Y.; Yin, P.; Zhang, G.-Q.; Du, K.; et al. An unexpected noncarpellate epigynous flower from the Jurassic of China. eLife 2018, 7, e38827. [Google Scholar] [CrossRef]
- Taylor, D.W.; Li, H. Paleobotany: Did flowering plants exist in the Jurassic period? eLife 2018, 7, e43421. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X. Floral ontogeny of Illicium lanceolatum (Schisandraceae) and its implications on carpel homology. Phytotaxa 2019, 416, 200–210. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, W.; Wang, X. How the ovules get enclosed in magnoliaceous carpels. PLoS ONE 2017, 12, e0174955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diez, J.B.; Broutin, J.; Grauvogel-Stamm, L.; Bourquin, S.; Bercovici, A.; Ferrer, J. Anisian floras from the NE Iberian Peninsula and Balearic Islands: A synthesis. Rev. Palaeobot. Palynol. 2010, 162, 522–542. [Google Scholar] [CrossRef]
- Schweitzer, H.-J.; Kirchner, M. Die Rhaeto-Jurassischen Floren des Iran und Afghanistans: 9. Coniferophyta. Paläontographica B 1996, 238, 77–139. [Google Scholar]
- Rao, A.R.; Bose, M.N. Further observations on Ontheodendron florini Sahni & Rao. Palaeobotanist 1958, 7, 29–31. [Google Scholar]
- Bose, M.N.; Maheshwari, H.K. The genus Ontheodendron Sahni and Rao. Palaeobotanist 1974, 23, 231–232. [Google Scholar]
- Fu, Q.; Liu, J.; Wang, X. Offspring development conditioning (ODC): A universal evolutionary trend in sexual reproduction of organisms. J. Northwest Univ. (Nat. Sci. Ed.) 2021, 51, 163–172. [Google Scholar]
- Canright, J.E. The comparative morphology and relationships of the Magnoliaceae. III. Carpels. Am. J. Bot. 1960, 47, 145–155. [Google Scholar] [CrossRef]
- Wang, X. The Dawn Angiosperms; Springer: Heidelberg, Germany, 2010. [Google Scholar]
- Wang, X. The Dawn Angiosperms, 2nd ed.; Springer: Cham, Switzerland, 2018; p. 407. [Google Scholar]
- Wang, X.; Liu, Z.-J.; Liu, W.; Liao, W.; Zhang, X.; Liu, Z.; Hu, G.; Guo, X.; Wang, Y. Stepping out of the shadow of Goethe: For a more scientific plant systematics. Chin. Bull. Bot. 2020, 55, 505–512. [Google Scholar]
- Roe, J.L.; Nemhauser, J.L.; Zambryski, P.C. TOUSLED participates in apical tissue formation during gynoecium development in Arabidopsis. Plant Cell 1997, 9, 335–353. [Google Scholar] [PubMed] [Green Version]
- Rounsley, S.D.; Ditta, G.S.; Yanofsky, M.F. Diverse roles for MADS box genes in Arabidopsis development. Plant Cell 1995, 7, 1259–1269. [Google Scholar] [PubMed] [Green Version]
- Skinner, D.J.; Hill, T.A.; Gasser, C.S. Regulation of ovule development. Plant Cell 2004, 16 (Suppl. 1), S32–S45. [Google Scholar] [CrossRef] [PubMed]
- Mathews, S.; Kramer, E.M. The evolution of reproductive structures in seed plants: A re-examination based on insights from developmental genetics. New Phytol. 2012, 194, 910–923. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.-Z.; Hilu, K.; Wang, Y.-L. From leaf and branch into a flower: Magnolia tells the story. Bot. Stud. 2014, 55, 28. [Google Scholar] [CrossRef]
- Wang, X.; Luo, B. Mechanical pressure, not genes, makes ovulate parts leaf-like in Cycas. Am. J. Plant Sci. 2013, 4, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Miao, Y.; Liu, Z.J.; Wang, M.; Wang, X. Fossil and living cycads say “No more megasporophylls”. J. Morphol. Anat. 2017, 1, 107. [Google Scholar]
- Melville, R. A new theory of the angiosperm flower: I. The gynoecium. Kew Bull. 1962, 16, 1–50. [Google Scholar] [CrossRef]
- Taylor, D.W. Angiosperm ovule and carpels: Their characters and polarities, distribution in basal clades, and structural evolution. Postilla 1991, 208, 1–40. [Google Scholar]
- Rümpler, F.; Theißen, G. Reconstructing the ancestral flower of extant angiosperms: The ‘war of the whorls’ is heating up. J. Exp. Bot. 2019, 70, 2615–2622. [Google Scholar] [CrossRef]
- Sokoloff, D.; Remizowa, M.V.; Bateman, R.M.; Rudall, P.J. Was the ancestral angiosperm flower whorled throughout? Am. J. Bot. 2018, 105, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledford, H. Deatabe blooms over anatomy of the world’s first flower. Nature 2018, 554, 153–154. [Google Scholar] [CrossRef] [Green Version]
- APG. APG IV: An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Endress, P.K. The morphological relationship between carpels and ovules in angiosperms: Pitfalls of morphological interpretation. Bot. J. Linn. Soc. 2019, 189, 201–227. [Google Scholar] [CrossRef] [Green Version]
- Florin, R. Evolution in cordaites and conifers. Acta Horti Berginal 1951, 15, 285–388. [Google Scholar]
- Schweitzer, H.-J. Der weibliches Zapfen von Pseudovoltzia liebeana und seine Bedeutung fuer die Phylogenie der Koniferen. Paläontographica B 1963, 113, 1–29. [Google Scholar]
- Wang, Z. A bizarre Palissya ovulate organ from Upper Triassic strata of the Zixing coal field, Hunan Province, China. Chin. Sci. Bull. 2012, 57, 1169–1177. [Google Scholar] [CrossRef] [Green Version]
- Pattemore, G.A.; Rigby, J.F.; Playford, G. Palissya: A global review and reassessment of Eastern Gondwanan material. Rev. Palaeobot. Palynol. 2014, 210, 50–61. [Google Scholar] [CrossRef]
- Delevoryas, T.; Hope, R.C. More evidence for conifer diversity in the upper Triassic of North Carolina. Am. J. Bot. 1981, 68, 1003–1007. [Google Scholar] [CrossRef]
- Arndt, S. Morphologie und Systematik ausgewählter Mesozoischer Koniferen. Paläontographica B 2002, 262, 1–23. [Google Scholar]
- Kerp, H.; Bomfleur, B. Photography of plant fossils-new techniques, old tricks. Rev. Palaeobot. Palynol. 2011, 166, 117–151. [Google Scholar] [CrossRef]
Figure 1.
General view and details of the holotype of Combina gen. nov. specimen number MPZ2009-425. (a) The organ including multiple helically arranged lateral units. Scale bar = 10 mm. (b) Detailed view of the lateral unit 3 in (a). Scale bar = 1 mm. (c) The same as in (b), under SEM. The ovule is colored artificially. Scale bar = 1 mm. (d) SEM view of the lateral unit 1 in (a), showing the adaxial suture (arrow) of the bract. The ovule base is colored artificially. Scale bar = 1 mm. (e) View of the lateral unit 1 in (a), showing the adaxial suture (arrow) of the bract. Scale bar = 1 mm. (f) Detailed view of the basal portion of the ovule (artificially colored) that is eclipsed by the adaxial part of the bract, under SEM, enlarged from (d). Scale bar = 0.5 mm. (g) View of the lateral unit 4 in (a), showing the ovule “sandwiched” adaxially and abaxially by its bract. Scale bar = 2 mm. (h) Detailed view of the ovule (arrows), enlarged from (g). Scale bar = 1 mm. (i) Detailed view of the ovule base, enlarged from (b), showing its physical connection (funiculus, between white arrows) to the organ axis (oa). Note the asymmetrical connection (black arrow) between the funiculus and the ovule. Refer to Figure 3a. Scale bar = 1 mm. (j) Detailed view of the lateral unit 2 in (a), showing the asymmetrical connection (arrow) between the ovule and its funiculus. Refer to Figure 3b. Scale bar = 1 mm.
Figure 1.
General view and details of the holotype of Combina gen. nov. specimen number MPZ2009-425. (a) The organ including multiple helically arranged lateral units. Scale bar = 10 mm. (b) Detailed view of the lateral unit 3 in (a). Scale bar = 1 mm. (c) The same as in (b), under SEM. The ovule is colored artificially. Scale bar = 1 mm. (d) SEM view of the lateral unit 1 in (a), showing the adaxial suture (arrow) of the bract. The ovule base is colored artificially. Scale bar = 1 mm. (e) View of the lateral unit 1 in (a), showing the adaxial suture (arrow) of the bract. Scale bar = 1 mm. (f) Detailed view of the basal portion of the ovule (artificially colored) that is eclipsed by the adaxial part of the bract, under SEM, enlarged from (d). Scale bar = 0.5 mm. (g) View of the lateral unit 4 in (a), showing the ovule “sandwiched” adaxially and abaxially by its bract. Scale bar = 2 mm. (h) Detailed view of the ovule (arrows), enlarged from (g). Scale bar = 1 mm. (i) Detailed view of the ovule base, enlarged from (b), showing its physical connection (funiculus, between white arrows) to the organ axis (oa). Note the asymmetrical connection (black arrow) between the funiculus and the ovule. Refer to Figure 3a. Scale bar = 1 mm. (j) Detailed view of the lateral unit 2 in (a), showing the asymmetrical connection (arrow) between the ovule and its funiculus. Refer to Figure 3b. Scale bar = 1 mm.

Figure 2.
An isolated part associated with the holotype of Combina gen. et sp. nov. (a) An adaxial view of an anatropous ovule (s) in a bract (b) axil. Specimen number R4/40/38/5. Scale bar = 1 mm. (b) Detailed view of the ovule base in (a), showing the asymmetrical funiculus (white arrow) and the micropyle (black arrow) of the anatropous ovule. Scale bar = 1 mm.
Figure 2.
An isolated part associated with the holotype of Combina gen. et sp. nov. (a) An adaxial view of an anatropous ovule (s) in a bract (b) axil. Specimen number R4/40/38/5. Scale bar = 1 mm. (b) Detailed view of the ovule base in (a), showing the asymmetrical funiculus (white arrow) and the micropyle (black arrow) of the anatropous ovule. Scale bar = 1 mm.
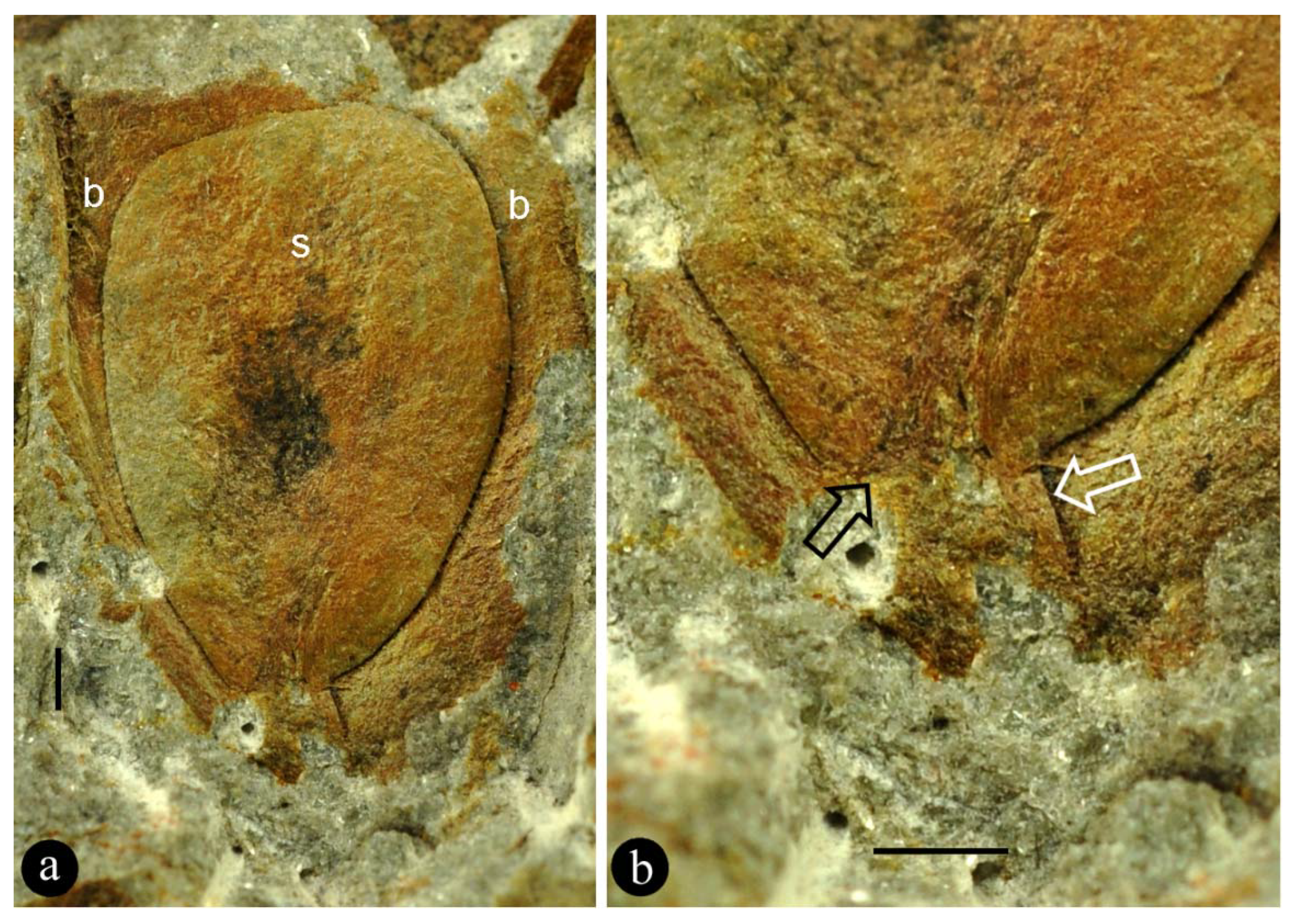
Figure 3.
Sketches of Combina. (a) Asymmetrical ovule base in Figure 1i. (b) Asymmetrical ovule base in Figure 1j.
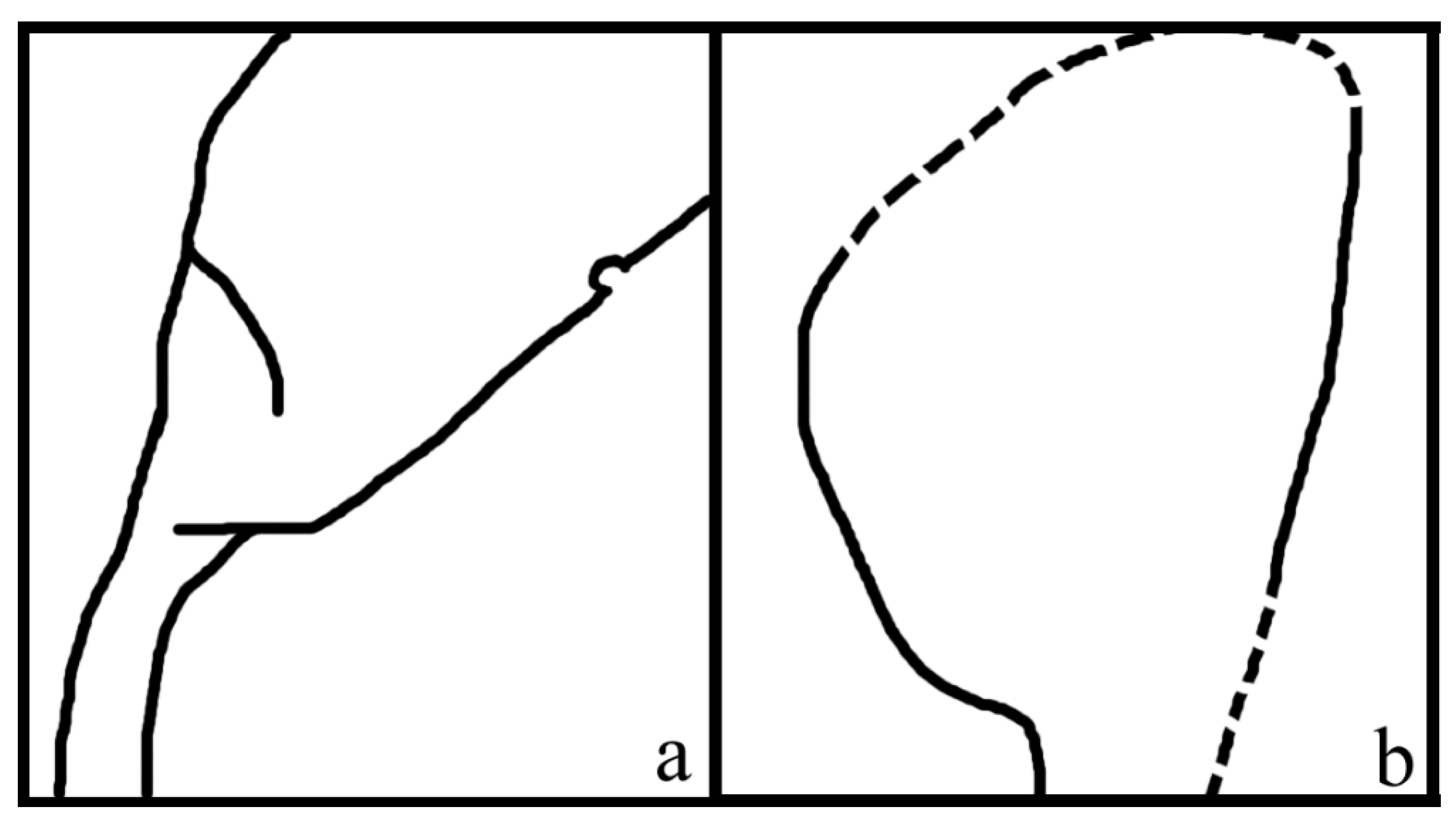
Figure 4.
Proposed possible evolutionary roadmap related to Combina gen. nov., from a conifer (Drepanolepis) to extant Illicium. The scale/funiculus is in dark gray, the ovule is in white, and the bract is in black. (a–c) Adaxial, lateral, and cross view of a lateral unit in Drepanolepis. (d–f) Adaxial, lateral, and cross view of a lateral unit in Elatides23. Note the wings of the bract curving adaxially. (g–i) Adaxial, lateral, and cross view of a lateral unit in Combina gen. nov. Note the axillary ovule is almost fully enclosed except along the ventral suture (gap). (j–l) Adaxial, lateral, and cross view of a carpel in extant Illicium (Illiciaceae).
Figure 4.
Proposed possible evolutionary roadmap related to Combina gen. nov., from a conifer (Drepanolepis) to extant Illicium. The scale/funiculus is in dark gray, the ovule is in white, and the bract is in black. (a–c) Adaxial, lateral, and cross view of a lateral unit in Drepanolepis. (d–f) Adaxial, lateral, and cross view of a lateral unit in Elatides23. Note the wings of the bract curving adaxially. (g–i) Adaxial, lateral, and cross view of a lateral unit in Combina gen. nov. Note the axillary ovule is almost fully enclosed except along the ventral suture (gap). (j–l) Adaxial, lateral, and cross view of a carpel in extant Illicium (Illiciaceae).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Santos, A.A.; Wang, X. Pre-Carpels from the Middle Triassic of Spain. Plants 2022, 11, 2833. https://doi.org/10.3390/plants11212833
AMA Style
Santos AA, Wang X. Pre-Carpels from the Middle Triassic of Spain. Plants. 2022; 11(21):2833. https://doi.org/10.3390/plants11212833
Chicago/Turabian StyleSantos, Artai A., and Xin Wang. 2022. "Pre-Carpels from the Middle Triassic of Spain" Plants 11, no. 21: 2833. https://doi.org/10.3390/plants11212833
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.