
Diversity of Arbuscular Mycorrhizal Fungi Associated with Maize in the Eastern Part of Uganda †

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
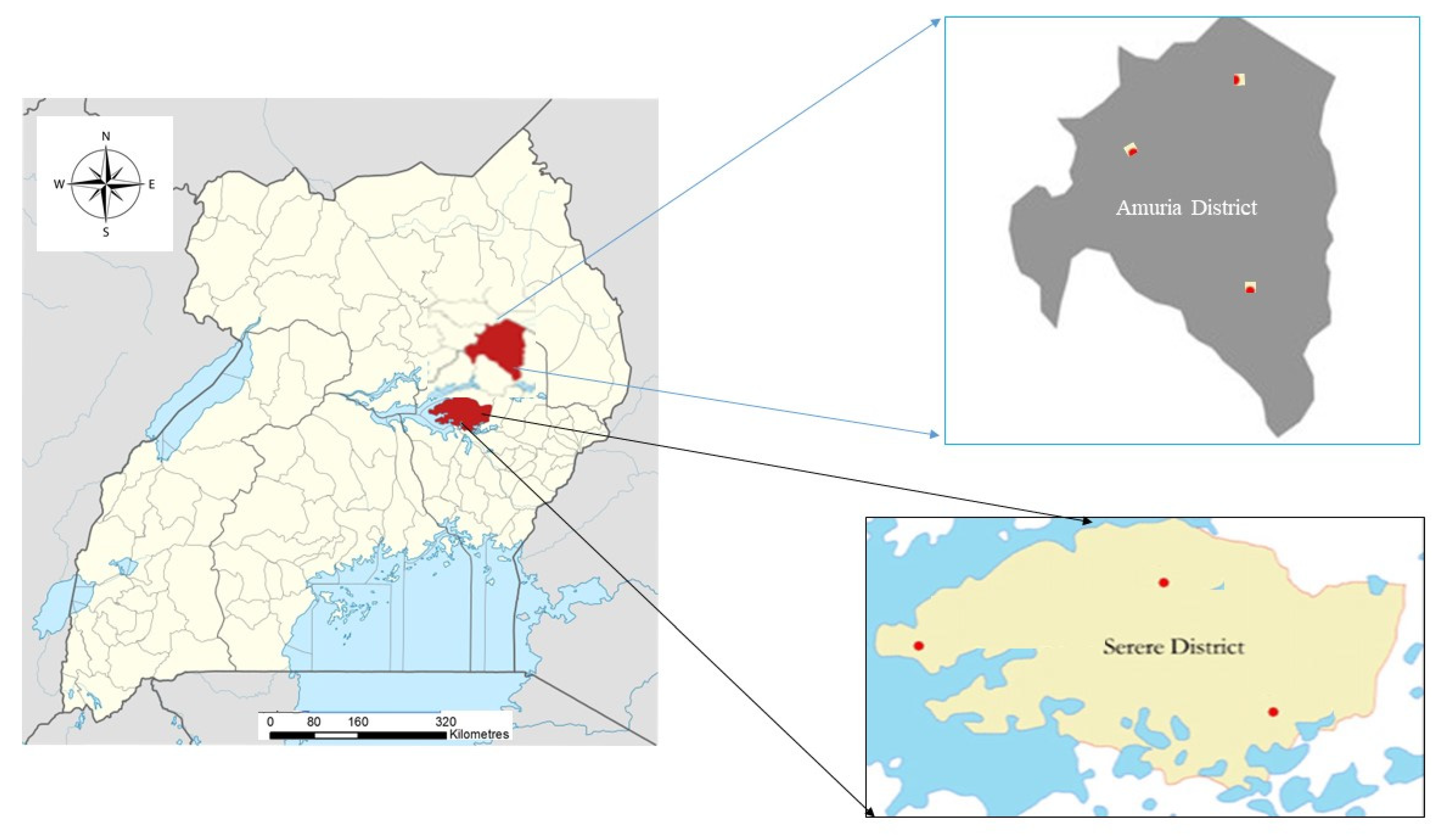
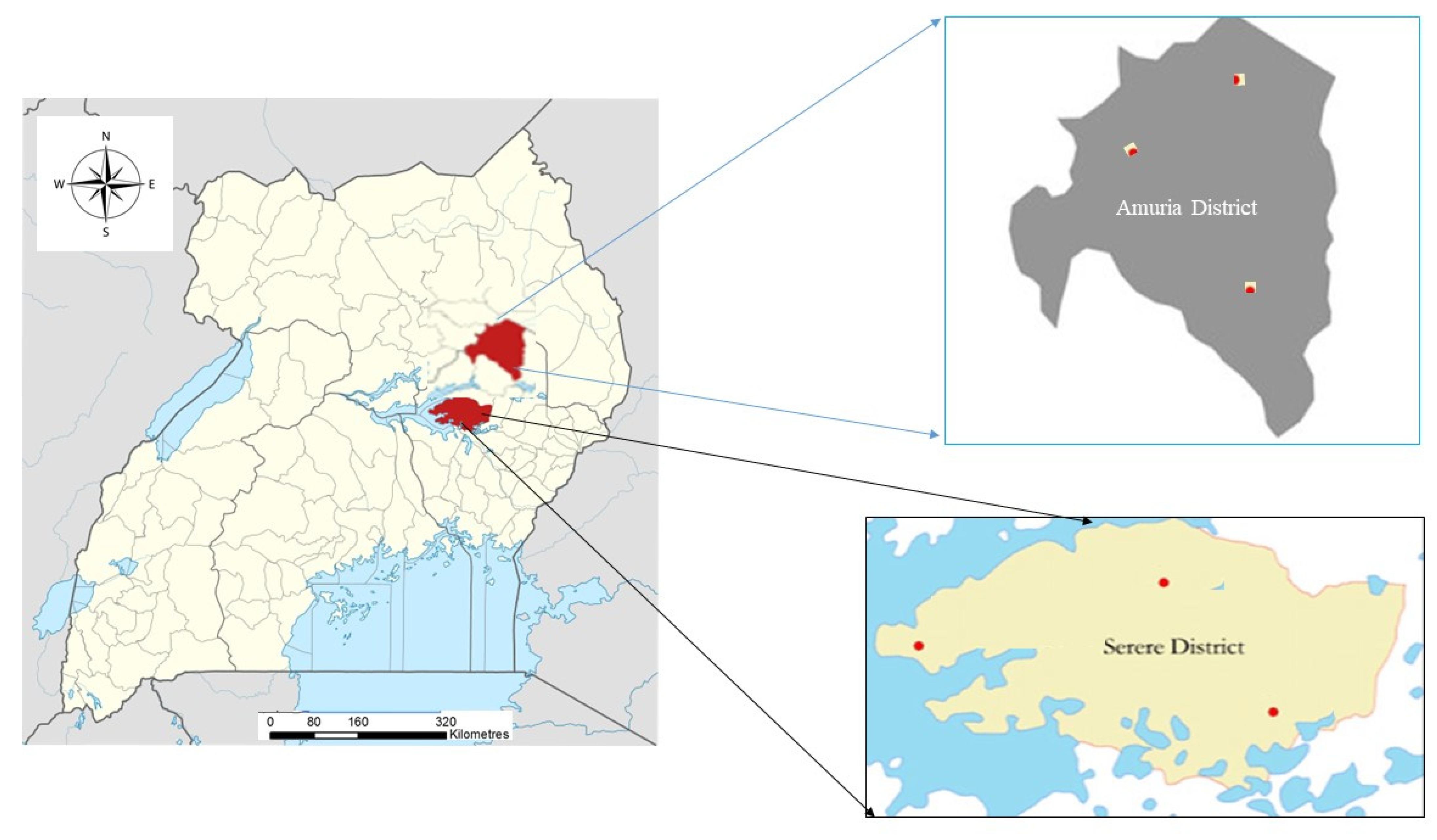
2.1. Study Area
2.2. Soils Sampling and Analysis
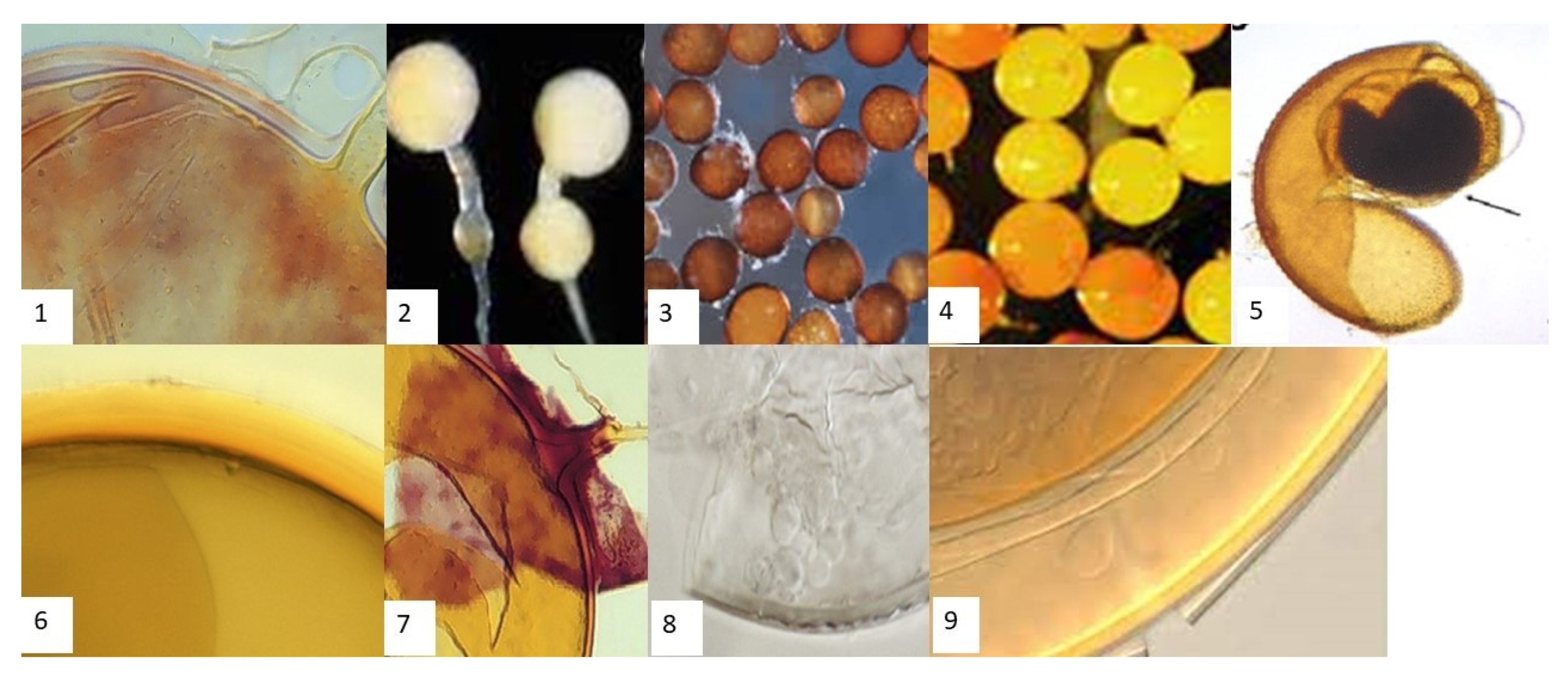
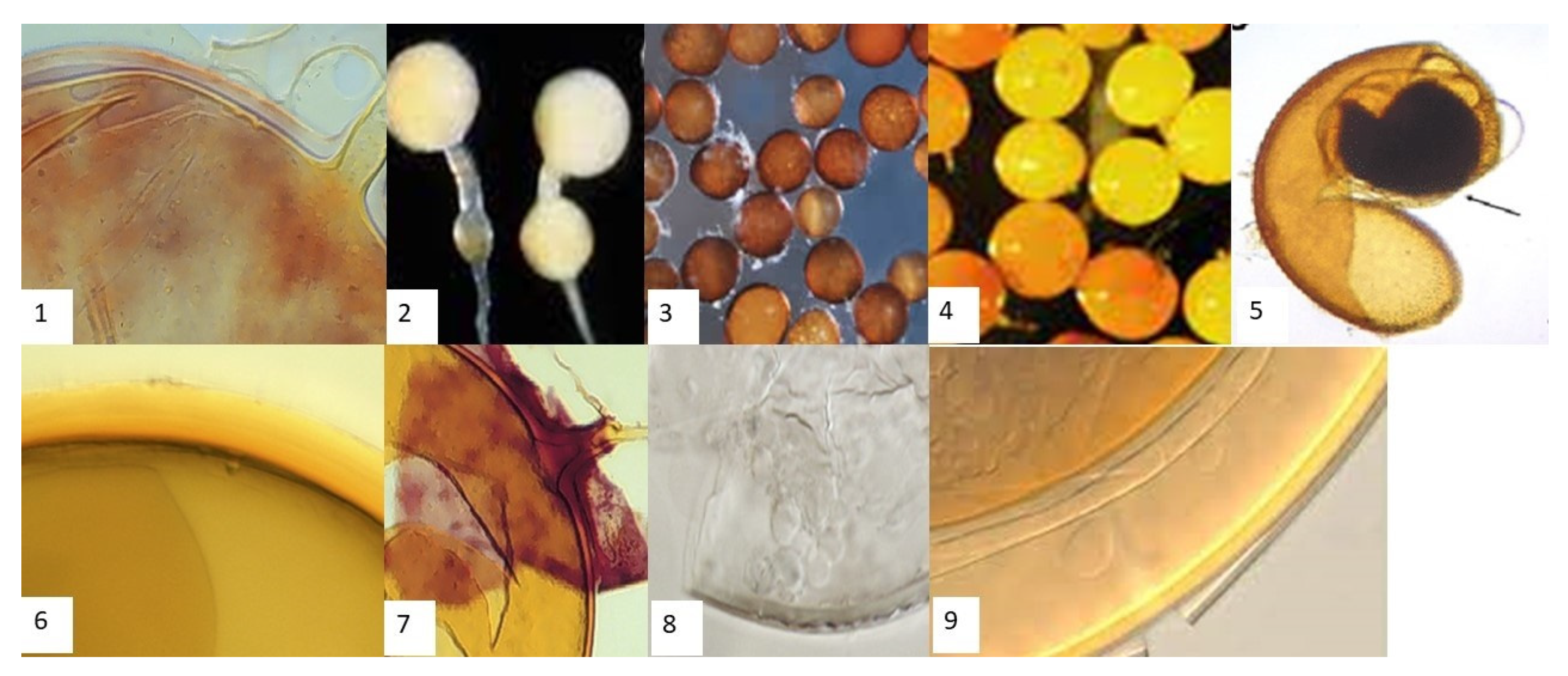
2.3. Extraction, Enumeration, and Morphological Identification of AMF Spores
2.4. AMF Roots Colonization Measurement
2.5. Statistical Analysis
3. Results
3.1. Soils Chemical and Physical Characterization
3.2. Intensity of Maize Mycorrhization
3.3. Diversity and Abundance of AMF Spores
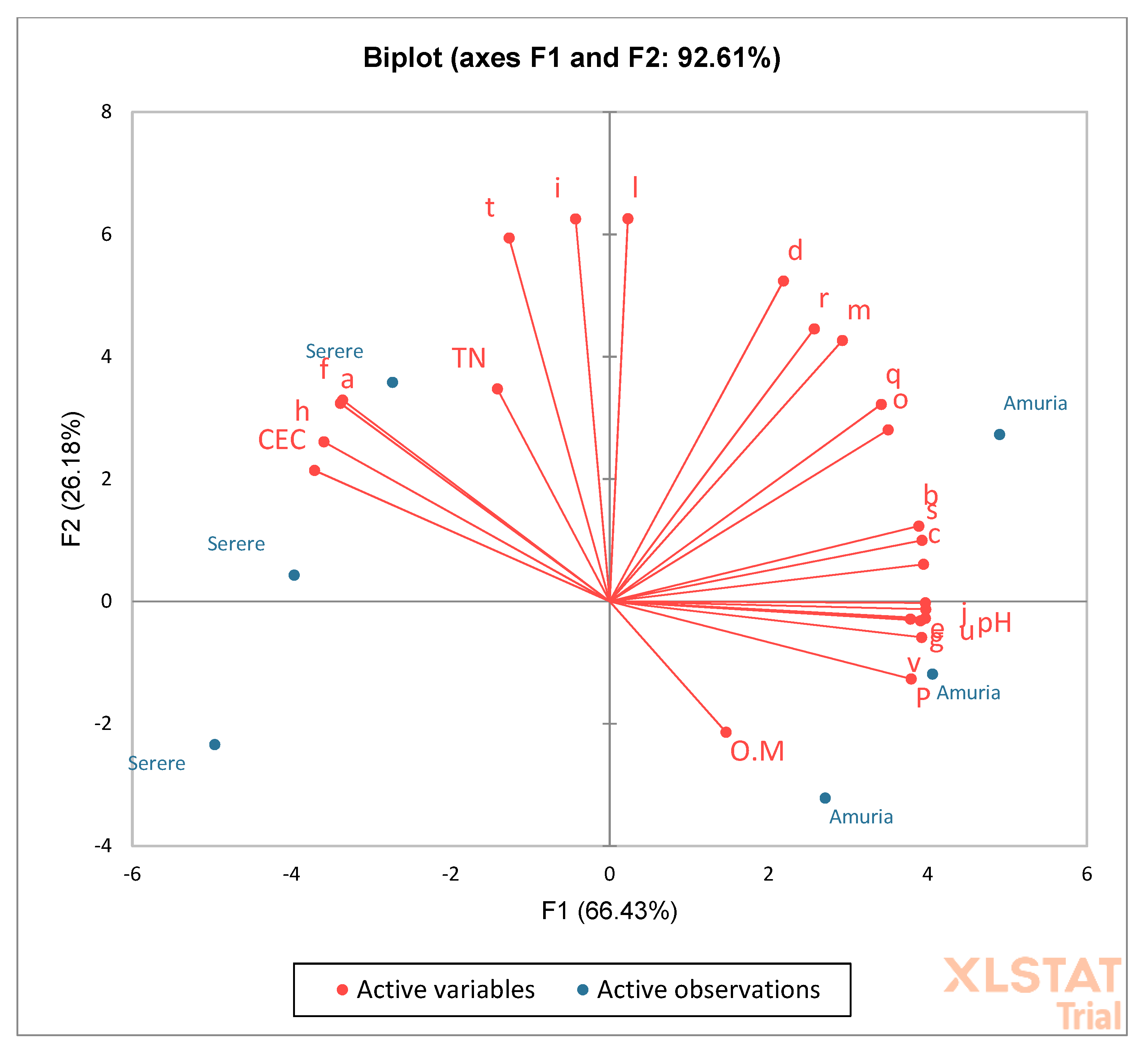
3.4. Relationship between Soil Chemical Properties and Species Diversity
4. Discussion
4.1. Intensity of Maize Mycorrhization
4.2. Diversity and Abundance of AMF Spores
4.3. Relationship between Soil Chemical Properties and Species Diversity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ekpa, O.; Palacios-Rojas, N.; Kruseman, G.; Fogliano, V.; Linnemann, A.R. Sub-Saharan African maize-based foods-processing practices, challe, needs, and opportunities. Food Rev. Int. 2019, 35, 609–639. [Google Scholar] [CrossRef] [Green Version]
- Mumo, L.; Yu, J.; Ojara, M.; Lukorito, C.; Kerandi, N. Assessing changes in climate suitability and yields of maize and sorghum crops over Kenya in the twenty-first century. Theor. Appl. Climatol. 2021, 146, 381–394. [Google Scholar] [CrossRef]
- Ochieng, I.O.; Gitari, H.I.; Mochoge, B.; Rezaei-Chiyaneh, E.; Gweyi-Onyango, J.P. Optimizing maize yield, nitrogen efficacy, and grain protein content under different N forms and rates. J. Soil Sci. Plant Nutr. 2021, 21, 1867–1880. [Google Scholar] [CrossRef]
- Wu, Y.; Messing, J. Proteome balancing of the maize seed for higher nutritional value. Front. Plant Sci. 2016, 5, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharon, B.; Michael, M.; Bwayo, M.F. Severity and prevalence of the destructive fall armyworm on maize in Uganda: A case of Bulambuli District. Afr. J. Agric. Res. 2020, 16, 777–784. [Google Scholar]
- UBOS. Uganda National Panel Survey 2015/2016, Technical Report 1224; Uganda Bureau of Statistics: Kampala, Uganda, 2016. [Google Scholar]
- FAO. World Food and Agriculture—Statistical Yearbook; FAO: Rome, Italy, 2020. [Google Scholar]
- Oloo, A. Constraints to Maize Production in Kamuli District, Eastern Uganda. Ph.D. Thesis, Makerere University, Kampala, Uganda, 2021. [Google Scholar]
- Okoboi, G.; Muwanga, J.; Mwebaze, T. Use of improved inputs and its effect on maize yield and profit in Uganda. Afr. J. Food Agric. Nutr. Dev. 2012, 12, 7. [Google Scholar] [CrossRef]
- Yadav, A.N. Plant Microbiomes for Sustainable Agriculture: Current Research and Future Challenges. In Sustainable Development and Biodiversity; Springer Nature: Cham, Switzerland, 2020; Volume 25, pp. 475–482. [Google Scholar]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef]
- Wu, Q.S. Arbuscular Mycorrhizas and Stress Tolerance of Plants; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Dowarah, B.; Gill, S.S.; Agarwala, N. Arbuscular Mycorrhizal Fungi in Conferring Tolerance to Biotic Stresses in Plants. J. Plant Growth Regul. 2022, 41, 1429–1444. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Y.; Jiang, S.; Deng, Y.; Christie, P.; Murray, P.J.; Li, X.; Zhang, J. Arbuscular mycorrhizal fungi in soil and roots respond differently to phosphorus inputs in an intensively managed calcareous agricultural soil. Sci. Rep. 2016, 6, 24902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leye, E.H.M.; Ndiaye, M.; Diouf, M.; Diop, T. Etude comparative de l’effet de souches de champignons mycorhiziens arbusculaires sur la croissance et la nutrition minérale du sésame cultivé au Sénégal. Afr. Crop Sci. J. 2015, 23, 211–219. [Google Scholar]
- Nyaga, J.; Jefwa, J.M.; Muthuri, C.W.; Matiru, V.N.; Wachira, P.M.; Okoth, S.A. Arbuscular mycorrhizal fungi with different soil fertility amendment practices in agricultural landscapes of Kenyan Highlands. Nutr. Cycl. Agroecosyst. 2015, 103, 229–240. [Google Scholar] [CrossRef]
- Msiska, Z. Arbuscular Mycorrhizal Fungi of Ugandan Banana Plantation Soils. Ph.D. Thesis, University of Pretoria, Pretoria, South Africa, 2007. [Google Scholar]
- Sebuliba, E.; Nyeko, P.; Majaliwa, J.G.M.; Kizza, L.C.; Eilu, G.; Adipala, E. Effect of selected arbuscular mycorrhiza fungi on the growth of Calliandra calothyrsus and Sorghum bicolor in eastern Uganda. In Proceedings of the Second RUFORUM Biennial Meeting, Entebbe, Uganda, 20–24 September 2010; pp. 20–24. [Google Scholar]
- Agwot, R.K. Contract Management and Service Delivery in Local Governments in Uganda: A case of Serere District Local Government. Ph.D. Thesis, Uganda Management Institute, Kampala, Uganda, 2018. [Google Scholar]
- Nsabagwa, M.; Mwije, A.; Nimusiima, A.; Odongo, R.I.; Ogwang, B.A.; Wasswa, P.; Mugume, I.; Basalirwa, C.; Nalwanga, F.; Kakuru, R.; et al. Examining the Ability of Communities to Cope with Food Insecurity due to Climate Change. Sustainability 2021, 13, 11047. [Google Scholar] [CrossRef]
- Egeru, A. Role of indigenous knowledge in climate change adaptation: A case study of the Teso Sub-Region, Eastern Uganda. Indian J. Tradit. Knowl. 2012, 11, 217–224. [Google Scholar]
- Okalebo, J.R.; Gathua, K.W.; Woomer, P.L. Laboratory Methods of Soil and Plant Analysis: A Working Manual, 2nd ed.; Sacred Africa: Nairobi, Kenya, 2002; Volume 21, pp. 25–26. [Google Scholar]
- Gerdemann, J.W.; Nicolson, T.H. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Oehl, F.; Sieverding, E.; Ineichen, K.; Mäder, P.; Boller, T.; Wiemken, A. Impact of land-use intensity on the species diversity of arbuscular mycorrhizal fungi in agroecosystems of Central Europe. Appl. Environ. Microbiol. 2003, 69, 2816–2824. [Google Scholar] [CrossRef] [Green Version]
- Sasvári, Z.; Magurno, F.; Galanics, D.; Hang, T.T.N.; Ha, T.T.H.; Luyen, N.D.; Posta, K. Isolation and identification of arbuscular mycorrhizal fungi from agricultural fields of Vietnam. Am. J. Plant Sci. 2012, 3, 1796–1801. [Google Scholar] [CrossRef] [Green Version]
- Redecker, D.; Schüßler, A.; Stockinger, H.; Stürmer, S.L.; Morton, J.B.; Walker, C. An evidence-based consensus for the classification of arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza 2013, 23, 515–531. [Google Scholar] [CrossRef]
- Trouvelot, A. Mesure du taux de mycorhization VA d’un système radiculaire. Recherche de méthodes d’estimation ayant une signification fonctionnelle. In Physiological and genetical aspects of mycorrhizae, Proceedings of the 1st European Symposium on Mycorrhizae, Dijon, France 1–5 July 1985; INRA: Paris, France, 1986; pp. 217–221. ISBN 2-85340-774-8. [Google Scholar]
- Morton, J.B.; Benny, G.L. Revised classification of arbuscular mycorrhizal fungi (Zygomycetes): A new order, Glomales, two new suborders, Glomineae and Gigasporineae, and two new families, Acaulosporaceae and Gigasporaceae, with an emendation of Glomaceae. Mycotaxon 1990, 37, 471–491. [Google Scholar]
- Houngnandan, P.; Yemadje, R.G.H.; Kane, A.; Boeckx, P.; Van Cleemput, O. Les glomales indigènes de la forêt claire à Isoberlinia doka (Craib et Stapf) à Wari-Maro au centre du Bénin. Tropicultura 2009, 27, 83–87. [Google Scholar]
- Tawaraya, K. Arbuscular mycorrhizal dependency of different plant species and cultivars. Soil Sci. Plant Nutr. 2003, 49, 655–668. [Google Scholar] [CrossRef]
- Garbaye, J. La Symbiose Mycorhizienne: Une Association Entre Les Plantes et Les Champignons. La Symbiose Mycorhizienne; Editions Quae: Versailles, France, 2013; pp. 1–280. ISBN 9782759219636. [Google Scholar]
- Fall, A.F.; Founoune-Mboup, H.; Diatta, S.; Diakhaté, S.; Ndoye, I. Arbuscular mycorrhizal fungi associated with the rhizosphere of Piliostigma reticulatum and Guiera senegalensis shrubs in Senegal. Afr. Crop. Sci. J. 2021, 29, 433. [Google Scholar] [CrossRef]
- Borriello, R.; Lumini, E.; Girlanda, M.; Bonfante, P.; Bianciotto, V. Effects of different management practices on arbuscular mycorrhizal fungal diversity in maize fields by a molecular approach. Biol. Fertil. Soils 2012, 48, 911–922. [Google Scholar] [CrossRef]
- Blaszkowski, J.; Czerniawska, B. Arbuscular mycorrhizal fungi (Glomeromycota) are associated with roots of Ammophila Arenaria growing in maritime dunes of Bornholm (Denmark). Acta Soc. Bot. Pol. 2011, 80, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Hijri, I.; Sýkorová, Z.; Oehl, F.; Ineichen, K.; Maeder, P.; Wiemken, A.; Redecker, D. Communities of arbuscular mycorrhizal fungi in arable soils are not necessarily low in diversity. Mol. Ecol. 2006, 15, 2277–2289. [Google Scholar] [CrossRef] [PubMed]
- Alguacil, M.M.; Lumini, E.; Roldan, A.; Salinas-Garcia, J.R.; Bonfante, P.; Bianciotto, V. The impact of tillage practices on arbuscular mycorrhizal fungal diversity in subtropical crops. Ecol. Appl. 2008, 18, 527–536. [Google Scholar] [CrossRef]
- Sasvári, Z.; Hornok, L.; Posta, K. The community structure of arbuscular mycorrhizal fungi in roots of maize grown in a 50-year monoculture. Biol. Fertil. Soils 2011, 47, 167–176. [Google Scholar] [CrossRef]
- Sanders, I.R. Specificity in the arbuscular mycorrhizal symbiosis. In Mycorrhizal Ecology; Springer: Berlin/Heidelberg, Germany, 2002; pp. 415–437. [Google Scholar]
- Jacquemyn, H.; Merckx, V.; Brys, R.; Tyteca, D.; Cammue, B.P.; Honnay, O.; Lievens, B. Analysis of network architecture reveals phylogenetic constraints on mycorrhizal specificity in the genus Orchis (Orchidaceae). New Phytol. 2011, 192, 518–528. [Google Scholar] [CrossRef]
- Hao, L.; Zhang, Z.; Hao, B.; Diao, F.; Zhang, J.; Bao, Z.; Guo, W. Arbuscular mycorrhizal fungi alter the microbiome structure of rhizosphere soil to enhance maize tolerance to La. Ecotoxicol. Environ. Saf. 2021, 212, 111996. [Google Scholar] [CrossRef]
- Chifflot, V.; Rivest, D.; Olivier, A.; Cogliastro, A.; Khasa, D. Molecular analysis of arbuscular mycorrhizal community structure and spores distribution in tree-based intercropping and forest systems. Agric. Ecosyst. Environ. 2009, 131, 32–39. [Google Scholar] [CrossRef]
- Vieira, C.K.; Marascalchi, M.N.; Rodrigues, A.V.; de Armas, R.D.; Stürmer, S.L. Morphological and molecular diversity of arbuscular mycorrhizal fungi in the revegetated iron-mining sites has the same magnitude as adjacent pristine ecosystems. J. Environ. Sci. 2018, 67, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Bossou, L.D.R.; Houngnandan, H.B.; Adandonon, A.; Zoundji, C.; Houngnandan, P. Diversité des champignons mycorhiziens arbusculaires associés à la culture du maïs (Zea mays L.) au Bénin. Int. J. Biol. Chem. Sci. 2019, 13, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Malembaka, R.E.B.; Onwonga, R.; Jefwa, J.; Ayuke, F.; Nabahungu, L. Diversity and distribution of arbuscular mycorrhizal fungi in maize (Zea mays) cropping fields in South Kivu, Democratic Republic of Congo. Afr. J. Agric. Res. 2021, 17, 604–617. [Google Scholar]
- Johnson, J.M.; Houngnandan, P.; Kane, A.; Sanon, K.B.; Neyra, M. Diversity patterns of indigenous arbuscular mycorrhizal fungi associated with the rhizosphere of cowpea (Vigna unguiculata (L.) Walp.) in Benin, West Africa. Pedobiologia 2013, 56, 121–128. [Google Scholar] [CrossRef]
- Mbogne, J.T.; Temegne, C.N.; Hougnandan, P.; Youmbi, E.; Tonfack, L.B.; Ntsomboh-Ntsefong, G. Biodiversity of arbuscular mycorrhizal fungi of pumpkins (Cucurbita spp.) under the influence of fertilizers in ferralitic soils of Cameroon and Benin. J. Appl. Biol. Biotechnol. 2015, 3, 1–10. [Google Scholar]
- Moreira, H.; Pereira, S.I.; Marques, A.P.; Rangel, A.O.; Castro, P.M. Mine land valorization through energy maize production enhanced by the application of plant growth-promoting rhizobacteria and arbuscular mycorrhizal fungi. Environ. Sci. Pollut. Res. 2016, 23, 6940–6950. [Google Scholar] [CrossRef] [PubMed]
- Crossay, T.; Cilia, A.; Cavaloc, Y.; Amir, H.; Redecker, D. Four new species of arbuscular mycorrhizal fungi (Glomeromycota) associated with endemic plants from ultramafic soils of New Caledonia. Mycol. Prog. 2018, 17, 729–744. [Google Scholar] [CrossRef]
- Songachan, L.S.; Kayang, H. Diversity and distribution of arbuscular mycorrhizal fungi in Solanum species growing in natural conditions. Agric. Res. 2012, 1, 258–264. [Google Scholar] [CrossRef]
- Bohrer, G.; Kagan-Zur, V.; Roth-Bejerano, N.; Ward, D.; Beck, G.; Bonifacio, E. Effects of different Kalahari-desert VA mycorrhizal communities on mineral acquisition and depletion from the soil by host plants. J. Arid Environ. 2003, 55, 193–208. [Google Scholar] [CrossRef]
- Rosendahl, S. Communities, Populations, and individuals of arbuscular mycorrhizal fungi. New Phytol. 2008, 178, 253–266. [Google Scholar] [CrossRef]
- Jansa, J.; Erb, A.; Oberholzer, H.R.; Šmilauer, P.; Egli, S. Soil and geography are more important determinants of indigenous arbuscular mycorrhizal communities than management practices in Swiss agricultural soils. Mol. Ecol. 2014, 23, 2118–2135. [Google Scholar] [CrossRef]
- Xiang, X.; Gibbons, S.M.; He, J.S.; Wang, C.; He, D.; Li, Q.; Ni, Y.; Chu, H. Rapid response of arbuscular mycorrhizal fungal communities to short-term fertilization in an alpine grassland on the Qinghai-Tibet Plateau. PeerJ 2016, 4, e2226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Yang, W.; Song, F.; Li, X. Diversity and composition of arbuscular mycorrhizal fungal communities in the cropland black soils of China. Glob. Ecol. Conserv. 2020, 22, e00964. [Google Scholar] [CrossRef]
- Séry, D.; Kouadjo, Z.G.; Voko, B.R.; Zeze, A. Selecting native arbuscular mycorrhizal fungi to promote cassava growth and increase yield under field conditions. Front. Microbiol. 2016, 7, 2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: A review. Plant Soil 2001, 237, 173–195. [Google Scholar] [CrossRef]
- Amijee, F.; Tinker, P.B.; Stribley, D.P. The development of endomycorrhizal root systems: VII. A detailed study of effects of soil phosphorus on colonization. New Phytol. 1989, 111, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Lagrange, A.; L’Huillier, L.; Amir, H. Mycorrhizal status of Cyperaceae from New Caledonian ultramafic soils: Effects of phosphorus availability on arbuscular mycorrhizal colonization of Costularia comosa under field conditions. Mycorrhiza 2013, 23, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Xiao, L.; Liu, R. Response of arbuscular mycorrhizal fungi and phosphorus solubilizing bacteria to remediation abandoned solid waste of coal mine. Int. J. Coal Sci. Technol. 2019, 6, 603–610. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Site | pH | %TN | %O.M | P (mg/kg) | K (mg/kg) | CEC | Soil Texture * |
|---|---|---|---|---|---|---|---|
| Amuria | 6.15 a | 0.047 a | 2.09 a | 9.52 a | 108.3 a | 8.6 a | Sandy loam |
| Serere | 5.0 b | 0.093 b | 1.952 a | 4.62 b | 73.7 b | 19.9 b | Clay loam |
| Amuria | Serere | LSD | |
|---|---|---|---|
| Number of AMF spores per gram of soil (average of three field replicates) | 5.1 a | 3.7 b | 2.34 |
| Total number of AMF species found at the field sites (sum of three field replicates) | 19 | 17 | - |
| H′ | 1.84 a | 1.81 a | 0.16 |
| Intensity of mycorrhization | 56% | 55% |
| Genus | Morpho-Species | Amuria | Serere |
|---|---|---|---|
| Acaulospora | Acaulospora spinose | 6.9 | 10.4 |
| Acaulospora foveata | 10.2 | 6.9 | |
| Acaulospora sp. | 12.6 | 7.3 | |
| Archaeospora | Archaeospora schenckii | 5.0 | 4.4 |
| Archaeospora sp. | 13.1 | 3.1 | |
| Entrophospora | Entrophospora colombiana | 6.6 | 10.7 |
| Entrophospora sp.1 | 15.3 | 6.8 | |
| Entrophospora sp.2 | 5.7 | 11.8 | |
| Gigaspora | Gigaspora gigantean | 8.6 | 9.0 |
| Gigaspora sp. | 12.3 | 4.6 | |
| Glomus | Glomus mossae | 7.3 | 7.8 |
| Glomus geosporum | 6.7 | 5.1 | |
| Glomus intraradices | 10.9 | 8.2 | |
| Glomus sp.1 | 6.3 | 4.4 | |
| Glomus sp.2 | 5.6 | 4.7 | |
| Scutellospora | Scutellospora. Pellucida | 13.3 | 8.2 |
| Scutellospora sp. | 6.0 | 7.3 | |
| Paraglomus | Paraglomus sp.1 | 5.1 | 0.0 |
| Paraglomus sp.2 | 5.3 | 0.0 | |
| The total number of spores identified | 310 | 226 |
| Morphotypes | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| a | b | c | d | e | f | g | h | i | j | l | m | o | q | r | s | t | u | v | |
| pH | −0.806 | 0.911 | 0.952 | 0.494 | 0.953 | −0.829 | 0.959 | −0.888 | −0.119 | 0.938 | 0.018 | 0.673 | 0.865 | 0.783 | 0.564 | 0.935 | −0.383 | 0.909 | 0.917 |
| TN | 0.511 | −0.198 | −0.299 | 0.188 | −0.357 | 0.546 | −0.397 | 0.518 | 0.511 | −0.314 | 0.463 | 0.117 | −0.136 | −0.006 | 0.105 | −0.268 | 0.679 | −0.300 | −0.345 |
| O.M | −0.500 | 0.262 | 0.298 | −0.045 | 0.361 | −0.403 | 0.357 | −0.387 | −0.392 | 0.357 | −0.325 | 0.041 | 0.066 | 0.145 | 0.223 | 0.309 | −0.350 | 0.345 | 0.388 |
| P | −0.917 | 0.886 | 0.933 | 0.346 | 0.958 | −0.896 | 0.951 | −0.938 | −0.302 | 0.953 | −0.171 | 0.578 | 0.741 | 0.711 | 0.518 | 0.911 | −0.485 | 0.929 | 0.947 |
| CEC | 0.981 | −0.852 | −0.889 | −0.229 | −0.940 | 0.967 | −0.944 | 0.982 | 0.447 | −0.938 | 0.283 | −0.457 | −0.654 | −0.635 | −0.368 | −0.867 | 0.600 | −0.948 | −0.964 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fall, A.F.; Nakabonge, G.; Ssekandi, J.; Founoune-Mboup, H.; Badji, A.; Balde, I.; Ndiaye, M. Diversity of Arbuscular Mycorrhizal Fungi Associated with Maize in the Eastern Part of Uganda. Biol. Life Sci. Forum 2022, 15, 12. https://doi.org/10.3390/IECD2022-12351
Fall AF, Nakabonge G, Ssekandi J, Founoune-Mboup H, Badji A, Balde I, Ndiaye M. Diversity of Arbuscular Mycorrhizal Fungi Associated with Maize in the Eastern Part of Uganda. Biology and Life Sciences Forum. 2022; 15(1):12. https://doi.org/10.3390/IECD2022-12351
Chicago/Turabian StyleFall, Abdoulaye Fofana, Grace Nakabonge, Joseph Ssekandi, Hassna Founoune-Mboup, Arfang Badji, Issa Balde, and Malick Ndiaye. 2022. "Diversity of Arbuscular Mycorrhizal Fungi Associated with Maize in the Eastern Part of Uganda" Biology and Life Sciences Forum 15, no. 1: 12. https://doi.org/10.3390/IECD2022-12351