
Tauroursodeoxycholic Acid Reduces Neuroinflammation but Does Not Support Long Term Functional Recovery of Rats with Spinal Cord Injury

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
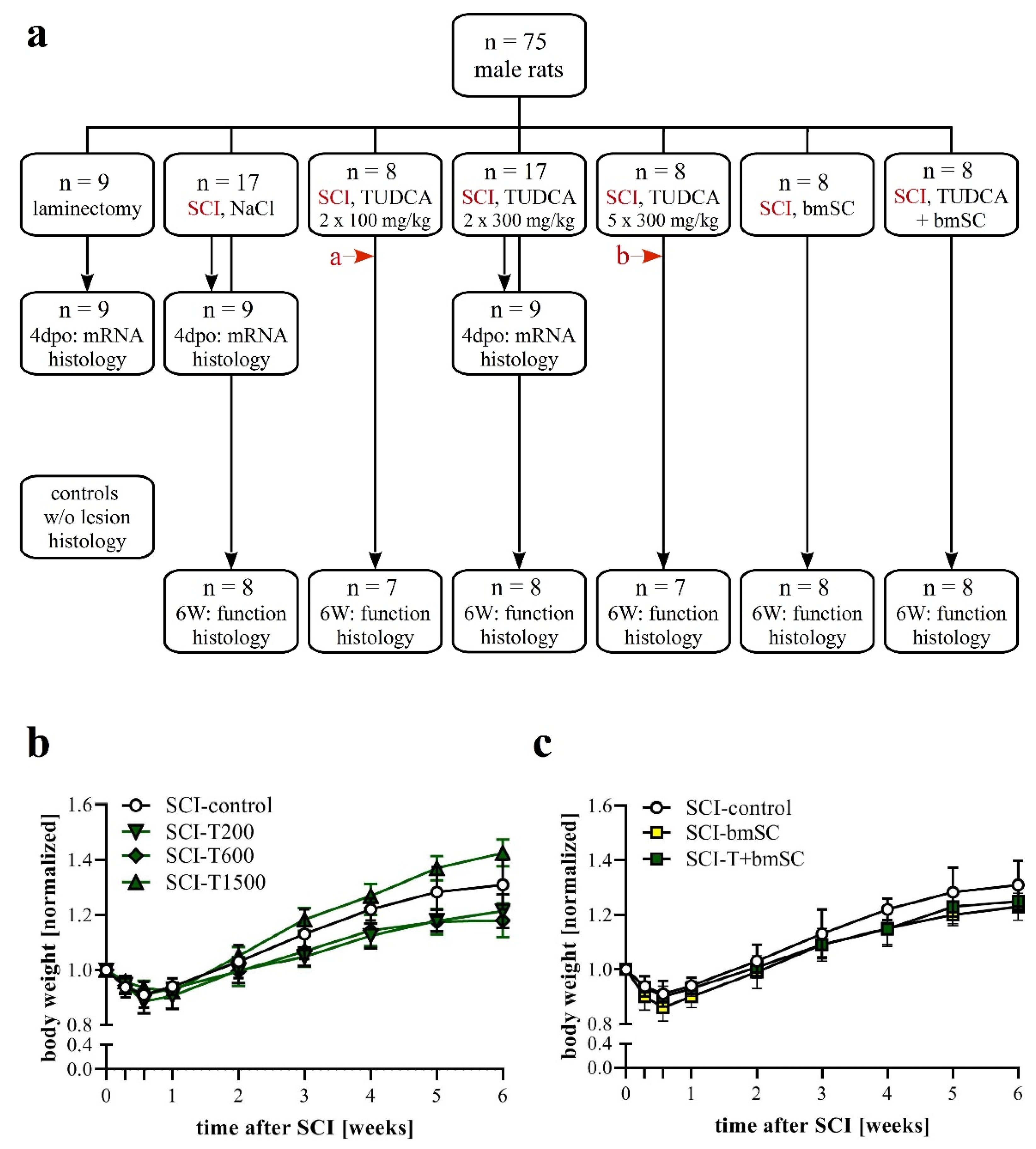
2.1. Experimental Animals and Study Design
2.2. Surgical Procedures and Postoperative Treatment
2.3. Experimental Groups
2.4. Preparation of bmSC
2.5. Bile Acid Treatment
2.6. Evaluation of Locomotor Functions
2.7. Von Frey Test of Mechanical Allodynia/Hyperalgesia
2.8. Quantification of Gene Expression
2.9. Tissue Preparation and Histological Staining
2.10. Immunofluorescence
2.11. Microscopy and Image Analysis
2.12. Statistical Analysis
3. Results
3.1. Effects of SCI, TUDCA, bmSC and TUDCA/bmSC Combinatorial Treatment on the General Health Status and Body Weight of the Animals
3.2. Expression of TGR5 after SCI
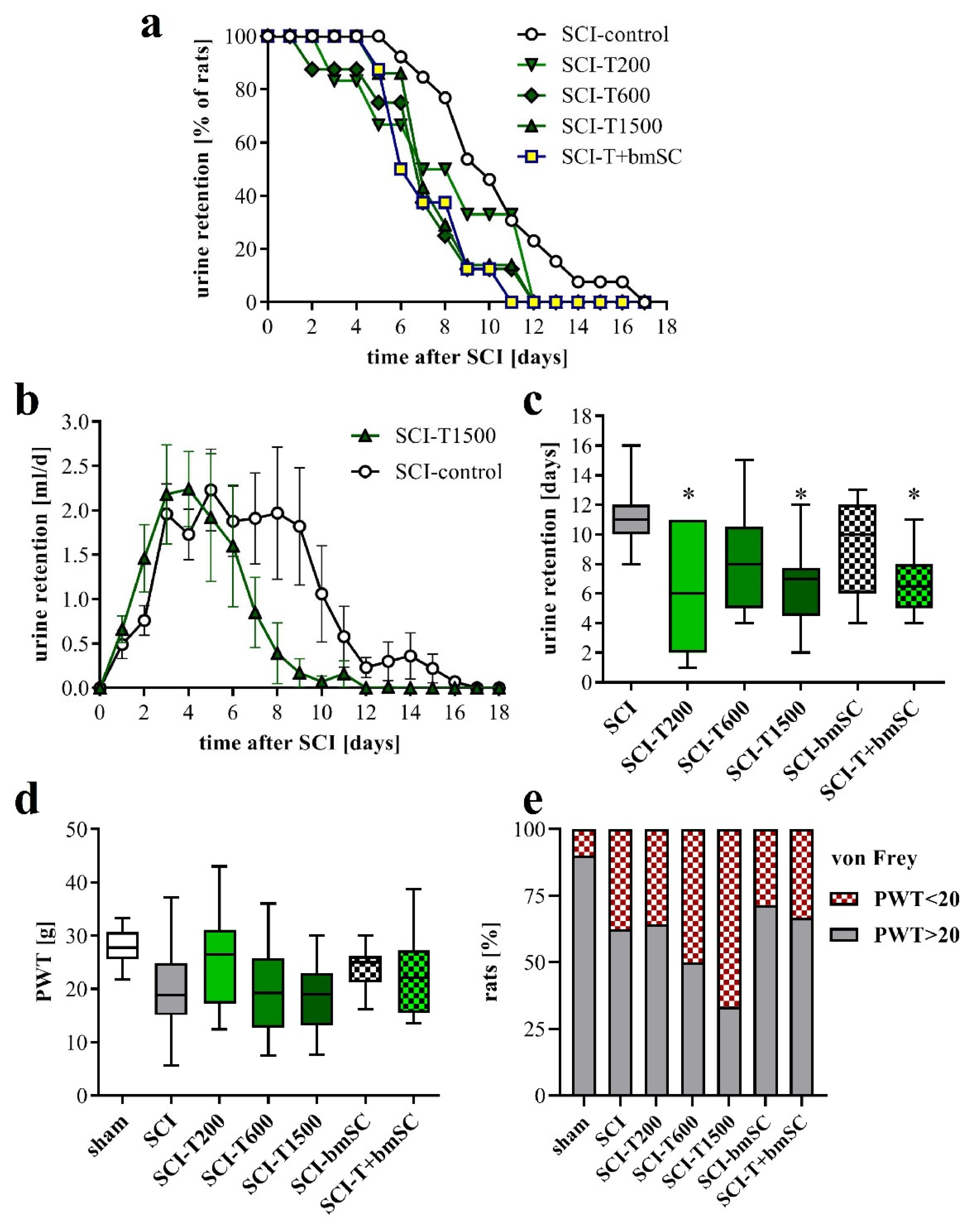
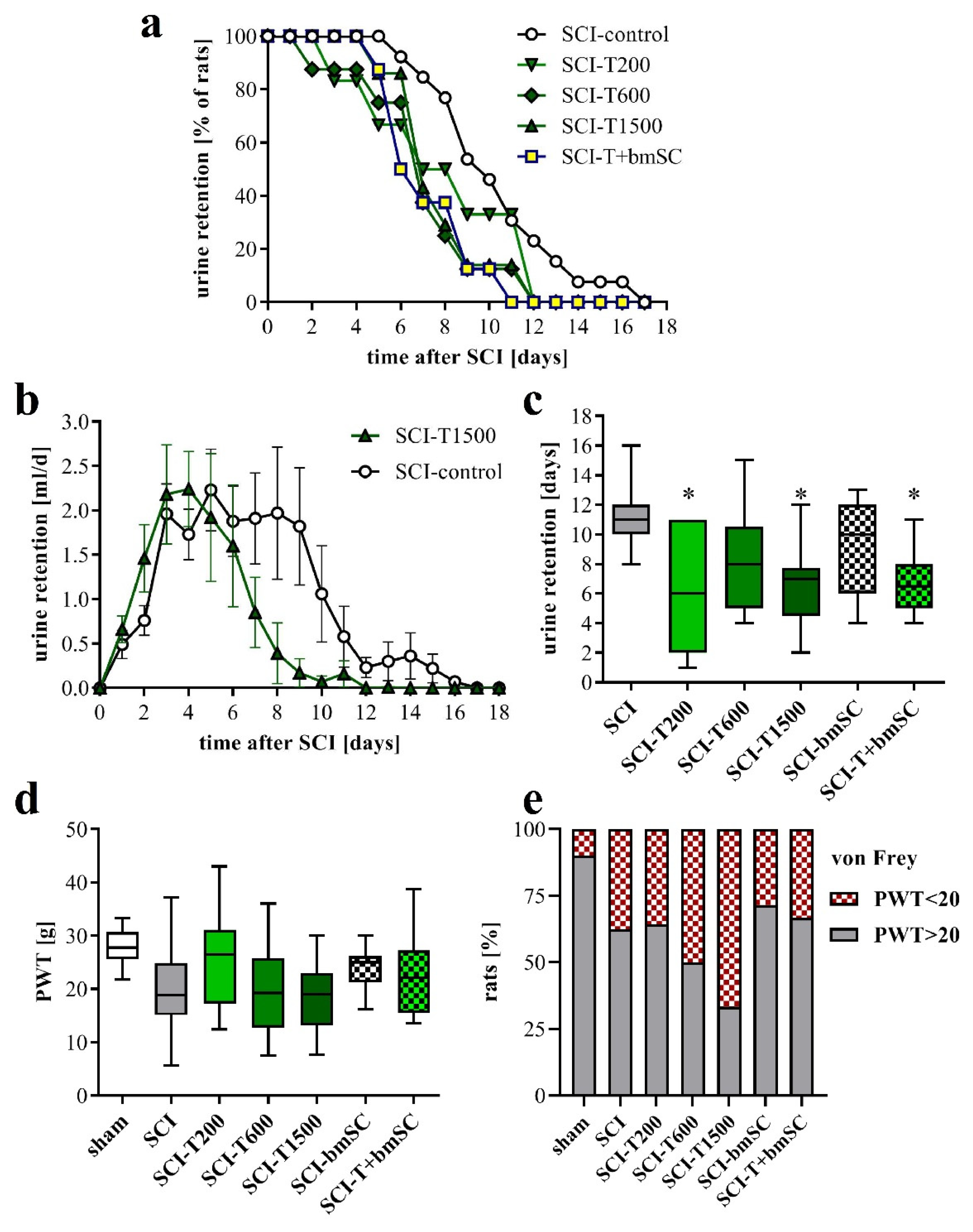
3.3. Faster Recovery of Bladder Control with TUDCA Treatment
3.4. Effect of TUDCA and bmSC Treatment on Allodynia/Hyperalgesia
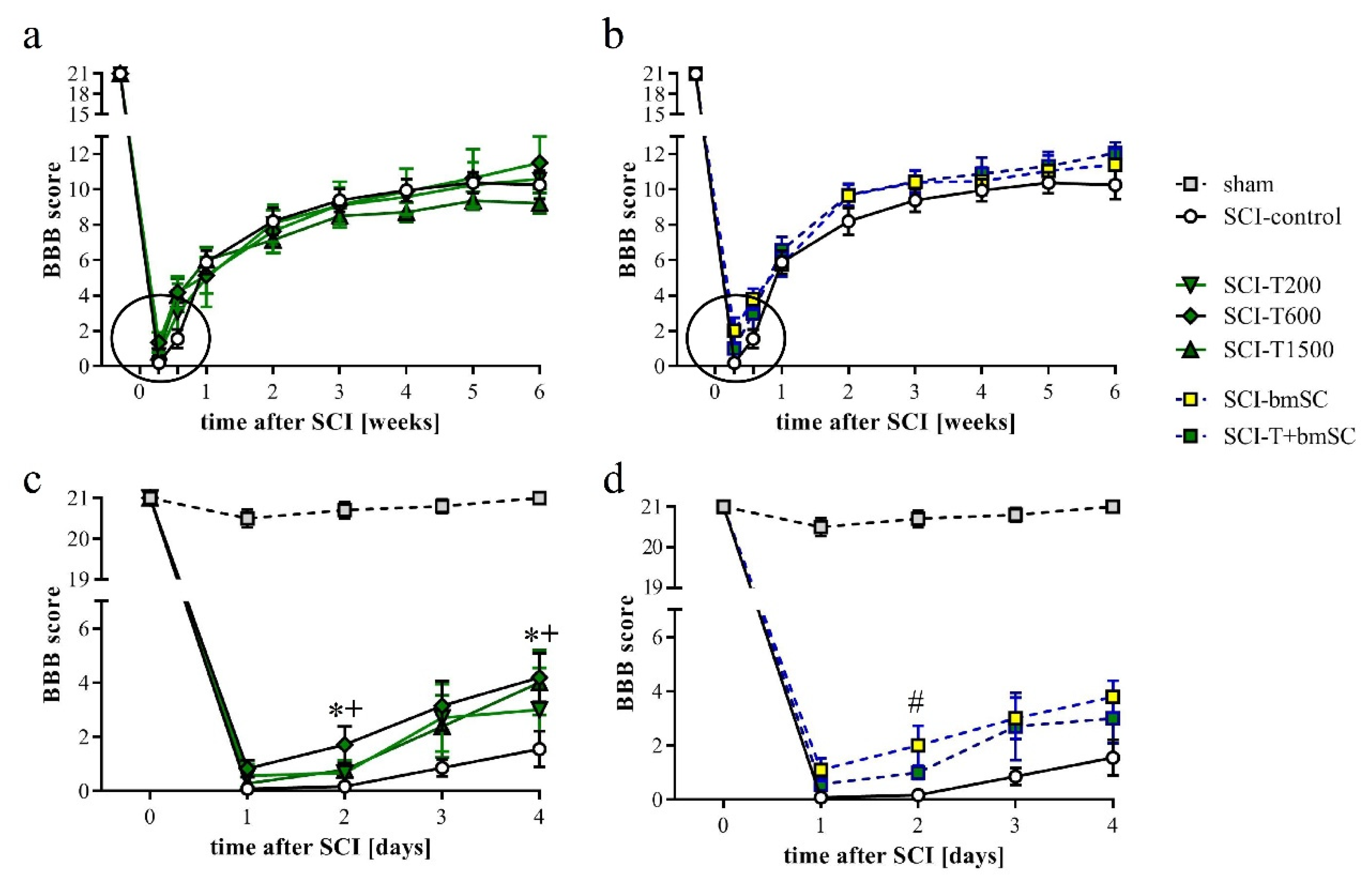
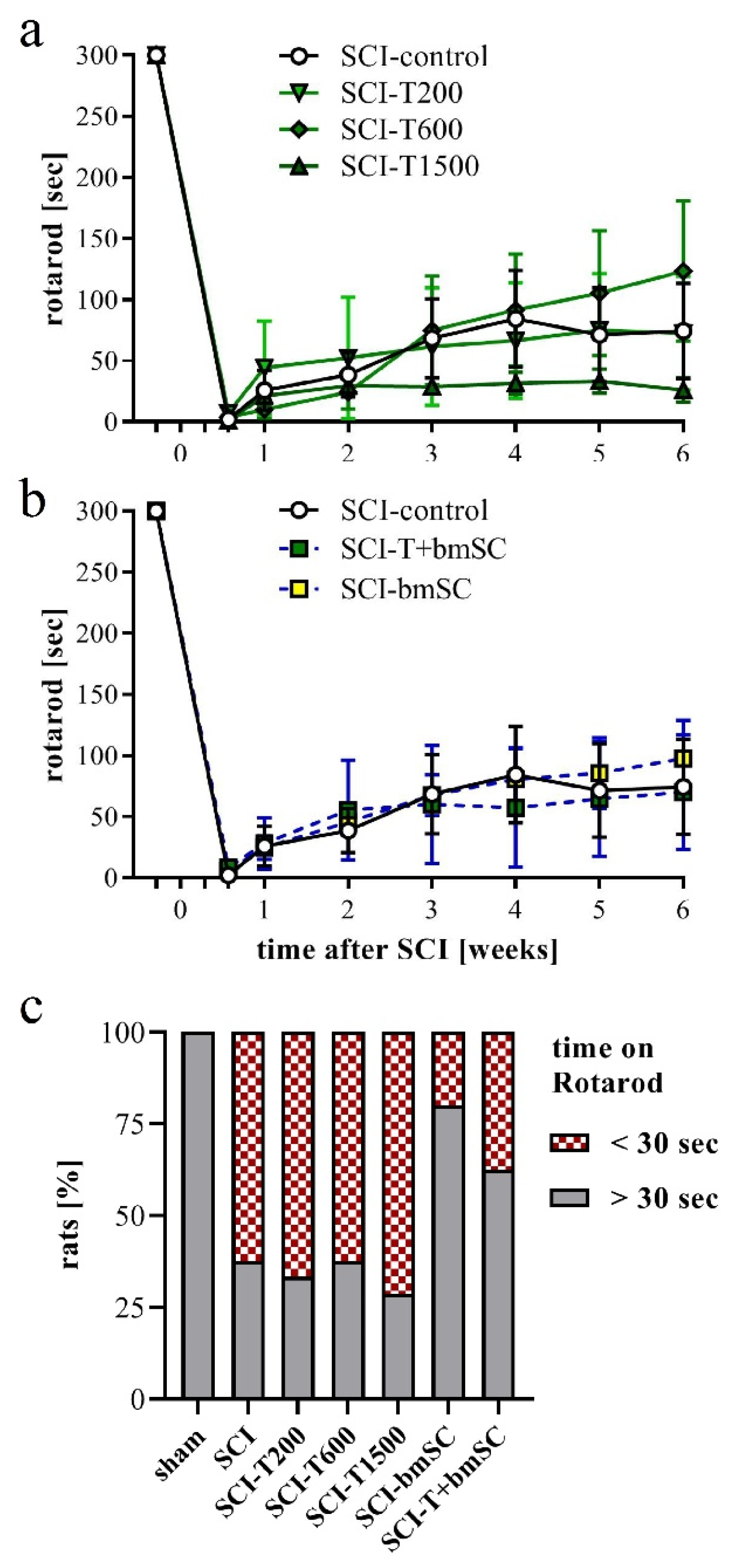
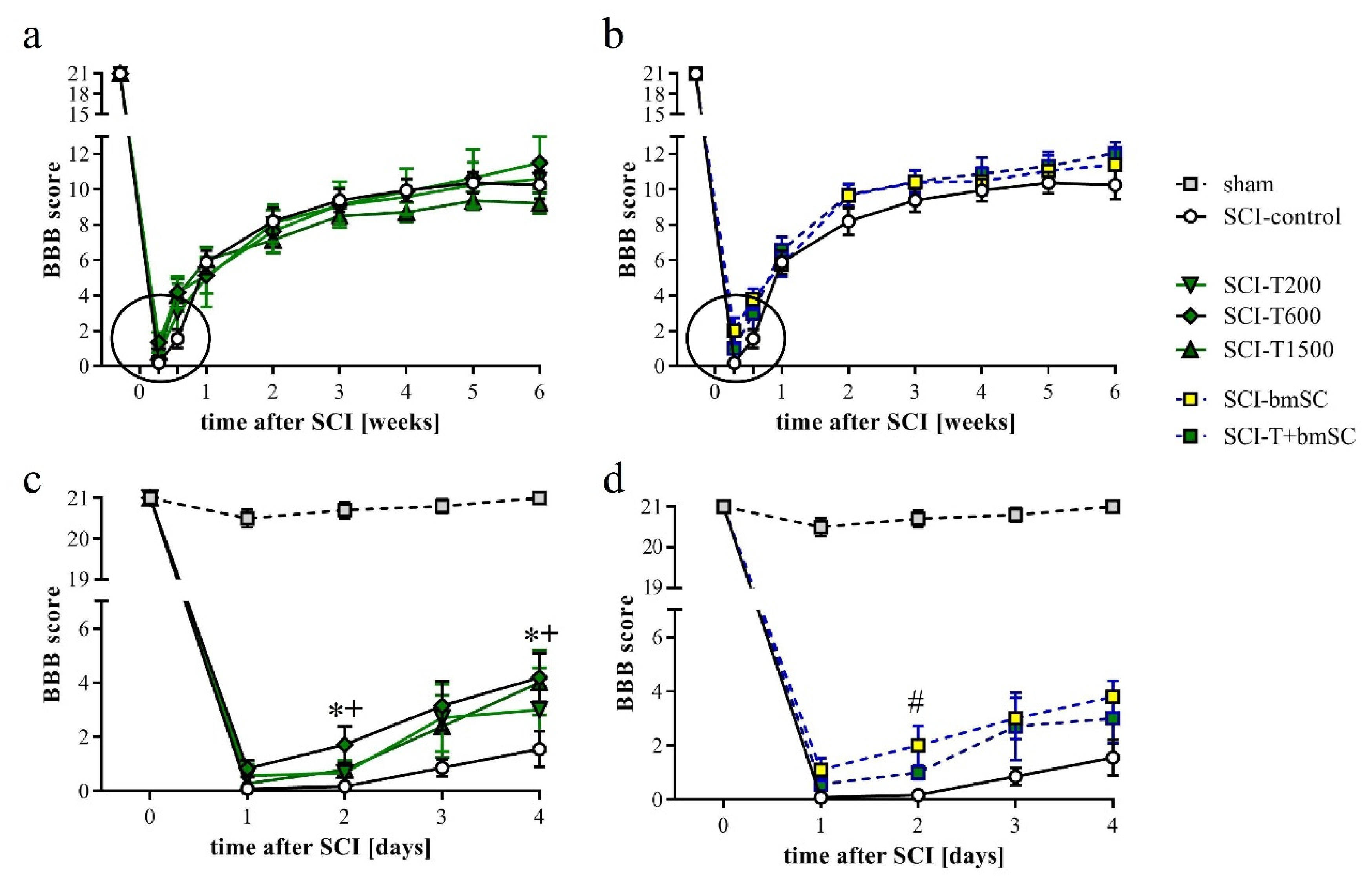
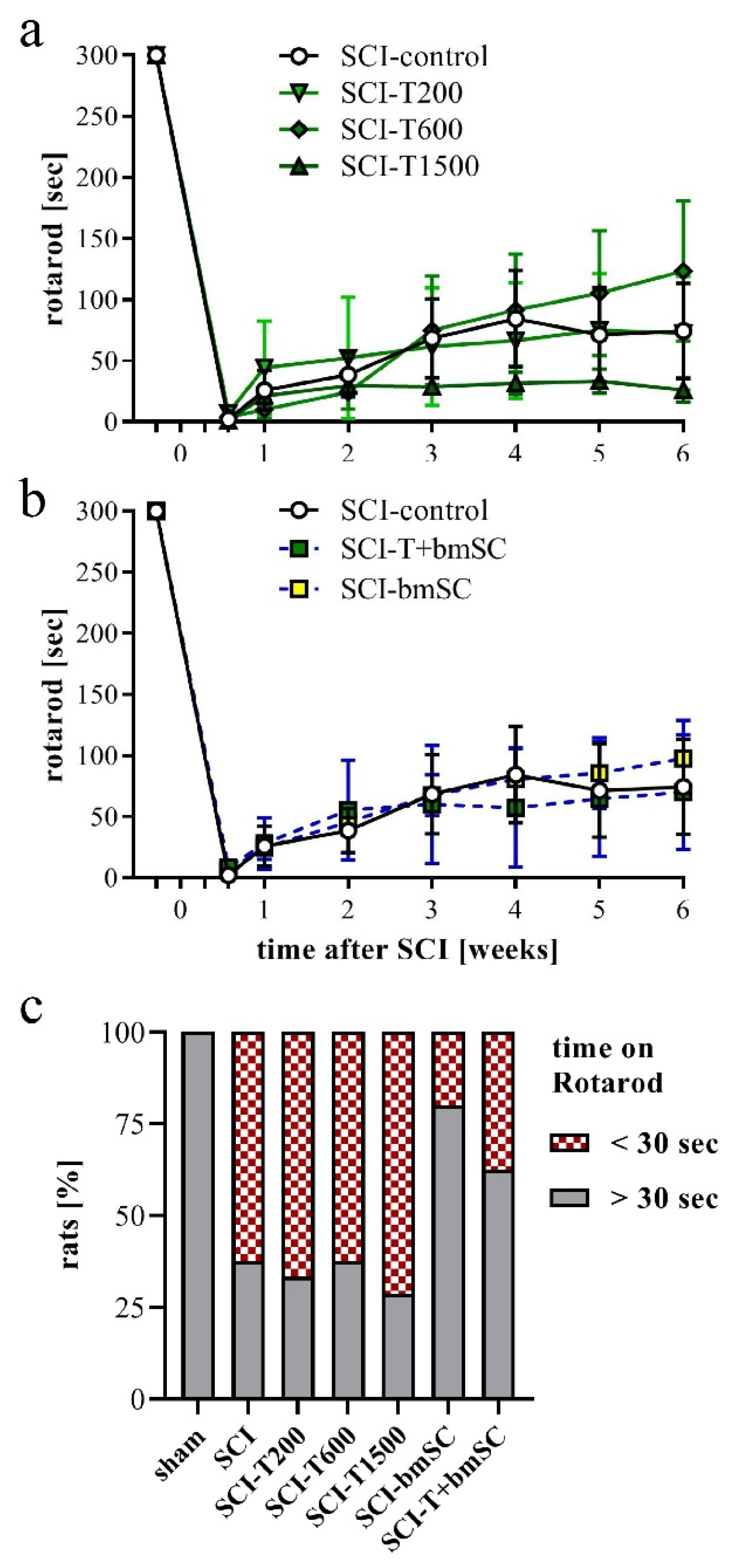
3.5. Effect of TUDCA and bmSC Treatment on Recovery of Sensory-Motor Functions
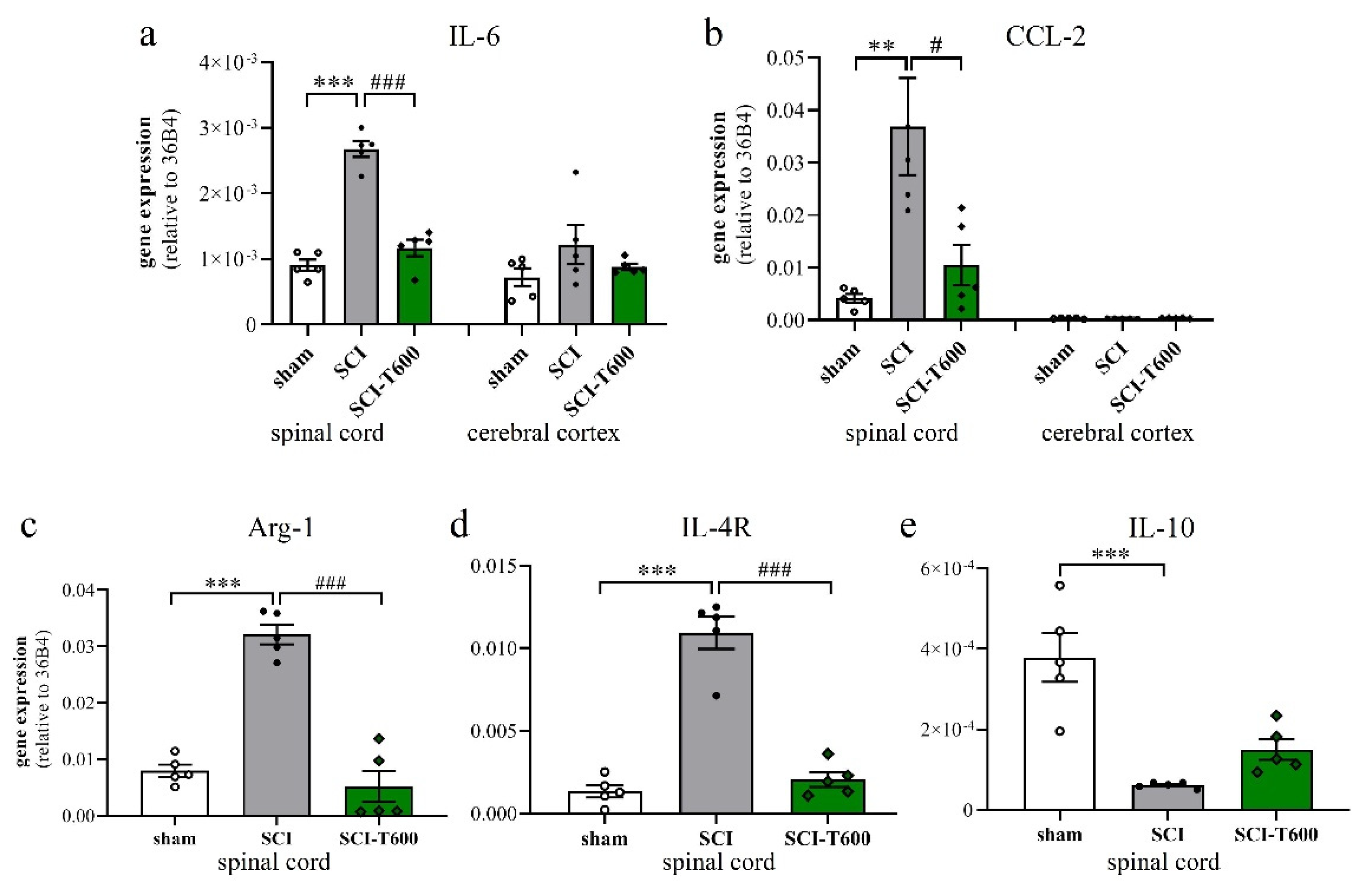
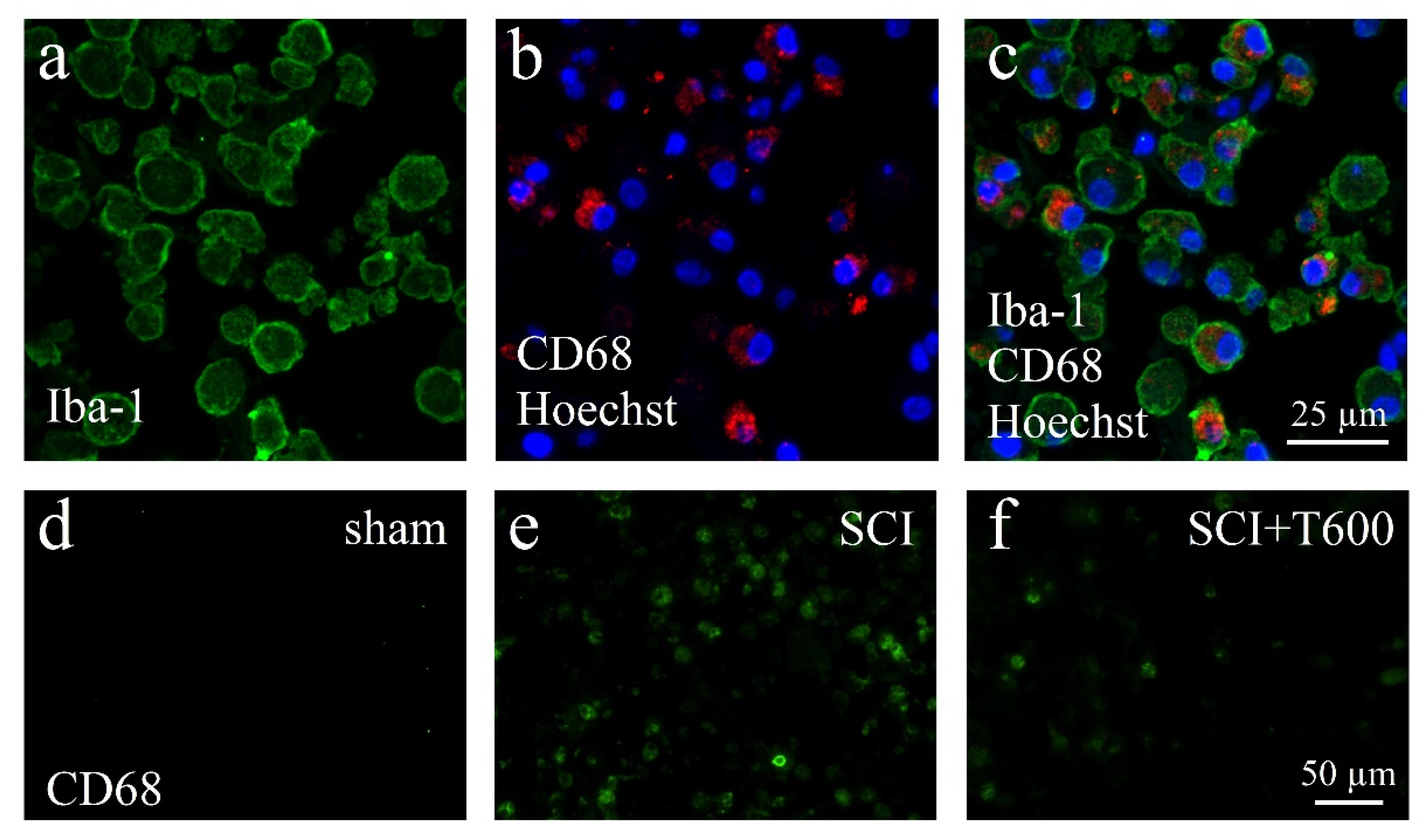
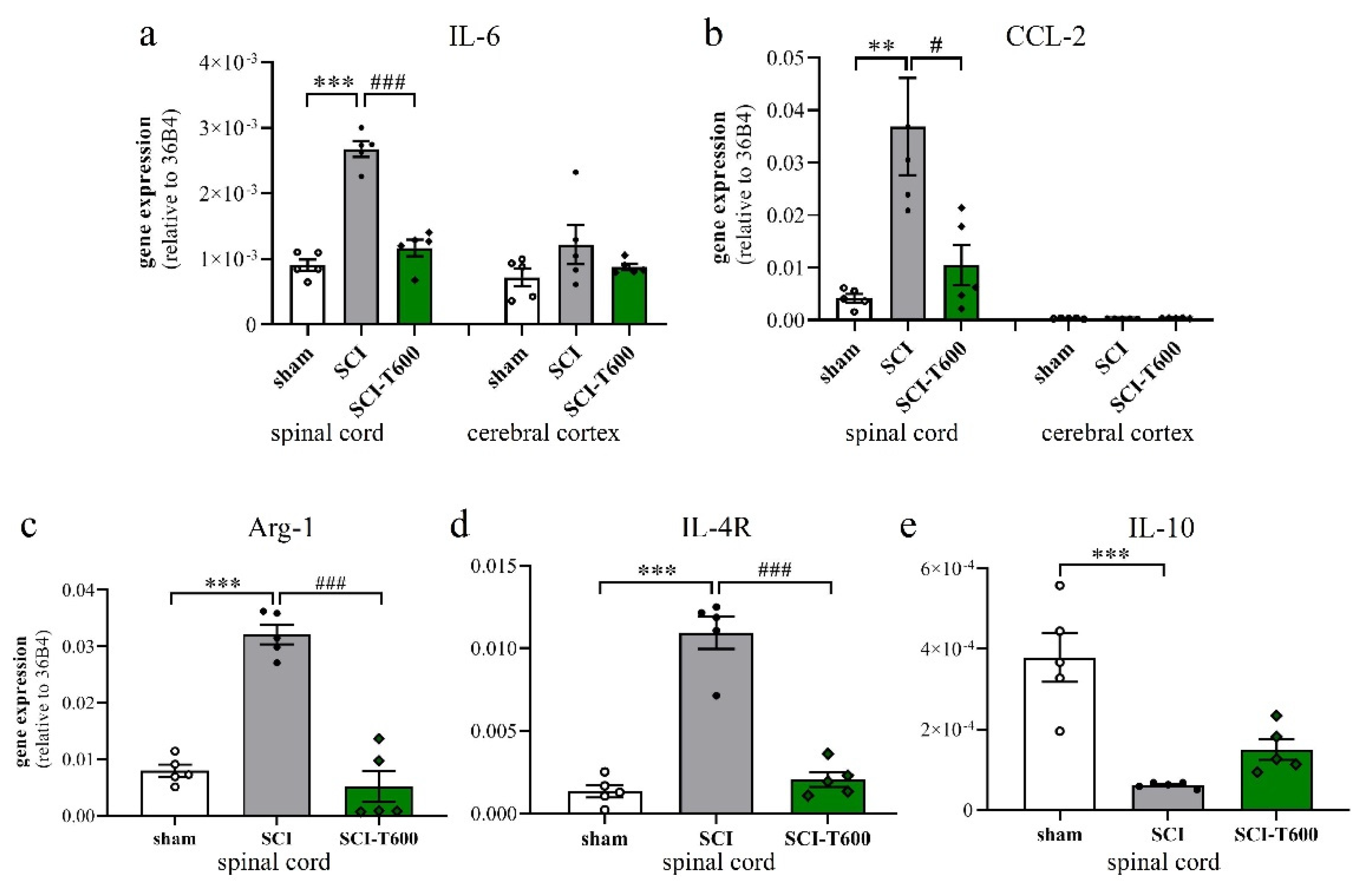
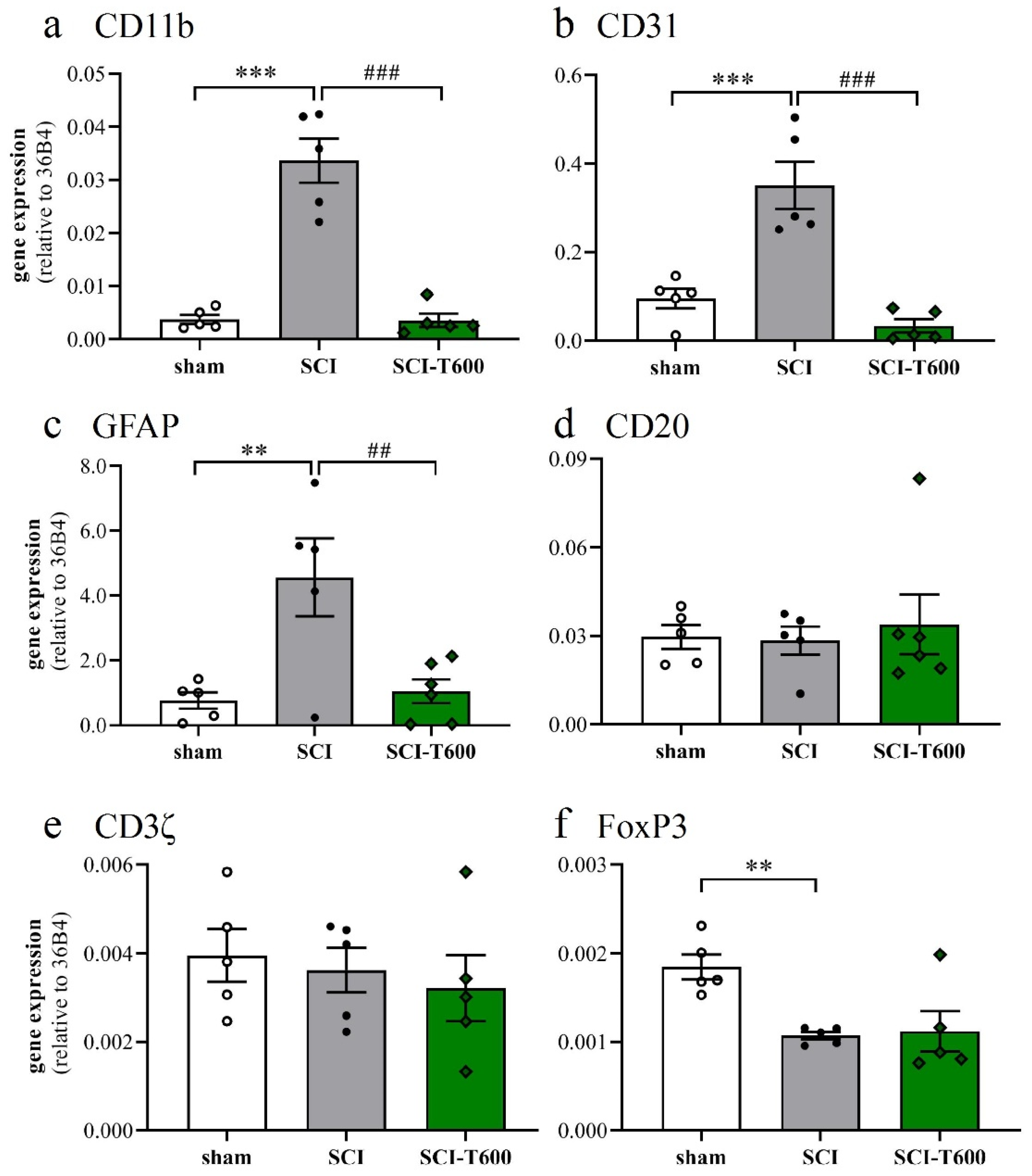
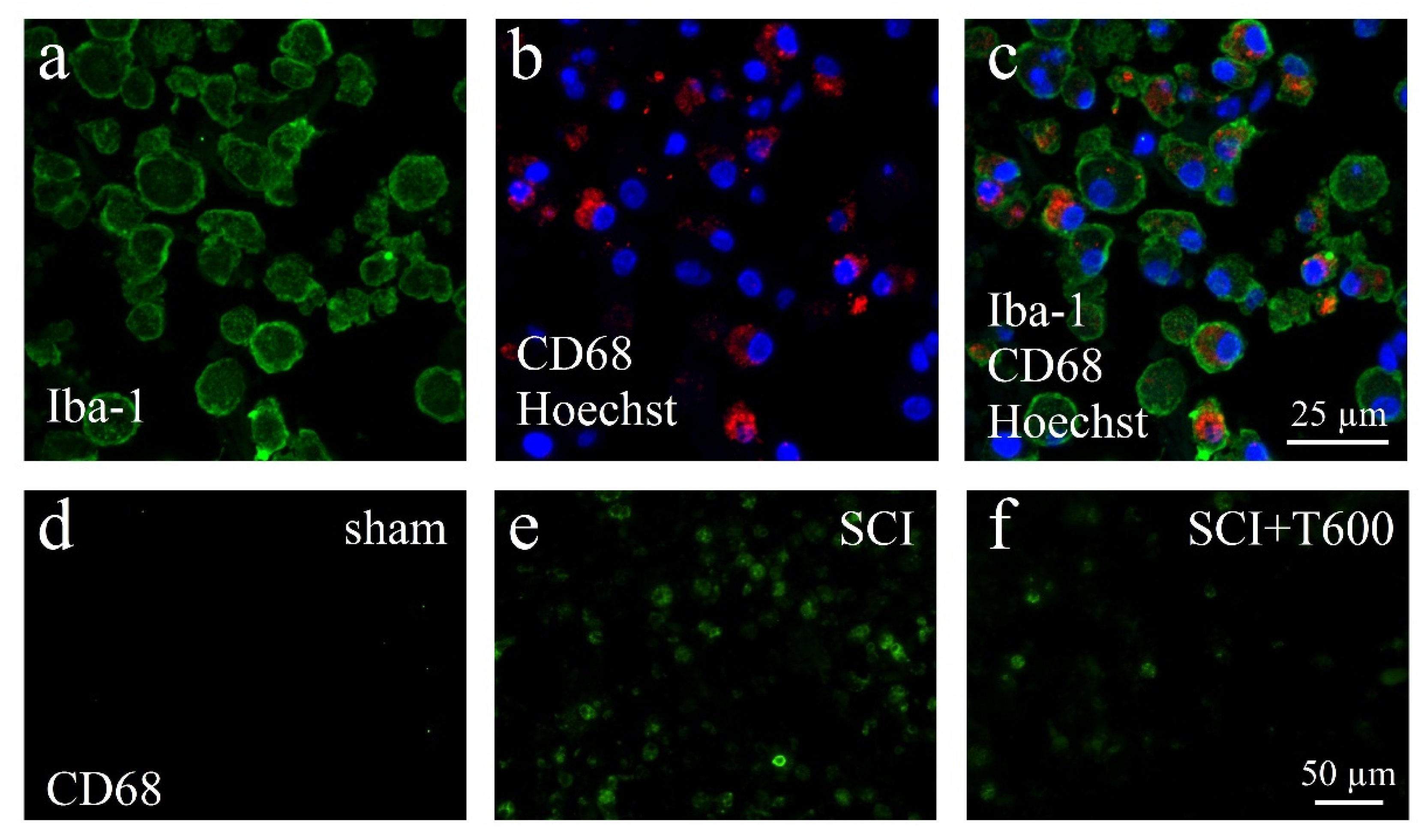
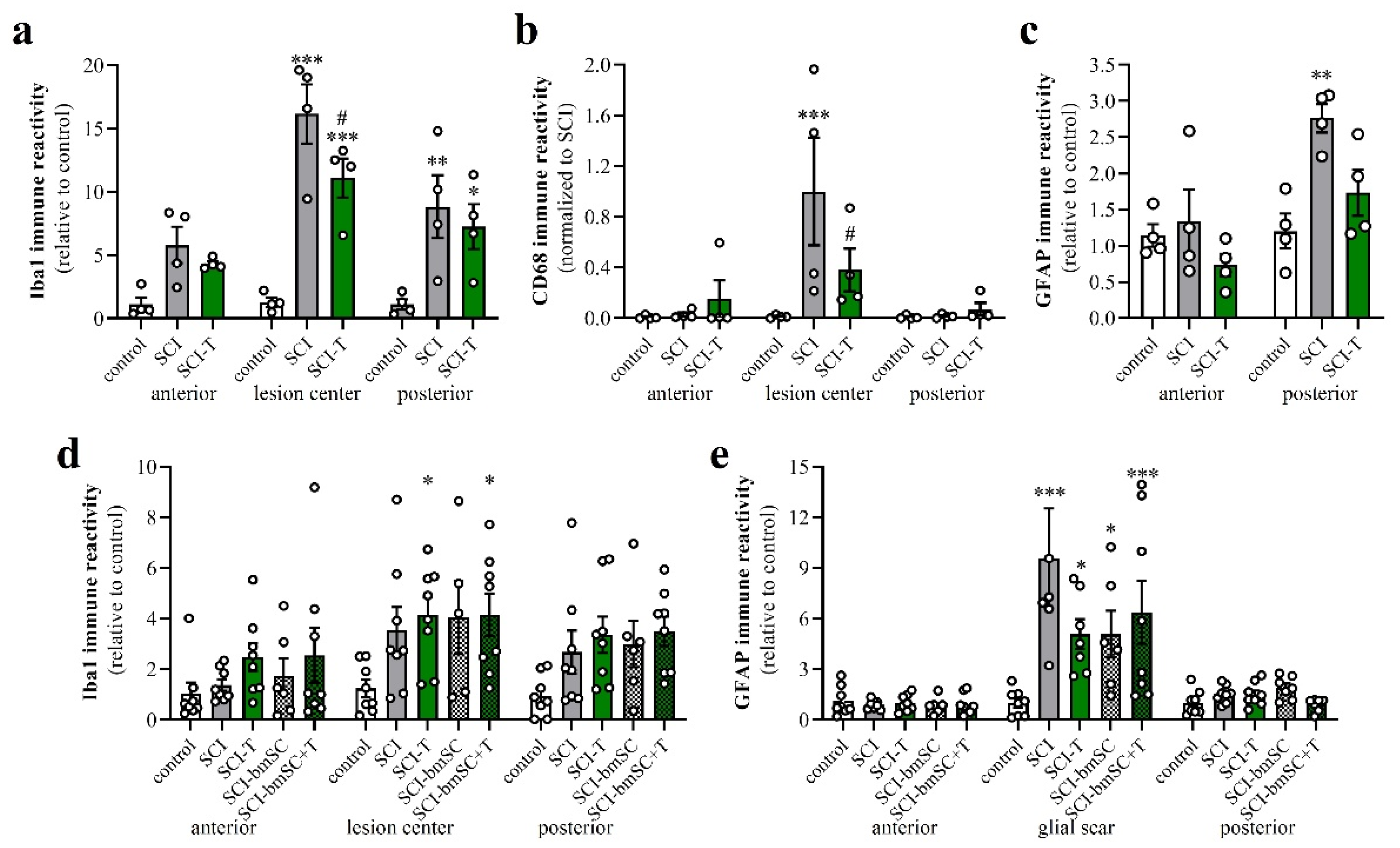
3.6. Effect of TUDCA Treatment on Neuroinflammation
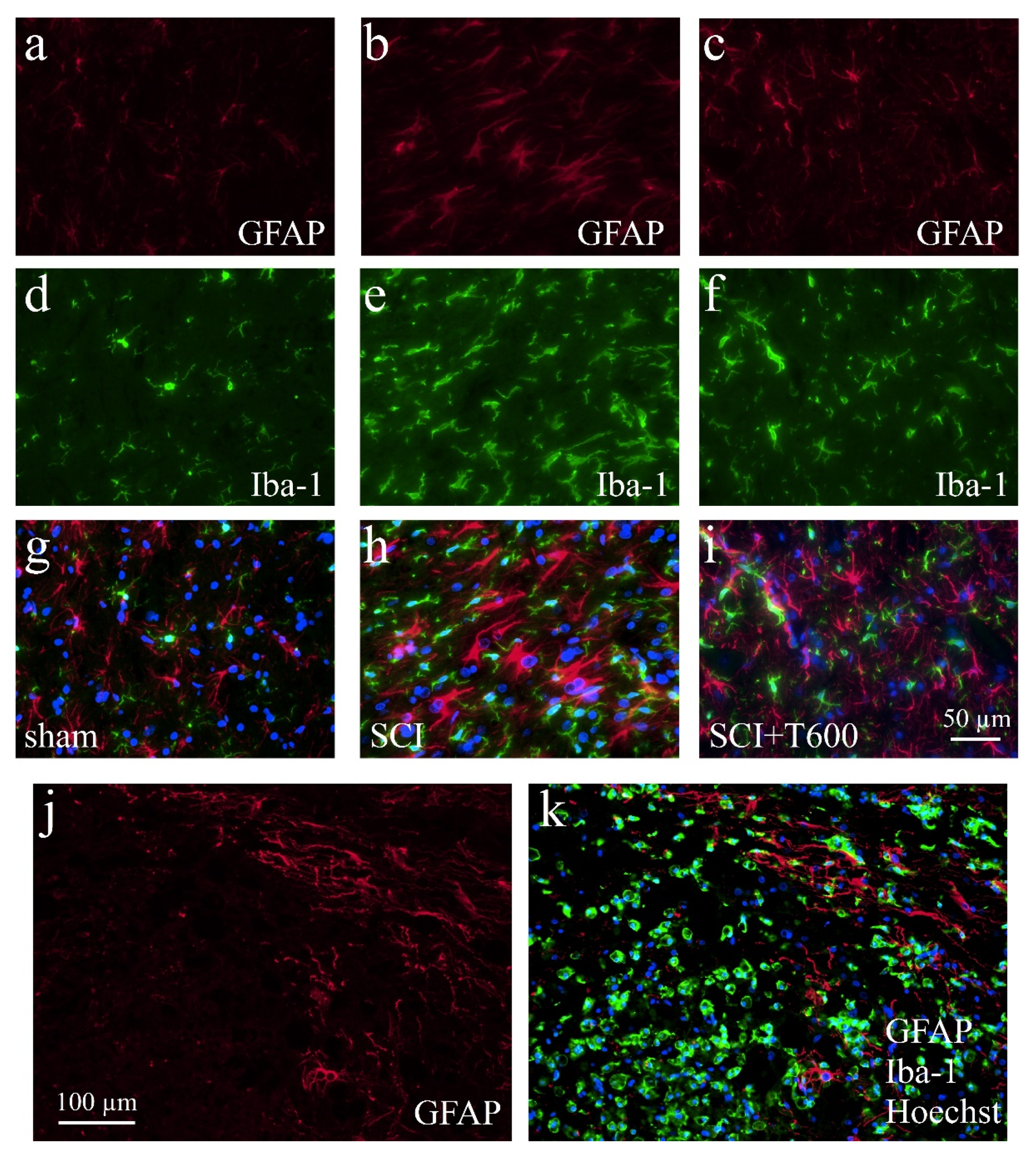
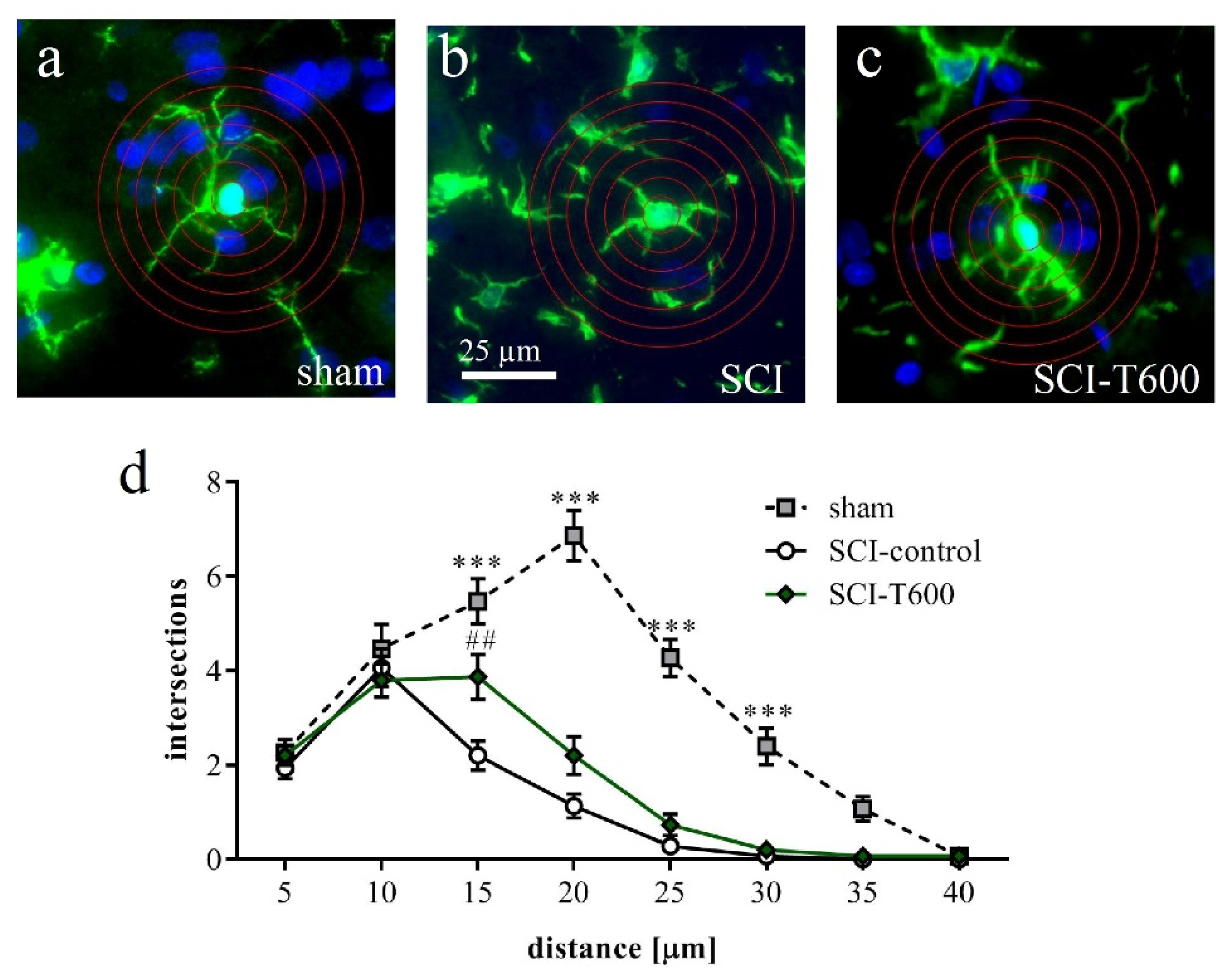
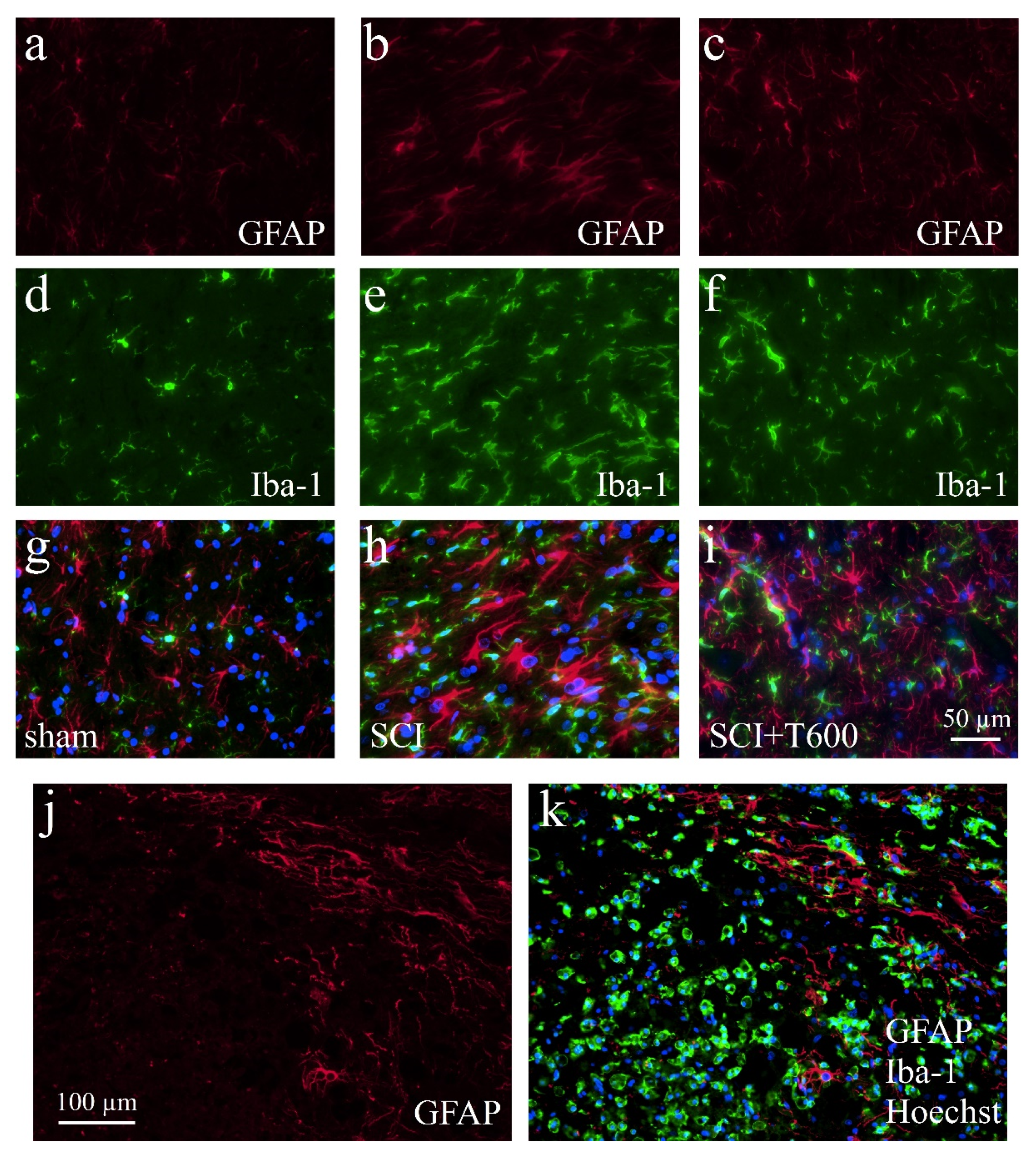
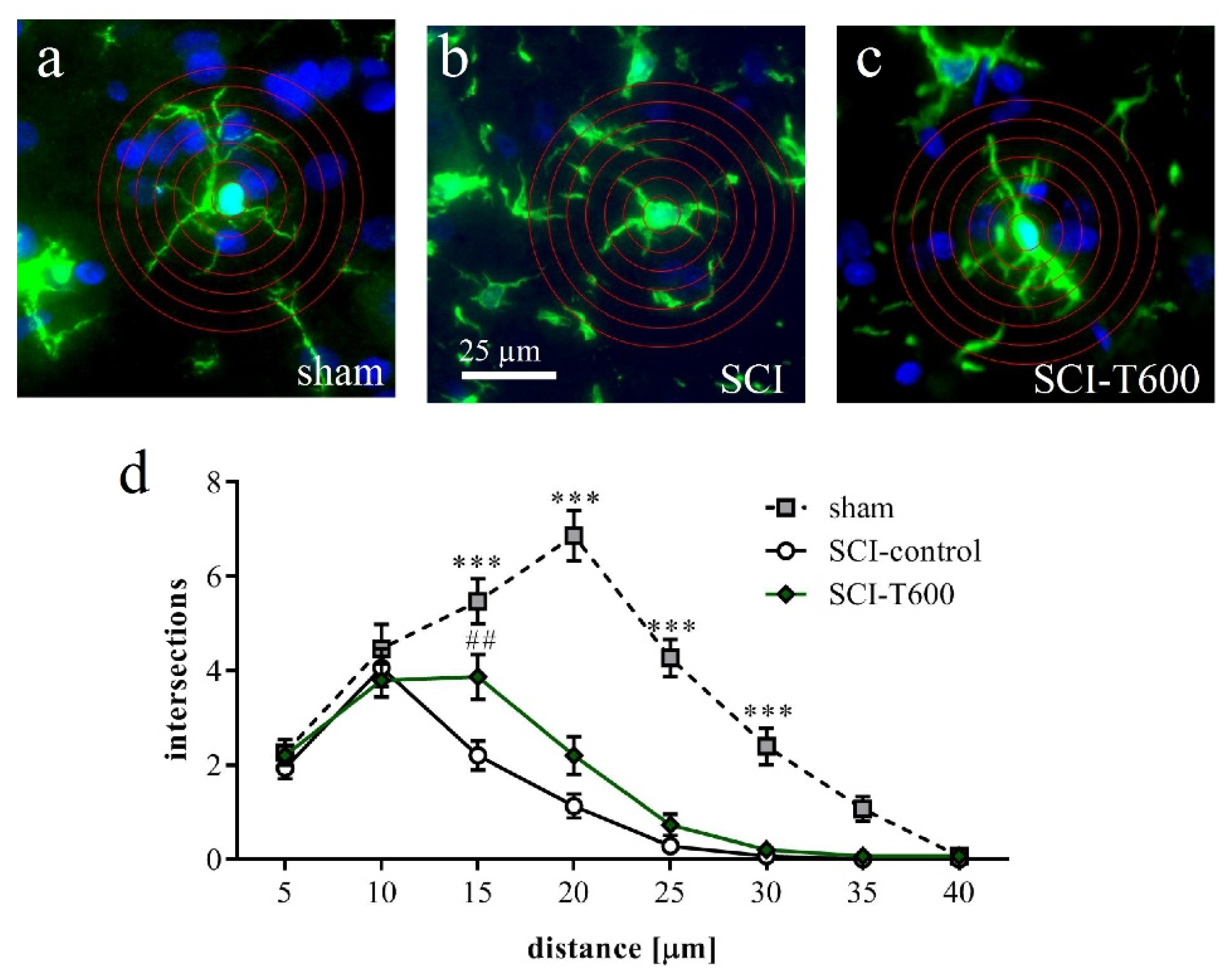
3.7. Effect of TUDCA Treatment on Glial Activation after SCI
3.8. Effect of TUDCA Treatment on SCI-Induced Apoptosis
4. Discussion
4.1. Are Bile Acids Promising for the Treatment of SCI?
4.2. How Does TUDCA Influence the Recovery of Urinary Function?
4.3. How Does TUDCA Affect the Inflammatory Phenotype after SCI?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 36B4 | acidic ribosomal phosphoprotein P0 |
| Akt | serine/threonine protein kinase encoded by the oncogene in retrovirus isolated from stock A strain k mouse thymoma cell line (protein kinase B) |
| ANOVA | analysis of variance |
| BBB | Basso/Beattie/Bresnahan locomotor rating scale for the assessment of hind limb motor function in rats |
| bmSC | bone marrow-derived stromal cells |
| CCL | chemokine C-C motif ligand |
| CD | cluster of differentiation |
| DMSO | dimethyl sulfoxide |
| dpo | days post operation |
| EDTA | ehylenediaminetetraacetic acid |
| GFAP | glial fibrillary acidic protein |
| GMP | good manufacturing practice |
| Iba-1 | ionized calcium-binding adapter molecule 1 |
| IFN | interferon |
| IF | immunofluorescence |
| IgG | immunoglobulin G |
| IH | Infinite Horizon spinal cord impactor |
| IL | interleukin |
| i.p. | intraperitoneal |
| IR | immunoreactivity |
| LPS | lipopolysaccharide |
| NFκB | nuclear factor kappa B |
| PBS | phosphate buffered saline |
| PCR | polymerase chain reaction |
| PFA | paraformaldehyde |
| PI3K | phosphoinositide 3-kinase |
| PKA | serine-threonine protein kinase A family |
| PWT | paw withdrawal threshold |
| ROI | region of interest |
| RT | room temperature |
| s.c. | subcutaneous |
| SCI | spinal cord injury |
| SEM | standard error of the mean |
| T200, T600, T1500 | treatment with TUDCA 2 × 100 mg/kg, 2 × 300 mg/k and 5 × 300 mg/kg respectively in the present study |
| TBS-T | Tris-buffered saline/Tween 20 |
| TGF | transforming growth factor |
| TGR5 | Takeda G protein-coupled receptor-5 |
| TNF | tumor necrosis factor |
| TRITC | tetramethyl rhodamine iso-thiocyanate |
| TUDCA | tauroursodeoxycholic acid |
| TUNEL | terminal deoxynucleotidyl transferase dUTP nick end labeling |
| W | weeks after SCI |
References
- Donnelly, D.J.; Popovich, P.G. Inflammation and its role in neuroprotection, axonal regeneration and functional recovery after spinal cord injury. Exp. Neurol. 2008, 209, 378–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falavigna, A.; Quadros, F.W.; Teles, A.R.W.C.C.; Barbagallo, G.; Brodke, D.; Al Mutair, A.; Riew, K.D. Worldwide steroid prescription for acute spinal cord injury. Glob. Spine J. 2018, 8, 303–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hugenholtz, H.; Gass, D.E.; Dvorak, M.F.; Fewer, D.H.; Fox, R.J.; Izukawa, D.M.; Lexchin, J.; Tuli, S.; Bharatwal, N.; Short, C. High-dose methylprednisolone for acute closed spinal cord injury--only a treatment option. Can. J. Neurol. Sci. 2002, 29, 227–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tetzlaff, W.; Okon, E.B.; Karimi-Abdolrezaee, S.; Hill, C.E.; Sparling, J.S.; Plemel, J.R.; Plunet, W.T.; Tsai, E.C.; Baptiste, D.; Smithson, L.J.; et al. A systematic review of cellular transplantation therapies for spinal cord injury. J. Neurotrauma 2011, 28, 1611–1682. [Google Scholar] [CrossRef] [PubMed]
- Shende, P.; Subedi, M. Pathophysiology, mechanisms and applications of mesenchymal stem cells for the treatment of spinal cord injury. Biomed. Pharmacother. 2017, 91, 693–706. [Google Scholar] [CrossRef]
- Tsintou, M.; Dalamagkas, K.; Makris, N. Taking central nervous system regenerative therapies to the clinic: Curing rodents versus nonhuman primates versus humans. Neural Regen. Res. 2019, 15, 425–437. [Google Scholar]
- Munter, J.D.; Beugels, J.; Munter, S.; Jansen, L.; Cillero-Pastor, B.; Moskvin, O.; Brook, G.; Pavlov, D.; Strekalova, T.; Kramer, B.W.; et al. Standardized human bone-marrow-derive stem cells infusion improves survival and rocovery in a rat model of spinal cord injury. J. Neurolog. Sci. 2019, 402, 16–29. [Google Scholar] [CrossRef]
- Romero-Ramírez, L.; Wu, S.; de Munter, J.; Wolters, E.C.; Kramer, B.W.; Mey, J. Treatment of rats with spinal cord injury using human bone marrow-derived stromal cells prepared by negative selection. J. Biomed. Sci. 2020, 27, 35. [Google Scholar] [CrossRef] [Green Version]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A sensitive and reliable locomotor rating scale for open field testing in rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef]
- Feng, Y.; Siu, K.; Wang, N.; Ng, K.-M.; Tsao, S.-W.; Nagamatsu, T.; Tong, Y. Bear bile: Dilemma of traditional medicinal use and animal protection. J. Ethnobiol. Ethnomed. 2009, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Grant, S.; DeMorrow, S. Bile acid signaling in neurodegenerative and neurological disorders. Int. J. Mol. Sci. 2020, 21, 5982. [Google Scholar] [CrossRef] [PubMed]
- Keitel, V.; Gorg, B.; Bidmon, H.J.; Zemtsova, I.; Spomer, L.; Zilles, K.; Haussinger, D. The bile acid receptor TGR5 (Gpbar-1) acts as a neurosteroid receptor in brain. Glia 2010, 58, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Eggink, H.M.; Soester, M.R.; Pols, T.W.H. TGR5 ligands as potential therapeutics in inflammatory diseases. Int. J. Interferon Cytokine Mediat. Res. 2014, 6, 27–38. [Google Scholar]
- Pols, T.W.H.; Noriega, L.G.; Nomura, M.; Auwerx, J.; Schoonjans, K. The bile acid membrane receptor TGR5: A valuable metabolic target. Dig. Dis. 2011, 29, 37–44. [Google Scholar] [CrossRef] [Green Version]
- De Marino, S.; Carino, A.; Masullo, D.; Finamore, C.; Marchianò, S.; Cipriani, S.; Di Leva, F.S.; Catalanotti, B.; Novellino, E.; Limongelli, V.; et al. Hyodeoycholic acid derivatives as liver X receptor α and G-protein-coupled bile acid receptor agonists. Sci. Rep. 2017, 7, 43290. [Google Scholar] [CrossRef] [Green Version]
- Chiang, J.Y.L. Bile acid metabolism and signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar]
- Yanguas-Casás, N.; Barreda-Manso, M.A.; Nieto-Sampedro, M.; Romero-Ramírez, L. TUDCA: An agonist of the bile acid receptor GPBAR1/TGR5 with anti-inflammatory effects in microglial cells. J. Cell Physiol. 2017, 232, 2231–2245. [Google Scholar] [CrossRef]
- Romero-Ramírez, L.; García-Rama, C.; Wu, S.; Mey, J. Bile acids attenuate PKM2 pathway activation in proinflammatory microglia. Sci. Rep. 2022, 12, 1459. [Google Scholar] [CrossRef]
- Rosa, A.I.; Duarte-Silva, S.; Silva-Fernandes, A.; Nunes, M.J.; Carvalho, A.N.; Rodrigues, E.; Gama, M.J.; Rodrigues, C.M.P.; Marciel, P.; Castro-Caldas, M. Tauroursodeoxycholic acid improves motor symptoms in a mouse model of Parkinson’s disease. Mol. Neurobiol. 2018, 55, 9139–9155. [Google Scholar] [CrossRef]
- Bhargava, P.; Smith, M.D.; Mische, L.; Harrington, E.; Fitzgerald, K.C.; Martin, K.; Kim, S.; Reyes, A.A.; González-Cardona, J.; Volsko, C.; et al. Bile acid metabolism is altered in multiple sclerosis and supplementation ameliorates neuroinflammation. J. Clin. Investig. 2020, 130, 3467–3482. [Google Scholar] [CrossRef] [Green Version]
- Elia, A.E.; Lalli, S.; Monsurro, M.R.; Sagnelli, A.; Taiello, A.C.; Reggiori, B.; La, B.V.; Tedeschi, G.; Albanese, A. Tauroursodeoxycholic acid in the treatment of patients with amyotrophic lateral sclerosis. Eur. J. Neurol. 2016, 23, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Çolak, A.; Kelten, B.; Sagmanligil, A.; Akdemir, O.; Karaoglan, A.; Sahan, E.; Çelik, Ö.; Barut, S. Tauroursodeoxycholic acid and secondary damage after spinal cord injury in rats. J. Clin. Neurosci. 2008, 15, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Miao, L.; Lin, L.; Ding, H. Neuroprotective effects impact on caspase-12 expression of tauroursodeoxycholic acid after acute spinal cord injury in rats. Int. J. Clin. Exp. Pathol. 2015, 8, 15871–15878. [Google Scholar] [PubMed]
- Miao, L.; Dong, L.; Zhou, F.-B.; Chang, Y.-L.; Suo, Z.G.; Ding, H.Q. Protective effect of tauroursodeoxycholic acid on the aotophagy of nerve cells in rats with acute spinal cord injury. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1133–1141. [Google Scholar]
- Zhang, Z.; Chen, J.; Chen, F.; Yu, D.; Li, R.; Lv, C.; Wang, H.; Li, H.; Li, J.; Cai, Y. Tauroursodeoxycholic acid alleviates secondary injury in the spinal cord via up-regulation of CIBZ gene. Cell Stress Chaperones 2018, 23, 560. [Google Scholar] [CrossRef]
- Dong, Y.; Yang, S.; Fu, B.; Zhou, S.; Ding, H.; Ma, W. Mechanism of tauroursodeoxycholic acid-mediated neuronal protection after acute spinal cord injury through AKT signaling pathway in rats. Int. J. Clin. Exp. Pathol. 2020, 13, 2218–2227. [Google Scholar]
- Mann, A.; Chesselet, M.-F. Techniques in motor assessment in rodents. In Movement Disorders—Genetics and Models; LeDouy, M.S., Ed.; Elsevier: London, UK, 2015. [Google Scholar]
- Pikov, V.; Wrathall, J.R. Coordination of the bladder detrusor and the external urethral sphincter in a rat model of spinal cord injury: Effects of injury severity. J. Neurosci. 2001, 21, 559–569. [Google Scholar] [CrossRef] [Green Version]
- Pols, T.W.H.; Nomura, M.; Harach, T.; Lo Sasso, G.; Ooosterveer, M.H.; Thomas, C.; Rizzo, G.; Gioiello, A.; Adorini, L.; Pellicari, R.; et al. TGR5 activation inhibits atherosclerosis by reducing macrophage inflammation and lipid loading. Cell Metab. 2011, 14, 747–757. [Google Scholar] [CrossRef] [Green Version]
- Yanguas-Casás, N.; Barreda-Manso, M.A.; Nieto-Sampedro, M.; Romero-Ramírez, L. Tauroursodeoxycholic acid reduces glial cell activation in an animal model of acute neuroinflammation. J. Neuroinflamm. 2014, 11, 50. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Ko, W.K.; Jo, M.J.; Arai, Y.; Choi, H.; Kumar, H.; Han, I.B.; Sohn, S. Anti-inflammatory effect of Tauroursodeoxycholic acid in RAW 264.7 macrophages, Bone marrow-derived macrophages, BV2 microglial cells, and spinal cord injury. Sci. Rep. 2018, 8, 3176. [Google Scholar] [CrossRef]
- Hou, Y.; Luan, J.; Huang, T.; Deng, T.; Li, X.; Xiao, Z.; Zhan, J.; Luo, D.; Hou, Y.; Xu, L.; et al. Tauroursodeoxycholic acid alleviates secondary injury in spinal cord injury mice by reducing oxidative stress, apoptosis, and inflammatory response. J. Neuroinflamm. 2021, 18, 216. [Google Scholar] [CrossRef] [PubMed]
- Hayda, E.; Elden, H. Acute spinal cord injury: A review of pathophysiology and potential of non-steroidal ant-inflammatory drugs for pharmacological intervention. J. Chem. Neuroanat. 2018, 87, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Benevento, B.T.; Sipski, M.L. Neurogenic bladder, neurogenic bowel, and sexual dysfunction in people with spinal cord injury. Phys. Ther. 2002, 82, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Fowler, C.J.; Griffiths, D.; de Groat, W.C. The neuronal control of micturition. Nat. Rev. Neurosci. 2008, 9, 453–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsui, T.; Shumsky, J.S.; Lepor, A.C.; Murray, M.; Fisher, I. Transplantation of neuronal and glial restricted precursors into contused spinal cord improves bladder and motor functions, decreases thermal hypersensitivity and modifies intraspinal circuitry. J. Neurosci. 2005, 25, 9624–9636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redshaw, J.D.; Lenjerr, S.M.; Elliott, S.P.; Stoffel, J.T.; Rosenbluth, J.P.; Presson, A.P.; Myers, J.B. Protocol for a randomized clinical trial investigating early sacral nerve stimnulation as an adjunct to standard neurogenic bladder management following acute spinal cord injury. BMC Urol. 2018, 18, 72. [Google Scholar] [CrossRef]
- Ditunno, J.F.; Little, J.W.; Tessler, A.; Burns, A.S. Spinal shock revisited: A four-phase model. Spinal Cord 2004, 42, 383–395. [Google Scholar] [CrossRef] [Green Version]
- Yanguas-Casás, N.; Barreda-Manso, M.A.; Perez-Rial, S.; Nieto-Sampedro, M.; Romero-Ramírez, L. TGFbeta Contributes to the Anti-inflammatory Effects of Tauroursodeoxycholic Acid on an Animal Model of Acute Neuroinflammation. Mol. Neurobiol. 2017, 54, 6737–6749. [Google Scholar] [CrossRef]
- Castro, J.; Harrington, A.M.; Lieu, T.; García-Caraballo, S.; Maddern, J.; Schober, G.; O’Donnell, T.; Grundy, L.; Lumsden, A.L.; Miller, P.; et al. Activation of pruritogenic TGR5, MRGPRA3, and MRGPRC11 on colon-innervating afferents induces visceral hypersensitivity. JCI Insight 2019, 4, e131712. [Google Scholar] [CrossRef]
- Freilich, R.W.; Woodbury, M.E.; Ikezu, T. Integrated expression profiles of mRNA and miRNA in polarized primary murine microglia. PLoS ONE 2013, 8, e79416. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, T.; Lively, S.; Schlichter, L.C. Complex molecular and functional outcomes of single versus sequential cytokine stimulation of rat microglia. J. Neuroinflamm. 2016, 13, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, M.; Xu, Y.; Pearse, D.D. Cyclic AMP is a key regulator of M1 to M2a phenotypic conversion of microglia in the presence of Th2 cytokines. J. Neuroinflamm. 2016, 13, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ransohoff, R.M. A polarizing question: Do M1 and M2 microglia exist? Nat. Neurosci. 2016, 19, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Gu, G.; Wang, J.; Chai, Y.; Fan, Y.; Yang, M.; Xu, X.; Gao, W.; Li, F.; Yin, D.; et al. Administration of taurourosdoxycholic acid attenuates early brain injury via Akt pathway activation. Front. Cell. Neurosci. 2017, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Chen, W.-D.; Wang, Y.-D. TGR5, not only a metabolic regulator. Front. Physiol. 2016, 7, 646. [Google Scholar] [CrossRef]
- Terui, R.; Yanase, Y.; Yokoo, H.; Suhara, Y.; Makishima, M.; Demizu, Y.; Misawa, T. Development of selective TGR5 ligands based on the 5,6,7,7-tetrahydro-5,5,8,8-tetramethylnaphthalene skeleton. ChemMedChem 2020, 16, 458–462. [Google Scholar] [CrossRef]
- Wang, J.-L.; Ren, C.-H.; Feng, J.; Ou, C.-H.; Liu, L. Oleanolic acid inhibits mouse spinal cord injury through suppressing inflammation and apoptosis via the blockage of p38 and JNK MAPKs. Biomed. Pharmacother. 2020, 123, 109752. [Google Scholar] [CrossRef]
- Ackerman, H.D.; Gerhard, G.S. Bile disorders in neurodegenerative disorders. Front. Aging Neurosci. 2016, 8, 263. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Chen, J.; Hollister, K.; Sowers, L.C.; Forman, B.M. Endogenous bile acids are ligands for the nuclear receptor FXR/BAR. Mol. Cell 1999, 3, 543–553. [Google Scholar] [CrossRef]
- Mey, J. A new target for CNS injury? The role of retinoic acid signaling after nerve lesions. J. Neurobiol. 2006, 66, 757–779. [Google Scholar] [CrossRef]
- Van Neerven, S.; Kampmann, E.; Mey, J. RAR/RXR and PPAR/RXR signaling in neurological and psychiatric diseases. Prog. Neurobiol. 2008, 85, 433–451. [Google Scholar] [CrossRef] [PubMed]
- Paterniti, I.; Genovese, T.; Mazzon, E.; Crisafulli, C.; Di Paola, R.; Galuppo, M.; Bramanti, P.; Cuzzocrea, S. Liver X receptor agonist treatment regulates inflammatory response after spinal cord trauma. J. Neurochem. 2010, 112, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Fandel, D.; Wasmuht, D.; Ávila-Martín, G.; Taylor, M.D.; Galán-Arriero, I.; Mey, J. Spinal cord injury-induced changes of nuclear receptors PPARalpha and LXRbeta and modulation with oleic acid/albumin treatment. Brain Res. 2013, 1535, 89–105. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene ID (NCBI Reference) | Primer Sequences | Product Tm [°C] | Product Size [bp] |
|---|---|---|---|---|
| 36B4 | AC130745.3 | sense: TTCCCACTGGCTGAAAAGGT antisense: CGCAGCCGCAAATGC | 60 | 60 |
| IL-6 | NM_012589 | sense: TAGTCCTTCCTACCCCAATTTCC antisense: TTGGTCCTTAGCCACTCCTTC | 60 | 76 |
| CCL-2 | NM_031530 | sense: TGCTGTCTCAGCCAGATGCAGTTA antisense: TACAGCTTCTTTGGGACACCTGCT | 64 | 131 |
| Arg-1 | NM_017134 | sense: GCAGAGACCCAGAAGAATGGAAC antisense: CGGAGTGTTGATGTCAGTGTGAGC | 62 | 144 |
| IL-4Rα | NM_133380 | sense: GATCTTCTGAGCCCGGTTGA antisense: CTCTCCGCTTGCTGCATT | 59 | 59 |
| IL-10 | NM_012854 | sense: GATGCCCCAGGCAGAGAA antisense: CCCAGGGAATTCAAATGCT | 61 | 57 |
| CD11b | NM_012711.1 | sense: CTGCCTCAGGGATCCGTAAAG antisense: CCTCTGCCTCAGGAATGACATC | 60 | 150 |
| CD31 | NM_031591 | sense: GAGGTATCGAATGGGCAGAA antisense: GTGGAAGACCCGAGACTGAG | 55 | 174 |
| GFAP | NM_017009 | sense: TGGCCACCAGTAACATGCAA antisense: CAGTTGGCGGCGATAGTCAT | 60 | 134 |
| CD20 | NM_001107578 | sense: TCTTGGGCATTCTGTCGGTG antisense: TCTACAACACCGGCTGTCAC | 60 | 70 |
| CD3ζ | NM_170789 | sense: TCATACCCCAGCCCAGTTCT antisense: CGGCTCTGGGGACTTTACAA | 60 | 148 |
| FoxP3 | NM_001108250 | sense: TCATCACTGGCTTTCTGCGT antisense: GCTTTTAGCCTGAACCCCCT | 60 | 95 |
| TGR5 | NM_177936 | sense: AAAGGTGGCTACAAGTGCTTC antisense: TTCAAGTCCAAGTCAGTGCTG | 58 | 103 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; García-Rama, C.; Romero-Ramírez, L.; de Munter, J.P.J.M.; Wolters, E.C.; Kramer, B.W.; Mey, J. Tauroursodeoxycholic Acid Reduces Neuroinflammation but Does Not Support Long Term Functional Recovery of Rats with Spinal Cord Injury. Biomedicines 2022, 10, 1501. https://doi.org/10.3390/biomedicines10071501
Wu S, García-Rama C, Romero-Ramírez L, de Munter JPJM, Wolters EC, Kramer BW, Mey J. Tauroursodeoxycholic Acid Reduces Neuroinflammation but Does Not Support Long Term Functional Recovery of Rats with Spinal Cord Injury. Biomedicines. 2022; 10(7):1501. https://doi.org/10.3390/biomedicines10071501
Chicago/Turabian StyleWu, Siyu, Concepción García-Rama, Lorenzo Romero-Ramírez, Johannes P. J. M. de Munter, Erik Ch. Wolters, Boris W. Kramer, and Jörg Mey. 2022. "Tauroursodeoxycholic Acid Reduces Neuroinflammation but Does Not Support Long Term Functional Recovery of Rats with Spinal Cord Injury" Biomedicines 10, no. 7: 1501. https://doi.org/10.3390/biomedicines10071501
APA StyleWu, S., García-Rama, C., Romero-Ramírez, L., de Munter, J. P. J. M., Wolters, E. C., Kramer, B. W., & Mey, J. (2022). Tauroursodeoxycholic Acid Reduces Neuroinflammation but Does Not Support Long Term Functional Recovery of Rats with Spinal Cord Injury. Biomedicines, 10(7), 1501. https://doi.org/10.3390/biomedicines10071501