
Deletion of Wnt10a Is Implicated in Hippocampal Neurodegeneration in Mice

 , ,
, ,
Abstract
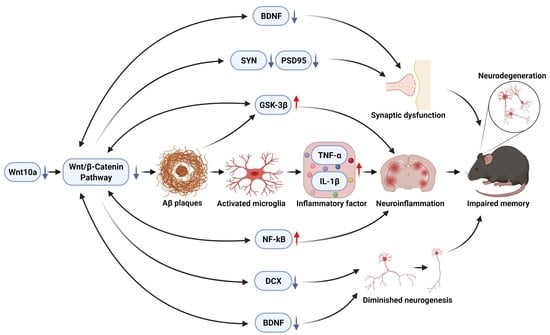
:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Neurobehavioral Tests
2.3. Histological Observation
2.4. Immunostaining
2.5. Transmission Electron Microscopy
2.6. Western Blot Analyses
2.7. Statistical Analyses
3. Results
3.1. Wnt10a-/- Mice were Smaller in Size and had Lower Hippocampal Volume
3.2. Wnt10a-/- Mice Showed Impaired Spatial Memory and Anxiety-like Behavior
3.3. Wnt10a-/- Mice Exhibited Lower Expression of β-Catenin in the Hippocampus
3.4. Wnt10a-/- Mice Displayed Lower Neurogenesis and BDNF Expression in the Hippocampus
3.5. Wnt10a-/- Mice Showed Synaptic Dysfunction in the Hippocampus
3.6. Wnt10a-/- Mice Exhibited Neuroinflammation in the Hippocampus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Knierim, J.J. The hippocampus. Curr. Biol. 2015, 25, R1116–R1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naslund, J.; Haroutunian, V.; Mohs, R.; Davis, K.L.; Davies, P.; Greengard, P.; Buxbaum, J.D. Correlation between elevated levels of amyloid beta-peptide in the brain and cognitive decline. JAMA 2000, 283, 1571–1577. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Zhou, Q.; Niwa, M.; Kubo, K.Y. Association between Mastication, the Hippocampus, and the HPA Axis: A Comprehensive Review. Int. J. Mol. Sci. 2017, 18, 1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, H.; Kurahashi, M.; Mori, D.; Iinuma, M.; Tamura, Y.; Mizutani, K.; Shimpo, K.; Sonoda, S.; Azuma, K.; Kubo, K.Y. Hippocampus-dependent spatial memory impairment due to molar tooth loss is ameliorated by an enriched environment. Arch. Oral Biol. 2016, 61, 1–7. [Google Scholar] [CrossRef]
- Varela-Nallar, L.; Inestrosa, N.C. Wnt signaling in the regulation of adult hippocampal neurogenesis. Front. Cell. Neurosci. 2013, 7, 100. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Wang, Z.; Hu, H.; Zhao, M.; Sun, L. Microglia in Alzheimer’s Disease: A Target for Therapeutic Intervention. Front. Cell. Neurosci. 2021, 15, 749587. [Google Scholar] [CrossRef]
- Aghaizu, N.D.; Jin, H.; Whiting, P.J. Dysregulated Wnt Signalling in the Alzheimer’s Brain. Brain Sci. 2020, 10, 902. [Google Scholar] [CrossRef]
- Moore, K.M.; Oelberg, W.L.; Glass, M.R.; Johnson, M.D.; Been, L.E.; Meisel, R.L. Glutamate Afferents From the Medial Prefrontal Cortex Mediate Nucleus Accumbens Activation by Female Sexual Behavior. Front. Behav. Neurosci. 2019, 13, 227. [Google Scholar] [CrossRef] [Green Version]
- Lie, D.C.; Colamarino, S.A.; Song, H.J.; Desire, L.; Mira, H.; Consiglio, A.; Lein, E.S.; Jessberger, S.; Lansford, H.; Dearie, A.R.; et al. Wnt signalling regulates adult hippocampal neurogenesis. Nature 2005, 437, 1370–1375. [Google Scholar] [CrossRef]
- Galli, S.; Lopes, D.M.; Ammari, R.; Kopra, J.; Millar, S.E.; Gibb, A.; Salinas, P.C. Deficient Wnt signalling triggers striatal synaptic degeneration and impaired motor behaviour in adult mice. Nat. Commun. 2014, 5, 4992. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Hu, J.; Qian, J.; Hackam, A.S. Expression of brain-derived neurotrophic factor is regulated by the Wnt signaling pathway. Neuroreport 2012, 23, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wang, K.; Wei, X.; Xie, T.; Lv, B.; Zhou, Q.; Wang, X. Interaction of NF-kappaB and Wnt/beta-catenin Signaling Pathways in Alzheimer’s Disease and Potential Active Drug Treatments. Neurochem. Res. 2021, 46, 711–731. [Google Scholar] [CrossRef] [PubMed]
- Nagu, P.; Sharma, V.; Behl, T.; Pathan, A.K.A.; Mehta, V. Molecular Insights to the Wnt Signaling During Alzheimer’s Disorder: A Potential Target for Therapeutic Interventions. J. Mol. Neurosci. 2022, 72, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Horrell, J.; Snitow, M.; Cui, J.; Gochnauer, H.; Syrett, C.M.; Kallish, S.; Seykora, J.T.; Liu, F.; Gaillard, D.; et al. WNT10A mutation causes ectodermal dysplasia by impairing progenitor cell proliferation and KLF4-mediated differentiation. Nat. Commun. 2017, 8, 15397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adaimy, L.; Chouery, E.; Megarbane, H.; Mroueh, S.; Delague, V.; Nicolas, E.; Belguith, H.; de Mazancourt, P.; Megarbane, A. Mutation in WNT10A is associated with an autosomal recessive ectodermal dysplasia: The odonto-onycho-dermal dysplasia. Am. J. Hum. Genet. 2007, 81, 821–828. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, M.; Guo, X.; Wang, K.Y.; Izumi, H.; Tsukamoto, M.; Nakashima, T.; Tasaki, T.; Kurose, N.; Uramoto, H.; Sasaguri, Y.; et al. Depletion of WNT10A Prevents Tumor Growth by Suppressing Microvessels and Collagen Expression. Int. J. Med. Sci. 2019, 16, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Tsukamoto, M.; Wang, K.Y.; Tasaki, T.; Murata, Y.; Okada, Y.; Yamanaka, Y.; Nakamura, E.; Yamada, S.; Izumi, H.; Zhou, Q.; et al. Findings as a starting point to unravel the underlying mechanisms of in vivo interactions involving Wnt10a in bone, fat and muscle. Bone 2019, 120, 75–84. [Google Scholar] [CrossRef]
- Wang, K.Y.; Yamada, S.; Izumi, H.; Tsukamoto, M.; Nakashima, T.; Tasaki, T.; Guo, X.; Uramoto, H.; Sasaguri, Y.; Kohno, K. Critical in vivo roles of WNT10A in wound healing by regulating collagen expression/synthesis in WNT10A-deficient mice. PLoS ONE 2018, 13, e0195156. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.-H.; Tasaki, T.; Tsukamoto, M.; Wang, K.-Y.; Azuma, K. Deficiency of Wnt10a causes female infertility via the β-catenin/Cyp19a1 pathway in mice. Int. J. Med. Sci. 2022, 19, 701–710. [Google Scholar] [CrossRef]
- Kim, C.S.; Chun, W.Y.; Shin, D.M. Dehydration Impairs Physical Growth and Cognitive Development in Young Mice. Nutrients 2020, 12, 670. [Google Scholar] [CrossRef] [Green Version]
- Struntz, K.H.; Siegel, J.A. Effects of methamphetamine exposure on anxiety-like behavior in the open field test, corticosterone, and hippocampal tyrosine hydroxylase in adolescent and adult mice. Behav. Brain Res. 2018, 348, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Jafari, Z.; Kolb, B.E.; Mohajerani, M.H. Chronic traffic noise stress accelerates brain impairment and cognitive decline in mice. Exp. Neurol. 2018, 308, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, Q.; Zhang, J.H.; Wang, K.Y.; Saito, T.; Saido, T.C.; Wang, X.; Gao, X.; Azuma, K. Microglia-Based Sex-Biased Neuropathology in Early-Stage Alzheimer’s Disease Model Mice and the Potential Pharmacologic Efficacy of Dioscin. Cells 2021, 10, 3261. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Suzuki, A.; Iinuma, M.; Wang, K.Y.; Kubo, K.Y.; Azuma, K. Effects of Maternal Chewing on Prenatal Stress-Induced Cognitive Impairments in the Offspring via Multiple Molecular Pathways. Int. J. Mol. Sci. 2020, 21, 5627. [Google Scholar] [CrossRef]
- Suzuki, A.; Iinuma, M.; Hayashi, S.; Sato, Y.; Azuma, K.; Kubo, K.Y. Maternal chewing during prenatal stress ameliorates stress-induced hypomyelination, synaptic alterations, and learning impairment in mouse offspring. Brain Res. 2016, 1651, 36–43. [Google Scholar] [CrossRef]
- Jahn-Eimermacher, A.; Lasarzik, I.; Raber, J. Statistical analysis of latency outcomes in behavioral experiments. Behav. Brain Res. 2011, 221, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.; Zlotnik, A.; Kofman, O.; Grinshpun, J.; Severynovska, O.; Brotfain, E.; Kut, R.; Natanel, D.; Melamed, I.; Boyko, M. Early life stress induces submissive behavior in adult rats. Behav. Brain Res. 2019, 372, 112025. [Google Scholar] [CrossRef]
- Holscher, C. Stress impairs performance in spatial water maze learning tasks. Behav. Brain Res. 1999, 100, 225–235. [Google Scholar] [CrossRef]
- Pitts, M.W. Barnes Maze Procedure for Spatial Learning and Memory in Mice. Bio-Protocol 2018, 8, e2744. [Google Scholar] [CrossRef] [Green Version]
- Kajimoto, K.; Hisada, C.; Ochi, S.; Yoshikawa, E.; Suzuki, A.; Tsugane, H.; Zhang, J.; Iinuma, M.; Kubo, K.Y.; Azuma, K. Maternal chewing improves prenatal stress-induced cognitive deficit and anxiety-like behavior associated with alterations of the apoptotic response and serotonin pathway in mouse offspring. Arch. Oral Biol. 2021, 130, 105245. [Google Scholar] [CrossRef]
- Inestrosa, N.C.; Tapia-Rojas, C.; Lindsay, C.B.; Zolezzi, J.M. Wnt Signaling Pathway Dysregulation in the Aging Brain: Lessons From the Octodon degus. Front. Cell Dev. Biol. 2020, 8, 734. [Google Scholar] [CrossRef] [PubMed]
- Mardones, M.D.; Andaur, G.A.; Varas-Godoy, M.; Henriquez, J.F.; Salech, F.; Behrens, M.I.; Couve, A.; Inestrosa, N.C.; Varela-Nallar, L. Frizzled-1 receptor regulates adult hippocampal neurogenesis. Mol. Brain 2016, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arredondo, S.B.; Valenzuela-Bezanilla, D.; Mardones, M.D.; Varela-Nallar, L. Role of Wnt Signaling in Adult Hippocampal Neurogenesis in Health and Disease. Front. Cell Dev. Biol. 2020, 8, 860. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Fu, X.H.; Zhou, D.; Li, J.M. The Role of Wnt/beta-Catenin Signaling Pathway in Disrupted Hippocampal Neurogenesis of Temporal Lobe Epilepsy: A Potential Therapeutic Target? Neurochem. Res. 2015, 40, 1319–1332. [Google Scholar] [CrossRef] [PubMed]
- Numakawa, T.; Odaka, H. Brain-Derived Neurotrophic Factor Signaling in the Pathophysiology of Alzheimer’s Disease: Beneficial Effects of Flavonoids for Neuroprotection. Int. J. Mol. Sci. 2021, 22, 5719. [Google Scholar] [CrossRef]
- Zhang, W.; Shi, Y.; Peng, Y.; Zhong, L.; Zhu, S.; Zhang, W.; Tang, S.J. Neuron activity-induced Wnt signaling up-regulates expression of brain-derived neurotrophic factor in the pain neural circuit. J. Biol. Chem. 2018, 293, 15641–15651. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.Y.; Wang, X.; Wang, Z.Y.; Wang, Y.Z.; Chen, L.W.; Luo, Z.J. Brain-derived neurotrophic factor stimulates proliferation and differentiation of neural stem cells, possibly by triggering the Wnt/beta-catenin signaling pathway. J. Neurosci. Res. 2013, 91, 30–41. [Google Scholar] [CrossRef]
- McLeod, F.; Salinas, P.C. Wnt proteins as modulators of synaptic plasticity. Curr. Opin. Neurobiol. 2018, 53, 90–95. [Google Scholar] [CrossRef]
- McQuate, A.; Latorre-Esteves, E.; Barria, A. A Wnt/Calcium Signaling Cascade Regulates Neuronal Excitability and Trafficking of NMDARs. Cell Rep. 2017, 21, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Rosso, S.B.; Inestrosa, N.C. WNT signaling in neuronal maturation and synaptogenesis. Front. Cell. Neurosci. 2013, 7, 103. [Google Scholar] [CrossRef] [Green Version]
- Calhoun, M.E.; Kurth, D.; Phinney, A.L.; Long, J.M.; Hengemihle, J.; Mouton, P.R.; Ingram, D.K.; Jucker, M. Hippocampal neuron and synaptophysin-positive bouton number in aging C57BL/6 mice. Neurobiol. Aging 1998, 19, 599–606. [Google Scholar] [CrossRef]
- Ramos-Fernandez, E.; Tapia-Rojas, C.; Ramirez, V.T.; Inestrosa, N.C. Wnt-7a Stimulates Dendritic Spine Morphogenesis and PSD-95 Expression Through Canonical Signaling. Mol. Neurobiol. 2019, 56, 1870–1882. [Google Scholar] [CrossRef] [PubMed]
- Assaf, N.; El-Shamarka, M.E.; Salem, N.A.; Khadrawy, Y.A.; El Sayed, N.S. Neuroprotective effect of PPAR alpha and gamma agonists in a mouse model of amyloidogenesis through modulation of the Wnt/beta catenin pathway via targeting alpha- and beta-secretases. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 97, 109793. [Google Scholar] [CrossRef] [PubMed]
- Farkhondeh, T.; Samarghandian, S.; Pourbagher-Shahri, A.M.; Sedaghat, M. The impact of curcumin and its modified formulations on Alzheimer’s disease. J. Cell. Physiol. 2019, 234, 16953–16965. [Google Scholar] [CrossRef]
- Hansen, D.V.; Hanson, J.E.; Sheng, M. Microglia in Alzheimer’s disease. J. Cell Biol. 2018, 217, 459–472. [Google Scholar] [CrossRef]
- Yirmiya, R.; Rimmerman, N.; Reshef, R. Depression as a microglial disease. Trends Neurosci. 2015, 38, 637–658. [Google Scholar] [CrossRef]
- Rana, T.; Behl, T.; Sehgal, A.; Srivastava, P.; Bungau, S. Unfolding the Role of BDNF as a Biomarker for Treatment of Depression. J. Mol. Neurosci. 2021, 71, 2008–2021. [Google Scholar] [CrossRef]
- Song, D.; Zhang, X.; Chen, J.; Liu, X.; Xue, J.; Zhang, L.; Lan, X. Wnt canonical pathway activator TWS119 drives microglial anti-inflammatory activation and facilitates neurological recovery following experimental stroke. J. Neuroinflamm. 2019, 16, 256. [Google Scholar] [CrossRef]
- Van Steenwinckel, J.; Schang, A.L.; Krishnan, M.L.; Degos, V.; Delahaye-Duriez, A.; Bokobza, C.; Csaba, Z.; Verdonk, F.; Montane, A.; Sigaut, S.; et al. Decreased microglial Wnt/beta-catenin signalling drives microglial pro-inflammatory activation in the developing brain. Brain 2019, 142, 3806–3833. [Google Scholar] [CrossRef]
- Vandenbark, A.A.; Offner, H.; Matejuk, S.; Matejuk, A. Microglia and astrocyte involvement in neurodegeneration and brain cancer. J. Neuroinflamm. 2021, 18, 298. [Google Scholar] [CrossRef]
- Dresselhaus, E.C.; Boersma, M.C.H.; Meffert, M.K. Targeting of NF-kappaB to Dendritic Spines Is Required for Synaptic Signaling and Spine Development. J. Neurosci. 2018, 38, 4093–4103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nejak-Bowen, K.; Kikuchi, A.; Monga, S.P. Beta-catenin-NF-κB interactions in murine hepatocytes: A complex to die for. Hepatology 2013, 57, 763–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonnies, E.; Trushina, E. Oxidative Stress, Synaptic Dysfunction, and Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 57, 1105–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angers, S.; Moon, R.T. Proximal events in Wnt signal transduction. Nat. Rev. Mol. Cell Biol. 2009, 10, 468–477. [Google Scholar] [CrossRef]
- Bayod, S.; Felice, P.; Andres, P.; Rosa, P.; Camins, A.; Pallas, M.; Canudas, A.M. Downregulation of canonical Wnt signaling in hippocampus of SAMP8 mice. Neurobiol. Aging 2015, 36, 720–729. [Google Scholar] [CrossRef]
- Ambacher, K.K.; Pitzul, K.B.; Karajgikar, M.; Hamilton, A.; Ferguson, S.S.; Cregan, S.P. The JNK- and AKT/GSK3beta- signaling pathways converge to regulate Puma induction and neuronal apoptosis induced by trophic factor deprivation. PLoS ONE 2012, 7, e46885. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | CAT# | Source | MW (kDa) | Dilution | Application |
|---|---|---|---|---|---|
| Amyloid-β | Ab201060 | Abcam | 38 | 1:1000 | WB |
| β-Catenin | Ab16051 | Abcam | 92 | 1:1000 | WB/IF |
| Wnt10a | * | 46 | 1:1000/1:5000 | WB/IF | |
| Doublecortin | Ab18732 | Abcam | 1:1000 | IF | |
| BDNF | Ab108319 | Abcam | 35 | 1:1000 | WB/IF |
| Iba1 | 013-27691 | Fujifilm | 1:1000 | IHC | |
| Iba1 | Ab48004 | Abcam | 17 | 1:1000 | WB |
| PSD95 | #2507 | CST | 95 | 1:1000 | WB |
| Synaptophysin | #36406 | CST | 38 | 1:1000 | WB |
| NF-κB | #8242 | CST | 65 | 1:1000 | WB |
| GSK-3β | #9832 | CST | 46 | 1:1000 | WB |
| IL-1β | #12426 | CST | 36 | 1:1000 | WB |
| GAPDH | #2118 | CST | 37 | 1:1000 | WB |
| TNF-α | #L1120 | SCBT | 27 | 1:500 | WB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.-H.; Tasaki, T.; Tsukamoto, M.; Wang, K.-Y.; Kubo, K.-y.; Azuma, K. Deletion of Wnt10a Is Implicated in Hippocampal Neurodegeneration in Mice. Biomedicines 2022, 10, 1500. https://doi.org/10.3390/biomedicines10071500
Zhang J-H, Tasaki T, Tsukamoto M, Wang K-Y, Kubo K-y, Azuma K. Deletion of Wnt10a Is Implicated in Hippocampal Neurodegeneration in Mice. Biomedicines. 2022; 10(7):1500. https://doi.org/10.3390/biomedicines10071500
Chicago/Turabian StyleZhang, Jia-He, Takashi Tasaki, Manabu Tsukamoto, Ke-Yong Wang, Kin-ya Kubo, and Kagaku Azuma. 2022. "Deletion of Wnt10a Is Implicated in Hippocampal Neurodegeneration in Mice" Biomedicines 10, no. 7: 1500. https://doi.org/10.3390/biomedicines10071500
APA StyleZhang, J.-H., Tasaki, T., Tsukamoto, M., Wang, K.-Y., Kubo, K.-y., & Azuma, K. (2022). Deletion of Wnt10a Is Implicated in Hippocampal Neurodegeneration in Mice. Biomedicines, 10(7), 1500. https://doi.org/10.3390/biomedicines10071500