Non-Genomic Effects of Xenoestrogen Mixtures

{kind=link}
Abstract
:1. Introduction
2. Mechanisms of Estrogenic Actions-Genomic & Non—Genomic Pathways
2.1. Different Types of Estrogen Receptors
2.2. Types of Non-Genomic Signaling Induced by Estrogens and Xenoestrogens and Their Functional Consequences
2.3. Non-Monotonic Dose Responses of Xenoestrogens
3. Types of Estrogens and Estrogen Mimetics
3.1. Physiologic Estrogens
3.2. Pharmaceutical and Personal Care Product Estrogens
3.3. Phytoestrogens
3.4. Synthetic/Anthropogenic Estrogens
4. Non-Genomic Actions of Estrogen/Xenoestrogen Mixtures
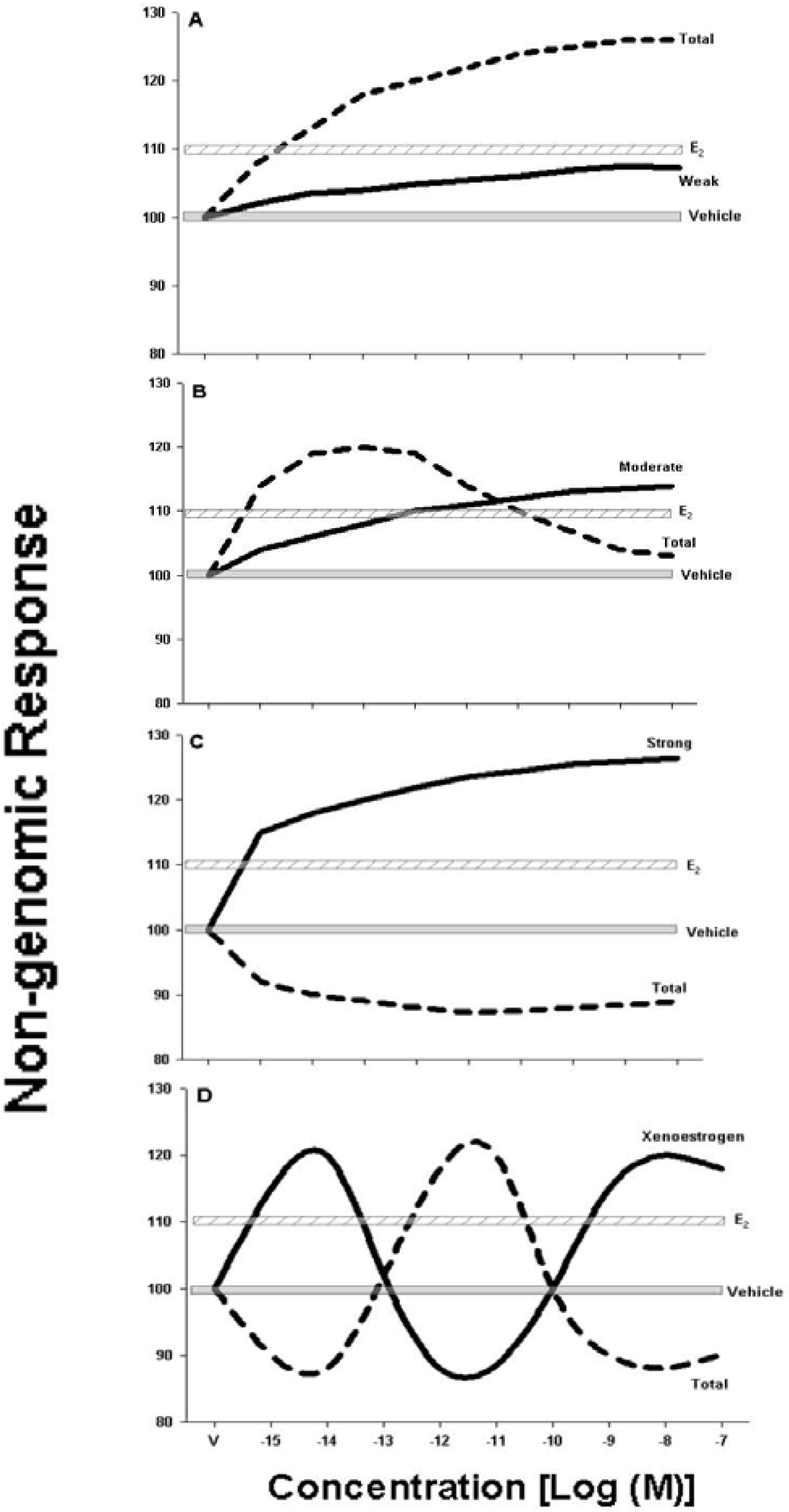
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Watson, C.S.; Jeng, Y.J.; Guptarak, J. Endocrine disruption via estrogen receptors that participate in nongenomic signaling pathways. J. Steroid Biochem. Mol. Biol. 2011, 127, 44–50. [Google Scholar] [CrossRef]
- Goncalves, C.R.; Cunha, R.W.; Barros, D.M.; Martinez, P.E. Effects of prenatal and postnatal exposure to a low dose of bisphenol A on behavior and memory in rats. Environ. Toxicol. Pharmacol. 2010, 30, 195–201. [Google Scholar]
- Li, D.K.; Zhou, Z.; Miao, M.; He, Y.; Wang, J.; Ferber, J.; Herrinton, L.J.; Gao, E.; Yuan, W. Urine bisphenol-A (BPA) level in relation to semen quality. Fertil. Steril. 2011, 95, 625–630. [Google Scholar] [CrossRef]
- Oehlmann, J.; Schulte-Oehlmann, U.; Kloas, W.; Jagnytsch, O.; Lutz, I.; Kusk, K.O.; Wollenberger, L.; Santos, E.M.; Paull, G.C.; Van Look, K.J.; Tyler, C.R. A critical analysis of the biological impacts of plasticizers on wildlife. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2047–2062. [Google Scholar]
- Sohoni, P.; Tyler, C.R.; Hurd, K.; Caunter, J.; Hetheridge, M.; Williams, T.; Woods, C.; Evans, M.; Toy, R.; Gargas, M.; Sumpter, J.P. Reproductive effects of long-term exposure to Bisphenol A in the fathead minnow (Pimephales promelas). Environ. Sci. Technol. 2001, 35, 2917–2925. [Google Scholar] [CrossRef]
- Wolstenholme, J.T.; Taylor, J.A.; Shetty, S.R.; Edwards, M.; Connelly, J.J.; Rissman, E.F. Gestational exposure to low dose bisphenol A alters social behavior in juvenile mice. PLoS One 2011, 6. [Google Scholar]
- Zhou, J.; Zhu, X.S.; Cai, Z.H. The impacts of bisphenol A (BPA) on abalone (Haliotis diversicolor supertexta) embryonic development. Chemosphere 2011, 82, 443–450. [Google Scholar] [CrossRef]
- Bouskine, A.; Nebout, M.; Brucker-Davis, F.; Benahmed, M.; Fenichel, P. Low doses of bisphenol A promote human seminoma cell proliferation by activating PKA and PKG via a membrane G-protein-coupled estrogen receptor. Environ. Health Perspect. 2009, 117, 1053–1058. [Google Scholar]
- Alonso-Magdalena, P.; Morimoto, S.; Ripoll, C.; Fuentes, E.; Nadal, A. The estrogenic effect of bisphenol A disrupts pancreatic beta-cell function in vivo and induces insulin resistance. Environ. Health Perspect. 2006, 114, 106–112. [Google Scholar]
- Midoro-Horiuti, T.; Tiwari, R.; Watson, C.S.; Goldblum, R.M. Maternal bisphenol a exposure promotes the development of experimental asthma in mouse pups. Environ. Health Perspect. 2010, 118, 273–277. [Google Scholar]
- Newbold, R.R.; Padilla-Banks, E.; Jefferson, W.N. Environmental estrogens and obesity. Mol. Cell Endocrinol. 2009, 304, 84–89. [Google Scholar] [CrossRef]
- Newbold, R.R. Developmental exposure to endocrine-disrupting chemicals programs for reproductive tract alterations and obesity later in life. Am. J. Clin. Nutr. 2011, 94, S1939–S1942. [Google Scholar] [CrossRef]
- Topp, E.; Starratt, A. Rapid mineralizatin of the enddocrine-disrupting chemical 4-nonylphenol in soil. Environ. Toxicol. Chem. 1999, 19, 313–318. [Google Scholar]
- Wang, J.; Xie, P.; Guo, N. Effects of nonylphenol on the growth and microcystin production of Microcystis strains. Environ. Res. 2007, 103, 70–78. [Google Scholar] [CrossRef]
- Kubwabo, C.; Kosarac, I.; Stewart, B.; Gauthier, B.R.; Lalonde, K.; Lalonde, P.J. Migration of bisphenol A from plastic baby bottles, baby bottle liners and reusable polycarbonate drinking bottles. Food Addit. Contam. A Chem. Anal. Control Expo Risk Assess. 2009, 26, 928–937. [Google Scholar]
- Dave, G.; Herger, G. Determination of detoxification to Daphnia magna of four pharmaceuticals and seven surfactants by activated sludge. Chemosphere 2012, 88, 459–466. [Google Scholar] [CrossRef]
- Dolar, D.; Gros, M.; Rodriguez-Mozaz, S.; Moreno, J.; Comas, J.; Rodriguez-Roda, I.; Barcelo, D. Removal of emerging contaminants from municipal wastewater with an integrated membrane system, MBR-RO. J. Hazard Mater. 2012. accepted. [Google Scholar]
- Kuruto-Niwa, R.; Nozawa, R.; Miyakoshi, T.; Shiozawa, T.; Terao, Y. Estrogenic activity of alkylphenols, bisphenol S, and their chlorinated derivatives using a GFP expression syste. Environ. Toxicol. Pharmacol. 2005, 19, 121–130. [Google Scholar] [CrossRef]
- Teuschler, L.; Klaunig, J.; Carney, E.; Chambers, J.; Conolly, R.; Gennings, C.; Giesy, J.; Hertzberg, R.; Klaassen, C.; Kodell, R.; Paustenbach, D.; Yang, R. Support of science-based decisions concerning the evaluation of the toxicology of mixtures: A new beginning. Regul. Toxicol. Pharmacol. 2002, 36, 34–39. [Google Scholar] [CrossRef]
- Alyea, R.A.; Laurence, S.E.; Kim, S.H.; Katzenellenbogen, B.S.; Katzenellenbogen, J.A.; Watson, C.S. The roles of membrane estrogen receptor subtypes in modulating dopamine transporters in PC-12 cells. J. Neurochem. 2008, 106, 1525–1533. [Google Scholar]
- Alyea, R.A.; Watson, C.S. Nongenomic mechanisms of physiological estrogen-mediated dopamine efflux. BMC Neurosci. 2009, 10. [Google Scholar]
- Jeng, Y.J.; Watson, C.S. Proliferative and anti-proliferative effects of dietary levels of phytoestrogens in rat pituitary GH3/B6/F10 cells—The involvement of rapidly activated kinases and caspases. BMC Cancer 2009, 9. [Google Scholar]
- Jeng, Y.J.; Kochukov, M.Y.; Watson, C.S. Membrane estrogen receptor-alpha-mediated nongenomic actions of phytoestrogens in GH3/B6/F10 pituitary tumor cells. J. Mol. Signal. 2009, 4. [Google Scholar]
- Jeng, Y.J.; Kochukov, M.; Watson, C.S. Combinations of physiologic estrogens with xenoestrogens alter calcium and kinase responses, prolactin release, and membrane estrogen receptor trafficking in rat pituitary cells. Environ. Health 2010, 9. [Google Scholar]
- Jeng, Y.J.; Watson, C.S. Combinations of physiologic estrogens with xenoestrogens alter ERK phosphorylation profiles in rat pituitary cells. Environ. Health Perspect. 2011, 119, 104–112. [Google Scholar]
- Kochukov, M.Y.; Jeng, Y.-J.; Watson, C.S. Alkylphenol xenoestrogens with varying carbon chain lengths differentially and potently activate signaling and functional responses in GH3/B6/F10 somatomammotropes. Environ. Health Perspect. 2009, 117, 723–730. [Google Scholar]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R., Jr.; Lee, D.H.; Shioda, T.; Soto, A.M.; Vom Saal, F.S.; Welshons, W.V.; Zoeller, R.T.; Myers, J.P. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef]
- Kortenkamp, A. Ten years of mixing cocktails: A review of combination effects of endocrine-disrupting chemicals. Environ. Health Perspect. 2007, 115(Suppl 1), 98–105. [Google Scholar] [CrossRef]
- Hayes, T.B.; Case, P.; Chui, S.; Chung, D.; Haeffele, C.; Haston, K.; Lee, M.; Mai, V.P.; Marjuoa, Y.; Parker, J.; Tsui, M. Pesticide mixtures, endocrine disruption, and amphibian declines: Are we underestimating the impact? Environ. Health Perspect. 2006, 114(Suppl 1), 40–50. [Google Scholar]
- Vandenberg, L.N.; Maffini, M.V.; Sonnenschein, C.; Rubin, B.S.; Soto, A.M. Bisphenol-A and the great divide: A review of controversies in the field of endocrine disruption. Endocr. Rev. 2009, 30, 75–95. [Google Scholar]
- Decherf, S.; Seugnet, I.; Fini, J.B.; Clerget-Froidevaux, M.S.; Demeneix, B.A. Disruption of thyroid hormone-dependent hypothalamic set-points by environmental contaminants. Mol. Cell Endocrinol. 2010, 323, 172–182. [Google Scholar] [CrossRef]
- Diamanti-Kandarakis, E.; Bourguignon, J.P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-disrupting chemicals: An Endocrine Society scientific statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef]
- Kretschmer, X.C.; Baldwin, W.S. CAR and PXR: Xenosensors of endocrine disrupters? Chem. Biol. Interact. 2005, 155, 111–128. [Google Scholar] [CrossRef]
- Qin, X.Y.; Zaha, H.; Nagano, R.; Yoshinaga, J.; Yonemoto, J.; Sone, H. Xenoestrogens down-regulate aryl-hydrocarbon receptor nuclear translocator 2 mRNA expression in human breast cancer cells via an estrogen receptor alpha-dependent mechanism. Toxicol. Lett. 2011, 206, 152–157. [Google Scholar] [CrossRef]
- Deroo, B.J.; Korach, K.S. Estrogen receptors and human disease. J. Clin. Invest. 2006, 116, 561–570. [Google Scholar] [CrossRef]
- Li, L.; Haynes, M.P.; Bender, J.R. Plasma membrane localization and function of the estrogen receptor alpha variant (ER46) in human endothelial cells. Proc. Natl. Acad. Sci. USA 2003, 100, 4807–4812. [Google Scholar] [CrossRef]
- Pappas, T.C.; Gametchu, B.; Yannariello-Brown, J.; Collins, T.J.; Watson, C.S. Membrane estrogen receptors in GH3/B6 cells are associated with rapid estrogen-induced release of prolactin. Endocrine 1994, 2, 813–822. [Google Scholar]
- Pietras, R.J.; Levin, E.R.; Szego, C.M. Estrogen receptors and cell signaling. Science 2005, 310, 51–53. [Google Scholar] [CrossRef]
- Pietras, R.J.; Szego, C.M. Cell membrane estrogen receptors resurface. Nat. Med. 1999, 5, 1330. [Google Scholar] [CrossRef]
- Kang, L.; Zhang, X.; Xie, Y.; Tu, Y.; Wang, D.; Liu, Z.; Wang, Z.Y. Involvement of estrogen receptor variant ER-alpha36, not GPR30, in nongenomic estrogen signaling. Mol. Endocrinol. 2010, 24, 709–721. [Google Scholar] [CrossRef]
- Thomas, P.; Pang, Y.; Filardo, E.J.; Dong, J. Identity of an estrogen membrane receptor coupled to a G protein in human breast cancer cells. Endocrinology 2005, 146, 624–632. [Google Scholar]
- Thomas, P.; Dong, J. Binding and activation of the seven-transmembrane estrogen receptor GPR30 by environmental estrogens: A potential novel mechanism of endocrine disruption. J. Steroid Biochem. Mol. Biol. 2006, 102, 175–179. [Google Scholar] [CrossRef]
- Watson, C.S.; Gametchu, B. Membrane-initiated steroid actions and the proteins that mediate them. Proc. Soc. Exp. Biol. Med. 1999, 220, 9–19. [Google Scholar]
- Watson, C.S.; Campbell, C.H.; Gametchu, B. Membrane estrogen receptors on rat pituitary tumor cells: Immunoidentification and responses to estradiol and xenoestrogens. Exp. Physiol. 1999, 84, 1013–1022. [Google Scholar] [CrossRef]
- Campbell, C.H.; Watson, C.S. A comparison of membrane vs. intracellular estrogen receptor-alpha in GH(3)/B6 pituitary tumor cells using a quantitative plate immunoassay. Steroids 2001, 66, 727–736. [Google Scholar] [CrossRef]
- Powell, C.E.; Soto, A.M.; Sonnenschein, C. Identification and characterization of membrane estrogen receptor from MCF7 estrogen-target cells. J. Steroid Biochem. Mol. Biol. 2001, 77, 97–108. [Google Scholar] [CrossRef]
- Norfleet, A.M.; Clarke, C.; Gametchu, B.; Watson, C.S. Antibodies to the estrogen receptor-α modulate prolactin release from rat pituitary tumor cells through plasma membrane estrogen receptors. FASEB J. 2000, 14, 157–165. [Google Scholar]
- Razandi, M.; Alton, G.; Pedram, A.; Ghonshani, S.; Webb, P.; Levin, E.R. Identification of a structural determinant necessary for the localization and function of estrogen receptor alpha at the plasma membrane. Mol. Cell Biol. 2003, 23, 1633–1646. [Google Scholar] [CrossRef]
- Schlegel, A.; Wang, C.G.; Katzenellenbogen, B.S.; Pestell, R.G.; Lisanti, M.P. Caveolin-1 potentiates estrogen receptor alpha (ER alpha) signaling—Caveolin-1 drives ligand-independent nuclear translocation and activation of ER alpha. J. Biol. Chem. 1999, 274, 33551–33556. [Google Scholar]
- Watson, C.S.; Jeng, Y.J.; Hu, G.; Wozniak, A.; Bulayeva, N.; Guptarak, J. Estrogen- and xenoestrogen-induced ERK signaling in pituitary tumor cells involves estrogen receptor-alpha interactions with G protein-alphai and caveolin I. Steroids 2011, 77, 424–432. [Google Scholar]
- Razandi, M.; Oh, P.; Pedram, A.; Schnitzer, J.; Levin, E.R. ERs associate with and regulate the production of caveolin: Implications for signaling and cellular actions. Mol. Endocrinol. 2002, 16, 100–115. [Google Scholar] [CrossRef]
- Pedram, A.; Razandi, M.; Levin, E.R. Nature of functional estrogen receptors at the plasma membrane. Mol. Endocrinol. 2006, 20, 1996–2009. [Google Scholar]
- Norfleet, A.M.; Thomas, M.L.; Watson, C.S. Modulation of Membrane Estrogen Receptor-α Levels by Nuclear Estrogen Receptor-α Antisense Oligodeoxynucleotides in the Rat Pituitary Tumor Cell Line, GH3/B6/F10. In Presented at Endocrine Society Meeting, San Diego, CA, USA, 12–15 June 1999.
- Norfleet, A.M.; Thomas, M.L.; Gametchu, B.; Watson, C.S. Estrogen receptor-α detected on the plasma membrane of aldehyde-fixed GH3/B6/F10 rat pituitary cells by enzyme-linked immunocytochemistry. Endocrinology 1999, 140, 3805–3814. [Google Scholar]
- Pedram, A.; Razandi, M.; Sainson, R.C.; Kim, J.K.; Hughes, C.C.; Levin, E.R. A conserved mechanism for steroid receptor translocation to the plasma membrane. J. Biol. Chem. 2007, 282, 22278–22288. [Google Scholar]
- Kuiper, G.G.; Carlsson, B.; Grandien, K.; Enmark, E.; Haggblad, J.; Nilsson, S.; Gustafsson, J.-Å. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 1997, 138, 863–870. [Google Scholar]
- Wozniak, A.L.; Bulayeva, N.N.; Watson, C.S. Xenoestrogens at picomolar to nanomolar concentrations trigger membrane estrogen receptor-alpha-mediated Ca2+ fluxes and prolactin release in GH3/B6 pituitary tumor cells. Environ. Health Perspect. 2005, 113, 431–439. [Google Scholar]
- Hunter, T. Protein kinases and phosphatases: The yin and yang of protein phosphorylation and signaling. Cell 1995, 80, 225–236. [Google Scholar] [CrossRef]
- Junttila, M.R.; Li, S.P.; Westermarck, J. Phosphatase-mediated crosstalk between MAPK signaling pathways in the regulation of cell survival. FASEB J. 2008, 22, 954–965. [Google Scholar]
- Nordstrom, E.; Fisone, G.; Kristensson, K. Opposing effects of ERK and p38-JNK MAP kinase pathways on formation of prions in GT1-1 cells. FASEB J. 2009, 23, 613–622. [Google Scholar]
- Xia, Z.; Dickens, M.; Raingeaud, J.; Davis, R.J.; Greenberg, M.E. Opposing effects of ERK and JNK-p38 MAP kinases on apoptosis. Science 1995, 270, 1326–1331. [Google Scholar]
- Jeng, Y.-J.; Watson, C.S.; Thomas, M.L. Identification of vitamin D-stimulated alkaline phosphatase in IEC-6 cells, a rat small intestine crypt cell line. Exp. Cell Res. 1994, 212, 338–343. [Google Scholar] [CrossRef]
- Bulayeva, N.N.; Wozniak, A.; Lash, L.L.; Watson, C.S. Mechanisms of membrane estrogen receptor-{alpha}-mediated rapid stimulation of Ca2+ levels and prolactin release in a pituitary cell line. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E388–E397. [Google Scholar] [CrossRef]
- Alyea, R.A.; Watson, C.S. Differential regulation of dopamine transporter function and location by low concentrations of environmental estrogens and 17beta-estradiol. Environ. Health Perspect. 2009, 117, 778–783. [Google Scholar] [CrossRef]
- Binda, F.; Dipace, C.; Bowton, E.; Robertson, S.D.; Lute, B.J.; Fog, J.U.; Zhang, M.; Sen, N.; Colbran, R.J.; Gnegy, M.E.; Gether, U.; Javitch, J.A.; Erreger, K.; Galli, A. Syntaxin 1A interaction with the dopamine transporter promotes amphetamine-induced dopamine efflux. Mol. Pharmacol. 2008, 74, 1101–1108. [Google Scholar]
- Foster, J.D.; Cervinski, M.A.; Gorentla, B.K.; Vaughan, R.A. Regulation of the dopamine transporter by phosphorylation. Handb. Exp. Pharmacol. 2006, 175, 197–214. [Google Scholar] [CrossRef]
- International Dose-Response Society. Available online: http://www.dose-response.org/ (accessed on 23 July 2012).
- U.S.Food and Drug Administration, Bisphenol A (BPA):Use in Food Contact Applications; FDA: Silver Spring, MD, USA, 2012.
- Fenner-Crisp, P.A. Endocrine modulators: Risk characterization and assessment. Toxicol. Pathol. 2000, 28, 438–440. [Google Scholar] [CrossRef]
- Lucier, G.W. Dose-response relationships for endocrine disruptors: What we know and what we don’t know. Regul. Toxicol. Pharmacol. 1997, 26, 34–35. [Google Scholar] [CrossRef]
- Myers, J.P.; Zoeller, R.T.; vom Saal, F.S. A clash of old and new scientific concepts in toxicity, with important implications for public health. Environ. Health Perspect. 2009, 117, 1652–1655. [Google Scholar]
- Teuschler, L.; Klaunig, J.; Carney, E.; Chambers, J.; Conolly, R.; Gennings, C.; Giesy, J.; Hertzberg, R.; Klaassen, C.; Kodell, R.; Paustenbach, D.; Yang, R. Support of science-based decisions concerning the evaluation of the toxicology of mixtures: A new beginning. Regul. Toxicol. Pharmacol. 2002, 36, 34–39. [Google Scholar] [CrossRef]
- Watson, C.S.; Jeng, Y.J.; Guptarak, J. Endocrine disruption via estrogen receptors that participate in nongenomic signaling pathways. J. Steroid Biochem. Mol. Biol. 2011, 127, 44–50. [Google Scholar] [CrossRef]
- Soto, A.M.; Rubin, B.S.; Sonnenschein, C. Interpreting endocrine disruption from an integrative biology perspective. Mol. Cell Endocrinol. 2009, 304, 3–7. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Iavicoli, I.; Calabrese, V. Hormesis: Why it is important to biogerontologists. Biogerontology 2012, 13, 215–235. [Google Scholar] [CrossRef]
- Calabrese, E.J. Getting the dose-response wrong: Why hormesis became marginalized and the threshold model accepted. Arch. Toxicol. 2009, 83, 227–247. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormesis is central to toxicology, pharmacology and risk assessment. Hum. Exp. Toxicol. 2010, 29, 249–261. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Wadia, P.R.; Schaeberle, C.M.; Rubin, B.S.; Sonnenschein, C.; Soto, A.M. The mammary gland response to estradiol: Monotonic at the cellular level, non-monotonic at the tissue-level of organization? J. Steroid Biochem. Mol. Biol. 2006, 101, 263–274. [Google Scholar] [CrossRef]
- Watson, C.S. The Identities of Membrane Steroid Receptors....and Other Proteins Mediating Nongenomic Steroid Action; Kluwer Academic Publishers: Boston, MA, USA, 2003. [Google Scholar]
- Bermudez, O.; Marchetti, S.; Pages, G.; Gimond, C. Post-translational regulation of the ERK phosphatase DUSP6/MKP3 by the mTOR pathway. Oncogene 2008, 27, 3685–3691. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, B.; Wang, M.; Carr, B.I. Cdc25A and ERK interaction: EGFR-independent ERK activation by a protein phosphatase Cdc25A inhibitor, compound 5. J. Cell Physiol. 2005, 204, 437–444. [Google Scholar] [CrossRef]
- Yu, L.G.; Packman, L.C.; Weldon, M.; Hamlett, J.; Rhodes, J.M. Protein phosphatase 2A, a negative regulator of the ERK signaling pathway, is activated by tyrosine phosphorylation of putative HLA class II-associated protein I (PHAPI)/pp32 in response to the antiproliferative lectin, jacalin. J. Biol. Chem. 2004, 279, 41377–41383. [Google Scholar]
- Zivadinovic, D.; Watson, C.S. Membrane estrogen receptor-alpha levels predict estrogen-induced ERK1/2 activation in MCF-7 cells. Breast Cancer Res. 2005, 7, R130–R144. [Google Scholar]
- Bulayeva, N.N.; Gametchu, B.; Watson, C.S. Quantitative measurement of estrogen-induced ERK 1 and 2 activation via multiple membrane-initiated signaling pathways. Steroids 2004, 69, 181–192. [Google Scholar] [CrossRef]
- Stormshak, F.; Leake, R.; Wertz, N.; Gorski, J. Stimulatory and inhibitory effects of estrogen on uterine DNA synthesis. Endocrinology 1976, 99, 1501–1511. [Google Scholar] [CrossRef]
- Wiklund, J.; Wertz, N.; Gorski, J. A comparison of estrogen effects on uterine and pituitary growth and prolactin synthesis in F344 and Holtzman rats. Endocrinology 1981, 109, 1700–1707. [Google Scholar] [CrossRef]
- Watson, C.S.; Jeng, Y.J.; Kochukov, M.Y. Nongenomic signaling pathways of estrogen toxicity. Toxicol. Sci. 2010, 115, 1–11. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormesis: Why it is important to toxicology and toxicologists. Environ. Toxicol. Chem. 2008, 27, 1451–1474. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormesis and mixtures. Toxicol. Appl. Pharmacol. 2008, 229, 262–263. [Google Scholar] [CrossRef]
- Yang, R.S.; Dennison, J.E. Initial analyses of the relationship between “Thresholds” of toxicity for individual chemicals and “Interaction Thresholds” for chemical mixtures. Toxicol. Appl. Pharmacol. 2007, 223, 133–138. [Google Scholar] [CrossRef]
- Sheehan, D.M.; Willingham, E.; Gaylor, D.; Bergeron, J.M.; Crews, D. No threshold dose for estradiol-induced sex reversal of turtle embryos: How little is too much? Environ. Health Perspect. 1999, 107, 155–159. [Google Scholar]
- Sheehan, D.M. No-threshold dose-response curves for nongenotoxic chemicals: Findings and applications for risk assessment. Environ. Res. 2006, 100, 93–99. [Google Scholar] [CrossRef]
- Cornwell, T.; Cohick, W.; Raskin, I. Dietary phytoestrogens and health. Phytochemistry 2004, 65, 995–1016. [Google Scholar]
- Watson, C.S.; Alyea, R.A.; Cunningham, K.A.; Jeng, Y.J. Estrogens of multiple classes and their role in mental health disease mechanisms. Int. J. Womens Health 2010, 2, 153–166. [Google Scholar]
- Greenspan, F.S.; Gardner, D.G. Appendix: Normal Hormone Reference Ranges. In Basic and Clinical Endocrinology, 7th; Greenspan, F.S., Gardner, D.G., Eds.; Lange Medical Books: McGraw Hill, NY, USA, 2004; pp. 925–926. [Google Scholar]
- Benn, P.A. Advances in prenatal screening for Down syndrome: I. General principles and second trimester testing. Clin. Chim. Acta 2002, 323, 1–16. [Google Scholar] [CrossRef]
- Shenhav, S.; Gemer, O.; Volodarsky, M.; Zohav, E.; Segal, S. Midtrimester triple test levels in women with severe preeclampsia and HELLP syndrome. Acta Obstet. Gynecol Scand. 2003, 82, 912–915. [Google Scholar] [CrossRef]
- Greenlee, H.; Chen, Y.; Kabat, G.C.; Wang, Q.; Kibriya, M.G.; Gurvich, I.; Sepkovic, D.W.; Bradlow, H.L.; Senie, R.T.; Santella, R.M.; Ahsan, H. Variants in estrogen metabolism and biosynthesis genes and urinary estrogen metabolites in women with a family history of breast cancer. Breast Cancer Res. Treat. 2007, 102, 111–117. [Google Scholar] [CrossRef]
- Riza, E.; dos Santos Silva, I.; De Stavola, B.; Bradlow, H.L.; Sepkovic, D.W.; Linos, D.; Linos, A. Urinary estrogen metabolites and mammographic parenchymal patterns in postmenopausal women. Cancer Epidemiol. Biomarkers Prev. 2001, 10, 627–634. [Google Scholar]
- Mermelstein, P.G.; Becker, J.B.; Surmeier, D.J. Estradiol reduces calcium currents in rat neostriatal neurons via a membrane receptor. J. Neurosci. 1996, 16, 595–604. [Google Scholar]
- Schwarz, S.; Pohl, P. Steroids and opioid receptors. J. Steroid Biochem. Mol. Biol. 1994, 48, 391–402. [Google Scholar] [CrossRef]
- Watson, C.S.; Jeng, Y.J.; Kochukov, M.Y. Nongenomic actions of estradiol compared with estrone and estriol in pituitary tumor cell signaling and proliferation. FASEB J. 2008, 22, 3328–3336. [Google Scholar] [CrossRef]
- Jean, J.; Perrodin, Y.; Pivot, C.; Trepo, D.; Perraud, M.; Droguet, J.; Tissot-Guerraz, F.; Locher, F. Identification and prioritization of bioaccumulable pharmaceutical substances discharged in hospital effluents. J. Environ. Manag. 2012, 103C, 113–121. [Google Scholar]
- Fent, K.; Weston, A.A.; Caminada, D. Ecotoxicology of human pharmaceuticals. Aquat. Toxicol. 2006, 76, 122–159. [Google Scholar] [CrossRef]
- Gall, H.E.; Sassman, S.A.; Lee, L.S.; Jafvert, C.T. Hormone discharges from a midwest tile-drained agroecosystem receiving animal wastes. Environ. Sci. Technol. 2011, 45, 8755–8764. [Google Scholar]
- Lu, G.; Yan, Z.; Wang, Y.; Chen, W. Assessment of estrogenic contamination and biological effects in Lake Taihu. Ecotoxicology 2011, 20, 974–981. [Google Scholar] [CrossRef]
- Soto, A.M.; Calabro, J.M.; Prechtl, N.V.; Yau, A.Y.; Orlando, E.F.; Daxenberger, A.; Kolok, A.S.; Guillette, L.J., Jr.; le Bizec, B.; Lange, I.G.; Sonnenschein, C. Androgenic and estrogenic activity in water bodies receiving cattle feedlot effluent in Eastern Nebraska, USA. Environ. Health Perspect. 2004, 112, 346–352. [Google Scholar]
- Touraud, E.; Roig, B.; Sumpter, J.P.; Coetsier, C. Drug residues and endocrine disruptors in drinking water: risk for humans? Int. J. Hyg. Environ. Health. 2011, 214, 437–441. [Google Scholar] [CrossRef]
- Zhou, Y.; Zha, J.; Xu, Y.; Lei, B.; Wang, Z. Occurrences of six steroid estrogens from different effluents in Beijing, China. Environ. Monit. Assess. 2012, 184, 1719–1729. [Google Scholar] [CrossRef]
- Mustafa, A.M.; Malintan, N.T.; Seelan, S.; Zhan, Z.; Mohamed, Z.; Hassan, J.; Pendek, R.; Hussain, R.; Ito, N. Phytoestrogens levels determination in the cord blood from Malaysia rural and urban populations. Toxicol. Appl. Pharmacol. 2007, 222, 25–32. [Google Scholar]
- Whitten, P.L.; Patisaul, H.B. Cross-species and interassay comparisons of phytoestrogen action. Environ. Health Perspect. 2001, 109, 5–20. [Google Scholar]
- Adlercreutz, H.; Fotsis, T.; Lampe, J.; Wahala, K.; Makela, T.; Brunow, G.; Hase, T. Quantitative determination of lignans and isoflavonoids in plasma of omnivorous and vegetarian women by isotope dilution gas chromatography-mass spectrometry. Scand. J. Clin. Lab. Invest. Suppl. 1993, 215, 5–18. [Google Scholar]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef]
- Lippi, G.; Franchini, M.; Favaloro, E.J.; Targher, G. Moderate red wine consumption and cardiovascular disease risk: Beyond the “French paradox”. Semin. Thromb Hemost. 2010, 36, 59–70. [Google Scholar] [CrossRef]
- Eden, J.A. Phytoestrogens for menopausal symptoms: A review. Maturitas 2012, 72, 157–159. [Google Scholar] [CrossRef]
- Pitkin, J. Alternative and complementary therapies for the menopause. Menopause Int. 2012, 18, 20–27. [Google Scholar] [CrossRef]
- Sunita, P.; Pattanayak, S.P. Phytoestrogens in postmenopausal indications: A theoretical perspective. Pharmacogn Rev. 2011, 5, 41–47. [Google Scholar] [CrossRef]
- Jeng, Y.J.; Kochukov, M.; Nauduri, D.; Kaphalia, B.S.; Watson, C.S. Subchronic exposure to phytoestrogens alone and in combination with diethylstilbestrol—Pituitary tumor induction in Fischer 344 rats. Nutr. Metab. (Lond.) 2010, 7. [Google Scholar]
- Jefferson, W.N.; Patisaul, H.B.; Williams, C.J. Reproductive consequences of developmental phytoestrogen exposure. Reproduction. 2012, 143, 247–260. [Google Scholar] [CrossRef]
- Adlercreutz, H.; Yamada, T.; Wahala, K.; Watanabe, S. Maternal and neonatal phytoestrogens in Japanese women during birth. Am. J. Obstet. Gynecol. 1999, 180, 737–743. [Google Scholar] [CrossRef]
- Cao, Y.; Calafat, A.M.; Doerge, D.R.; Umbach, D.M.; Bernbaum, J.C.; Twaddle, N.C.; Ye, X.; Rogan, W.J. Isoflavones in urine, saliva, and blood of infants: Data from a pilot study on the estrogenic activity of soy formula. J. Expo. Sci. Environ. Epidemiol. 2009, 19, 223–234. [Google Scholar] [CrossRef]
- Setchell, K.D.; Zimmer-Nechemias, L.; Cai, J.; Heubi, J.E. Isoflavone content of infant formulas and the metabolic fate of these phytoestrogens in early life. Am. J. Clin. Nutr. 1998, 68, S1453–S1461. [Google Scholar]
- Gaido, K.W.; Leonard, L.S.; Lovell, S.; Gould, J.C.; Babai, D.; Portier, C.J.; McDonnell, D.P. Evaluation of chemicals with endocrine modulating activity in a yeast-based steroid hormone receptor gene transcription assay. Toxicol. Appl. Pharmacol. 1997, 143, 205–212. [Google Scholar] [CrossRef]
- Mazerolles, G.; Preys, S.; Bouchut, C.; Meudec, E.; Fulcrand, H.; Souquet, J.M.; Cheynier, V. Combination of several mass spectrometry ionization modes: A multiblock analysis for a rapid characterization of the red wine polyphenolic composition. Anal. Chim. Acta 2010, 678, 195–202. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, Y.; Lu, J. Phenolic contents and compositions in skins of red wine grape cultivars among various genetic backgrounds and originations. Int. J. Mol. Sci. 2012, 13, 3492–3510. [Google Scholar] [CrossRef]
- Mnif, W.; Hassine, A.I.; Bouaziz, A.; Bartegi, A.; Thomas, O.; Roig, B. Effect of endocrine disruptor pesticides: A review. Int. J. Environ. Res. Public Health 2011, 8, 2265–2303. [Google Scholar] [CrossRef]
- Soto, A.M.; Chung, K.L.; Sonnenschein, C. The pesticides endosulfan, toxaphene, and dieldrin have estrogenic effects on human estrogen-sensitive cells. Environ. Health Perspect. 1994, 102, 380–383. [Google Scholar] [CrossRef]
- Shaw, J.; deCatanzaro, D. Estrogenicity of parabens revisited: Impact of parabens on early pregnancy and an uterotrophic assay in mice. Reprod. Toxicol. 2009, 28, 26–31. [Google Scholar] [CrossRef]
- Bonefeld-Jorgensen, E.C.; Long, M.; Hofmeister, M.V.; Vinggaard, A.M. Endocrine-disrupting potential of bisphenol A, bisphenol A dimethacrylate, 4-n-nonylphenol, and 4-n-octylphenol in vitro: New data and a brief review. Environ. Health Perspect. 2007, 115(Suppl 1), 69–76. [Google Scholar]
- Isidori, M.; Lavorgna, M.; Palumbo, M.; Piccioli, V.; Parrella, A. Influence of alkylphenols and trace elements in toxic, genotoxic, and endocrine disruption activity of wastewater treatment plants. Environ. Toxicol. Chem. 2007, 26, 1686–1694. [Google Scholar] [CrossRef]
- Kujawinski, E.B.; Kido Soule, M.C.; Valentine, D.L.; Boysen, A.K.; Longnecker, K.; Redmond, M.C. Fate of dispersants associated with the deepwater horizon oil spill. Environ. Sci. Technol. 2011, 45, 1298–1306. [Google Scholar]
- Ahel, M.; McEvoy, J.; Giger, W. Bioaccumulation of the lipophilic metabolites of nonionic surfactants in freshwater organisms. Environ. Pollut. 1993, 79, 243–248. [Google Scholar] [CrossRef]
- Deblonde, T.; Cossu-Leguille, C.; Hartemann, P. Emerging pollutants in wastewater: A review of the literature. Int. J. Hyg. Environ. Health. 2011, 214, 442–448. [Google Scholar] [CrossRef]
- Fleisch, A.F.; Sheffield, P.E.; Chinn, C.; Edelstein, B.L.; Landrigan, P.J. Bisphenol A and related compounds in dental materials. Pediatrics. 2010, 126, 760–768. [Google Scholar]
- Geens, T.; Goeyens, L.; Covaci, A. Are potential sources for human exposure to bisphenol-A overlooked? Int. J. Hyg. Environ. Health 2011, 214, 339–347. [Google Scholar] [CrossRef]
- Huang, Y.Q.; Wong, C.K.; Zheng, J.S.; Bouwman, H.; Barra, R.; Wahlstrom, B.; Neretin, L.; Wong, M.H. Bisphenol A (BPA) in China: A review of sources, environmental levels, and potential human health impacts. Environ. Int. 2012, 42, 91–99. [Google Scholar] [CrossRef]
- Myers, D.E.; Hutz, R.J. Current status of potential bisphenol toxicity in dentistry. Gen. Dent. 2011, 59, 262–265. [Google Scholar]
- Alonso-Magdalena, P.; Vieira, E.; Soriano, S.; Menes, L.; Burks, D.; Quesada, I.; Nadal, A. Bisphenol A exposure during pregnancy disrupts glucose homeostasis in mothers and adult male offspring. Environ. Health Perspect. 2010, 118, 1243–1250. [Google Scholar]
- Soto, A.M.; Vandenberg, L.N.; Maffini, M.V.; Sonnenschein, C. Does breast cancer start in the womb? Basic Clin. Pharmacol. Toxicol. 2008, 102, 125–133. [Google Scholar] [CrossRef]
- The Scientis -Jef Akst. US Doesn’t Ban BPA—The FDA Announces that BPA will Continue to be Permitted in Food and Beverage Containers. Available online: http://the-scientist.com/2012/04/02/us-doesnt-ban-bpa/ (accessed on 2 April 2012).
- Canada Gazette. Order Adding Toxic Substances to Schedule 1 to the Canadian Environmental Protection Act. 1999. Available online: http://www.gazette.gc.ca/rp-pr/p2/2012/2012-03-28/html/sor-dors40-eng.html (accessed on 26 April 2012).
- European Union. Amending Directive 2002/72/EC as Regards the Restriction of Use of Bisphenol A in Plastic Feeding Bottles. 2002. Available online: http://eurlex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2011:026:0011:0014:EN:PDF (accessed on 21 July 2012).
- Canada Bans BPA from Baby Bottles. Available online: http://www.washingtonpost.com/wp-dyn/content/article/2008/04/18/AR2008041803036.html (accessed on 26 April 2012).
- Bouskine, A.; Nebout, M.; Mograbi, B.; Brucker-Davis, F.; Roger, C.; Fenichel, P. Estrogens promote human testicular germ cell cancer through a membrane-mediated activation of extracellular regulated kinase and protein kinase A. Endocrinology 2008, 149, 565–573. [Google Scholar]
- Bulayeva, N.N.; Watson, C.S. Xenoestrogen-induced ERK-1 and ERK-2 activation via multiple membrane-initiated signaling pathways. Environ. Health Perspect. 2004, 112, 1481–1487. [Google Scholar] [CrossRef]
- Otto, C.; Fuchs, I.; Altmann, H.; Klewer, M.; Schwarz, G.; Bohlmann, R.; Nguyen, D.; Zorn, L.; Vonk, R.; Prelle, K.; Osterman, T.; Malmstrom, C.; Fritzemeier, K.H. In vivo characterization of estrogen receptor modulators with reduced genomic versus nongenomic activity in vitro. J. Steroid Biochem. Mol. Biol. 2008, 111, 95–100. [Google Scholar] [CrossRef]
- Yang, J.; Cao, J.; Sun, X.; Feng, Z.; Hao, D.; Zhao, X.; Sun, C. Effects of long-term exposure to low levels of organophosphorous pesticides and their mixture on altered antioxidative defense mechanisms and lipid peroxidation in rat liver. Cell Biochem. Funct. 2012, 30, 122–128. [Google Scholar] [CrossRef]
- Reffstrup, T.K.; Larsen, J.C.; Meyer, O. Risk assessment of mixtures of pesticides. Current approaches and future strategies. Regul. Toxicol. Pharmacol. 2010, 56, 174–192. [Google Scholar] [CrossRef]
- Watson, C.S.; Bulayeva, N.N.; Wozniak, A.L.; Alyea, R.A. Xenoestrogens are potent activators of nongenomic estrogenic responses. Steroids 2007, 72, 124–134. [Google Scholar] [CrossRef]
- Boobis, A.; Budinsky, R.; Collie, S.; Crofton, K.; Embry, M.; Felter, S.; Hertzberg, T.; Kopp, D.; Mihlan, G.; Mumtaz, M.; Price, P.; Solomon, K.; Teuschler, L.; Yang, R.; Zaleski, R. Critical analysis of literature on low-dose synergy for use in screening chemical mixtures for risk assessment. Crit. Rev. Toxicol. 2011, 41, 369–383. [Google Scholar] [CrossRef]
- ATSDR. DDT, DDE, and DDD. Available online: http://www.atsdr.cdc.gov/PHS/PHS.asp?id=79&tid=20 (accessed on 21 July 2012).
- US EPA. Supplementary Guidance for Conducting Health Risk Assessment of Chemical Mixtures; EPA/630/R-00/002. Available online: http://cfpub.epa.gov/ncea/cfm/recordisplay.cfm?deid=20533 (accessed on 21 July 2012).
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Viñas, R.; Jeng, Y.-J.; Watson, C.S. Non-Genomic Effects of Xenoestrogen Mixtures. Int. J. Environ. Res. Public Health 2012, 9, 2694-2714. https://doi.org/10.3390/ijerph9082694
Viñas R, Jeng Y-J, Watson CS. Non-Genomic Effects of Xenoestrogen Mixtures. International Journal of Environmental Research and Public Health. 2012; 9(8):2694-2714. https://doi.org/10.3390/ijerph9082694
Chicago/Turabian StyleViñas, René, Yow-Jiun Jeng, and Cheryl S. Watson. 2012. "Non-Genomic Effects of Xenoestrogen Mixtures" International Journal of Environmental Research and Public Health 9, no. 8: 2694-2714. https://doi.org/10.3390/ijerph9082694
APA StyleViñas, R., Jeng, Y.-J., & Watson, C. S. (2012). Non-Genomic Effects of Xenoestrogen Mixtures. International Journal of Environmental Research and Public Health, 9(8), 2694-2714. https://doi.org/10.3390/ijerph9082694