Elevated Oestrogen Receptor Splice Variant ERαΔ5 Expression in Tumour-adjacent Hormone-responsive Tissue


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Tissue Collection and Storage
2.3. Quantitative Real-time Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
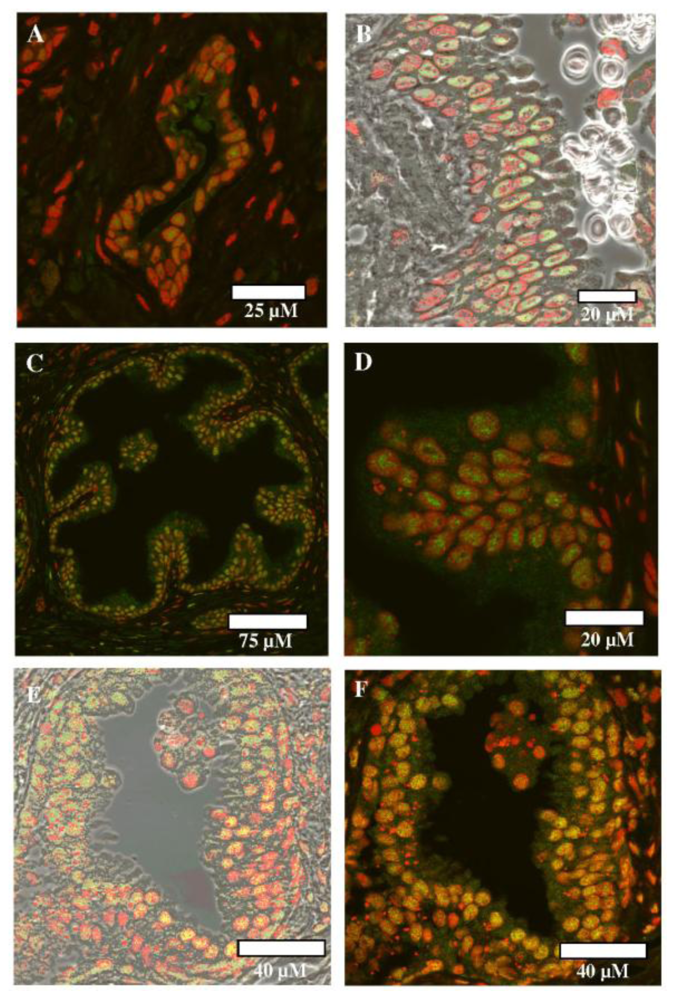
2.4. Immunofluorescence
3. Results
3.1. Proliferative Endometrium vs. Grade 2 Endometrioid TA Tissue
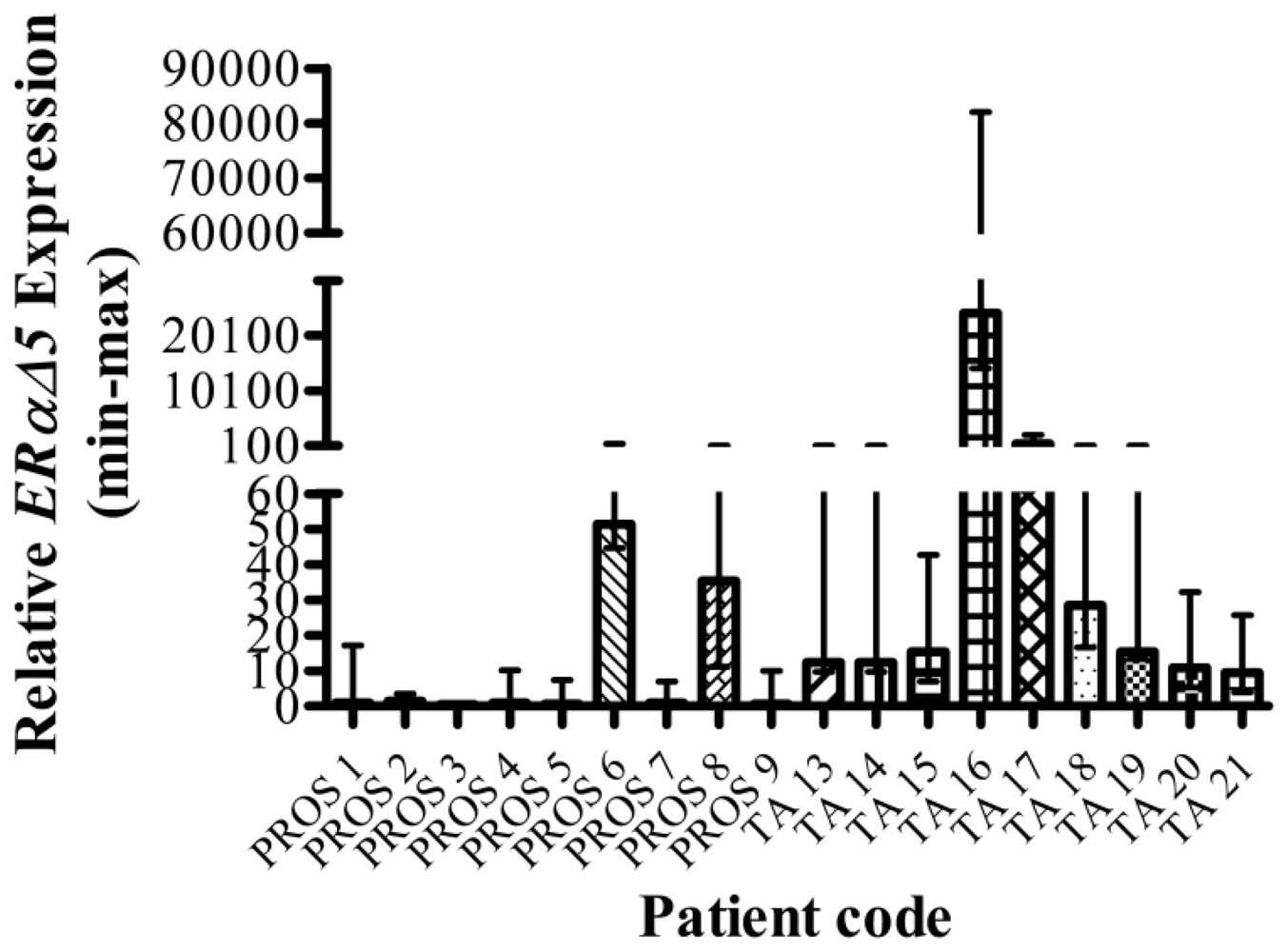
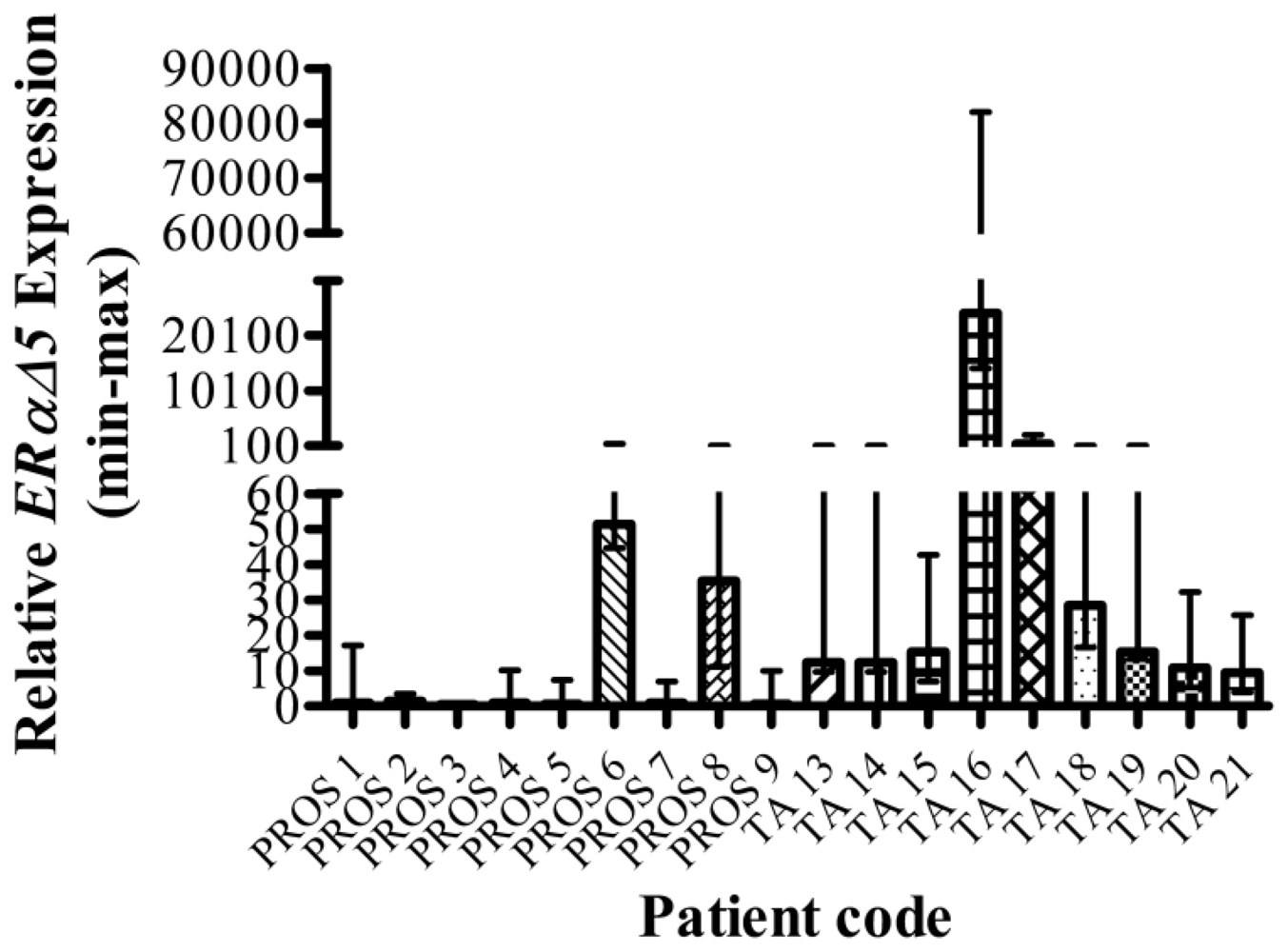
3.2. Benign Prostate vs. Tumour-adjacent Prostate
3.3. UK vs. India Benign Prostate
4. Discussion
Acknowledgements
Appendix

{kind=link}
{kind=link}
| Code | ERα | ERαΔ3 | ERαΔ5 | ERβ1 | ERβ2 | ERβ5 |
|---|---|---|---|---|---|---|
| N1 | 25.0 (19.2) | 30.6 (19.2) | 31.2 (19.2) | 33.9 (18.7) | 33.4 (18.7) | 37.0 (18.7) |
| N2 | 24.0 (19.2) | 30.0 (19.2) | 31.5 (19.2) | 32.8 (18.5) | 32.6 (18.5) | 35.1 (18.5) |
| N3 | 23.9 (19.4) | 28.6 (19.4) | 30.2 (19.4) | 34.1 (18.8) | 33.4 (18.8) | 36.9 (18.8) |
| TA1 | 24.7 (19.0) | 30.4 (19.0) | 31.9 (19.0) | 31.1 (18.7) | 32.9 (18.7) | 33.9 (18.7) |
| TA2 | 23.0 (17.7) | 28.3 (17.7) | 30.2 (17.7) | 32.4 (17.7) | 30.9 (17.7) | 34.6 (17.7) |
| TA3 | 24.4 (18.8) | 30.1 (18.8) | 31.2 (18.8) | 34.0 (18.8) | 32.7 (18.8) | 37.1 (18.8) |
| N4 | 26.4 (20.8) | 31.1 (20.8) | 31.5 (20.8) | 30.1 (19.4) | 33.5 (19.4) | 39.4 (19.4) |
| N5 | 29.0 (22.8) | 32.5 (22.8) | 33.7 (22.8) | 20.7 (21.3) | 34.5 (21.3) | 40.3 (21.3) |
| N6 | 26.0 (20.3) | 30.8 (20.3) | 30.8 (20.3) | 34.1 (19.0) | 33.7 (19.0) | 37.7 (19.0) |
| TA4 | 24.6 (20.2) | 29.6 (20.2) | 29.5 (20.2) | 24.7 (18.3) | 32.9 (18.3) | 34.3 (18.3) |
| TA5 | 24.3 (20.3) | 29.5 (20.3) | 29.8 (20.3) | 35.3 (18.6) | 33.4 (18.6) | 37.0 (18.6) |
| TA6 | 27.2 (22.7) | 32.2 (22.7) | 33.3 (22.7) | 37.2 (21.4) | 33.0 (21.4) | 38.2 (21.4) |
| N7 | 24.5 (19.0) | 29.4 (19.0) | 30.5 (19.0) | 33.1 (18.9) | 33.5 (18.9) | 37.2 (18.9) |
| N8 | 24.4 (19.1) | 30.6 (19.1) | 30.9 (19.1) | 34.2 (18.8) | 32.3 (18.8) | 58.7 (18.8) |
| N9 | 26.0 (19.9) | 31.0 (19.9) | 32.4 (19.9) | 36.1 (20.0) | 33.8 (20.0) | 39.2 (20.0) |
| TA7 | 24.0 (19.4) | 29.7 (19.4) | 31.3 (19.4) | 32.1 (19.1) | 32.5 (19.1) | 36.6 (19.1) |
| TA8 | 24.1 (18.5) | 30.2 (18.5) | 31.0 (18.5) | 32.8 (18.5) | 33.2 (18.5) | 37.2 (18.5) |
| TA9 | 23.2 (18.2) | 28.6 (18.2) | 30.0 (18.2) | 32.6 (17.9) | 31.8 (17.9) | 38.9 (17.9) |
| Code | ERα | ERαΔ3 | ERαΔ5 | ERβ1 | ERβ2 | ERβ5 |
|---|---|---|---|---|---|---|
| PROS 1 | 36.4 (23.9) | 39.5 (23.9) | 43.9 (23.9) | 37.0 (24.4) | 37.2 (24.4) | 57.1 (24.4) |
| PROS 2 | 34.4 (24.9) | 36.6 (24.9) | 44.4 (24.9) | 34.8 (23.3) | 35.0 (23.3) | 53.2 (23.3) |
| PROS 3 | 33.8 (20.7) | 40.1 (20.7) | 41.6 (20.7) | 36.5 (20.3) | 33.1 (20.3) | 37.5 (20.3) |
| TA 13 | 29.1 (19.7) | 35.2 (19.7) | 36.0 (19.7) | 36.3 (19.5) | 32.8 (19.5) | 38.6 (19.5) |
| TA 14 | 28.8 (19.0) | 34.5 (19.0) | 34.2 (19.0) | 35.4 (18.3) | 31.0 (18.3) | 36.4 (18.3) |
| TA 15 | 30.0 (21.0) | 35.3 (21.0) | 37.1 (21.0) | 36.5 (21.2) | 33.5 (21.2) | 38.8 (21.2) |
| PROS 4 | 41.3 (29.8) | 43.0 (29.8) | 56.1 (29.8) | Insuff | Insuff | Insuff |
| PROS 5 | 39.0 (26.3) | 48.2 (26.3) | 53.2 (26.3) | Insuff | Insuff | Insuff |
| PROS 6 | 37.7 (29.7) | 39.3 (29.7) | 50.3 (29.7) | Insuff | Insuff | Insuff |
| TA 16 | 32.6 (29.4) | 34.6 (29.4) | 41.1 (29.4) | Insuff | Insuff | Insuff |
| TA 17 | 37.3 (26.1) | 40.0 (26.1) | 43.8 (26.1) | Insuff | Insuff | Insuff |
| TA 18 | 37.7 (26.6) | 40.1 (26.6) | 48.1 (26.6) | Insuff | Insuff | Insuff |
| PROS 7 | 36.2 (25.8) | 38.6 (25.8) | 44.7 (25.8) | 37.2 (25.7) | 34.6 (25.7) | 56.0 (25.7) |
| PROS 8 | 31.6 (25.3) | 32.9 (25.3) | 39.1 (25.3) | 31.2 (25.2) | 31.7 (25.2) | 42.1 (25.2) |
| PROS 9 | 33.4 (21.9) | 36.5 (21.9) | 41.3 (21.9) | 35.0 (21.2) | 34.2 (21.2) | 38.5 (21.2) |
| TA14 19 | 33.0 (25.3) | 35.5 (25.3) | 40.4 (25.3) | 36.0 (25.5) | 23.3 (25.5) | 48.2 (25.5) |
| TA17 20 | 31.0 (21.7) | 33.5 (21.7) | 37.2 (21.7) | 32.6 (21.5) | 30.8 (21.5) | 38.4 (21.5) |
| TA21 21 | 32.1 (22.4) | 34.9 (22.4) | 38.2 (22.4) | Insuff | Insuff | Insuff |
| TA8 22 | Insuff | Insuff | Insuff | 33.8 (20.1) | 30.4 (20.1) | 36.8 (20.1) |
| Code | ERα | ERαΔ3 | ERαΔ5 | ERβ1 | ERβ2 | ERβ5 |
|---|---|---|---|---|---|---|
| PROS 10 | 39.2 (26.9) | 40.2 (26.9) | 41.5 (26.9) | 39.8 (25.7) | 35.8 (25.7) | 45.8 (25.7) |
| PROS 11 | 36.1 (24.7) | 39.1 (24.7) | 41.1 (24.7) | 38.4 (23.4) | 35.1 (23.4) | 40.6 (23.4) |
| PROS 12 | 33.9 (22.2) | 37.0 (22.2) | 38.2 (22.2) | 37.5 (21.1) | 33.2 (21.1) | 41.0 (21.1) |
| IND11 | Insuff | Insuff | Insuff | 37.8 (22.9) | 34.3 (22.9) | 45.2 (22.9) |
| IND12 | 34.2 (21.8) | 38.8 (21.8) | 38.3 (21.8) | 40.4 (21.6) | 35.6 (21.6) | 47.7 (21.6) |
| IND13 | Insuff | Insuff | Insuff | 33.8 (25.7) | 35.8 (25.7) | 43.3 (25.7) |
| IND14 | 34.6 (22.0) | 37.9 (22.0) | 41.6 (22.0) | Insuff | Insuff | Insuff |
| IND15 | 36.8 (27.0) | 38.2 (27.0) | 40.9 (27.0) | Insuff | Insuff | Insuff |
| PROS 7 | 35.7 (26.2) | 37.3 (26.2) | 45.4 (26.2) | 36.2 (26.1) | 48.7 (26.1) | 51.4 (26.1) |
| PROS 8 | 30.2 (25.9) | 32.8 (25.9) | 38.0 (25.9) | 30.8 (25.6) | 41.9 (25.6) | 41.6 (25.6) |
| PROS 9 | 32.9 (22.3) | 35.9 (22.3) | 39.0 (22.3) | 34.0 (22.6) | 50.0 (22.6) | 36.7 (22.6) |
| IND1 | 33.7 (23.1) | 39.1 (23.1) | 42.0 (23.1) | 39.6 (21.8) | 53.9 (21.8) | 55.1 (21.8) |
| IND2 | 35.7 (28.6) | 38.1 (28.6) | 42.4 (28.6) | 35.4 (26.9) | 48.1 (26.9) | 50.6 (26.9) |
| IND3 | 33.7 (24.9) | 35.9 (24.9) | ND | 33.0 (23.4) | 48.5 (23.4) | 50.5 (23.4) |
| PROS 1 | Insuff | Insuff | Insuff | 36.6 (23.8) | 37.8 (23.8) | 41.3 (23.8) |
| PROS 2 | Insuff | Insuff | Insuff | 34.4 (24.6) | 34.1 (24.6) | 43.2 (24.6) |
| PROS 3 | Insuff | Insuff | Insuff | 36.5 (20.5) | 33.6 (20.5) | 40.6 (20.5) |
| IND4 | Insuff | Insuff | Insuff | 40.3 (23.4) | 37.1 (23.4) | 46.2 (23.4) |
| IND5 | Insuff | Insuff | Insuff | ND | 46.0 (30.0) | 44.4 (30.0) |
| IND6 | Insuff | Insuff | Insuff | 32.1 (23.6) | 34.4 (23.6) | 38.9 (23.6) |
| PROS 4 | Insuff | Insuff | Insuff | 38.1 (27.3) | 37.3 (27.3) | 44.4 (27.3) |
| PROS 5 | Insuff | Insuff | Insuff | 38.7 (23.2) | 36.2 (23.2) | 45.9 (23.2) |
| PROS 6 | Insuff | Insuff | Insuff | 35.3 (27.8) | 36.4 (27.8) | 39.8 (27.8) |
| IND7 | Insuff | Insuff | Insuff | 32.4 (25.3) | 35.4 (25.3) | 39.8 (25.3) |
| IND8 | Insuff | Insuff | Insuff | 37.2 (21.0) | 33.4 (21.0) | 37.0 (21.0) |
| IND10 | Insuff | Insuff | Insuff | 38.8 (23.4) | 35.0 (23.4) | 47.6 (23.4) |
| Code | Age (y) | PSA (ng/mL) | Gleason grade |
|---|---|---|---|
| PROS 1 | 71 | 3.75 | NA |
| PROS 2 | 81 | 4.99 | NA |
| PROS 3 | 72 | NK | NA |
| PROS 4 | 72 | 4.96 | NA |
| PROS 5 | 73 | 2.49 | NA |
| PROS 6 | 74 | 6.25 | NA |
| PROS 7 | 81 | 5 | NA |
| PROS 8 | 62 | 5.7 | NA |
| PROS 9 (open) | 82 | 34 | NA |
| PROS 10 | 79 | 3.75 | NA |
| PROS 11 | 71 | 4.19 | NA |
| PROS 12 (open) | 73 | 5 | NA |
| TA 13 | 60 | 11.5 | 3 + 3 |
| TA 14 | 61 | 8.2 | 3 + 3 |
| TA 15 | 64 | 8.7 | 4 + 4 |
| TA 16 | 65 | 6.5 | 3 + 3 |
| TA 17 | 56 | 2.7 | benign |
| TA 18 | 67 | NK | benign |
| TA 19 | NK | NK | NK |
| TA 20 | 66 | 9 | 3 + 3 |
| TA 21 | 57 | 5.3 | 3 + 3 |
| TA 22 | 63 | 5.8 | 3 + 4 |
| IND 1 | 58 | 1.2 | NA |
| IND 2 | 60 | 2.4 | NA |
| IND 3 | 63 | 0.3 | NA |
| IND 4 | 76 | 3.2 | NA |
| IND 5 | 75 | 0.3 | NA |
| IND 6 | 65 | 0.2 | NA |
| IND 7 | 79 | 2.5 | NA |
| IND 8 | 74 | 0.3 | NA |
| IND 9 | 65 | 0.6 | NA |
| IND 10 | NK | NK | NA |
| IND 11 | 62 | 3.3 | NA |
| IND 12 | 47 | 2.4 | NA |
| IND 13 | 64 | 2.4 | NA |
| IND 14 | 60 | 0.3 | NA |
| IND 15 | 74 | 1.4 | NA |
| Code | Age (y) | Histology | Stage | |||
|---|---|---|---|---|---|---|
| N1 | 39 | proliferative | NA | |||
| N2 | 46 | early proliferative | NA | |||
| N3 | 42 | proliferative | NA | |||
| N4 | 39 | proliferative | NA | |||
| N5 | 46 | proliferative | NA | |||
| N6 | 51 | proliferative | NA | |||
| N7 | 42 | proliferative | NA | |||
| N8 | 43 | proliferative to early secretory with simple hyperplasia | NA | |||
| N9 | 43 | proliferative | NA | |||
| TA1 | 58 | G2 endometrioid | 3a | |||
| TA2 | 74 | G2 endometrioid | 1c | |||
| TA3 | 67 | G2 endometrioid | 1b | |||
| TA4 | 84 | G2 endometrioid | 1a | |||
| TA5 | 57 | G2 endometrioid | 1a | |||
| TA6 | 77 | G2 endometrioid | 1c | |||
| TA7 | 70 | G2 endometrioid | 1b | |||
| TA8 | 62 | G2 endometrioid | 1b | |||
| TA9 | 62 | G2 endometrioid | 2a | |||
Abbreviations
| C | calibrator control |
| CaP | prostate cancer |
| CT | threshold cycle |
| ER | oestrogen receptor |
| GSPBS | 5% normal goat serum in PBS |
| PI | propidium iodide |
| PMT | photomultiplier tubes |
| PROS | benign prostate tissues |
| PSA | prostate-specific antigen |
| RRP | radical retro-pubic prostatectomy |
| RT-PCR | real time polymerase chain reaction |
| TA | tumour-adjacent |
| TURP | trans-urethral resection of the prostate |
References
- Whittemore, AS; Kolonel, LN; Wu, AH; John, EM; Gallagher, RP; Howe, GR; Burch, JD; Hankin, J; Dreon, DM; West, DW; Teh, C-Z; Paffenbarger, RS, Jr. Prostate cancer in relation to diet, physical activity, and body size in Blacks, Whites, and Asians in the United States and Canada. J. Natl. Cancer Inst 1995, 87, 652–661. [Google Scholar]
- Dey, S; Hablas, A; Seifeldin, IA; Ismail, K; Ramadan, M; El-Hamzawy, H; Wilson, ML; Banerjee, M; Boffetta, P; Harford, J; Merajver, SD; Soliman, AS. Urban-rural differences of gynaecological malignancies in Egypt (1999–2002). BJOG 2010, 117, 348–355. [Google Scholar]
- Yang, C-Y; Hsieh, Y-L. The Relationship between Population Density and Cancer Mortality in Taiwan. Jpn. J. Cancer Res 1998, 89, 355–360. [Google Scholar]
- Khan, N; Afaq, F; Mukhtar, H. Lifestyle as risk factor for cancer: Evidence from human studies. Cancer Lett 2010, 293, 133–143. [Google Scholar]
- Rieck, G; Fiander, A. The effect of lifestyle factors on gynaecological cancer. Best Pract. Res. Clin. Obstet. Gynaecol 2006, 20, 227–251. [Google Scholar]
- Sorosky, J. Endometrial Cancer. Obstet. Gynecol 2008, 111, 436–477. [Google Scholar]
- Singh, PB; Matanhelia, SS; Martin, FL. A potential paradox in prostate adenocarcinoma progression: Oestrogen as the initiating driver. Eur. J. Cancer 2008, 44, 928–936. [Google Scholar]
- Setlur, SR; Mertz, KD; Hoshida, Y; Demichelis, F; Lupien, M; Perner, S; Sboner, A; Pawitan, Y; Andren, O; Johnson, LA; Tang, J; Adami, H-O; Calza, S; Chinnaiyan, AM; Rhodes, D; Tomlins, S; Fall, K; Mucci, LA; Kantoff, PW; Stampfer, MJ; Andersson, S-O; Varenhorst, E; Johansson, J-E; Brown, M; Golub, TR; Rubin, MA. Estrogen-dependent signaling in a molecularly distinct subclass of aggressive prostate cancer. J. Natl. Cancer Inst 2008, 100, 815–825. [Google Scholar]
- Simpson, ER; Clyne, C; Rubin, G; Boon, WC; Robertson, K; Britt, K; Speed, C; Jones, M. Aromatase - a brief overview. Annu. Rev. Physiol 2002, 64, 93–127. [Google Scholar]
- Taylor, SE; Martin-Hirsch, PL; Martin, FL. Oestrogen receptor splice variants in the pathogenesis of disease. Cancer Lett 2010, 288, 133–148. [Google Scholar]
- Koduri, S; Goldhar, AS; Vonderhaar, BK. Activation of vascular endothelial growth factor (VEGF) by the ER-α variant, ERΔ3. Breast Cancer Res. Treat 2006, 95, 37–43. [Google Scholar]
- Bollig, A; Miksicek, RJ. An estrogen receptor-α splicing variant mediates both positive and negative effects on gene transcription. Mol. Endocrinol 2000, 14, 634–649. [Google Scholar]
- Leung, Y-K; Mak, P; Hassan, S; Ho, S-M. Estrogen receptor (ER)-β isoforms: a key to understanding ER-β signalling. Proc. Natl. Acad. Sci. USA 2006, 103, 13162–13167. [Google Scholar]
- Girault, I; Andrieu, C; Tozlu, S; Spyratos, F; Bièche, I; Lidereau, R. Altered expression pattern of alternatively spliced estrogen receptor β transcripts in breast carcinoma. Cancer Lett 2004, 215, 101–112. [Google Scholar]
- Poola, I; Abraham, J; Baldwin, K; Saunders, A; Bhatnagar, R. Estrogen receptors beta4 and beta5 are full length functionally distinct ERβ isoforms. Endocrine 2005, 27, 227–238. [Google Scholar]
- Rey, JM; Pujol, P; Dechaud, H; Edouard, E; Hedon, B; Maudelonde, T. Expression of oestrogen receptor-α splicing variants and oestrogen receptor-β in endometrium of infertile patients. Mol. Hum. Reprod 1998, 4, 641–647. [Google Scholar]
- Marshburn, PB; Zhang, J; Bahrani–Mostafavi, Z; Mostafavi, BZ; Marroum, M-C; Mougeot, J-LC; Roshon, MJ. Estrogen receptor-α messenger RNA variants that lack exon 5 or exon 7 are coexpressed with wild-type form in human endometrium during all phases of the menstrual cycle. Am. J. Obstet Gynecol 2004, 191, 626–633. [Google Scholar]
- Skrzypczak, M; Bieche, I; Szymczak, S; Tozlu, S; Lewan Ddowowski, S; Girault, I; Radwanska, K; Szczylik, C; Jakowicki, J; LidereauI, R; Kaczmarek, L. Evaluation of mRNA expression of estrogen receptor β and its isoforms in human normal and neoplastic endometrium. Int. J. Cancer 2004, 110, 783–787. [Google Scholar]
- Leung, Y-K; Lam, H-M; Wu, S; Song, D; Levin, L; Cheng, L; Wu, C-L; Ho, S-M. Estrogen receptor β2 and β5 are associated with poor prognosis in prostate cancer, and promote cancer cell migration and invasion. Endocr. Relat. Cancer 2010, 17, 675–689. [Google Scholar]
- Singh, MN; Stringfellow, HF; Taylor, SE; Ashton, KM; Ahmad, M; Abdo, KR; El-Agnaf, OMA; Martin-Hirsch, PL; Martin, FL. Elevated expression of CYP1A1 and γ-SYNUCLEIN in human ectopic (ovarian) endometriosis compared with eutopic endometrium. Mol. Hum. Reprod 2008, 14, 655–663. [Google Scholar]
- Singh, PB; Ragavan, N; Ashton, KM; Basu, P; Nadeem, SM; Nicholson, CM; Krishna, RKG; Matanhelia, SS; Martin, FL. Quantified gene expression levels for phase I/II metabolizing enzyme and estrogen receptor levels in benign prostate from cohorts designated as high-risk (UK) versus low-risk (India) for adenocarcinoma at this organ site: a preliminary study. Asian J. Androl 2010, 12, 203–214. [Google Scholar]
- Witek, A; Paul-Samojedny, M; Stojko, R; Seifert, B; Mazurek, U. Coexpression index of estrogen receptor alpha mRNA isoforms in simple, complex hyperplasia without atypia, complex atypical hyperplasia and adenocarcinoma. Gynecol. Oncol 2007, 106, 407–412. [Google Scholar]
- Yared, E; McMillan, TJ; Martin, FL. Genotoxic effects of oestrogens in breast cells detected by the micronucleus assay and the Comet assay. Mutagenesis 2002, 17, 345–352. [Google Scholar]
- John, K; Ragavan, N; Pratt, MM; Singh, PB; Al-Buheissi, S; Matanhelia, SS; Phillips, DH; Poirier, MC; Martin, FL. Quantification of phase I/II metabolizing enzyme gene expression and polycyclic aromatic hydrocarbon–DNA adduct levels in human prostate. Prostate 2009, 69, 505–519. [Google Scholar]
- Martin, FL; Patel, II; Sozeri, O; Singh, PB; Ragavan, N; Nicholson, CM; Frei, E; Meinl, W; Glatt, H; Phillips, DH; Arlt, VM. Constitutive expression of bioactivating enzymes in normal human prostate suggests a capability to activate pro-carcinogens to DNA-damaging metabolites. Prostate 2010, 70, 1586–1599. [Google Scholar]
- Martin, FL; Kelly, JG; Llabjani, V; Martin–Hirsch, PL; Patel, II; Trevisan, J; Fullwood, NJ; Walsh, MJ. Distinguishing cell types or populations based on the computational analysis of their infrared spectra. Nat. Protoc 2010, 5, 1748–1760. [Google Scholar]
| Gene Symbol | Primer | Sequence (5′–3′) |
|---|---|---|
| ERα | ERα-F | TGG ACA GGA ACC AGG GAA AAT |
| ERα-R | GAG ATG ATG TAG CCA GCA GCA T | |
| ERαΔ3 | ERαΔ3-F | AGA AGT ATT CAA GGG ATA CGA AAA G |
| ERαΔ3-R | ATC ATC TCT CTG GCG CTT GT | |
| ERαΔ5 | ERαΔ5-F | AGG GTG CCA GGA ACC A |
| ERαΔ5-R | GAT GTA GCC AGC AGC ATG TC | |
| ERβ1 | ERβ-F | TGT AAA CAG AGA GAC ACT GAA AAG GAA |
| ERβ-R | CCT CTT TGA ACC TGG ACC AGT AA | |
| ERβ2 | ERβ2-F | GCA TGC GAG GGC AGA A |
| ERβ2-R | TTC TTT AGG CCA CCG AGT TG | |
| ERβ5 | ERβ5-F | CAC GGA GGG TGA AGT GAT TT |
| ERβ5-R | ATT CCA AAT GAG GCA TTC ATT | |
| β-ACTIN | β-ACTIN-F | CCT GGC ACC CAG CAC AAT |
| β-ACTIN-R | GCC GAT CCA CAC GGA GTA CT | |
| Patient code | ERα | ERαΔ3 | ERαΔ5 | ERβ1 | ERβ2 | ERβ5 |
|---|---|---|---|---|---|---|
| N 1 | 1 (c)* (0.69–1.46) | 1 (c) (0.71–1.41) | 1 (c) (0.71–1.42) | 1 (c) (0.71–1.41) | 1 (c) (0.64–1.57) | 1 (c) (0.50–2.02) |
| N 2 | 1.06 (0.92–1.23) | 1.54 (1.32–1.81) | 0.83 (0.72–0.96) | 1.95 (1.28–2.96) | 1.53 (0.95–2.46) | 3.41 (2.58–4.51) |
| N 3 | 2.29 (1.68–3.13) | 4.31 (3.11–5.97) | 2.24 (1.71–2.93) | 0.90 (0.52–1.58) | 1.08 (0.65–1.81) | 1.15 (0.62–2.16) |
| TA 1 | 1.07 (0.77–1.49) | 1.02 (0.78–1.33) | 0.55 (0.40–0.75) | 7.36 (4.75–11.41) | 1.47 (0.99–2.19) | 8.90 (6.53–12.13) |
| TA 2 | 1.43 (1.29–1.58) | 1.77 (1.54–2.02) | 0.73 (0.65–0.83) | 1.44 (1.01–2.05) | 2.82 (1.88–4.24) | 2.64 (1.61–4.33) |
| TA 3 | 1.12 (0.88–1.43) | 1.03 (0.82–1.29) | 0.74 (0.52–1.04) | 0.98 (0.79–1.22) | 1.77 (1.34–2.33) | 1.00 (0.59–1.69) |
| N 4 | 1 (c) (0.91–1.10) | 1 (c) (0.88–1.14) | 1 (c) (0.88–1.14) | 1 (c) (0.96–1.05) | 1 (c) (0.70–1.43) | 1 (c) (0.74–1.34) |
| N 5 | 0.61 (0.49–0.76) | 1.42 (1.08–1.87) | 0.85 (0.59–1.23) | 2.32 (2.10–2.57) | 1.82 (1.32–2.49) | 2.06 (0.43–9.80) |
| N 6 | 0.93 (0.72–1.20) | 0.87 (0.53–1.42) | 1.14 (0.85–1.54) | 0.05 (0.03–0.06) | 0.62 (0.31–1.27) | 2.56 (1.36–4.82) |
| TA 4 | 2.13 (1.91–2.38) | 1.77 (1.62–1.94) | 2.49 (2.19–2.82) | 0.02 (0.01–0.03) | 0.70 (0.38–1.31) | 16.19 (10.38–25.23) |
| TA 5 | 2.88 (2.56–3.25) | 2.13 (1.89–2.40) | 2.24 (1.88–2.65) | 0.01 (0.01–0.02) | 0.57 (0.43–0.76) | 2.92 (1.28–4.28) |
| TA 6 | 2.16 (1.88–2.48) | 1.73 (1.49–2.02) | 1.11 (0.86–1.44) | 0.03 (0.02–0.05) | 5.70 (4.08–7.96) | 9.76 (4.02–23.71) |
| N 7 | 1 (c) (0.53–1.88) | 1 (c) (0.54–1.87) | 1 (c) (0.45–2.22) | 1 (c) (0.62–1.61) | 1 (c) (0.49–2.06) | 1 (c) (0.39–2.59) |
| N 8 | 1.12 (0.64–1.97) | 0.49 (0.29–0.85) | 0.80 (0.39–1.67) | 0.41 (0.23–0.75) | 2.15 (1.36–3.41) | NQ |
| N 9 | 0.66 (0.48–0.91) | 0.63 (0.45–0.86) | 0.49 (0.29–0.83) | 0.26 (0.16–0.44) | 1.69 (0.84–3.42) | 0.51 (0.16–1.63) |
| TA 7 | 1.80 (1.34–2.40) | 1.15 (0.78–1.69) | 0.78 (0.59–1.03) | 2.37 (1.79–3.13) | 2.42 (1.88–3.12) | 1.77 (1.01–3.08) |
| TA 8 | 0.98 (0.68–1.40) | 0.42 (0.32–0.56) | 0.51 (0.37–0.71) | 0.87 (0.62–1.22) | 0.91 (0.55–1.51) | 0.76 (0.50–1.17) |
| TA 9 | 1.37 (0.99–1.89) | 1.05 (0.81–1.34) | 0.79 (0.62–1.00) | 0.68 (0.41–1.13) | 1.59 (0.76–3.32) | 0.15 (0.003–7.94) |
| Patient code | ERα | ERαΔ3 | ERαΔ5 | ERβ1 | ERβ2 | ERβ5 |
|---|---|---|---|---|---|---|
| PROS 1 | 1 (c)* (0.71–1.41) | 1 (c) (0.25–4.10) | 1 (c) (0.06–16.2) | 5.95 (5.65–6.27) | 0.51 (0.36–0.71) | ND |
| PROS 2 | 8.04 (5.59–11.56) | 15.6 (8.64–28.17) | 1.41 (0.95–2.10) | 57.95 (47.1–71.4) | 5.04 (3.62–7.03) | ND |
| PROS 3 | 0.70 (0.52–0.93) | 0.08 (0.02–0.29) | 0.56 (0.06–5.63) | 1 (c) (0.58–1.72) | 1 (c) (0.87–1.15) | 1 (c) (0.78–1.29) |
| TA 13 | 8.13 (3.15–21.0) | 1.07 (0.60–1.91) | 12.38 (2.75–55.78) | 0.65 (0.54–0.78) | 0.74 (0.29–1.9) | 0.28 (0.23–0.33) |
| TA 14 | 6.59 (3.37–12.88) | 1.13 (0.60–2.13) | 28.44 (13.79–58.66) | 0.54 (0.12–2.49) | 1.18 (0.22–6.52) | 0.59 (0.11–3.08) |
| TA 15 | 10.9 (7.15–16.6) | 2.57 (1.61–4.10) | 15.4 (8.40–27.27) | 1.81 (0.86–3.82) | 1.44 (0.77–2.69) | 0.78 (0.23–2.61) |
| PROS 4 | 1 (c) (0.24–4.25) | 1 (c) (0.31–3.22) | 1 (c) (0.11–9.14) | Insuff | Insuff | Insuff |
| PROS 5 | 0.44 (0.08–2.43) | 0.002 (0.001–0.007) | 0.63 (0.06–6.76) | Insuff | Insuff | Insuff |
| PROS 6 | 11.39 (3.58–36.29) | 11.69 (3.72–36.73) | 51.4 (6.78–389.6) | Insuff | Insuff | Insuff |
| TA 16 | 300.3 (202.4–445.4) | 250.2 (180.3–347.1) | 24,154.4 (10,085.4–57,849.9) | Insuff | Insuff | Insuff |
| TA 17 | 1.26 (0.86–1.86) | 0.61 (0.3–1.27) | 385.79 (88.89–1,674.4) | Insuff | Insuff | Insuff |
| TA 18 | 1.35 (0.46–3.96) | 0.81 (0.3–2.21) | 28.4 (11.9–68) | Insuff | Insuff | Insuff |
| PROS 7 | 1 (c) (0.65–1.54) | 1 (c) (0.56–1.8) | 1 (c) (0.17–5.91) | 4.94 (3.49–6.98) | 18.24 (15.1–22.12) | ND |
| PROS 8 | 17.39 (13.87–21.8) | 37.01 (30.5–44.93) | 35.34 (24.23–51.56) | 227.5 (207.9–249.0) | 95.56 (69.65–131.1) | 1.35 (0.31–5.93) |
| PROS 9 | 0.48 (0.26–0.89) | 0.29 (0.12–0.68) | 0.7 (0.05–9.21) | 1 (c) (0.79–1.27) | 1 (c) (0.54–1.86) | 1 (c) (0.64–1.55) |
| TA 19 | 6.76 (2.83–16.14) | 6.53 (2.79–15.3) | 15.31 (2.02–116.23) | 10.13 (7.19–14.26) | 77.62 (28.89–208.6) | 0.02 (0.003–0.21) |
| TA 20 | 2.1 (1.83–2.42) | 2.07 (1.5–2.86) | 10.85 (5.53–21.29) | 6.04 (3.58–10.18) | 13.01 (6.59–25.68) | 1.28 (0.31–5.23) |
| TA 21 | 1.67 (1.5–1.86) | 1.29 (0.81–2.06) | 9.47 (5.43–16.53) | Insuff | Insuff | Insuff |
| TA 22 | Insuff | Insuff | Insuff | 1.07 (0.81–1.41) | 6.58 (5.18–8.36) | 1.41 (0.68–2.93) |
| Patient code | ERα | ERαΔ3 | ERαΔ5 | ERβ1 | ERβ2 | ERβ5 |
|---|---|---|---|---|---|---|
| PROS 10 | 1 (c)* (0.2–4.93) | 1 (c) (0.15–6.88) | 1 (c) (0.57–1.74) | 1 (c) (0.74–1.36) | 1 (c) (0.48–2.09) | 1 (c) (0.47–2.13) |
| PROS 11 | 1.86 (1.31–2.64) | 0.45 (0.22–0.95) | 0.3 (0.15–0.59) | 0.54 (0.39–0.74) | 0.33 (0.2–0.55) | 8.16 (2.0–33.22) |
| PROS 12 | 1.40 (0.87–2.24) | 0.34 (0.26–0.44) | 0.35 (0.26–0.48) | 0.21 (0.1–0.42) | 0.26 (0.13–0.55) | 1.22 (0.17–8.8) |
| IND 11 | Insuff | Insuff | Insuff | 0.58 (0.21–1.65) | 0.42 (0.22–0.8) | 0.23 (0.01–6.1) |
| IND 12 | 0.82 (0.58–1.18) | 0.07 (0.02–0.3) | 0.26 (0.2–0.34) | 0.04 (0.02–0.06) | 0.07 (0.05–0.1) | 0.02 (0.01–0.03) |
| IND 13 | Insuff | Insuff | Insuff | 65.12 (11.2–375.4) | 0.99 (0.16–6.24) | 6.12 (0.98–38.19) |
| IND 14 | 0.79 (0.61–1.04) | 0.16 (0.1–0.26) | 0.03 (0.02–0.05) | Insuff | Insuff | Insuff |
| IND 15 | 5.54 (3.53–8.69) | 4.23 (2.91–6.16) | 1.58 (0.19–12.85) | Insuff | Insuff | Insuff |
| PROS 7 | 1 (c) (0.63–1.59) | 1 (c) (0.78–1.28) | 1 (c) (0.60–1.67) | 1 (c) (0.53–1.88) | 1 (c) (0.35–2.83) | 1 (c) (0.45–2.23) |
| PROS 8 | 36.59 (31.22–42.88) | 19.12 (16.01–22.82) | 142.7 (86.53–235.3) | 29.24 (19.35–44.2) | 78.43 (21.56–285.3) | 654.8 (61.56–6,966.2) |
| PROS 9 | 0.48 (0.31–0.74) | 0.18 (0.13–0.25) | 5.74 (1.11–29.65) | 0.41 (0.25–0.67) | 0.04 (0.02–0.09) | 2,341.7(1,122.7–4,884.3) |
| IND 1 | 0.44 (0.31–0.62) | 0.03 (0.01–0.09) | 1.19 (0.14–10.4) | 0.005 (0.004–0.007) | 0.001 (0–0.011) | 0.004 (0.003–0.005) |
| IND 2 | 5.27 (2.79–9.95) | 3.09 (1.81–5.26) | ND | 3.0 (2.16–4.16) | 2.69 (0.47–15.37) | 3.07 (0.57–16.45) |
| IND 3 | 1.53 (1.22–1.92) | 1.11 (0.62–1.99) | 3.23 (1.61–6.5) | 1.33 (0.81–2.16) | 0.18 (0.004–8.7) | 0.29 (0.01–6.63) |
| PROS 1 | Insuff | Insuff | Insuff | 1 (c) (0.6–1.67) | 1 (c) (0.1–10.14) | 1 (c) (0.19–5.37) |
| PROS 2 | Insuff | Insuff | Insuff | 8.46 (7.15–10.01) | 23.5 (21.1–26.17) | 0.45 (0.1–2.04) |
| PROS 3 | Insuff | Insuff | Insuff | 0.12 (0.08–0.17) | 1.96 (1.4–2.75) | 0.16 (0.02–1.71) |
| IND 4 | Insuff | Insuff | Insuff | 0.23 (0.1–0.54) | 4.79 (3.62–6.34) | 0.09 (0.06–0.13) |
| IND 5 | Insuff | Insuff | Insuff | ND | 0.25 (0.03–2.24) | 8.61 (2.59–28.69) |
| IND 6 | Insuff | Insuff | Insuff | 20.02 (15.34–26.12) | 9.47 (5.66–15.85) | 4.37 (2.59–7.35) |
| PROS 4 | Insuff | Insuff | Insuff | 1 (c) (0.67–1.49) | 1 (c) (0.43–2.33) | 1 (c) (0.19–5.32) |
| PROS 5 | Insuff | Insuff | Insuff | 0.04 (0.02–0.08) | 0.13 (0.09–0.17) | 0.02 (0.02–0.02) |
| PROS 6 | Insuff | Insuff | Insuff | 10.15 (4.1–25.11) | 2.87 (1.31–6.26) | 35.7 (15.2–83.8) |
| IND 7 | Insuff | Insuff | Insuff | 13.64 (10.48–17.77) | 0.99 (0.66–1.49) | 6.26 (1.06–37.0) |
| IND 8 | Insuff | Insuff | Insuff | 0.02 (0.02–0.03) | 0.2 (0.14–0.29) | 2.26 (1.31–3.89) |
| IND 10 | Insuff | Insuff | Insuff | 0.04 (0.01–0.13) | 0.33 (0.19–0.56) | 0.01 (0.002–0.03) |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Taylor, S.E.; Patel, I.I.; Singh, P.B.; Nicholson, C.M.; Stringfellow, H.F.; Krishna, R.K.G.; Matanhelia, S.S.; Martin-Hirsch, P.L.; Martin, F.L. Elevated Oestrogen Receptor Splice Variant ERαΔ5 Expression in Tumour-adjacent Hormone-responsive Tissue. Int. J. Environ. Res. Public Health 2010, 7, 3871-3889. https://doi.org/10.3390/ijerph7113871
Taylor SE, Patel II, Singh PB, Nicholson CM, Stringfellow HF, Krishna RKG, Matanhelia SS, Martin-Hirsch PL, Martin FL. Elevated Oestrogen Receptor Splice Variant ERαΔ5 Expression in Tumour-adjacent Hormone-responsive Tissue. International Journal of Environmental Research and Public Health. 2010; 7(11):3871-3889. https://doi.org/10.3390/ijerph7113871
Chicago/Turabian StyleTaylor, Siân E., Imran I. Patel, Paras B. Singh, Caroline M. Nicholson, Helen F. Stringfellow, R. K. Gopala Krishna, Shyam S. Matanhelia, Pierre L. Martin-Hirsch, and Francis L. Martin. 2010. "Elevated Oestrogen Receptor Splice Variant ERαΔ5 Expression in Tumour-adjacent Hormone-responsive Tissue" International Journal of Environmental Research and Public Health 7, no. 11: 3871-3889. https://doi.org/10.3390/ijerph7113871
APA StyleTaylor, S. E., Patel, I. I., Singh, P. B., Nicholson, C. M., Stringfellow, H. F., Krishna, R. K. G., Matanhelia, S. S., Martin-Hirsch, P. L., & Martin, F. L. (2010). Elevated Oestrogen Receptor Splice Variant ERαΔ5 Expression in Tumour-adjacent Hormone-responsive Tissue. International Journal of Environmental Research and Public Health, 7(11), 3871-3889. https://doi.org/10.3390/ijerph7113871