
Effects of Voluntary Wheel Running Exercise on Chemotherapy-Impaired Cognitive and Motor Performance in Mice

 , , and
, , and 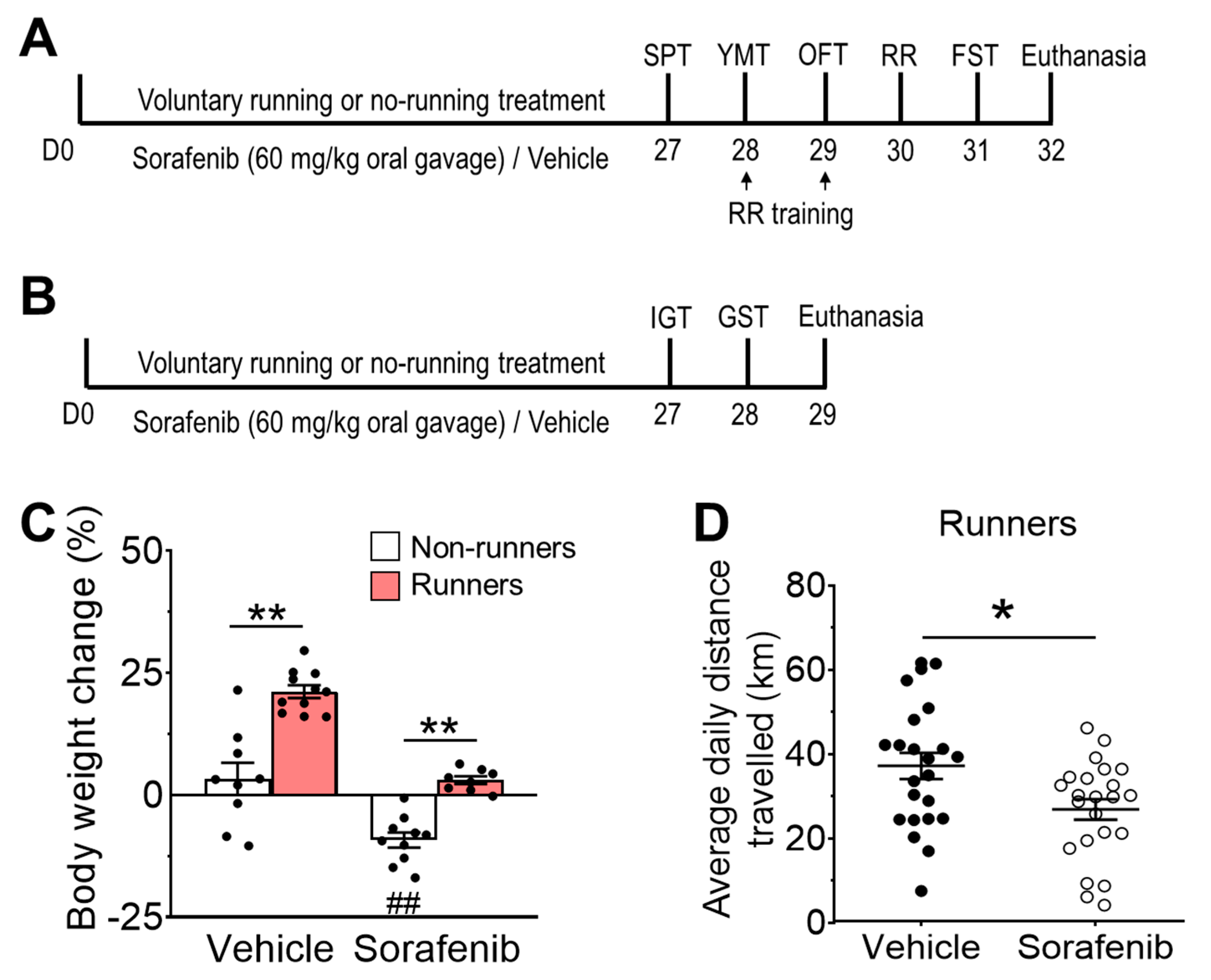{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Drug Preparation and Treatment
2.3. Voluntary Wheel Running Exercise
2.4. Behavioural Assessments
2.4.1. Y-Maze Task (YMT)
2.4.2. Sucrose Preference Test (SPT)
2.4.3. Open Field Test (OFT)
2.4.4. Forced Swim Test (FST)
2.4.5. Rotarod Test (RR)
2.5. Grip Strength Test (GST)
2.6. Inverted Grid Hanging Test (IGT)
2.7. Immunohistochemistry
2.8. Quantifications of Ki-67 and DCX-Immunopositive Cells
2.9. Isolation of Skeletal Muscle Tissues
2.10. Statistical Analyses
3. Results
3.1. Voluntary Running Exercise Attenuated Sorafenib-Induced Weight Loss
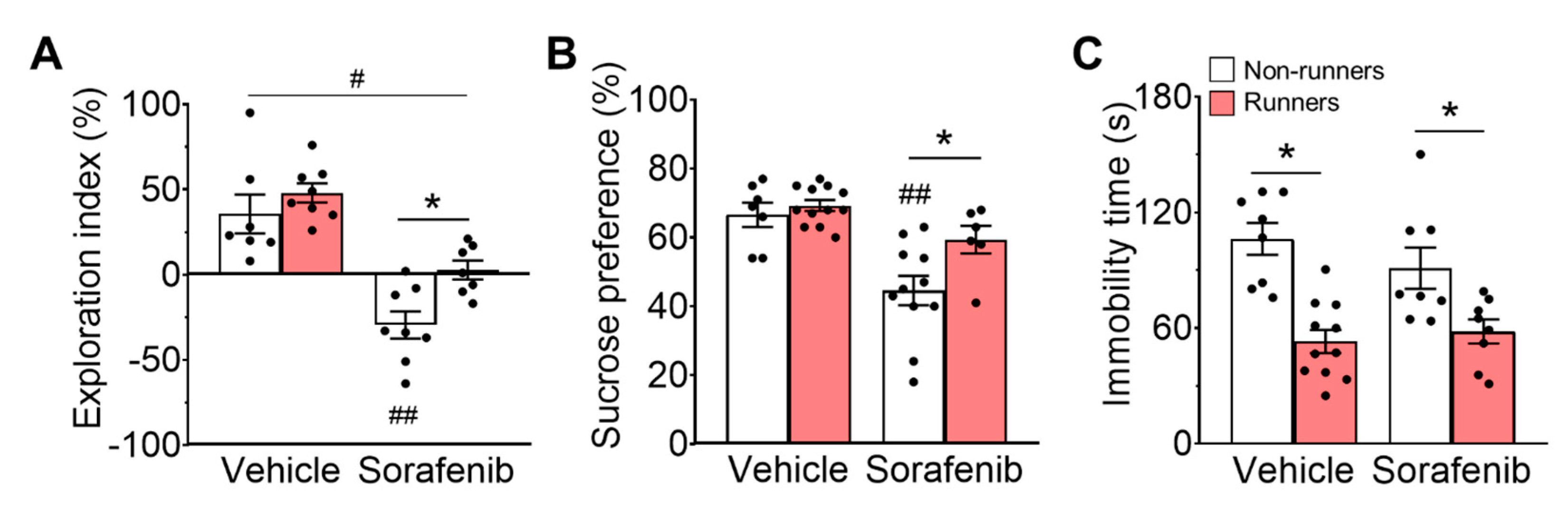
3.2. Sorafenib Administration Impaired Spatial Memory
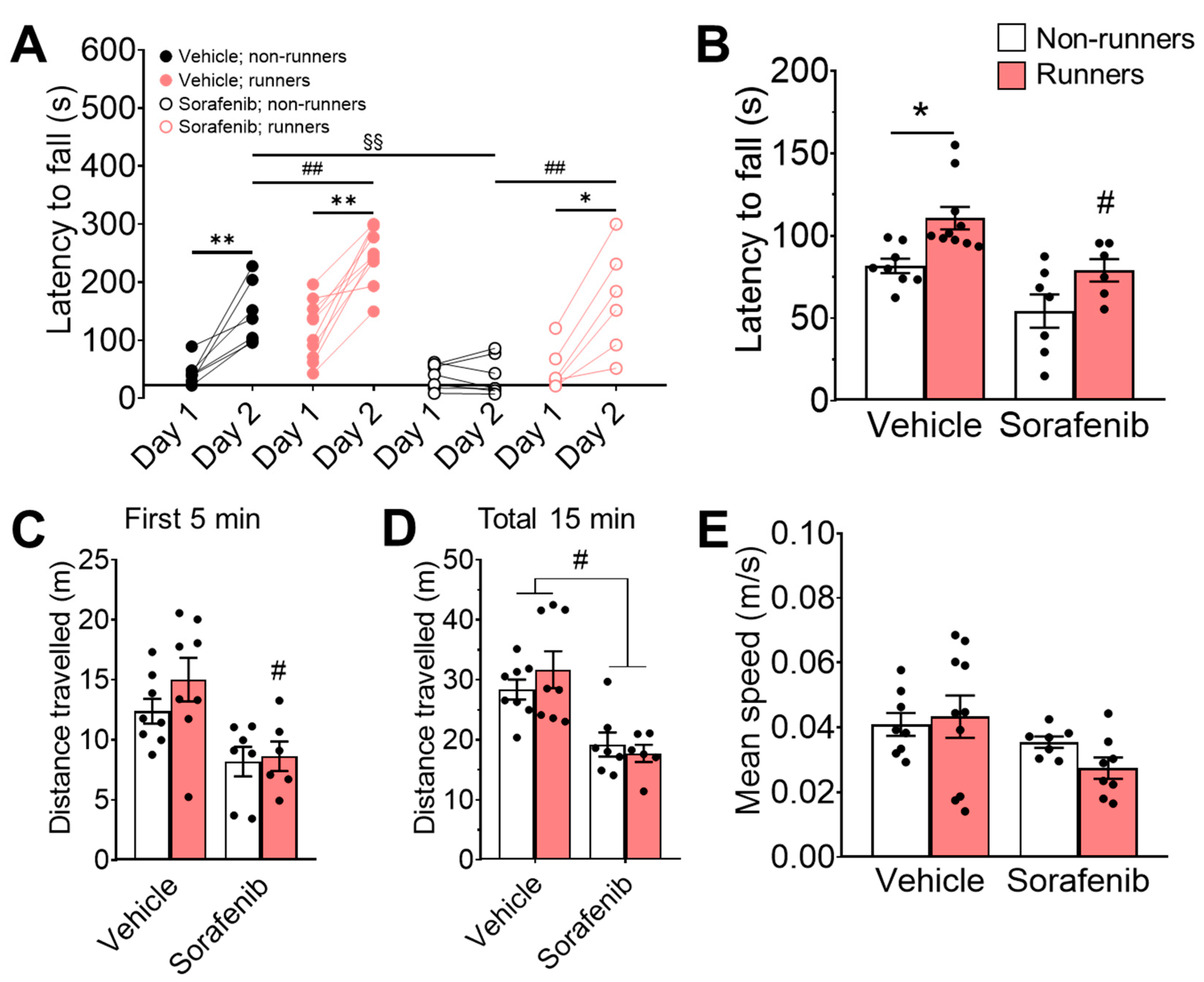
3.3. Voluntary Running Restored Motor Learning Deficit Induced by Sorafenib
3.4. Neither Sorafenib Administration nor Voluntary Wheel Running Exercise Resulted in Any Changes in Muscle Weight or Strength
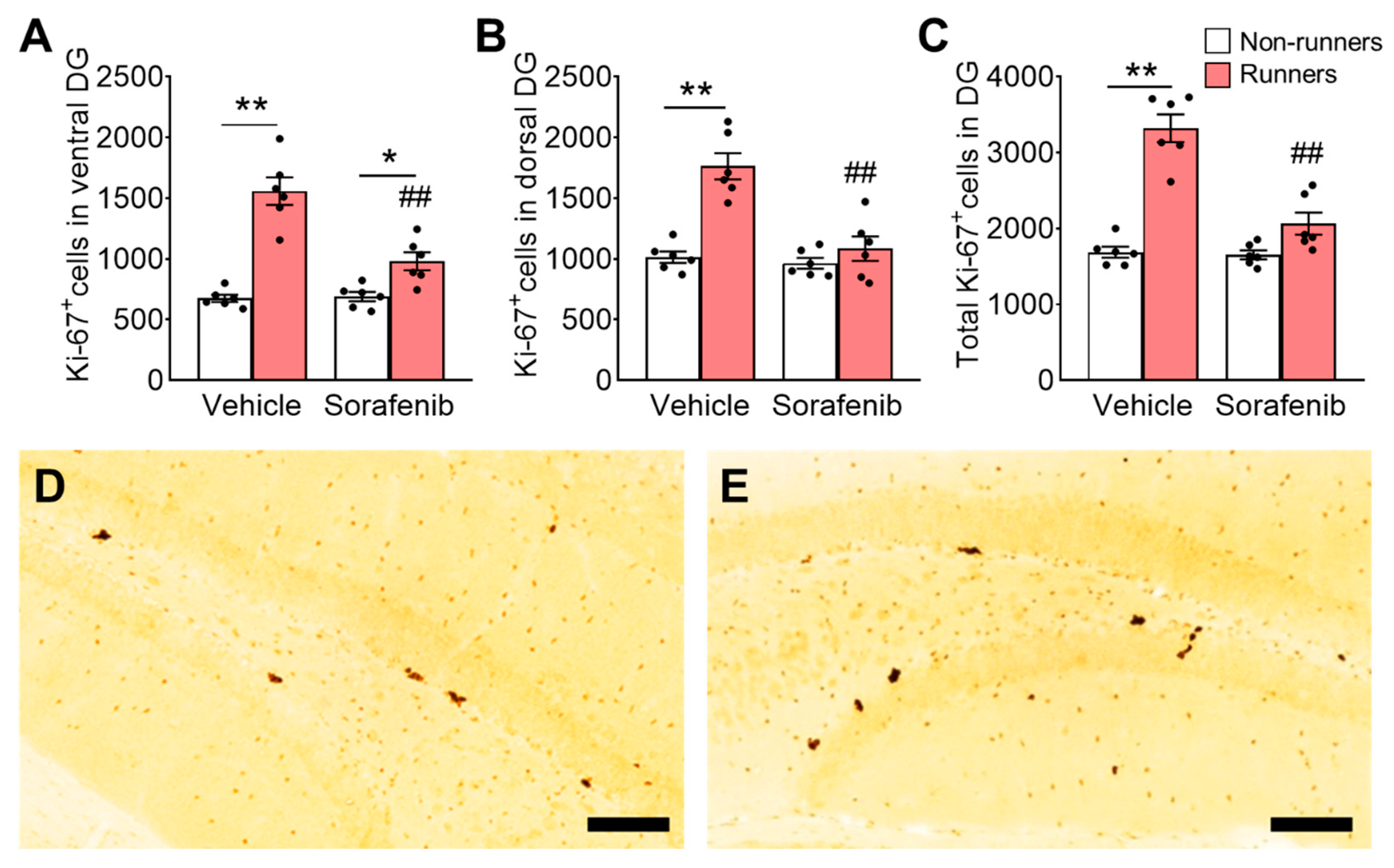
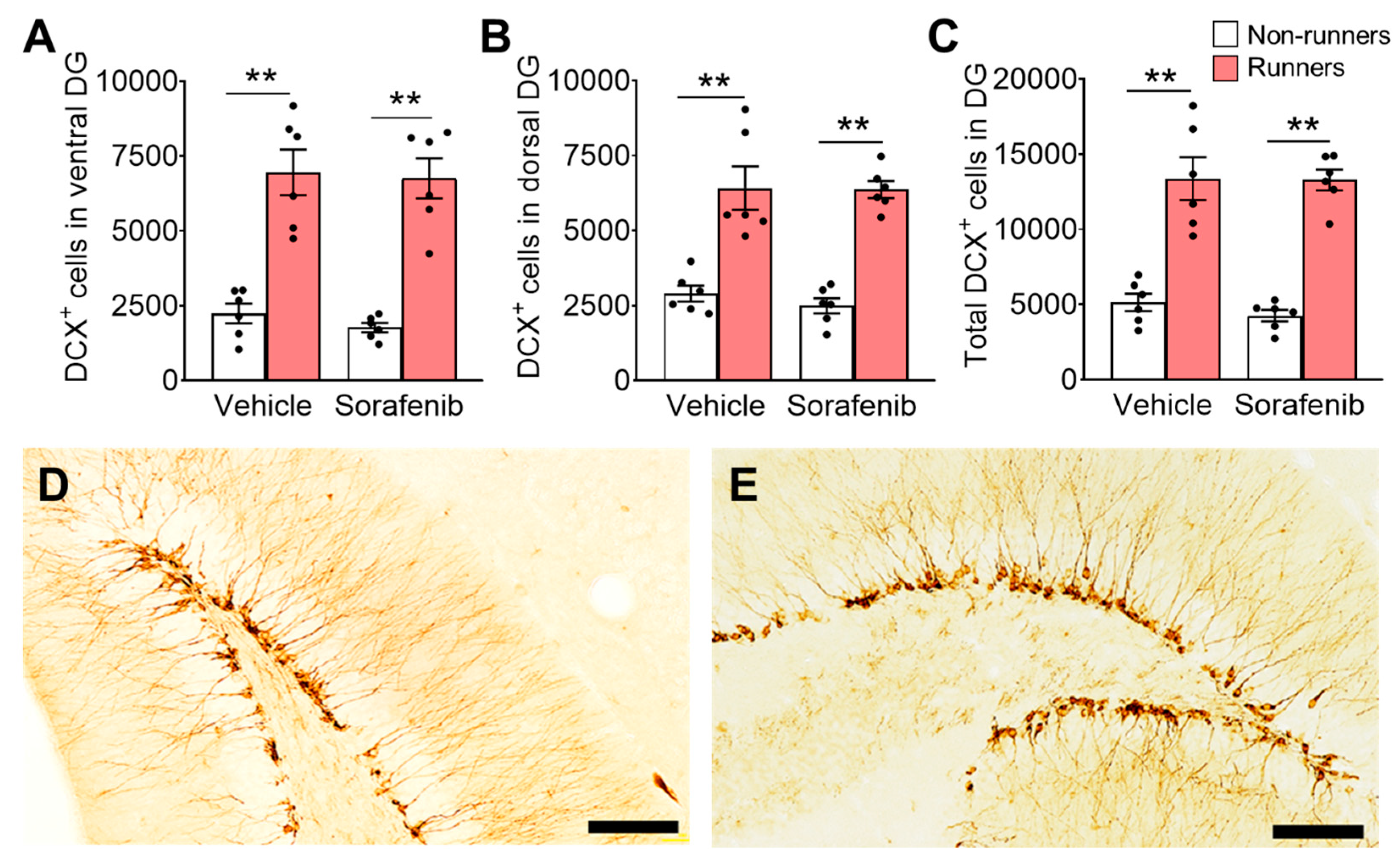
3.5. Voluntary Wheel Running Increased Number of Immature Neurons in the Hippocampus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gibson, E.M.; Monje, M. Treating cancer therapy–related cognitive impairment. Nat. Med. 2020, 26, 1174–1175. [Google Scholar] [CrossRef]
- Rausch, V.; Sala, V.; Penna, F.; Porporato, P.E.; Ghigo, A. Understanding the common mechanisms of heart and skeletal muscle wasting in cancer cachexia. Oncogenesis 2021, 10, 1. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, Z.; Ding, H.; Zhou, Y.; Doan, H.A.; Sin, K.W.T.; Zhu, Z.J.; Flores, R.; Wen, Y.; Gong, X. Tumor induces muscle wasting in mice through releasing extracellular Hsp70 and Hsp90. Nat. Commun. 2017, 8, 589. [Google Scholar] [CrossRef] [PubMed]
- Essex, A.L.; Pin, F.; Huot, J.R.; Bonewald, L.F.; Plotkin, L.I.; Bonetto, A. Bisphosphonate Treatment Ameliorates Chemotherapy-Induced Bone and Muscle Abnormalities in Young Mice. Front. Endocrinol. 2019, 10, 809. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Joly, F.; Vardy, J.; Ahles, T.; Dubois, M.; Tron, L.; Winocur, G.; De Ruiter, M.B.; Castel, H. Cancer-related cognitive impairment: An update on state of the art, detection, and management strategies in cancer survivors. Ann. Oncol. 2019, 30, 1925–1940. [Google Scholar] [CrossRef]
- Collins, B.; Mackenzie, J.; Tasca, G.A.; Scherling, C.; Smith, A. Persistent cognitive changes in breast cancer patients 1 year following completion of chemotherapy. J. Int. Neuropsychol. Soc. 2014, 20, 370–379. [Google Scholar] [CrossRef]
- Lindner, O.C.; Phillips, B.; McCabe, M.G.; Mayes, A.; Wearden, A.; Varese, F.; Talmi, D. A meta-analysis of cognitive impairment following adult cancer chemotherapy. Neuropsychology 2014, 28, 726–740. [Google Scholar] [CrossRef] [PubMed]
- Makino, M.; Shoji, H.; Takemoto, D.; Honboh, T.; Nakamura, S.; Kurayoshi, K.; Kaibara, N. Comparative study between daily and 5-days-a-week administration of oral 5-fluorouracil chemotherapy in mice: Determining the superior regimen. Cancer Chemother. Pharmacol. 2001, 48, 370–374. [Google Scholar] [CrossRef]
- Farhang-Sardroodi, S.; La Croix, M.A.; Wilkie, K.P. Chemotherapy-induced cachexia and model-informed dosing to preserve lean mass in cancer treatment. PLoS Comput. Biol. 2022, 18, e1009505. [Google Scholar] [CrossRef]
- Ross, P.; Ashley, S.; Norton, A.; Priest, K.; Waters, J.; Eisen, T.; Smith, I.; O’Brien, M. Do patients with weight loss have a worse outcome when undergoing chemotherapy for lung cancers? Br. J. Cancer 2004, 90, 1905–1911. [Google Scholar] [CrossRef]
- Hess, L.; Barakat, R.; Tian, C.; Ozols, R.; Alberts, D. Weight change during chemotherapy as a potential prognostic factor for stage III epithelial ovarian carcinoma: A Gynecologic Oncology Group study. Gynecol. Oncol. 2007, 107, 260–265. [Google Scholar] [CrossRef]
- Takayoshi, K.; Uchino, K.; Nakano, M.; Ikejiri, K.; Baba, E. Weight loss during initial chemotherapy predicts survival in patients with advanced gastric cancer. Nutr. Cancer 2017, 69, 408–415. [Google Scholar] [CrossRef]
- Chen, Z.-X.; Jian, Z.-W.; Wu, X.-W.; Wang, J.-C.; Peng, J.-Y.; Huang, C.-Y.; Lao, X.-M. Impact of overweightness and critical weight loss on overall survival in patients with hepatocellular carcinoma initially treated with chemoembolisation. Gastroenterol. Rep. 2020, 8, 125–133. [Google Scholar] [CrossRef]
- Carnie, L.; Abraham, M.; McNamara, M.G.; Hubner, R.A.; Valle, J.W.; Lamarca, A. Impact on prognosis of early weight loss during palliative chemotherapy in patients diagnosed with advanced pancreatic cancer. Pancreatology 2020, 20, 1682–1688. [Google Scholar] [CrossRef]
- Gouspillou, G.; Scheede-Bergdahl, C.; Spendiff, S.; Vuda, M.; Meehan, B.; Mlynarski, H.; Archer-Lahlou, E.; Sgarioto, N.; Purves-Smith, F.M.; Konokhova, Y. Anthracycline-containing chemotherapy causes long-term impairment of mitochondrial respiration and increased reactive oxygen species release in skeletal muscle. Sci. Rep. 2015, 5, 8717. [Google Scholar] [CrossRef]
- Campelj, D.G.; Goodman, C.A.; Rybalka, E. Chemotherapy-induced myopathy: The dark side of the cachexia sphere. Cancers 2021, 13, 3615. [Google Scholar] [CrossRef]
- Smith, H.R. Depression in cancer patients: Pathogenesis, implications and treatment. Oncol. Lett. 2015, 9, 1509–1514. [Google Scholar] [CrossRef] [PubMed]
- John, J.; Kinra, M.; Mudgal, J.; Viswanatha, G.; Nandakumar, K. Animal models of chemotherapy-induced cognitive decline in preclinical drug development. Psychopharmacology 2021, 238, 3025–3053. [Google Scholar] [CrossRef] [PubMed]
- Braun, T.P.; Szumowski, M.; Levasseur, P.R.; Grossberg, A.J.; Zhu, X.; Agarwal, A.; Marks, D.L. Muscle atrophy in response to cytotoxic chemotherapy is dependent on intact glucocorticoid signaling in skeletal muscle. PLoS ONE 2014, 9, e106489. [Google Scholar] [CrossRef] [PubMed]
- Hain, B.A.; Xu, H.; Waning, D.L. Loss of REDD1 prevents chemotherapy-induced muscle atrophy and weakness in mice. J. Cachexia Sarcopenia Muscle 2021, 12, 1597–1612. [Google Scholar] [CrossRef]
- Escudier, B.; Eisen, T.; Stadler, W.M.; Szczylik, C.; Oudard, S.; Siebels, M.; Negrier, S.; Chevreau, C.; Solska, E.; Desai, A.A. Sorafenib in advanced clear-cell renal-cell carcinoma. N. Engl. J. Med. 2007, 356, 125–134. [Google Scholar] [CrossRef]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.-F.; De Oliveira, A.C.; Santoro, A.; Raoul, J.-L.; Forner, A. Sorafenib in advanced hepatocellular carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef]
- Liu, L.; Cao, Y.; Chen, C.; Zhang, X.; McNabola, A.; Wilkie, D.; Wilhelm, S.; Lynch, M.; Carter, C. Sorafenib blocks the RAF/MEK/ERK pathway, inhibits tumor angiogenesis, and induces tumor cell apoptosis in hepatocellular carcinoma model PLC/PRF/5. Cancer Res. 2006, 66, 11851–11858. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.M.; Carter, C.; Tang, L.; Wilkie, D.; McNabola, A.; Rong, H.; Chen, C.; Zhang, X.; Vincent, P.; McHugh, M. BAY 43-9006 exhibits broad spectrum oral antitumor activity and targets the RAF/MEK/ERK pathway and receptor tyrosine kinases involved in tumor progression and angiogenesis. Cancer Res. 2004, 64, 7099–7109. [Google Scholar] [CrossRef]
- Zhang, T.; Ding, X.; Wei, D.; Cheng, P.; Su, X.; Liu, H.; Wang, D.; Gao, H. Sorafenib improves the survival of patients with advanced hepatocellular carcinoma: A meta-analysis of randomised trials. Anti-Cancer Drugs 2010, 21, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.-C.; He, Y.-Q.; Qiu, Y.-S.; Ni, C.-X.; Shen, F.-M.; Li, D.-J. Zinc supplementation ameliorates sorafenib-induced cognitive impairment through ROS/JNK signaling pathway. Biol. Trace Elem. Res. 2023, 201, 324–337. [Google Scholar] [CrossRef]
- Damaraju, V.L.; Kuzma, M.; Cass, C.E.; Putman, C.T.; Sawyer, M.B. Multitargeted kinase inhibitors imatinib, sorafenib and sunitinib perturb energy metabolism and cause cytotoxicity to cultured C2C12 skeletal muscle derived myotubes. Biochem. Pharmacol. 2018, 155, 162–171. [Google Scholar] [CrossRef]
- Brandi, G.; de Rosa, F.; Calzà, L.; Girolamo, S.D.; Tufoni, M.; Ricci, C.S.; Cirignotta, F.; Caraceni, P.; Biasco, G. Can the tyrosine kinase inhibitors trigger metabolic encephalopathy in cirrhotic patients? Liver Int. 2013, 33, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Mulder, S.F.; Bertens, D.; Desar, I.M.; Vissers, K.C.; Mulders, P.F.; Punt, C.J.; van Spronsen, D.-J.; Langenhuijsen, J.F.; Kessels, R.P.; van Herpen, C.M. Impairment of cognitive functioning during Sunitinib or Sorafenib treatment in cancer patients: A cross sectional study. BMC Cancer 2014, 14, 219. [Google Scholar] [CrossRef]
- Antoun, S.; Birdsell, L.; Sawyer, M.B.; Venner, P.; Escudier, B.; Baracos, V.E. Association of skeletal muscle wasting with treatment with sorafenib in patients with advanced renal cell carcinoma: Results from a placebo-controlled study. J. Clin. Oncol. 2010, 28, 1054–1060. [Google Scholar] [CrossRef]
- Antoun, S.; Baracos, V.; Birdsell, L.; Escudier, B.; Sawyer, M. Low body mass index and sarcopenia associated with dose-limiting toxicity of sorafenib in patients with renal cell carcinoma. Ann. Oncol. 2010, 21, 1594–1598. [Google Scholar] [CrossRef] [PubMed]
- Karbownik, A.; Stanisławiak-Rudowicz, J.; Stachowiak, A.; Romański, M.; Grześkowiak, E.; Szałek, E. The Influence of Paracetamol on the Penetration of Sorafenib and Sorafenib N-Oxide Through the Blood–Brain Barrier in Rats. Eur. J. Drug Metab. Pharmacokinet. 2020, 45, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Dahlmanns, M.; Yakubov, E.; Chen, D.; Sehm, T.; Rauh, M.; Savaskan, N.; Wrosch, J.K. Chemotherapeutic xCT inhibitors sorafenib and erastin unraveled with the synaptic optogenetic function analysis tool. Cell Death Discov. 2017, 3, 17030. [Google Scholar] [CrossRef]
- Deng, W.; Aimone, J.B.; Gage, F.H. New neurons and new memories: How does adult hippocampal neurogenesis affect learning and memory? Nat. Rev. Neurosci. 2010, 11, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Sahay, A.; Hen, R. Adult hippocampal neurogenesis in depression. Nat. Neurosci. 2007, 10, 1110–1115. [Google Scholar] [CrossRef]
- Tsitkanou, S.; Murach, K.A.; Washington, T.A.; Greene, N.P. Exercise counteracts the deleterious effects of cancer cachexia. Cancers 2022, 14, 2512. [Google Scholar] [CrossRef]
- Hojman, P.; Fjelbye, J.; Zerahn, B.; Christensen, J.F.; Dethlefsen, C.; Lonkvist, C.K.; Brandt, C.; Gissel, H.; Pedersen, B.K.; Gehl, J. Voluntary exercise prevents cisplatin-induced muscle wasting during chemotherapy in mice. PLoS ONE 2014, 9, e109030. [Google Scholar] [CrossRef]
- Ballarò, R.; Beltrà, M.; De Lucia, S.; Pin, F.; Ranjbar, K.; Hulmi, J.J.; Costelli, P.; Penna, F. Moderate exercise in mice improves cancer plus chemotherapy-induced muscle wasting and mitochondrial alterations. FASEB J. 2019, 33, 5482–5494. [Google Scholar] [CrossRef]
- Slivicki, R.A.; Mali, S.S.; Hohmann, A.G. Voluntary exercise reduces both chemotherapy-induced neuropathic nociception and deficits in hippocampal cellular proliferation in a mouse model of paclitaxel-induced peripheral neuropathy. Neurobiol. Pain 2019, 6, 100035. [Google Scholar] [CrossRef]
- Alves de Lima Jr, E.; Teixeira, A.A.d.S.; Biondo, L.A.; Diniz, T.A.; Silveira, L.S.; Coletti, D.; Busquets Rius, S.; Rosa Neto, J.C. Exercise reduces the resumption of tumor growth and proteolytic pathways in the skeletal muscle of mice following chemotherapy. Cancers 2020, 12, 3466. [Google Scholar] [CrossRef]
- Wakefield, Z.R.; Tanaka, M.; Pampo, C.; Lepler, S.; Rice, L.; Guingab-Cagmat, J.; Garrett, T.J.; Siemann, D.W. Normal tissue and tumor microenvironment adaptations to aerobic exercise enhance doxorubicin anti-tumor efficacy and ameliorate its cardiotoxicity in retired breeder mice. Oncotarget 2021, 12, 1737. [Google Scholar] [CrossRef]
- Fardell, J.E.; Vardy, J.; Shah, J.D.; Johnston, I.N. Cognitive impairments caused by oxaliplatin and 5-fluorouracil chemotherapy are ameliorated by physical activity. Psychopharmacology 2012, 220, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Winocur, G.; Wojtowicz, J.M.; Huang, J.; Tannock, I.F. Physical exercise prevents suppression of hippocampal neurogenesis and reduces cognitive impairment in chemotherapy-treated rats. Psychopharmacology 2014, 231, 2311–2320. [Google Scholar] [CrossRef]
- Winocur, G.; Wojtowicz, J.M.; Tannock, I.F. Memory loss in chemotherapy-treated rats is exacerbated in high-interference conditions and related to suppression of hippocampal neurogenesis. Behav. Brain Res. 2015, 281, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.-V.; Hsu, T.-H.; Wu, W.-T.; Huang, K.-C.; Han, D.-S. Association between sarcopenia and cognitive impairment: A systematic review and meta-analysis. J. Am. Med. Dir. Assoc. 2016, 17, 1164.e7–1164.e15. [Google Scholar] [CrossRef]
- Basile, G.; Sardella, A. From cognitive to motor impairment and from sarcopenia to cognitive impairment: A bidirectional pathway towards frailty and disability. Aging Clin. Exp. Res. 2021, 33, 469–478. [Google Scholar] [CrossRef]
- Sui, S.X.; Hordacre, B.; Pasco, J.A. Are sarcopenia and cognitive dysfunction comorbid after stroke in the context of brain–muscle crosstalk? Biomedicines 2021, 9, 223. [Google Scholar] [CrossRef] [PubMed]
- Mangiameli, D.P.; Blansfield, J.A.; Kachala, S.; Lorang, D.; Schafer, P.H.; Muller, G.W.; Stirling, D.I.; Libutti, S.K. Combination therapy targeting the tumor microenvironment is effective in a model of human ocular melanoma. J. Transl. Med. 2007, 5, 38. [Google Scholar] [CrossRef]
- Yau, S.Y.; Li, A.; Hoo, R.L.; Ching, Y.P.; Christie, B.R.; Lee, T.M.; Xu, A.; So, K.-F. Physical exercise-induced hippocampal neurogenesis and antidepressant effects are mediated by the adipocyte hormone adiponectin. Proc. Natl. Acad. Sci. USA 2014, 111, 15810–15815. [Google Scholar] [CrossRef]
- Lee, T.H.; Christie, B.R.; van Praag, H.; Lin, K.; Siu, P.M.-F.; Xu, A.; So, K.-F.; Yau, S.-y. AdipoRon treatment induces a dose-dependent response in adult hippocampal neurogenesis. Int. J. Mol. Sci. 2021, 22, 2068. [Google Scholar] [CrossRef]
- Sclafani, A. Starch and sugar tastes in rodents: An update. Brain Res. Bull. 1991, 27, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Gorwood, P. Neurobiological mechanisms of anhedonia. Dialogues Clin. Neurosci. 2022, 10, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Commons, K.G.; Cholanians, A.B.; Babb, J.A.; Ehlinger, D.G. The rodent forced swim test measures stress-coping strategy, not depression-like behavior. ACS Chem. Neurosci. 2017, 8, 955–960. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.; Roberts, D. The quantitative measurement of motor inco-ordination in naive mice using an accelerating rotarod. J. Pharm. Pharmacol. 1968, 20, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Toledo, M.; Busquets, S.; Sirisi, S.; Serpe, R.; Orpi, M.; Coutinho, J.; Martinez, R.; Lopez-Soriano, F.J.; Argiles, J.M. Cancer cachexia: Physical activity and muscle force in tumour-bearing rats. Oncol. Rep. 2011, 25, 189–193. [Google Scholar]
- Deacon, R.M. Measuring the strength of mice. JoVE J. Vis. Exp. 2013, 76, e2610. [Google Scholar]
- Yau, S.-Y.; Lee, T.H.-Y.; Formolo, D.A.; Lee, W.-L.; Li, L.C.-K.; Siu, P.M.; Chan, C.C. Effects of maternal voluntary wheel running during pregnancy on adult hippocampal neurogenesis, temporal order memory, and depression-like behavior in adult female and male offspring. Front. Neurosci. 2019, 13, 470. [Google Scholar] [CrossRef]
- Collins, C.A.; Zammit, P.S. Isolation and grafting of single muscle fibres. Stem Cells Regen. Med. 2009, 482, 319–330. [Google Scholar]
- Van Waart, H.; Stuiver, M.M.; van Harten, W.H.; Geleijn, E.; Kieffer, J.M.; Buffart, L.M.; de Maaker-Berkhof, M.; Boven, E.; Schrama, J.; Geenen, M.M. Effect of low-intensity physical activity and moderate-to high-intensity physical exercise during adjuvant chemotherapy on physical fitness, fatigue, and chemotherapy completion rates: Results of the PACES randomised clinical trial. J. Clin. Oncol. 2015, 33, 1918–1927. [Google Scholar] [CrossRef]
- Park, J.S.; Kim, S.; Hoke, A. An exercise regimen prevents development paclitaxel induced peripheral neuropathy in a mouse model. J. Peripher. Nerv. Syst. 2015, 20, 7–14. [Google Scholar] [CrossRef]
- Licht, T.; Goshen, I.; Avital, A.; Kreisel, T.; Zubedat, S.; Eavri, R.; Segal, M.; Yirmiya, R.; Keshet, E. Reversible modulations of neuronal plasticity by VEGF. Proc. Natl. Acad. Sci. USA 2011, 108, 5081–5086. [Google Scholar] [CrossRef] [PubMed]
- Karakilic, A.; Yuksel, O.; Kizildag, S.; Hosgorler, F.; Topcugil, B.; Ilgin, R.; Gumus, H.; Guvendi, G.; Koc, B.; Kandis, S. Regular aerobic exercise increased VEGF levels in both soleus and gastrocnemius muscles correlated with hippocampal learning and VEGF levels. Acta Neurobiol. Exp. 2021, 81, 1–9. [Google Scholar] [CrossRef]
- Nguyen, L.D.; Ehrlich, B.E. Cellular mechanisms and treatments for chemobrain: Insight from aging and neurodegenerative diseases. EMBO Mol. Med. 2020, 12, e12075. [Google Scholar] [CrossRef] [PubMed]
- Seigers, R.; Schagen, S.B.; Beerling, W.; Boogerd, W.; Van Tellingen, O.; Van Dam, F.S.; Koolhaas, J.M.; Buwalda, B. Long-lasting suppression of hippocampal cell proliferation and impaired cognitive performance by methotrexate in the rat. Behav. Brain Res. 2008, 186, 168–175. [Google Scholar] [CrossRef] [PubMed]
- ELBeltagy, M.; Mustafa, S.; Umka, J.; Lyons, L.; Salman, A.; Tu, C.-Y.G.; Bhalla, N.; Bennett, G.; Wigmore, P.M. Fluoxetine improves the memory deficits caused by the chemotherapy agent 5-fluorouracil. Behav. Brain Res. 2010, 208, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Nokia, M.S.; Anderson, M.L.; Shors, T.J. Chemotherapy disrupts learning, neurogenesis and theta activity in the adult brain. Eur. J. Neurosci. 2012, 36, 3521–3530. [Google Scholar] [CrossRef]
- Anacker, C.; Luna, V.M.; Stevens, G.S.; Millette, A.; Shores, R.; Jimenez, J.C.; Chen, B.; Hen, R. Hippocampal neurogenesis confers stress resilience by inhibiting the ventral dentate gyrus. Nature 2018, 559, 98–102. [Google Scholar] [CrossRef]
- Tye, K.M.; Mirzabekov, J.J.; Warden, M.R.; Ferenczi, E.A.; Tsai, H.-C.; Finkelstein, J.; Kim, S.-Y.; Adhikari, A.; Thompson, K.R.; Andalman, A.S. Dopamine neurons modulate neural encoding and expression of depression-related behaviour. Nature 2013, 493, 537–541. [Google Scholar] [CrossRef]
- Markovic, T.; Pedersen, C.E.; Massaly, N.; Vachez, Y.M.; Ruyle, B.; Murphy, C.A.; Abiraman, K.; Shin, J.H.; Garcia, J.J.; Yoon, H.J. Pain induces adaptations in ventral tegmental area dopamine neurons to drive anhedonia-like behavior. Nat. Neurosci. 2021, 24, 1601–1613. [Google Scholar] [CrossRef]
- Molina-Luna, K.; Pekanovic, A.; Röhrich, S.; Hertler, B.; Schubring-Giese, M.; Rioult-Pedotti, M.-S.; Luft, A.R. Dopamine in motor cortex is necessary for skill learning and synaptic plasticity. PLoS ONE 2009, 4, e7082. [Google Scholar] [CrossRef]
- Rioult-Pedotti, M.-S.; Pekanovic, A.; Atiemo, C.O.; Marshall, J.; Luft, A.R. Dopamine promotes motor cortex plasticity and motor skill learning via PLC activation. PLoS ONE 2015, 10, e0124986. [Google Scholar] [CrossRef]
- Panigrahi, B.; Martin, K.A.; Li, Y.; Graves, A.R.; Vollmer, A.; Olson, L.; Mensh, B.D.; Karpova, A.Y.; Dudman, J.T. Dopamine is required for the neural representation and control of movement vigor. Cell 2015, 162, 1418–1430. [Google Scholar] [CrossRef]
- Hosp, J.A.; Pekanovic, A.; Rioult-Pedotti, M.S.; Luft, A.R. Dopaminergic projections from midbrain to primary motor cortex mediate motor skill learning. J. Neurosci. 2011, 31, 2481–2487. [Google Scholar] [CrossRef]
- Bastioli, G.; Arnold, J.C.; Mancini, M.; Mar, A.C.; Gamallo-Lana, B.; Saadipour, K.; Chao, M.V.; Rice, M.E. Voluntary exercise boosts striatal dopamine release: Evidence for the necessary and sufficient role of BDNF. J. Neurosci. 2022, 42, 4725–4736. [Google Scholar] [CrossRef] [PubMed]
- Leemburg, S.; Canonica, T.; Luft, A. Motor skill learning and reward consumption differentially affect VTA activation. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Kung, J.; Dong, J.; Chang, L.; Xie, C.; Habib, A.; Hawes, S.; Yang, N.; Chen, V.; Liu, Z. Distinct connectivity and functionality of aldehyde dehydrogenase 1a1-positive nigrostriatal dopaminergic neurons in motor learning. Cell Rep. 2019, 28, 1167–1181.e1167. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M. Measuring motor coordination in mice. JoVE J. Vis. Exp. 2013, 75, e2609. [Google Scholar]
- Delavar, H.; Nogueira, L.; Wagner, P.D.; Hogan, M.C.; Metzger, D.; Breen, E.C. Skeletal myofiber VEGF is essential for the exercise training response in adult mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, R586–R595. [Google Scholar] [CrossRef] [PubMed]
- Huot, J.R.; Essex, A.L.; Gutierrez, M.; Barreto, R.; Wang, M.; Waning, D.L.; Plotkin, L.I.; Bonetto, A. Chronic Treatment with Multi-Kinase Inhibitors Causes Differential Toxicities on Skeletal and Cardiac Muscles. Cancers 2019, 11, 571. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, H.J.; Lee, W.J.; Seong, J.K. A comparison of the metabolic effects of treadmill and wheel running exercise in mouse model. Lab. Anim. Res. 2020, 36, 3. [Google Scholar] [CrossRef]
- Tauer, J.T.; Canevazzi, G.H.R.; Schiettekatte-Maltais, J.; Rauch, F.; Bergeron, R.; Veilleux, L.-N. Muscle-bone properties after prolonged voluntary wheel running in a mouse model of dominant severe osteogenesis imperfecta. J. Musculoskelet. Neuronal Interact. 2021, 21, 517. [Google Scholar]
- Yang, M.; Kim, J.-S.; Kim, J.; Jang, S.; Kim, S.-H.; Kim, J.-C.; Shin, T.; Wang, H.; Moon, C. Acute treatment with methotrexate induces hippocampal dysfunction in a mouse model of breast cancer. Brain Res. Bull. 2012, 89, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Kim, J.; Kim, J.-S.; Kim, S.-H.; Kim, J.-C.; Kang, M.-J.; Jung, U.; Shin, T.; Wang, H.; Moon, C. Hippocampal dysfunctions in tumor-bearing mice. Brain Behav. Immun. 2014, 36, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Winocur, G.; Berman, H.; Nguyen, M.; Binns, M.A.; Henkelman, M.; van Eede, M.; Piquette-Miller, M.; Sekeres, M.J.; Wojtowicz, J.M.; Yu, J. Neurobiological mechanisms of chemotherapy-induced cognitive impairment in a transgenic model of breast cancer. Neuroscience 2018, 369, 51–65. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, T.H.; Devaki, M.; Formolo, D.A.; Rosa, J.M.; Cheng, A.S.K.; Yau, S.-Y. Effects of Voluntary Wheel Running Exercise on Chemotherapy-Impaired Cognitive and Motor Performance in Mice. Int. J. Environ. Res. Public Health 2023, 20, 5371. https://doi.org/10.3390/ijerph20075371
Lee TH, Devaki M, Formolo DA, Rosa JM, Cheng ASK, Yau S-Y. Effects of Voluntary Wheel Running Exercise on Chemotherapy-Impaired Cognitive and Motor Performance in Mice. International Journal of Environmental Research and Public Health. 2023; 20(7):5371. https://doi.org/10.3390/ijerph20075371
Chicago/Turabian StyleLee, Thomas H., Malegaddi Devaki, Douglas A. Formolo, Julia M. Rosa, Andy S. K. Cheng, and Suk-Yu Yau. 2023. "Effects of Voluntary Wheel Running Exercise on Chemotherapy-Impaired Cognitive and Motor Performance in Mice" International Journal of Environmental Research and Public Health 20, no. 7: 5371. https://doi.org/10.3390/ijerph20075371
APA StyleLee, T. H., Devaki, M., Formolo, D. A., Rosa, J. M., Cheng, A. S. K., & Yau, S.-Y. (2023). Effects of Voluntary Wheel Running Exercise on Chemotherapy-Impaired Cognitive and Motor Performance in Mice. International Journal of Environmental Research and Public Health, 20(7), 5371. https://doi.org/10.3390/ijerph20075371