
Current and Future Potential of Shellfish and Algae Mariculture Carbon Sinks in China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sources
2.2. Assessment of Carbon Sink Capacity of Shellfish and Algae
2.3. Accounting for the Economic Value of the Shellfish and Algae Carbon Sinks
3. Results
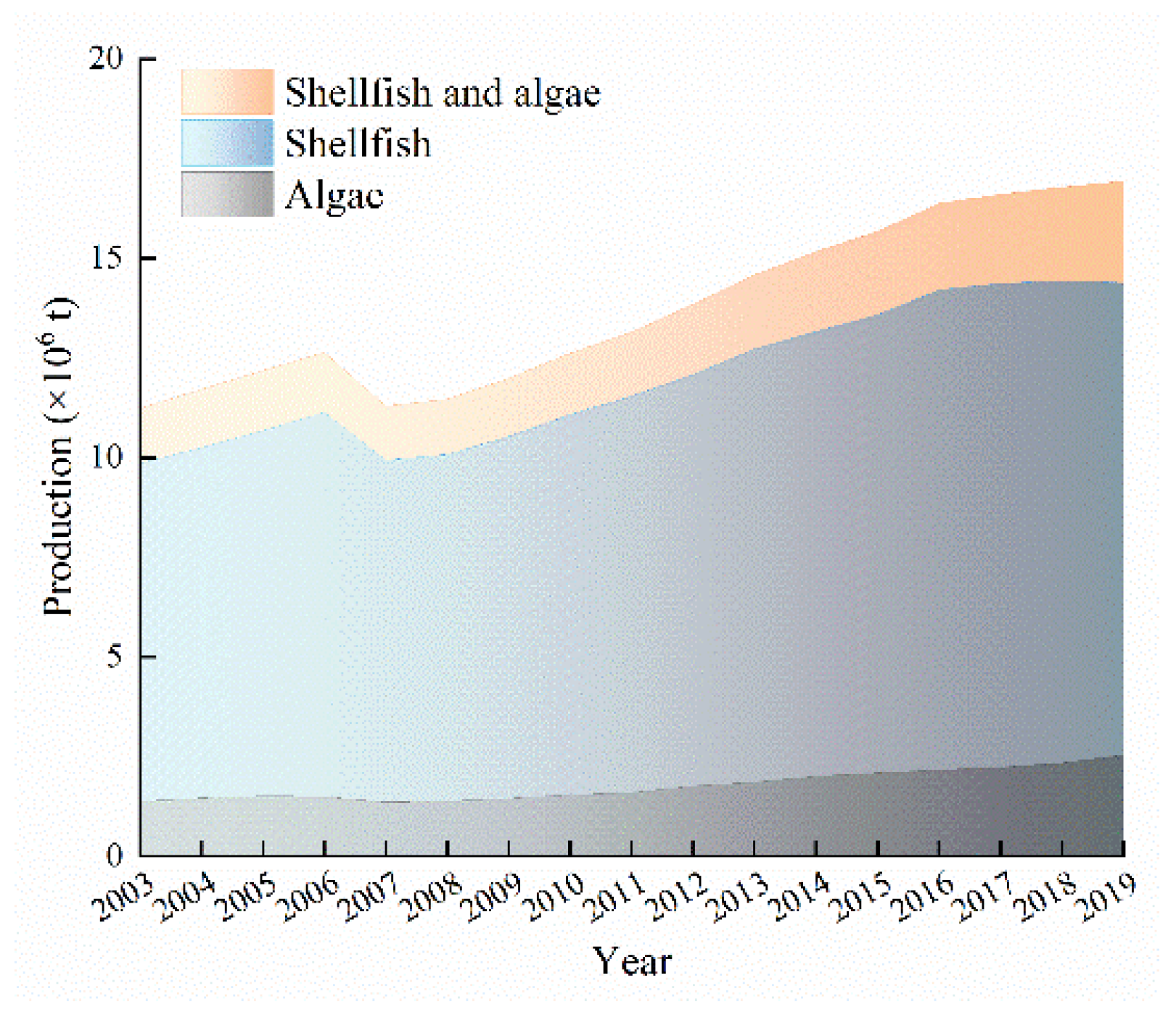
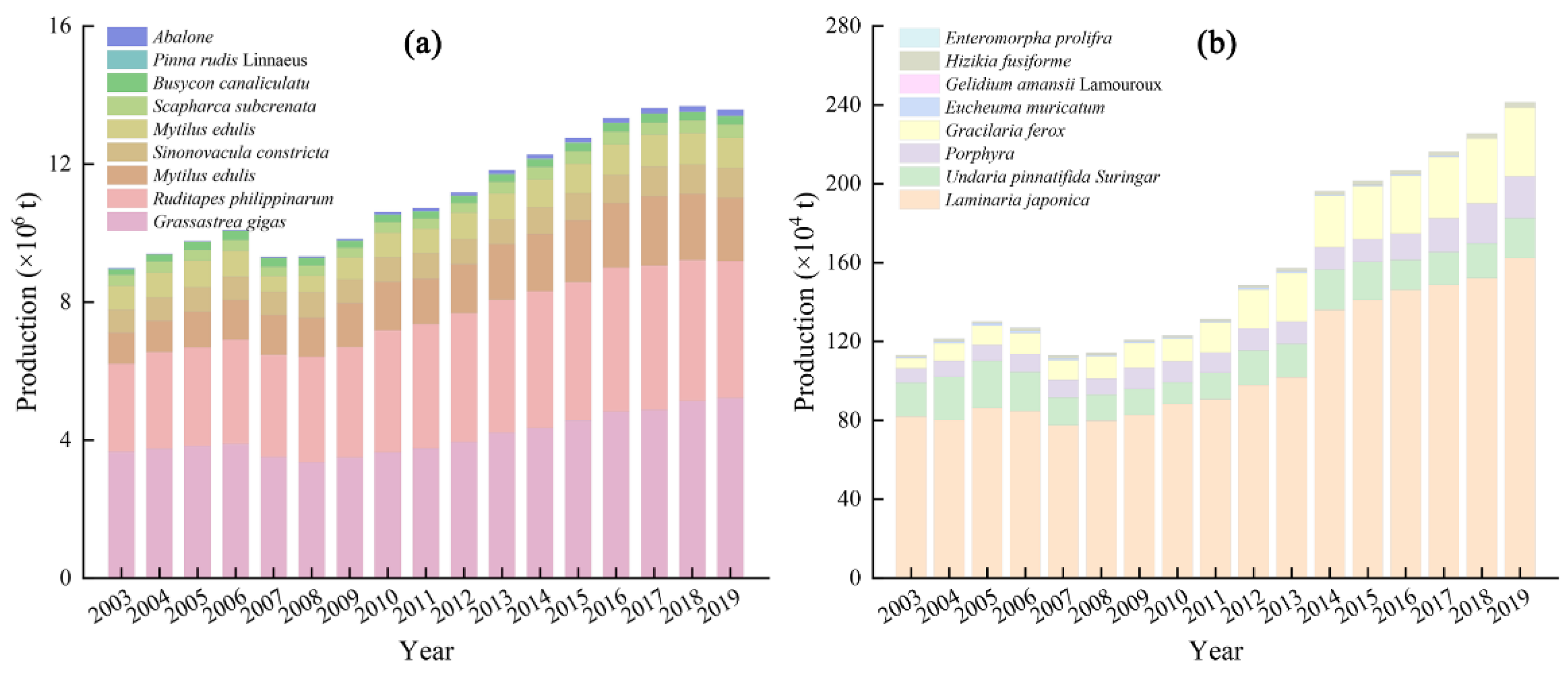
3.1. Status of Shellfish and Algae Culture
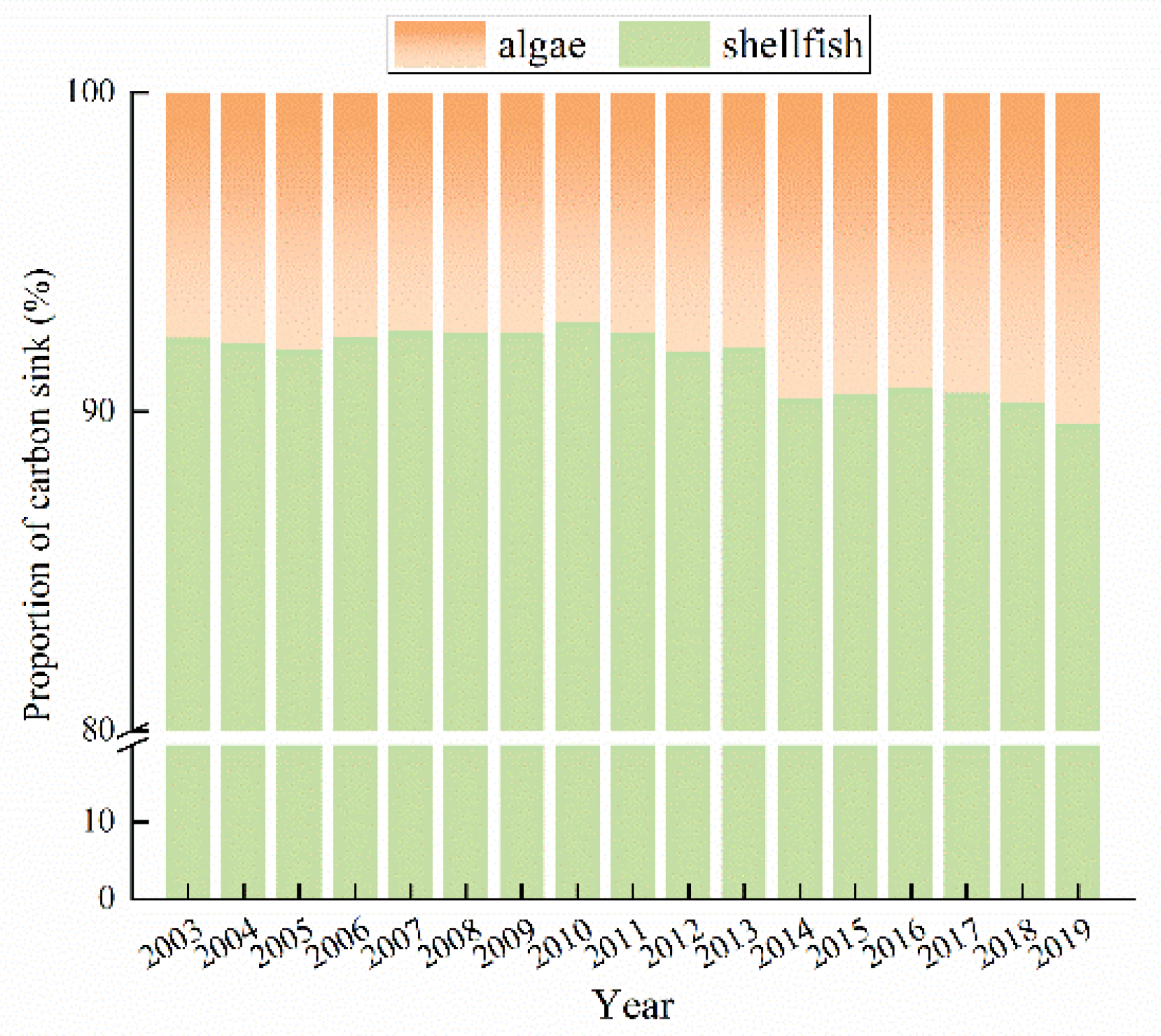
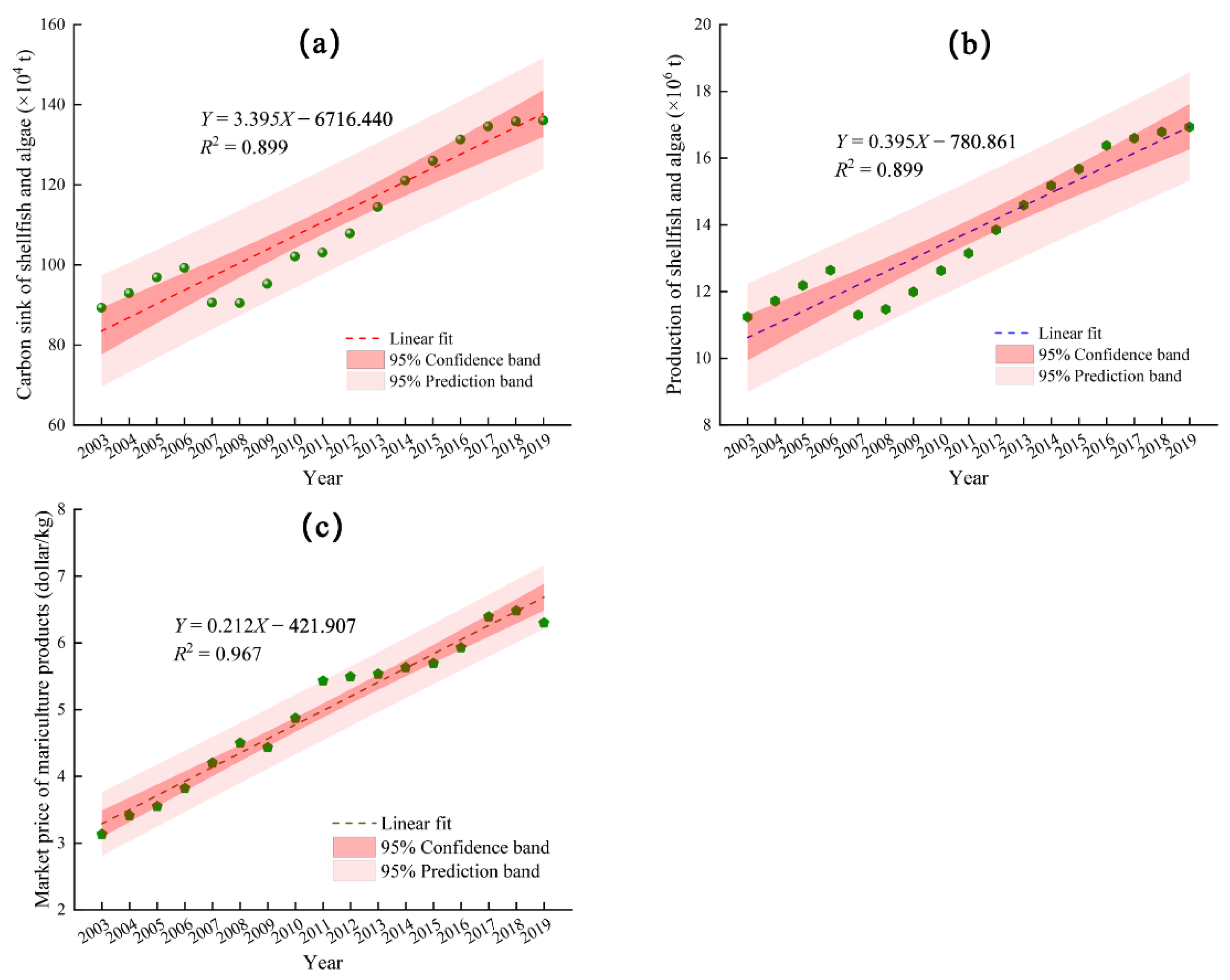
3.2. Carbon Sink Capacity of Shellfish and Algae Mariculture
3.3. Economic Value of the Shellfish and Algae Carbon Sinks
4. Discussion
4.1. Carbon Sink Conversion Efficiency of Shellfish and Algae
4.2. Carbon Sink Potential of Shellfish and Algae
4.3. Amplification of Carbon Sink Capacity of Shellfish and Algae
- (1)
- Expand breeding space and improve breeding yield
- (2)
- Promote multi-trophic-level, comprehensive breeding models
- (3)
- Implementation of artificial marine upwelling exchange enhancement project
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frame, D.J. Assessment of the first consensus prediction on climate change. Nat. Clim. Change 2013, 3, 357–359. [Google Scholar] [CrossRef]
- Schleussner, C.; Rogelj, J.; Schaeffer, M.; Lissner, T.; Licker, R.; Fischer, E.M.; Knutti, R.; Levermann, A.; Frieler, K.; Hare, W. Science and policy characteristics of the Paris Agreement temperature goal. Nat. Clim. Change 2016, 6, 827–835. [Google Scholar] [CrossRef] [Green Version]
- Peters, G.P.; Andrew, R.M.; Canadell, J.G.; Fuss, S.; Jackson, R.B.; Korsbakken, J.I.; Le Quéré, C.; Nakicenovic, N. Key indicators to track current progress and future ambition of the Paris Agreement. Nat. Clim. Change 2017, 7, 118–122. [Google Scholar] [CrossRef]
- Wan, B.; Tian, L.; Fu, M.; Zhang, G. Green development growth momentum under carbon neutrality scenario. J. Clean. Prod. 2021, 316, 128327. [Google Scholar] [CrossRef]
- Macreadie, P.I. The future of Blue Carbon science. Nat. Commun. 2019, 10, 3998. [Google Scholar] [CrossRef] [Green Version]
- Pautova, L.A. The biological calcium carbonate pump in the Norwegian and Barents Seas; regulation mechanisms. Dokl. Earth Sci. 2020, 490, 46–50. [Google Scholar] [CrossRef]
- Hamme, R.C. Using noble gases to assess the ocean’s carbon pumps. Annu. Rev. Mar. Sci. 2019, 11, 75–103. [Google Scholar] [CrossRef]
- Barange, M. The cost of reducing the north atlantic ocean biological carbon pump. Front. Mar. Sci. 2017, 3, 290. [Google Scholar] [CrossRef] [Green Version]
- Jiao, N. Microbial production of recalcitrant dissolved organic matter: Long-term carbon storage in the global ocean. Nat. Rev. Microbiol. 2010, 8, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, S.W. Stirring times in the Southern Ocean. Nature 2000, 407, 685–686. [Google Scholar] [CrossRef]
- Jones, A.R.; Alleway, H.K.; McAfee, D.; Reis-Santos, P.; Theuerkauf, S.J.; Jones, R.C. Climate-friendly seafood: The potential for emissions reduction and carbon capture in marine aquaculture. Bioscience 2022, 72, 123–143. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Ward, J.D.; Thompson, S.; Saint, C.P.; Diana, J.S. Blue-green water nexus in aquaculture for resilience to climate change. Rev. Fish. Sci. Aquac. 2018, 26, 139–154. [Google Scholar] [CrossRef]
- Ren, W. Study on the removable carbon sink estimation and decomposition of influencing factors of mariculture shellfish and algae in China—A two-dimensional perspective based on scale and structure. Environ. Sci. Pollut. Res. 2021, 28, 21528–21539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Liang, Y.; Li, H.; Li, G.; Chen, X.; Zhao, P.; Jiang, Z.; Zou, D.; Liu, X.; et al. Carbon sequestration processes and mechanisms in coastal mariculture environments in China. Sci. China Earth Sci. 2017, 60, 2097–2107. [Google Scholar] [CrossRef]
- Pessarrodona, A. Carbon assimilation and transfer through kelp forests in the NE Atlantic is diminished under a warmer ocean climate. Glob. Change Biol. 2018, 24, 4386–4398. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.M.; Wu, J.; Xiao, X.; Bruhn, A.; Krause-Jensen, D. Can seaweed farming play a role in climate change mitigation and adaptation? Front. Mar. Sci. 2017, 4, 100. [Google Scholar] [CrossRef]
- Fodrie, F.J. Oyster reefs as carbon sources and sinks. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170891. [Google Scholar] [CrossRef]
- Bertolini, C.; Bernardini, I.; Brigolin, D.; Matozzo, V.; Milan, M.; Pastres, R. A bioenergetic model to address carbon sequestration potential of shellfish farming: Example from Ruditapes philippinarum in the Venice lagoon. ICES J. Mar. Sci. 2021, 78, 2082–2091. [Google Scholar] [CrossRef]
- Hu, C.; Wang, M.; Lapointe, B.E.; Brewton, R.A.; Hernandez, F.J. On the Atlantic pelagic Sargassum’s role in carbon fixation and sequestration. Sci. Total Environ. 2021, 781, 146801. [Google Scholar] [CrossRef]
- Wang, M.; Hu, C.; Cannizzaro, J.; English, D.; Han, X.; Naar, D.; Lapointe, B.; Brewton, R.; Hernandez, F. Remote Sensing of Sargassum Biomass, Nutrients, and Pigments. Geophys. Res. Lett. 2018, 45, 12359–12367. [Google Scholar] [CrossRef]
- Mueller, K. Differing methods of accounting ocean carbon sequestration efficiency. J. Geophys. Res. 2004, 109, C12018. [Google Scholar] [CrossRef] [Green Version]
- Suplicy, F.M. A review of the multiple benefits of mussel farming. Rev. Aquacult. 2020, 12, 204–223. [Google Scholar] [CrossRef]
- Filgueira, R.; Guyondet, T.; Bacher, C.; Comeau, L.A. Informing Marine Spatial Planning (MSP) with numerical modelling: A case-study on shellfish aquaculture in Malpeque Bay (Eastern Canada). Mar. Pollut. Bull. 2015, 100, 200–216. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Zhang, Z.; Chen, L.; Wang, D.; Liu, H.; Wang, Q.; Wang, M.; Yu, D. Changes in ecosystem services values in the south and north Yellow Sea between 2000 and 2010. Ocean. Coast. Manag. 2021, 202, 105497. [Google Scholar] [CrossRef]
- Soloviy, I.; Kuryltsiv, R.; Hernik, J.; Kryshenyk, N.; Kuleshnyk, T. Integrating ecosystem services valuation into land use planning: Case of the ukrainian agricultural landscapes. Forests 2021, 12, 1465. [Google Scholar] [CrossRef]
- Zan, X.; Xu, B.; Zhang, C.; Ren, Y. Annual variations of biogenic element contents of manila clam (Ruditapes philippinarum) bottom-cultivated in Jiaozhou Bay, China. J. Ocean Univ. China 2014, 13, 637–646. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Littler, M.M.; Littler, D.S. Nutrient availability to marine macroalgae versus carbonate-rich coastal waters in siliciclastic. Estuaries 1992, 15, 75–82. [Google Scholar] [CrossRef]
- Ge, C.; Wang, H.; Kan, M.; Chai, Y. Carbon sequestration within silica bodies extracted from kelp cultured in the East China Sea. Silicon Neth 2017, 9, 613–618. [Google Scholar] [CrossRef]
- Schiener, P.; Black, K.D.; Stanley, M.S.; Green, D.H. The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. J. Appl. Phycol. 2015, 27, 363–373. [Google Scholar] [CrossRef]
- Aller-Rojas, O.; Moreno, B.; Aponte, H.; Zavala, J. Carbon storage estimation of Lessonia trabeculata kelp beds in Southern Peru: An analysis from the San Juan de Marcona region. Carbon Manag. 2020, 11, 525–532. [Google Scholar] [CrossRef]
- Šulc, R.; Ditl, P. A technical and economic evaluation of two different oxygen sources for a small oxy-combustion unit. J. Clean. Prod. 2021, 309, 127427. [Google Scholar] [CrossRef]
- Paraguay-Delgado, F.; Carreno-Gallardo, C.; Estrada-Guel, L.; Zabala-Arceo, A.; Alexander Martinez-Rodriguez, H.; Lardizabal-Gutierrez, D. Pelagic Sargassum spp. capture CO2 and produce calcite. Environ. Sci. Pollut. Res. 2020, 27, 25794–25800. [Google Scholar] [CrossRef] [PubMed]
- Muller-Karger, F.E. The importance of continental margins in the global carbon cycle. Geophys. Res. Lett. 2005, 32, L01602. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Liu, H.; Gao, Y.; Jiang, Z.; Lin, F.; Chang, L.; Zhang, Y. Food web structure and ecosystem attributes of integrated multi-trophic aquaculture waters in Sanggou Bay. Aquacult. Rep. 2020, 16, 100279. [Google Scholar] [CrossRef]
- Clark, M.S.; Peck, L.S.; Arivalagan, J.; Backeljau, T.; Berland, S.; Cardoso, J.C.R.; Caurcel, C.; Chapelle, G.; De Noia, M.; Dupont, S.; et al. Deciphering mollusc shell production: The roles of genetic mechanisms through to ecology, aquaculture and biomimetics. Biol. Rev. 2022, 95, 1812–1837. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, H. Variation in population structure and standing stocks of kelp along multiple environmental gradients and implications for ecosystem services. Front. Mar. Sci 2021, 8, 578629. [Google Scholar] [CrossRef]
- Theuerkauf, S.J.; Barrett, L.T.; Alleway, H.K.; Costa Pierce, B.A.; St. Gelais, A.; Jones, R.C. Habitat value of bivalve shellfish and seaweed aquaculture for fish and invertebrates: Pathways, synthesis and next steps. Rev. Aquacult. 2022, 14, 54–72. [Google Scholar] [CrossRef]
- Zheng, Y.; Jin, R.; Zhang, X.; Wang, Q.; Wu, J. The considerable environmental benefits of seaweed aquaculture in China. Stoch. Environ. Res. Risk Assess. 2019, 33, 1203–1221. [Google Scholar] [CrossRef]
- Zhang, C.; Ju, W.; Chen, J.; Fang, M.; Wu, M.; Chang, X.; Wang, T.; Wang, X. Sustained biomass carbon sequestration by China’s forests from 2010 to 2050. Forests 2018, 9, 689. [Google Scholar] [CrossRef] [Green Version]
- Xiao, D.; Deng, L.; Kim, D.G.; Huang, C.; Tian, K. Carbon budgets of wetland ecosystems in China. Glob. Change Biol. 2019, 25, 2061–2076. [Google Scholar] [CrossRef]
- Jiao, N.; Liang, Y.; Zhang, Y.; Liu, J.; Zhang, Y.; Zhang, R.; Zhao, M.; Dai, M.; Zhai, W.; Gao, K.; et al. Carbon pools and fluxes in the China Seas and adjacent oceans. Sci. China Earth Sci. 2018, 61, 1535–1563. [Google Scholar] [CrossRef]
- Gao, G.; Gao, L.; Jiang, M.; Jian, A.; He, L. The potential of seaweed cultivation to achieve carbon neutrality and mitigate deoxygenation and eutrophication. Environ. Res. Lett. 2022, 17, 14018. [Google Scholar] [CrossRef]
- Pedersen, M.F. Carbon sequestration potential increased by incomplete anaerobic decomposition of kelp detritus. Mar. Ecol. Prog. Ser. 2021, 660, 53–67. [Google Scholar] [CrossRef]
- Krause-Jensen, D. Substantial role of macroalgae in marine carbon sequestration. Nat. Geosci. 2016, 9, 737–742. [Google Scholar] [CrossRef]
- Bourque, A.S. Impacts of physical disturbance on ecosystem structure in subtropical seagrass meadows. Mar. Ecol. Prog. Ser. 2015, 540, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Jiao, N.; Robinson, C.; Azam, F.; Thomas, H.; Baltar, F.; Dang, H.; Hardman-Mountford, N.J.; Johnson, M.; Kirchman, D.L.; Koch, B.P.; et al. Mechanisms of microbial carbon sequestration in the ocean: Future research directions. Biogeosciences 2014, 11, 5285–5306. [Google Scholar] [CrossRef] [Green Version]
- Kieber, D.J.; Keene, W.C.; Frossard, A.A.; Long, M.S.; Maben, J.R.; Russell, L.M.; Kinsey, J.D.; Tyssebotn, I.M.B.; Quinn, P.K.; Bates, T.S. Coupled ocean-atmosphere loss of marine refractory dissolved organic carbon. Geophys. Res. Lett. 2016, 43, 2765–2772. [Google Scholar] [CrossRef] [Green Version]
- Tang, Q.; Zhang, J.; Fang, J. Shellfish and seaweed mariculture increase atmospheric CO2 absorption by coastal ecosystems. Mar. Ecol. Prog. Ser. 2011, 424, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Crawford, C.M.; Macleod, C.K.A.; Mitchell, I.M. Effects of shellfish farming on the benthic environment. Aquaculture 2003, 224, 117–140. [Google Scholar] [CrossRef] [Green Version]
- Paine, E.R.; Schmid, M.; Boyd, P.W.; Diaz Pulido, G.; Hurd, C.L. Rate and fate of dissolved organic carbon release by seaweeds: A missing link in the coastal ocean carbon cycle. J. Phycol. 2021, 57, 1375–1391. [Google Scholar] [CrossRef]
- Lovelock, C.E. Dimensions of Blue Carbon and emerging perspectives. Biol. Lett. 2019, 15, 20180781. [Google Scholar] [CrossRef] [PubMed]
- Gagern, A.; Manley, J.; Kapsenberg, L. Ocean-based carbon dioxide removal: A new frontier in the blue economy. Mar. Technol. Soc. J. 2022, 56, 40–48. [Google Scholar] [CrossRef]
- Lu, F.; Hu, H.; Sun, W.; Zhu, J.; Liu, G.; Zhou, W.; Zhang, Q.; Shi, P.; Liu, X.; Wu, X.; et al. Effects of national ecological restoration projects on carbon sequestration in China from 2001 to 2010. Proc. Natl. Acad. Sci. USA 2018, 115, 4039–4044. [Google Scholar] [CrossRef] [Green Version]
- Gjedrem, T.; Rye, M. Selection response in fish and shellfish: A review. Rev. Aquacult. 2018, 10, 168–179. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Y.; Liang, Y.; Chen, J.; Zhu, Y.; Zhao, Y.; Jiao, N. Impacts of maricultural activities on characteristics of dissolved organic carbon and nutrients in a typical raft-culture area of the Yellow Sea, North China. Mar. Pollut. Bull. 2018, 137, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; Shi, R.; Qi, Z.; Huang, H.; Liang, Q.; Liu, H. Interactive effects of oyster and seaweed on seawater dissolved inorganic carbon systems: Implications for integrated multi-trophic aquaculture. Aquacult. Environ. Interact. 2017, 9, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wu, W.; Ren, J.S.; Lin, F. A model for the growth of mariculture kelp Saccharina japonica in Sanggou Bay, China. Aquacult. Environ. Interact. 2016, 8, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Reid, G.K.; Lefebvre, S.; Filgueira, R.; Robinson, S.M.C.; Broch, O.J.; Dumas, A.; Chopin, T.B.R. Performance measures and models for open-water integrated multi-trophic aquaculture. Rev. Aquacult. 2019, 12, 47–75. [Google Scholar] [CrossRef]
- Baumann, M.; Taucher, J.; Paul, A.J.; Heinemann, M.; Vanharanta, M.; Bach, L.T.; Spilling, K.; Ortiz, J.; Aristegui, J.; Hernandez-Hernandez, N.; et al. Effect of intensity and mode of artificial upwelling on particle flux and carbon export. Front. Mar. Sci. 2021, 8, 742142. [Google Scholar] [CrossRef]
- Pan, Y.; Fan, W.; Huang, T.; Wang, S.; Chen, C.A. Evaluation of the sinks and sources of atmospheric CO2 by artificial upwelling. Sci. Total Environ. 2015, 511, 692–702. [Google Scholar] [CrossRef]
- Fan, W.; Zhang, Z.; Yao, Z.; Xiao, C.; Zhang, Y.; Zhang, Y.; Liu, J.; Di, Y.; Chen, Y.; Pan, Y. A sea trial of enhancing carbon removal from Chinese coastal waters by stimulating seaweed cultivation through artificial upwelling. Appl. Ocean Res. 2020, 101, 102260. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Dry Mass Specific Gravity | Carbon Contents (%) | |||
|---|---|---|---|---|---|
| Crassostrea gigas | 65.10 | 93.86 | 6.14 | 12.68 | 45.89 |
| Busycon canaliculatu | 64.21 | 87.91 | 12.09 | 11.98 | 44.99 |
| Scapharca subcrenata | 64.21 | 53.47 | 46.53 | 11.29 | 45.86 |
| Mytilus edulis | 75.28 | 91.53 | 8.47 | 11.76 | 44.40 |
| Pinna rudis Linnaeus | 64.21 | 88.59 | 11.41 | 11.44 | 43.87 |
| Chlamys farreri | 63.89 | 85.65 | 14.35 | 11.40 | 43.90 |
| Ruditapes philippinarum | 52.55 | 98.02 | 1.98 | 11.52 | 44.90 |
| Sinonovacula constricta | 70.48 | 96.74 | 3.26 | 13.24 | 44.99 |
| Species | ||
|---|---|---|
| Laminaria japonica | 20.00 | 31.20 |
| Undaria pinnatifida Suringar | 20.00 | 30.70 |
| Porphyra | 20.00 | 27.39 |
| Gracilaria ferox | 20.00 | 20.60 |
| Others 1 | 20.00 | 27.76 |
| Year | Market Price of Mariculture Products | Economic Value/Million Dollars | Product Value | Carbon Storage Value | Oxygen Release Value | |||
|---|---|---|---|---|---|---|---|---|
| Value Quantity/Million Dollars | Proportion /% | Value Quantity/Million Dollars | Proportion /% | Value Quantity/Million Dollars | Proportion/% | |||
| 2003 | 3.13 | 35,657.37 | 35,143.61 | 98.56 | 491.39 | 1.38 | 22.37 | 0.06 |
| 2004 | 3.41 | 40,469.99 | 39,934.99 | 98.68 | 511.25 | 1.26 | 23.74 | 0.06 |
| 2005 | 3.55 | 43,805.81 | 43,247.36 | 98.73 | 532.98 | 1.22 | 25.47 | 0.06 |
| 2006 | 3.82 | 48,839.94 | 48,269.19 | 98.83 | 546.02 | 1.12 | 24.73 | 0.05 |
| 2007 | 4.20 | 47,992.30 | 47,472.30 | 98.92 | 498.05 | 1.04 | 21.95 | 0.05 |
| 2008 | 4.50 | 52,073.67 | 51,554.21 | 99.00 | 497.31 | 0.96 | 22.15 | 0.04 |
| 2009 | 4.43 | 53,593.21 | 53,046.02 | 98.98 | 523.83 | 0.98 | 23.36 | 0.04 |
| 2010 | 4.87 | 62,096.86 | 61,511.48 | 99.06 | 561.50 | 0.90 | 23.88 | 0.04 |
| 2011 | 5.43 | 71,993.01 | 71,400.78 | 99.18 | 566.89 | 0.79 | 25.34 | 0.04 |
| 2012 | 5.49 | 76,604.59 | 75,982.69 | 99.19 | 593.45 | 0.77 | 28.45 | 0.04 |
| 2013 | 5.53 | 81,263.03 | 80,603.59 | 99.19 | 629.56 | 0.77 | 29.88 | 0.04 |
| 2014 | 5.63 | 86,168.12 | 85,464.43 | 99.18 | 666.01 | 0.77 | 37.68 | 0.04 |
| 2015 | 5.69 | 89,912.11 | 89,180.51 | 99.19 | 692.93 | 0.77 | 38.67 | 0.04 |
| 2016 | 5.93 | 97,814.06 | 97,052.34 | 99.22 | 722.18 | 0.74 | 39.55 | 0.04 |
| 2017 | 6.39 | 106,873.66 | 106,092.27 | 99.27 | 740.12 | 0.69 | 41.27 | 0.04 |
| 2018 | 6.48 | 109,571.06 | 108,780.98 | 99.28 | 747.13 | 0.68 | 42.95 | 0.04 |
| 2019 | 6.30 | 107,431.64 | 106,637.33 | 99.26 | 748.28 | 0.70 | 46.03 | 0.04 |
| Total | / | 1,212,160.44 | 1,201,374.08 | / | 10,268.89 | / | 517.47 | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, Q.; Ma, J.; He, F.; Zhang, A.; Pei, D.; Yu, M. Current and Future Potential of Shellfish and Algae Mariculture Carbon Sinks in China. Int. J. Environ. Res. Public Health 2022, 19, 8873. https://doi.org/10.3390/ijerph19148873
Lai Q, Ma J, He F, Zhang A, Pei D, Yu M. Current and Future Potential of Shellfish and Algae Mariculture Carbon Sinks in China. International Journal of Environmental Research and Public Health. 2022; 19(14):8873. https://doi.org/10.3390/ijerph19148873
Chicago/Turabian StyleLai, Qiuying, Jie Ma, Fei He, Aiguo Zhang, Dongyan Pei, and Minghui Yu. 2022. "Current and Future Potential of Shellfish and Algae Mariculture Carbon Sinks in China" International Journal of Environmental Research and Public Health 19, no. 14: 8873. https://doi.org/10.3390/ijerph19148873
APA StyleLai, Q., Ma, J., He, F., Zhang, A., Pei, D., & Yu, M. (2022). Current and Future Potential of Shellfish and Algae Mariculture Carbon Sinks in China. International Journal of Environmental Research and Public Health, 19(14), 8873. https://doi.org/10.3390/ijerph19148873