
Spatial Aggregation and Biometric Variability of the Grass Calamagrostis epigejos (L.) Roth during Different Expansion Stages in Mesic Mountain Meadows


Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Statistical Analysis
3. Results
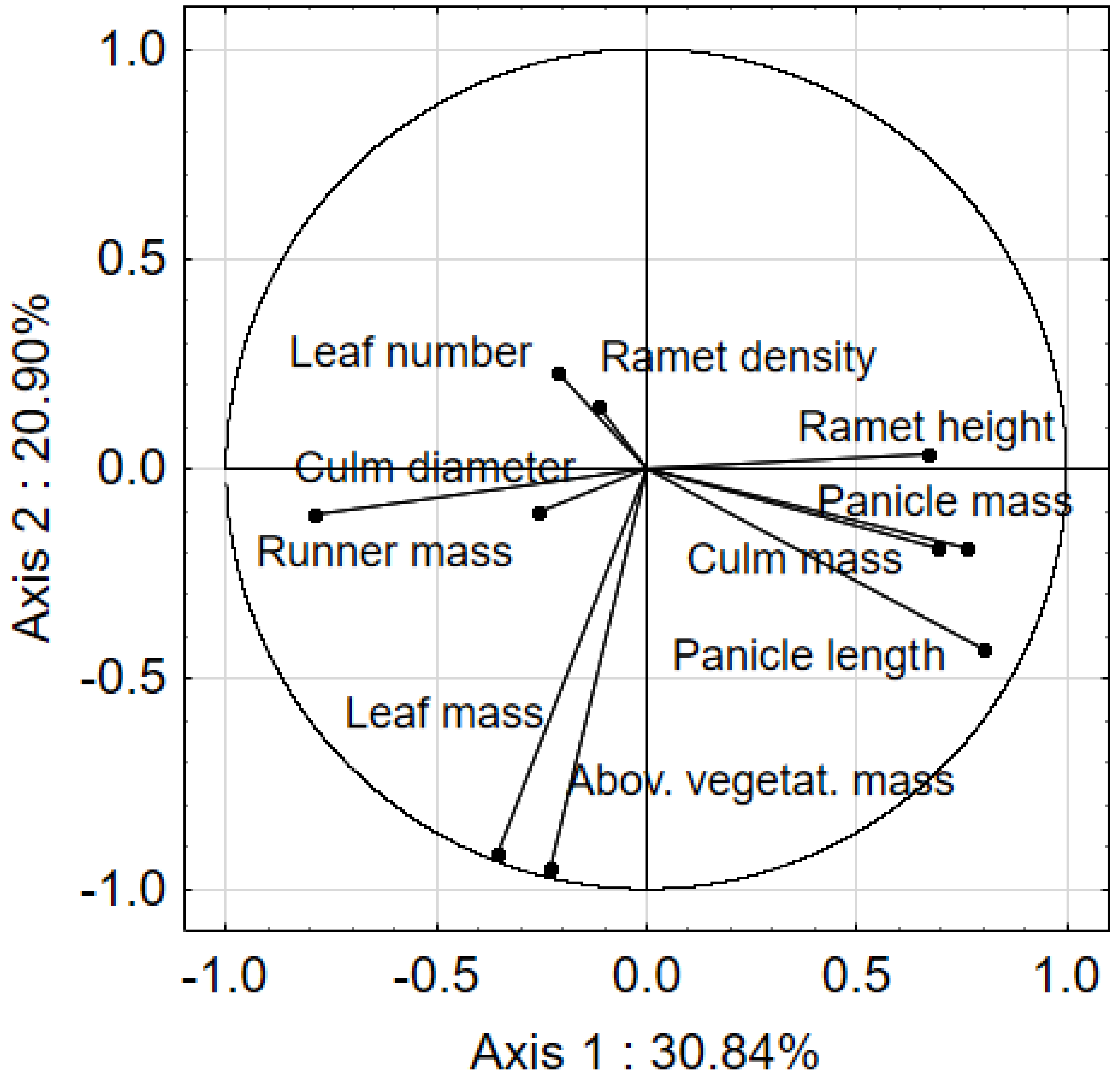
3.1. Clonal Architecture of Calamagrostis epigejos
3.2. Relation between Clone Architecture and Species Diversity in Various Spatial Scales
3.3. Relationship between Environmental Variables and Clonal Architecture
3.4. Spatial Pattern of Calamagrostis epigejos Ramets
4. Discussion
4.1. Clonal Architecture of Calamagrostis epigejos
4.2. Influence of Environmental Factors on Clonal Architecture
4.3. Spatial Pattern of Calamagrostis epigejos Ramets and Its Influence on Species Diversity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berger, U.; Piou, C.; Schiffers, K.; Grimm, V. Competition among plants: Concepts, individual-based modeling approaches, and a proposal for a future research strategy. Perspect. Plant Ecol. Evol. Syst. 2008, 9, 121–135. [Google Scholar] [CrossRef]
- Lovett-Doust, L. Population dynamics and local specialization in a clonal perennial (Ranunculus repens). J. Ecol. 1981, 69, 743–755. [Google Scholar] [CrossRef]
- Schmid, B.; Harper, J.L. Clonal growth in grassland perennials: I. Density and pattern-dependent competition between plants with different growth forms. J. Ecol. 1985, 73, 793–808. [Google Scholar] [CrossRef]
- Sackville Hamilton, N.R.; Schmid, B.; Harper, J.L. Life history concepts and the population biology of clonal organisms. Proc. R. Soc. London 1987, 232, 35–57. [Google Scholar]
- Humphrey, L.D.; Pyke, D.A. Demographic and growth responses of a guerrilla and a phalanx perennial grass in competitive mixtures. J. Ecol. 1998, 86, 854–865. [Google Scholar] [CrossRef]
- De Kroon, H.; Hutchings, M.J. Morphological plasticity in clonal plants: The foraging concept reconsidered. J. Ecol. 1995, 83, 143–152. [Google Scholar] [CrossRef]
- Rebele, F.; Lehmann, C. Biological Flora of Central Europe: Calamagrostis epigejos (L.) Roth. Flora 2001, 196, 325–344. [Google Scholar] [CrossRef]
- Żołnierz, L. Grasslands on Serpentines in Lower Silesia (SW Poland)—Some Aspects of Their Ecology. Wydaw. Uniw. Przyr. We Wrocławiu 2007, 557, 231, (In Polish with English Outline). Available online: https://www.dbc.wroc.pl/dlibra/publication/4380/edition/4255/content?ref=L3B1YmxpY2F0aW9uLzYxNzIvZWRpdGlvbi81NzQ3 (accessed on 1 March 2022).
- Pruchniewicz, D. Abandonment of traditionally managed mesic mountain meadows affects plant species composition and diversity. Basic Appl. Ecol. 2017, 20, 10–18. [Google Scholar] [CrossRef]
- Pruchniewicz, D.; Żołnierz, L. The influence of Calamagrostis epigejos expansion on the species composition and soil properties of mountain mesic meadows. Acta Soc. Bot. Pol. 2017, 86, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pruchniewicz, D.; Żołnierz, L. Relationship between litter produced by Calamagrostis epigejos and seedling recruitment of mesic meadow species in mountain conditions. Commun. Ecol. 2017, 18, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Aiken, S.G.; Lefkovitch, L.P.; Armstrong, K.C. Calamagrostis epigejos (Poaceae) in North America, especially Ontario. Can. J. Bot. 1989, 67, 3205–3218. [Google Scholar] [CrossRef]
- Sedláková, I.; Fiala, K. Ecological problems of degradation of alluvial meadows due to expanding Calamagrostis epigejos. Ekologia 2001, 20, 226–233. [Google Scholar]
- Süß, K.; Storm, C.; Zehm, A.; Schwabe, A. Succession inland sand ecosystems: Which factors determine the occurrenceof the tall grass species Calamagrostis epigejos (L.) Roth and Stippa capillata L. Plant Biol. 2004, 6, 465–476. [Google Scholar]
- Gloser, V.; Košvancová, M.; Gloser, J. Changes in growth parameters and content of N-storage compounds in roots and rhizomes of Calamagrostis epigejos after repeated defoliation. Biol. Bratisl. 2004, 59 (Suppl. 13), 179. [Google Scholar]
- Pruchniewicz, D.; Żołnierz, L.; Andonovski, V. Habitat factors influencing competitive ability of Calamagrostis epigejos (L.) Roth in mountain plant communities. Turk. J. Bot. 2017, 41, 579–587. [Google Scholar] [CrossRef]
- Baležentienė, L.; Šėžienė, V. Germination Responses on Phenols Content of Pine Forest Clear Cutting Dominants. Rural. Dev. Sustain. Dev. For. 2009, 2009, 161–164. [Google Scholar]
- Pruchniewicz, D.; Halarewicz, A. Allelopathic effects of wood small-reed (Calamagrostis epigejos) on germination and growth of selected grassland species. Pol. J. Ecol. 2019, 67, 122–136. [Google Scholar] [CrossRef]
- Pruchniewicz, D.; Żołnierz, L. The influence of environmental factors and management methods on the vegetation of mesic grasslands in a central European mountain range. Flora 2014, 209, 687–692. [Google Scholar] [CrossRef]
- Klimeš, L.; Klimešová, J. A database of clonal plants in central Europe. Plant Ecol. 1999, 141, 9–19. [Google Scholar] [CrossRef]
- Pottier, J.; Evette, A. On the Relationship Between Clonal Traits and Small-Scale Spatial Patterns of Three Dominant Grasses and its Consequences on Community Diversity. Folia Geobot. 2010, 45, 59–75. [Google Scholar] [CrossRef]
- Gloser, V.; Košvancová, M.; Gloser, J. Regrowth dynamics of Calamagrostis epigejos after defoliation as affected by nitrogen availability. Biol. Plant. 2007, 51, 501–506. [Google Scholar] [CrossRef]
- Allen, S.E. Chemical Analysis of Ecological Materials, 2nd ed.; Blackwell Scientific Publications: Oxford, UK; London, UK; Edinburgh, UK, 1989. [Google Scholar]
- Radojević, M.; Bashkin, V.N. Practical Environmental Analysis; The Royal Society of Chemistry: Cambridge, UK, 2006. [Google Scholar]
- McCune, B. Improved estimates of incident radiation and heat load using non-parametric regression against topographic variables. J. Veg. Sci. 2007, 18, 751–754. [Google Scholar] [CrossRef]
- TIBCO Software Inc. Statistica (Data Analysis Software System), 2017, Version 13. Available online: http://statistica.io (accessed on 1 March 2022).
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, PBC: Boston, MA, USA, 2022; Available online: http://www.rstudio.com/ (accessed on 1 March 2022).
- GeoDa. Available online: https://geodacenter.github.io/download.html (accessed on 1 March 2022).
- Aubry, P.; Piégay, H. Pratique de L’analyse de L’autocorrélation Spatiale en Géomorphologie: Definitions Opératoires et Tests. Géogr. Phys. Quat. 2001, 55, 111–129. [Google Scholar]
- Kovach Computing Services. MVSP v3.131; Kovach Computing Services: Anglesey, UK, 2004. [Google Scholar]
- Marshall, C.; Price, E.A.C. Sectoriality and its implications for physiological integration. In The Ecology and Evolution of Clonal Plants; De Kroon, H., van Groenendael, J., Eds.; Backhuys Publishers: Leiden, The Netherlands, 1997; pp. 79–107. [Google Scholar]
- Price, J.N.; Hiiesalu, I.; Gerhold, P.; Pärtel, M. Small-scale grassland assembly patterns differ above and below the soil surface. Ecology 2012, 93, 1290–1296. [Google Scholar] [CrossRef] [PubMed]
- Gloser, V.; Gloser, J. Nitrogen and base cation uptake in seedlings of Acer pseudoplatanus and Calamagrostis villosa exposed to an acidified environment. Plant Soil 2000, 226, 71–77. [Google Scholar] [CrossRef]
- Rebele, F. Konkurrenz und Koexistenz bei ausdauernden Ruderalpflanzen; Verlag Dr. Kovac: Hamburg, Germany, 1996. [Google Scholar]
- Lammerts, E.J.; Pegtel, D.M.; Grootjans, A.P.; van der Veen, A. Nutrient limitation and vegetation changes in a coastal dune slack. J. Veg. Sci. 1999, 10, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, C.; Rebele, F. Phenotypic plasticity in Calamagrostis epigejos (Poaceae): Response capacities of genotypes from different populations of contrasting habitats to a range of soil fertility. Acta Oecol. 2005, 28, 127–140. [Google Scholar]
- Tůma, I.; Holub, P.; Fiala, K. Soil nutrient heterogeneity and competitive ability of three grass species (Festuca ovina, Arrhenatherum elatius and Calamagrostis epigejos) in experimental conditions. Biologia 2009, 64, 694–704. [Google Scholar] [CrossRef]
- Holub, P.; Sedláková, I.; Fiala, K.; Tůma, I.; Záhora, J.; Tesařová, M. Reasons and consequences of expansion of Calamagrostis epigejos in meadows of the Dyje river floodplain. Verh. Ges. Okol. 2004, 34, 167. [Google Scholar]


{kind=link}
{kind=link}
| Stage of Expansion: | |||
|---|---|---|---|
| Initial (1 m) | Intermediate (5 m) | Advanced (10 m) | |
| Ramet density | 219.20 ± 32.62 a | 509.00 ± 53.46 b | 552.90 ± 33.12 b |
| Ramet height | 89.61 ± 6.82 | 92.28 ± 6.47 | 95.86 ± 6.82 |
| Culm mass | 1.14 ± 0.05 | 1.01 ± 0.06 | 1.16 ± 0.09 |
| Culm diameter | 2.31 ± 0.18 | 2.60 ± 0.14 | 2.36 ± 0.13 |
| Leaf mass | 0.73 ± 0.49 | 0.23 ± 0.03 | 0.69 ± 0.46 |
| Leaf number | 2.07 ± 0.08 | 2.20 ± 0.10 | 2.17 ± 0.15 |
| Aboveground vegetative mass | 1.87 ± 0.48 | 1.24 ± 0.07 | 1.84 ± 0.44 |
| Panicle length | 16.90 ± 1.47 | 15.13 ± 1.22 | 16.93 ± 1.26 |
| Panicle mass | 0.37 ± 0.05 | 0.35 ± 0.04 | 0.35 ± 0.05 |
| Runner mass | 347.78 ± 24.14 | 462.72 ± 69.83 | 437.12 ± 56.54 |
| BETA | r | R2 | F | df | p | |
|---|---|---|---|---|---|---|
| Total Number of Species (s) | ||||||
| Ramet density | −0.542 | −0.600 | 0.020 | 5.949 | 4.25 | 0.0017 |
| Ramet culm mass | −0.532 | −0.535 | 0.274 | |||
| BETA | r | R2 | F | df | p | |
|---|---|---|---|---|---|---|
| Calamagrostis epigejos coverage | ||||||
| Graminoid biomass | −0.632 | −0.632 | 0.399 | 18.630 | 1.280 | <0.001 |
| Ramet density | ||||||
| Graminoid biomass | −0.463 | −0.531 | 0.514 | 5.087 | 5.240 | 0.002 |
| pH | −0.667 | −0.576 | ||||
| Ramet height | ||||||
| Ca | 0.282 | 0.412 | 0.822 | 17.770 | 6.230 | <0.001 |
| Necromass | −0.823 | −0.730 | ||||
| Soil depth | 0.879 | 0.670 | ||||
| pH | −0.407 | −0.466 | ||||
| Ramet culm mass | ||||||
| Necromass | −0.709 | −0.591 | 0.451 | 7.120 | 3.260 | 0.001 |
| Soil depth | 1.315 | 0.621 | ||||
| K | −0.795 | −0.467 | ||||
| Culm diameter | ||||||
| Non-significant (ns) | ns | ns | 0.474 | 7.804 | 3.260 | <0.001 |
| Leaf mass | ||||||
| Soil depth | −0.982 | −0.399 | 0.329 | 3.075 | 4.250 | 0.034 |
| Leaf number | ||||||
| N | 0.623 | 0.572 | 0.404 | 5.873 | 3.260 | 0.003 |
| Forbs biomass | 0.444 | 0.447 | ||||
| Heat load | 0.349 | 0.409 | ||||
| Vegetative mass | ||||||
| Non-significant (ns) | ns | ns | 0.225 | 2.524 | 3.260 | 0.080 |
| Panicle length | ||||||
| Heat load | −1.402 | −0.674 | 0.779 | 16.911 | 5.240 | <0.001 |
| pH | 0.287 | 0.394 | ||||
| Panicle mass | ||||||
| Heat-load | −3.164 | −0.719 | 0.789 | 6.130 | 11.180 | <0.001 |
| Necromass | 2.197 | 0.697 | ||||
| K | −0.938 | −0.630 | ||||
| Forbs biomass | 0.524 | 0.555 | ||||
| N | 0.471 | 0.551 | ||||
| Runner mass | ||||||
| pH | −0.673 | −0.770 | 0.863 | 19.783 | 7.220 | <0.001 |
| Heat load | 0.477 | 0.657 | ||||
| Total graminoids and forbs biomass | −0.312 | −0.612 | ||||
| Soil depth | −1.256 | −0.770 | ||||
| K | 0.722 | 0.649 | ||||
| p | −0.564 | −0.549 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pruchniewicz, D.; Żołnierz, L. Spatial Aggregation and Biometric Variability of the Grass Calamagrostis epigejos (L.) Roth during Different Expansion Stages in Mesic Mountain Meadows. Int. J. Environ. Res. Public Health 2022, 19, 7903. https://doi.org/10.3390/ijerph19137903
Pruchniewicz D, Żołnierz L. Spatial Aggregation and Biometric Variability of the Grass Calamagrostis epigejos (L.) Roth during Different Expansion Stages in Mesic Mountain Meadows. International Journal of Environmental Research and Public Health. 2022; 19(13):7903. https://doi.org/10.3390/ijerph19137903
Chicago/Turabian StylePruchniewicz, Daniel, and Ludwik Żołnierz. 2022. "Spatial Aggregation and Biometric Variability of the Grass Calamagrostis epigejos (L.) Roth during Different Expansion Stages in Mesic Mountain Meadows" International Journal of Environmental Research and Public Health 19, no. 13: 7903. https://doi.org/10.3390/ijerph19137903
APA StylePruchniewicz, D., & Żołnierz, L. (2022). Spatial Aggregation and Biometric Variability of the Grass Calamagrostis epigejos (L.) Roth during Different Expansion Stages in Mesic Mountain Meadows. International Journal of Environmental Research and Public Health, 19(13), 7903. https://doi.org/10.3390/ijerph19137903