1. Introduction
Microcystins (MCs) are a group of natural cyclic peptides produced by multiple cyanobacterial species, with more than 279 analogues found so far [
1,
2], of which MC-LR and MC-RR are most common toxins [
3,
4,
5,
6]. According to published reports, MCs are specific hepatotoxic agents capable of causing acute liver damage in wild and domestic animals exposed to high doses and play a role in tumor promotion in long-term tested animals exposed to lower doses of these toxins [
7,
8,
9,
10,
11]. To date, a large number of studies have investigated hepatic toxicity resulting from MCs [
12,
13,
14,
15]. Some studies have also reported nephrotoxicity, reproductive toxicity, and gastrointestinal toxicity caused by MCs in rats, fish and other species [
1,
16,
17,
18,
19,
20,
21,
22,
23]. Regarding nephrotoxicity, whether the kidneys of different animals are all responsive to MCs is unclear.
Rats and mice are common animal models for toxicological studies in mammals. Previous studies reported consistent conclusions to support the nephrotoxicity of MCs in rats [
19,
20,
24,
25,
26,
27,
28,
29,
30,
31,
32,
33,
34,
35], but had different views on mice in terms of whether MCs could injure mouse kidneys. Qin, studying the toxicity of MC-LR in mice, found negative outcomes for the kidney, though they did not exclude possible endoplasmic reticulum stress (EMS) in kidney cells [
26]. Jayaraj’s study reported that no inhibition of protein phosphatase activity was detected in the kidneys of mice under acute MC-LR exposure [
27]. Al-Jassabi’s study suggested that MC-LR resulted in kidney tubule injury and renal oxidative damage in mice [
28]. Xu’s research also suggested that MC-LR induced nephrotoxicity in mice [
29]. In addition, Yi’s study reported histopathological changes in mouse kidney after three months of exposure to MC-LR [
30]. However, most researchers focus on the tissue and cell injury and ignore the metabolic abnormalities. Due to water metabolism and electrolyte balance being tightly associated with kidney function [
31], abnormal water metabolism is usually described as a symptom of kidney impairment and is often omitted when there are no obvious organ changes in the kidney. The effect of MCs on water metabolism in different test animals is unclear. Whether the changes of water intake and output in test animals induced by MCs are definitively caused by kidney damage also needs to be explored.
Over the past decades, most research has been focused on MC-LR and relatively little on MC-RR. These two analogues are cyclic heptapeptides that differ in one amino acid at two variable L-amino acid units (X and Y), MC-LR with a leucine and MC-RR with an arginine respectively [
32]. This chemical differentiation results in differences in their nature and toxicity. Although the biotoxicity of MC-RR is a little bit lower than MC-LR [
33,
34,
35], MC-RR is often detected in bodies of water and accounts for a higher proportion in the analogues. The level of MC-RR in these environments can be larger than MC-LR sometimes [
36,
37], so research on MC-RR for its toxicity is of great importance.
The present investigation aimed to understand the effect of MC-RR on water metabolism in the rat and mouse. Considering water metabolism is relevant to kidney function, the nephrotoxicity of MC-RR in the rat and mouse was also evaluated to explore the possible causes of water metabolic changes. As the liver is the main target of MCs and involves much metabolic reaction, some basic liver-related indices were monitored at the same time. This study was undertaken in two steps: a single dose toxicity test and a continuous exposure experiment, to explore and compare the effects of MC-RR on the two animals’ water metabolism and their related organ tropism.
2. Materials and Methods
2.1. Animals
Specific pathogen free (SPF) male Sprague–Dawley (SD) rats and KunMing (KM) mice were bought from the Laboratory Animal Center of Huazhong University of Science and Technology (Wuhan, China). All animals prior to the experiment were allowed to acclimatize to their surrounding conditions for a week with sufficient food and water ad libitum under a 12 h light/dark cycle and at 25 ± 2 °C room temperature.
2.2. MC-RR Extraction and Purification
MC-RR was extracted from Microcystis blooms occurring in Zhongshan Park, Wuhan (China). The extraction and purification procedures were as follows: 70% methanol aqueous solution was used to extract MCs from the collected cyanobacterial slurry with magnetic stirring at 120 r/min for 40 min at room temperature. The ratio of methanol aqueous solution (mL) to cyanobacterial slurry (g) was 10:1. The extracting solution was centrifuged at 9000× g for 8 min to collect the supernatant. To precipitate the phycocyanin, the supernatant was adjusted to pH 4 and placed stably for 120 min. Then, the phycocyanin was removed by centrifugation and filtration. After the phycocyanin was removed, the supernatant was adjusted to pH 7 and then methanol was removed by rotary evaporation. The crude extract was filtered through 0.45 μm pore size membrane filters to prepare for primary purification. Primary purification was performed by solid phase extraction (SPE) using an Oasis HLB column (Waters, Milford, MA, USA). The columns were activated by methanol and distilled water, and washed by 20 and 30% methanol aqueous solution successively after loading the crude extract. Then, 100% methanol was used to wash the columns for collection of the final eluent. The final eluent was concentrated to 2 mL by rotary evaporation for subsequent steps.
Secondary purification was performed by a Sephadex LH-20 column (500 mm × 26 mm i.d.) with 100% methanol eluted at 1 mL/min flow rate [
38]. The eluent was collected every 5 mL per tube, and tubes that contained MC-RR (identified by high-performance liquid chromatography (HPLC)) were selected for the third purification steps. MC-RR was separated through the third purification process with a DEAE-52 (DEAE-cellulose) column (300 mm × 26 mm i.d.). In this process, distilled water and 3 mmol/L sodium solution eluted the column successively with a flow rate of 1 mL/min. The eluents were identified and quantified by HPLC according to the MC-RR standard (Enzo Life Sciences Inc., Lausen, Switzerland). Eluent with a purity of MC-RR > 90% was collected for use in the animal experiments (
Supplementary Material Figure S1).
2.3. Experimental Procedures on Animals
2.3.1. Single-Dose Toxicity Test
Blocking Tests of Rat Nephrotoxicity
The rats were separated into 5 groups. While being given the high doses of toxins described above, the rats were simultaneously administered different doses of dexamethasone (DXM, purity ≥97%, purchased from Sigma-Aldrich, St. Louis., MO, USA): high dose group DXM 50.0 mg/kg + MC-RR 574.7 μg/kg; medium dose group DXM 10.0 mg/kg + MC-RR 574.7 μg/kg; low dose group DXM 2.0 mg/kg + MC-RR 574.7 μg/kg; positive control group DXM 0.0 mg/kg + MC-RR 574.7 μg/kg, and saline control group. Twenty-four hours after administration, urine, blood and body organ samples were taken for biochemical and pathologic measurement and analysis to understand the changes in the structure and function of the kidney and liver.
2.3.2. Continuous Exposure Experiment of MC-RR in Mice for Water Metabolism
Based on the results of single-dose toxicity of MC-RR in mice, a short-term toxicity test was performed to explore the possible impact on water metabolism and related organs. KM mice were randomly divided into 4 groups, with 35 in each group. The mice were fed in metabolic cages, with 7 in each cage in order to collect the urine effectively. The 4 groups were labeled in terms of toxin dosage given: high dose: 140.0 μg/kg; medium dose: 70.0 μg/kg; low dose: 35.0 μg/kg; and physiological saline as negative control. The animals were given intraperitoneal injection daily for 28 days, and continued to be observed until the 42nd day. The animals’ daily weight and drinking and urine volume were recorded. On the 7th, 14th, 28th, 35th, and 42nd days, 7 subjects in each group were sampled and euthanized for toxicological examination, including urinalysis, sodium and potassium ion levels in urine and serum, antidiuretic hormone (ADH) in blood, serum enzymes, kidney and liver weight, and histological examination.
2.3.3. Sample Collection and Process
The urine volumes were pooled and recorded in each metabolic cage. The urine used for urinalysis, including specific gravity, Na+ and K+, was collected directly from the bladders. Blood samples were obtained from the optical orbits of the animals. The blood was naturally clotted and then centrifuged at 2500× g at 4 °C for 10 min to harvest the serum for following analysis. After blood collection was finished, the animals were euthanized to sample the organs, such as kidneys and liver. The organs were weighed and fixed in 4% polyformaldehyde for further histological examination.
2.4. Urinalysis
For routine urine and some chemical indices, semiquantitative colorimetric reagent strips (dry chemical method) (Acon, Hangzhou, China) were used for detection in urine. Urine specific gravity (USG) was determined using a LSUD-digital urinometer (Guangzhou Mingrui Electronic Technology Co., Ltd, Guangzhou, China).
2.5. Detection of Na+and K+ Levels in Urine and Blood
The urine and blood samples were diluted 200- and 100-fold, respectively, using deionized water prior to instrument detection for Na+ and K+ concentration. Samples were analyzed by an AA240Fs fast sequential atomic absorption spectrometer (Agilent, Santa Clara, CA, USA) coupled with flame atomization and sodium and potassium hollow cathode lamps with the current operating at 5 mA and air/C2H2 as gas form. The instrument working conditions were as follows: wavelength for sodium 589.6 nm, slit width 0.2 nm; wavelength for potassium 769.9 nm, slit width 1.0 nm.
2.6. Serum Biochemical Index Assay
A Mindray BS-200 Chemistry Analyzer (Shenzhen Mindray Bio-Medical Electronics Co., Ltd., Shenzhen, China) was applied for serum index detection, including albumin (ALB), alkaline phosphatase (ALP), alanine aminotransferase (ALT), glutamic oxaloacetic transaminase (aspartate aminotransferase, AST), creatinine (CREA), serum lactate dehydrogenase (LDH), glucose (Glu), total cholesterol (TC), total protein (TP), uric acid (UA), and urea nitrogen (BUN). ADH was measured using ELISA kits (Anoric Bio-technology Co., Ltd., Tianjin, China) according to the manufacturer’s instructions.
2.7. Histopathological Examination
The kidney and liver tissues fixed by polyformaldehyde were embedded in paraffin, cut to a thickness of 2 µm, stained with hematoxylin-eosin (HE), and examined using a light microscope. Periodic acid-Schiff (PAS) stain was applied to the kidney tissues to observe histological changes in glomerular mesangium, or basement membrane.
2.8. Statistical Analysis
Statistical analysis was performed with SPSS for Windows software version 19.0 (SPSS Inc, Chicago, IL, USA). The homogeneity of variances was tested using Levene’s test. One-way analysis of variance (ANOVA) was used for homogeneous data, followed by Dunnett’s test, and Dunnett’s T3 test was performed to compare the heterogeneous data. p < 0.05 was considered to be significant.
4. Discussion
As we know, microcystins are hepatoxins that have been confirmed in many species and reported in a number of studies [
14,
39,
40,
41,
42]. Some papers have reported that the toxins also targeted other organs of tested animals [
24,
43,
44,
45]. However, more data are needed to support firm conclusions on the toxicity to other organs, such as kidneys. The kidney is a critical organ to keep water metabolism normal. Abnormal water metabolism is often identified by water intake and output, and attributed to kidney injury. However, water metabolism is a complicated process that involves several organizational systems. Kidney injury is not the only way to change water metabolism. Previous studies had various conclusions on the nephrotoxicity of MCs, but paid less attention to water metabolic changes. In the present study, the effects of MC-RR on water metabolism and the relevant nephrotoxicity in the two animals (SD rats and KM mice) were examined and compared at the same time.
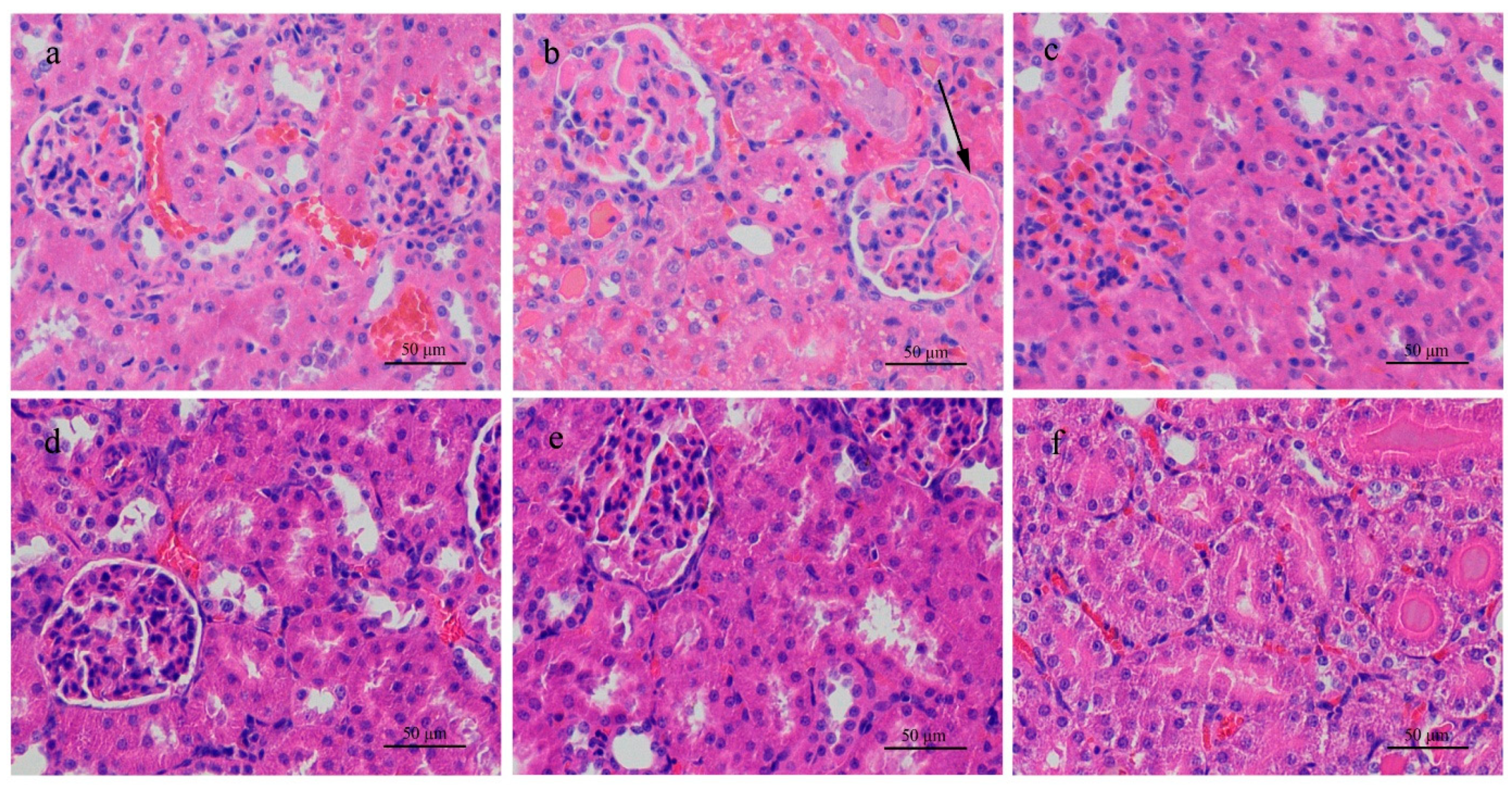
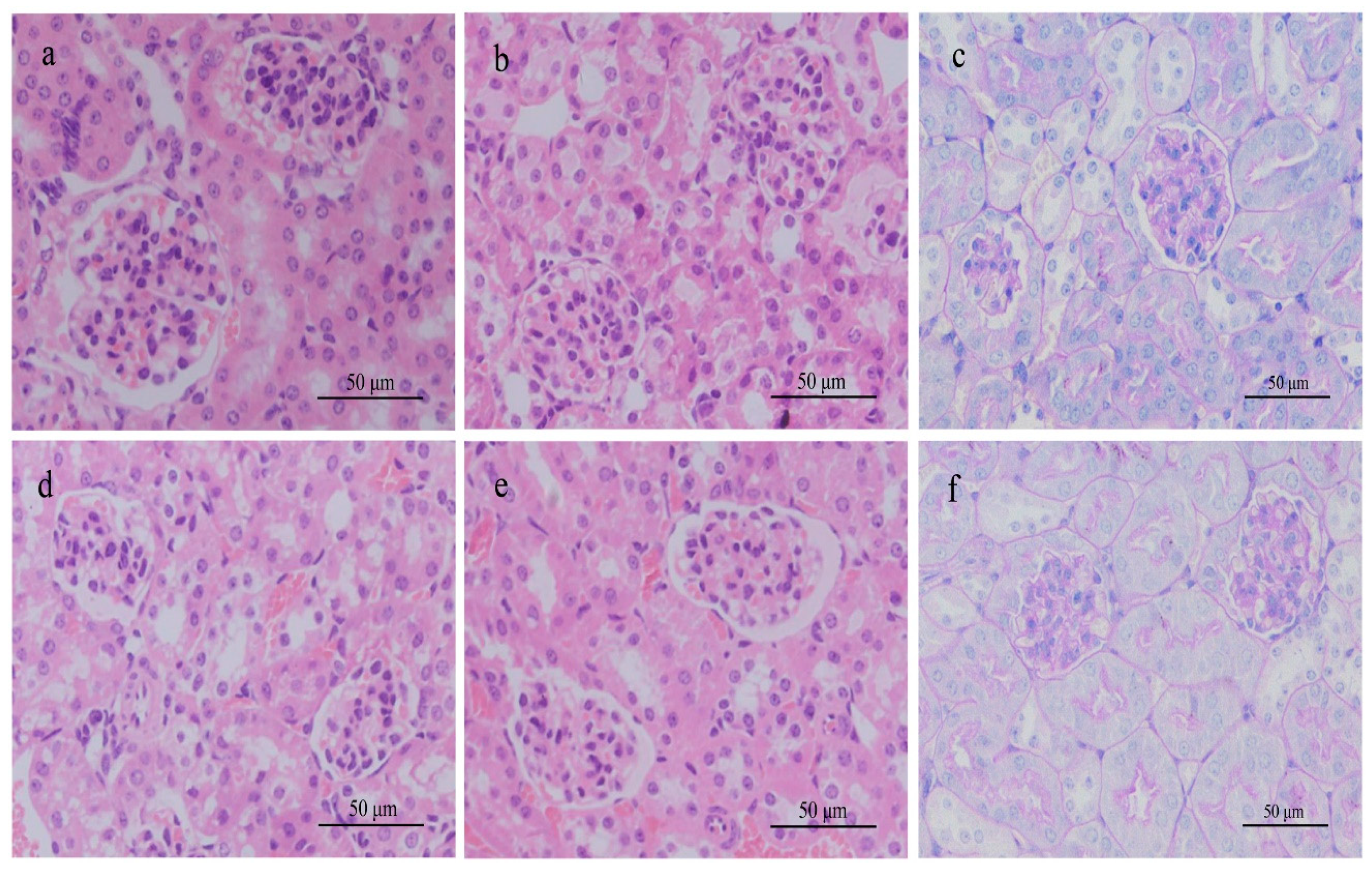
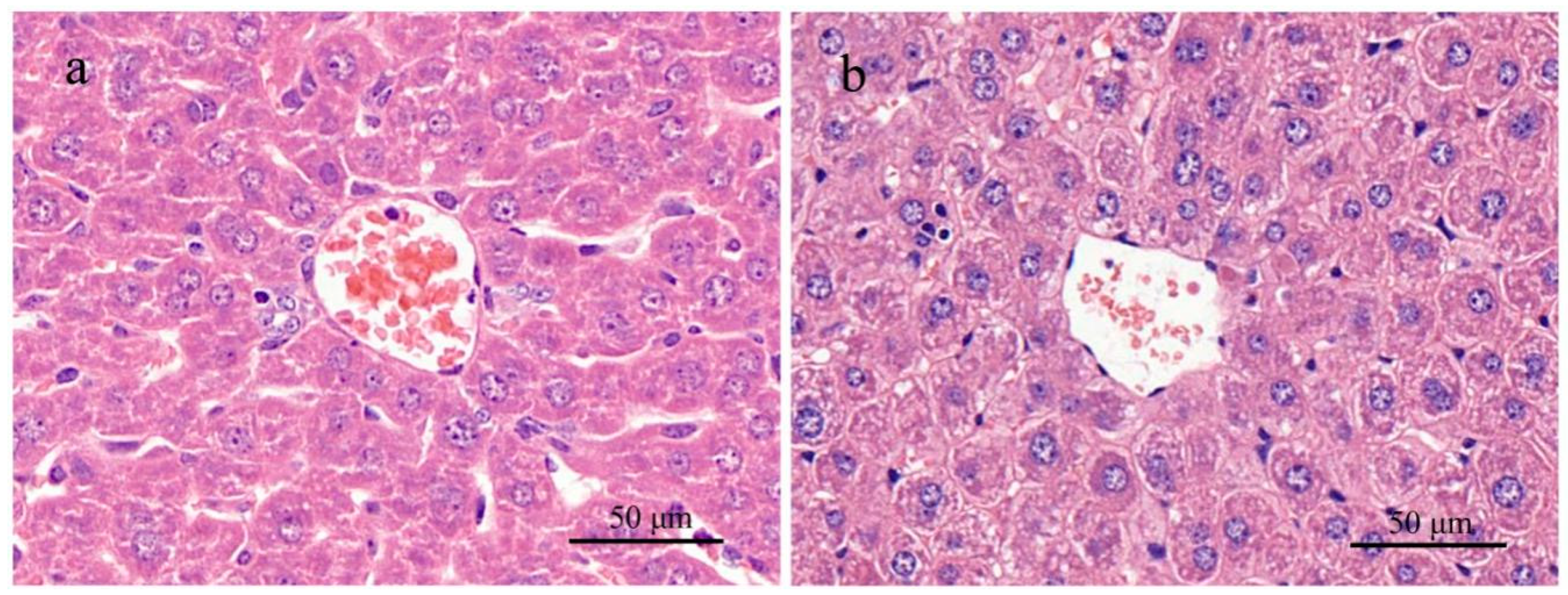
In the single-dose test, the highest dose of the MC-RR was 3/4 LD50 with which we could better observe possible nephrotoxicity and avoid the death of the subjects. Both animals were sensitive to the given toxin and showed a series of pathological changes within 24 h. For example, rats administered 574.7 μg/kg of MC-RR (high dose) showed abnormal behaviors, such as lethargy state and anorexia. However, only rats in the high-dose group showed abnormal water metabolism, polyuria, and polydipsia. The slowed down weight gain in this group was mainly attributed to the increased urine excretion 6 h after administration started and loss of appetite. Together with the unusual urinary excretion, gross hematuria appeared in rats 12 h after administration. This means that damage might be present in the urinary system.
Further urinalysis showed a notable increase in bilirubin/urobilinogen, microalbumin, protein, and creatinine in rats given a high dose, suggesting that the toxin injured both the liver and kidney. The increased bilirubin/urobilinogen in urine came from the liver damage and the increased microalbumin, protein, and creatinine in urine pointed to kidney injury. The histologic examination strongly supported this judgment. Kidney sections stained with H&E from high-dose rats demonstrated various histopathologic changes, such as necrosis and detachment of cells, and destruction of normal kidney structures. Milutinović’s research reported damage to the kidney cortex and medulla of rats with a long-term MC-LR and MC-YR treatment in a single low dose [
25]. Although the kinds of MC and the experimental method in Milutinović’s study differ from this study, kidney damage was observed after MC treatment in both. The kidney tubule is the place where water and salt reabsorption occur [
46]. Damage to the kidney tubules resulted in polyuria and high Na
+ with low K
+ in urine in this study, especially in the high-dose group. Serious polyuria then caused the polydipsia.
In the medium-dose (287.3 μg/kg) group of rats, urine protein and occult blood were detected with histologic alteration of inflammable cell infiltration, but such toxic effect was definitely mild compared to the high-dose group. Although the symptoms and histopathologic changes were not observed in rats given the low dose (143.7 μg/kg) group, the abnormal higher concentration of Na
+ and lower K
+ in urine found in this group may be more sensitive indices that reflected the early or slight damage of the kidney by MC-RR. Within 24 h, from clinical symptoms to histopathology and laboratory analysis, all examination evidence proved that the rat kidneys were targeted by high, medium, and low doses of MC-RR to some extent. These results are similar to Lowe’s report, in which he used only a single dose of MC-LR to observe the nephrotoxicity [
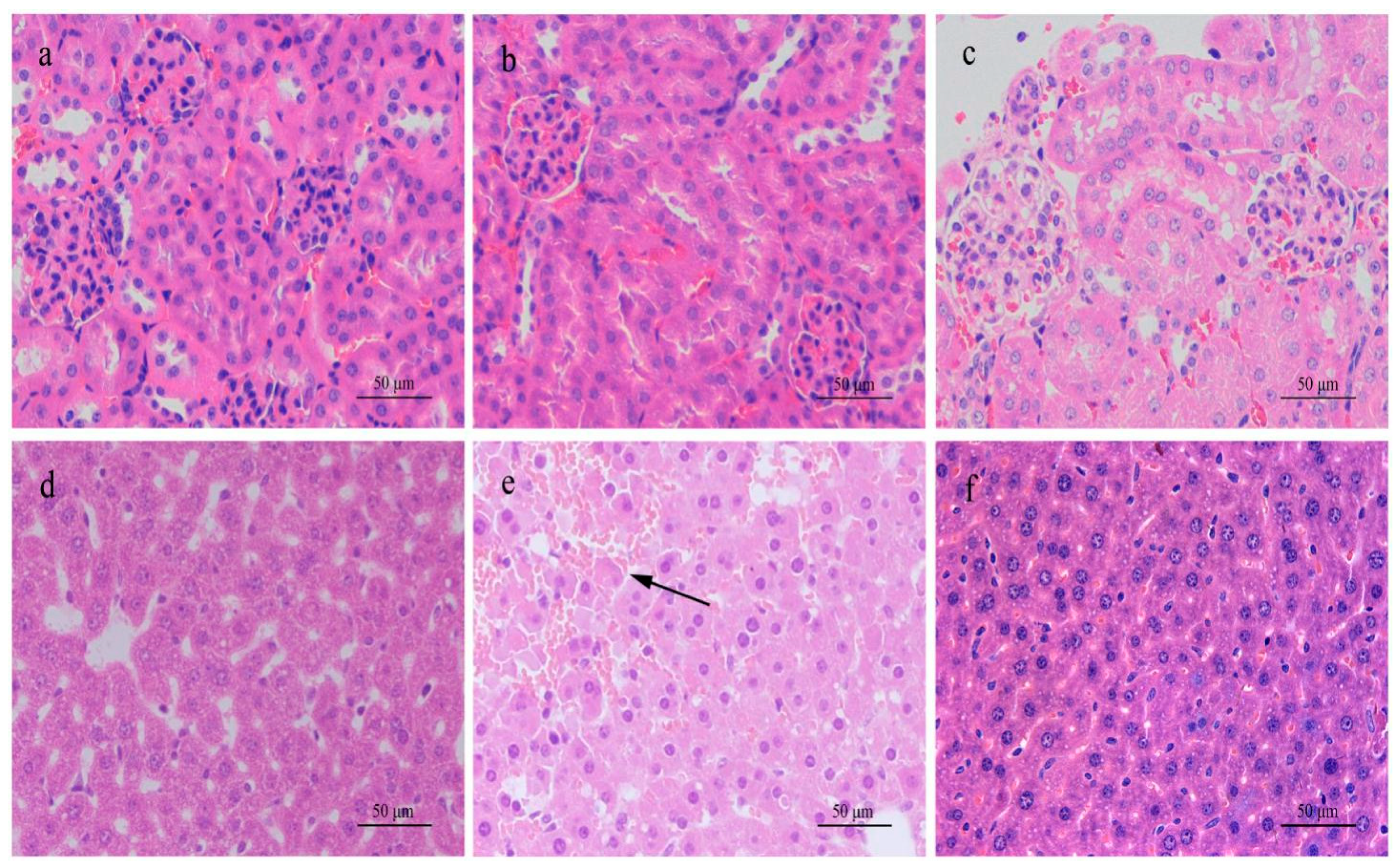
47]. Although the toxin and dosage used were different between his study and this one, the pathologic changes in rats were similar. In this study, gradient dosages were used so that a dose effect could be observed. However, no dysfunction or pathologic alteration in the mouse urinary system and kidney were observed with any toxin dosage given, whereas liver damage was prominent.
The serologic analysis showed that many key enzymatic activities reflecting the function of the kidney and liver in rats were higher or lower in the high- and medium-dose groups than the control, indicating that the related organs were damaged and the enzymes were released into the blood. For mice, nonspecific enzymes, such as ALT, AST, ALP, and LDH were abnormally in the blood serum in the high-dose group, suggested certain organ damage. Combined with the increased TC and decreased GLU in the high-dose group, the liver injury was mostly confirmed. The histopathologic liver changes in the high-dose group also confirmed liver damage. However, no kidney-related specificity index was found to have an abnormal level in mice. The Na+ in blood of the high-dose rat group was lower and the K+ level was higher than the control. Such a trend echoed their levels in urine, but this urine change could be observed in all rat groups, not exclusively limited to the high dose-group. The Na+ and K+ changes in blood lagged behind those in urine. The large amount of water and Na+ lost from the urine in the high-dose group was the main reason why the changes of Na+ and K+ levels in blood in this group were more obvious than in other groups.
In terms of the pathogenic symptoms and possible mechanisms, the intervention measure, DXM, was used to examine the offset effects on pathogenic phenomena in the present investigation. When different doses of DXM (50.0, 10.0, and 2.0 mg/kg) combined with 574.7 μg/kg of MC-RR was administered to rats, the overall pathologic symptoms and histopathologic lesions were improved, showing a good dose-response relationship. These results suggest that DXM, a universal anti-inflammatory drug, could counter the acute damage caused by MC-RR. From another angle, the polydipsia polyuria relief occuring under DXM administration also suggests that the polydipsia polyuria observed in rats was mainly caused by kidney injury. There was corroboration between this part of the results and Nobre’s study, which reported that the DXM could block the increased glomerular filtration rate and urinary flow caused by MC-LR in the isolated perfused rat kidney [
19]. The differences in the kidney effect between rats and mice under single dose MC-RR treatment were obvious and in sharp contrast. Though the high dose for the two animals was 3/4 LD
50, the nephrotoxocity was explicit in rats but was not reflected in mice, implying that SD rat kidney is more sensitive than KM mouse kidney to MC-RR exposure. The reasons for this difference in the two animals have not been fully clarified. Previous studies suggested that the MCs enter the cells or tissues through organic anion transporting polypeptides (Oatps), which include several members [
43,
48,
49,
50]. The kinds of Oatps distributed in rat and mouse kidneys are not exactly the same [
51,
52,
53,
54,
55]. Maybe the types and content of Oatps in the two animals’ kidneys are part of the reasons for the different response to MC-RR.
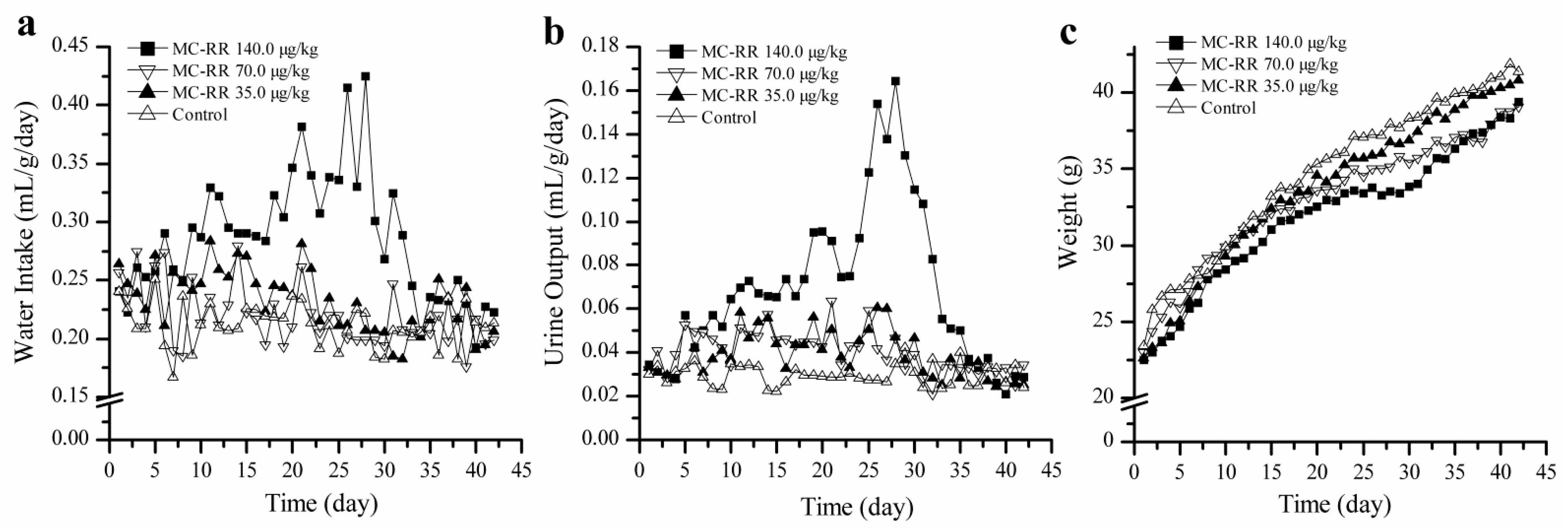
Since abnormal water metabolism in rats was found to be mainly caused by the kidney damage in the single dose toxicity experiment, the continuous exposure test was performed on the mice in the present study. In the continuous exposure experiment, the weight gain of mice in receiving a high dose (140.0 μg/kg) started to slow down on day 9 and water intake increased on the same day, because the urine output was increased on day 8 ahead of the weight loss and increased water intake. The three symptoms became worse with repeated administration until the toxin was stopped on day 29, indicating that there was a problem in water metabolism. It seems that the urinary system and kidney function should be responsible for such pathologic phenomena, since the symptoms were quite like rat intoxication. However, further urine and serum analysis did not support this assumption, because the indices reflecting kidney function in urine and blood were almost all within normal ranges, except specific gravity and levels of Na+ and K+ in mouse urine; in the urine of mice in the high-dose group, they were significant lower than the control simply due to the large volume of urine diluting them.
A different scenario could be seen with rats in this experiment, in that their levels of Na
+ and K
+ were quite opposite to their abnormal level, caused by kidney damage. The histological examination of mice given a high dose showed that the structure of the kidney under the microscope was intact and well arranged just like the control group. Therefore, the evidence to associate kidney damage with water metabolic dysfunction was not yet found in mice, even though the toxicity test extended to the 42nd day. Hooser and Qin’s research may support this part of the results, that no significant kidney injury was found in mice after MC administration in the acute experiment [
56] or long-term exposure [
26], although these studies used MC-LR and paid no attention to the related water metabolism. The dysfunction of water metabolism in rats and mice treated with MC-RR was compared for the first time in the present study.
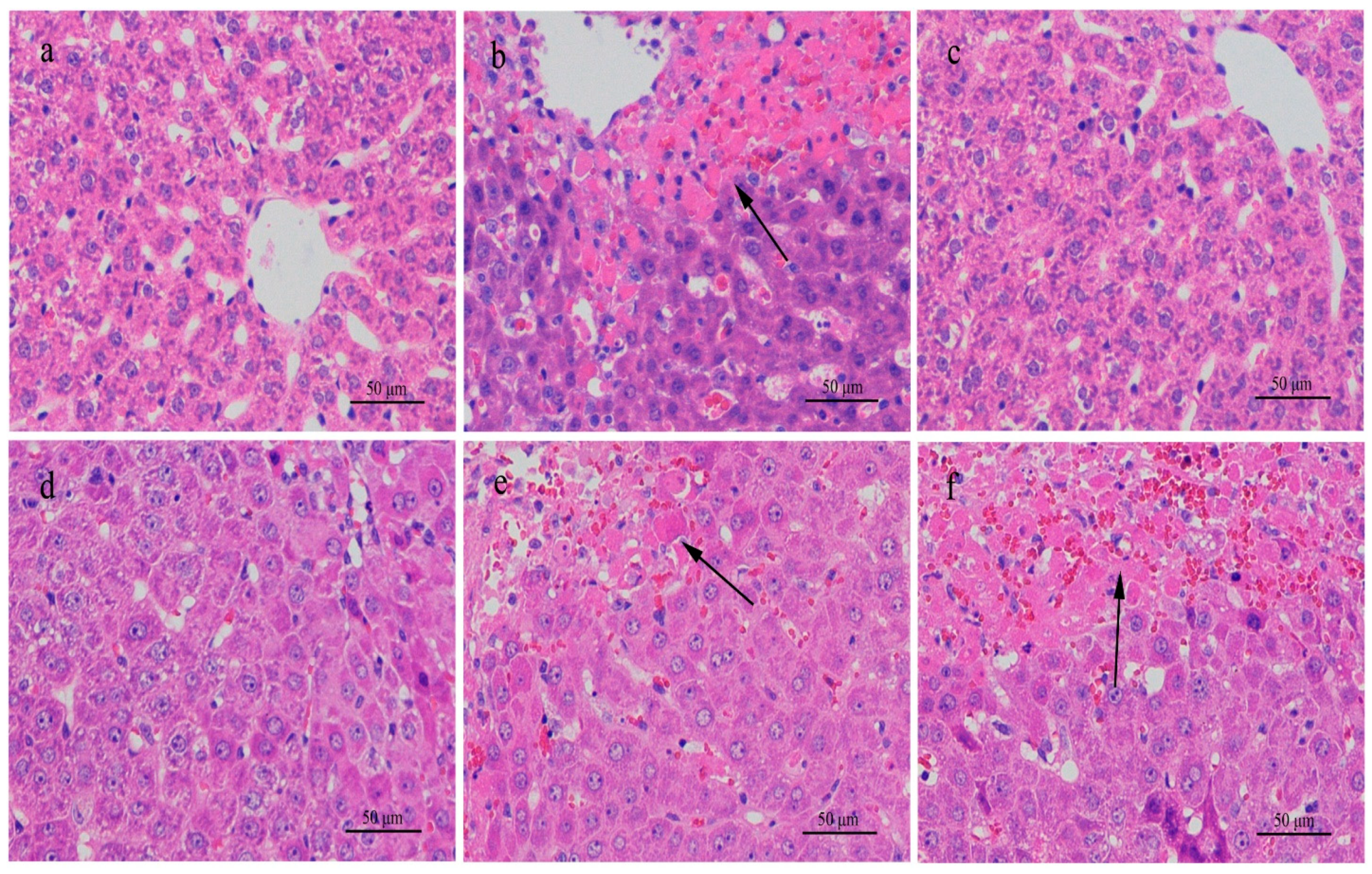
We expected that the toxin would affect the livers of mice in the high-dose group when samples were collected for the first time on day 7, showing increasing serum ALT. Other serum indices (AST, ALP, and TC) started to increase around day 14 and returned to normal or were relieved one or two weeks after administration was stopped on day 29. This trend was also shown in histologic liver changes. When the toxin was given, the liver sections from mice in the high-dose group showed various lesions; however, when the toxin was stopped, the histologic liver changes did not show evident lesions any more. The continuous exposure experiment demonstrated that the liver damage in mice by high-dose toxin was reversible. As long as the mice were not exposed to the MC-RR, all the abnormal values went back to normal levels.
The polydipsia polyuria of the mice in the high-dose group showed the same change trend as the liver damage, suggesting that the abnormal water metabolism was synchronous with the MC-RR exposure and more related to the abnormal liver system rather than the kidney. The changes of ADH in the high-dose group were mainly considered as feedback regulation. Water metabolism involves several organs and systems, not only the kidney [
57]. When water metabolism is found to be abnormal, it implies injury to the relative systems. In Zhong’s study, water metabolism dysfunction induced by MC-RR in mice was thought to be caused by liver damage [
58]. Zhao’s research reported similar polydipsia induced by MC-LR and attributed the polydipsia to the thyroid dysfunction, but they did not pay attention to the urine volume [
59]. Those two reports suggested that the polydipsia was caused by dysfunction of the thyroid or liver, not the kidney. On the other hand, Yi evaluated the influence of chronic exposure to MC-LR on mouse kidney, and found kidney injury after three months [
30]. Thus, although the kidney damage indices were not found in our study, the possibility of dysfunction originating from the kidney cannot be easily excluded. The real mechanism of polydipsia polyuria that occurred in mice intoxicated by MC-RR is not yet clear. The normal level K
+ in blood indicated that the polydipsia polyuria was not a result of hypokalemia and aldosteronism. The low blood glucose levels and increased ADH in mice also exclude diabetes and diabetes insipidus, which are common causes of polydipsia polyuria [
60,
61]. The outcomes from the present study offer two possible causes: dysfunction originating in the kidney without parenchymal damage, and liver-related metabolic dysfunction. More extensive and detailed research is needed to find the mechanism.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}