The 4R’s Framework of Nutritional Strategies for Post-Exercise Recovery: A Review with Emphasis on New Generation of Carbohydrates

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
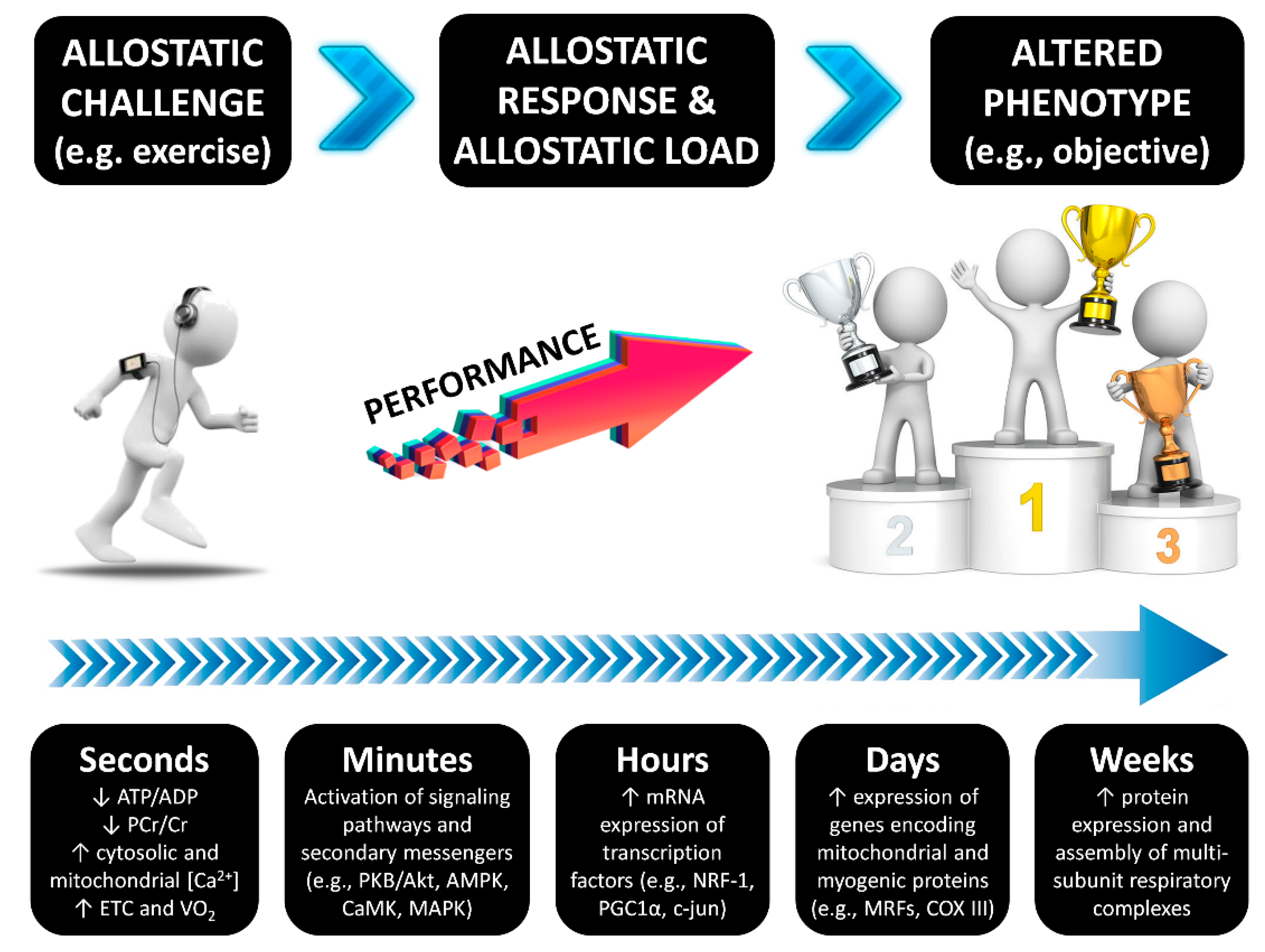
1. Introduction
2. Methods
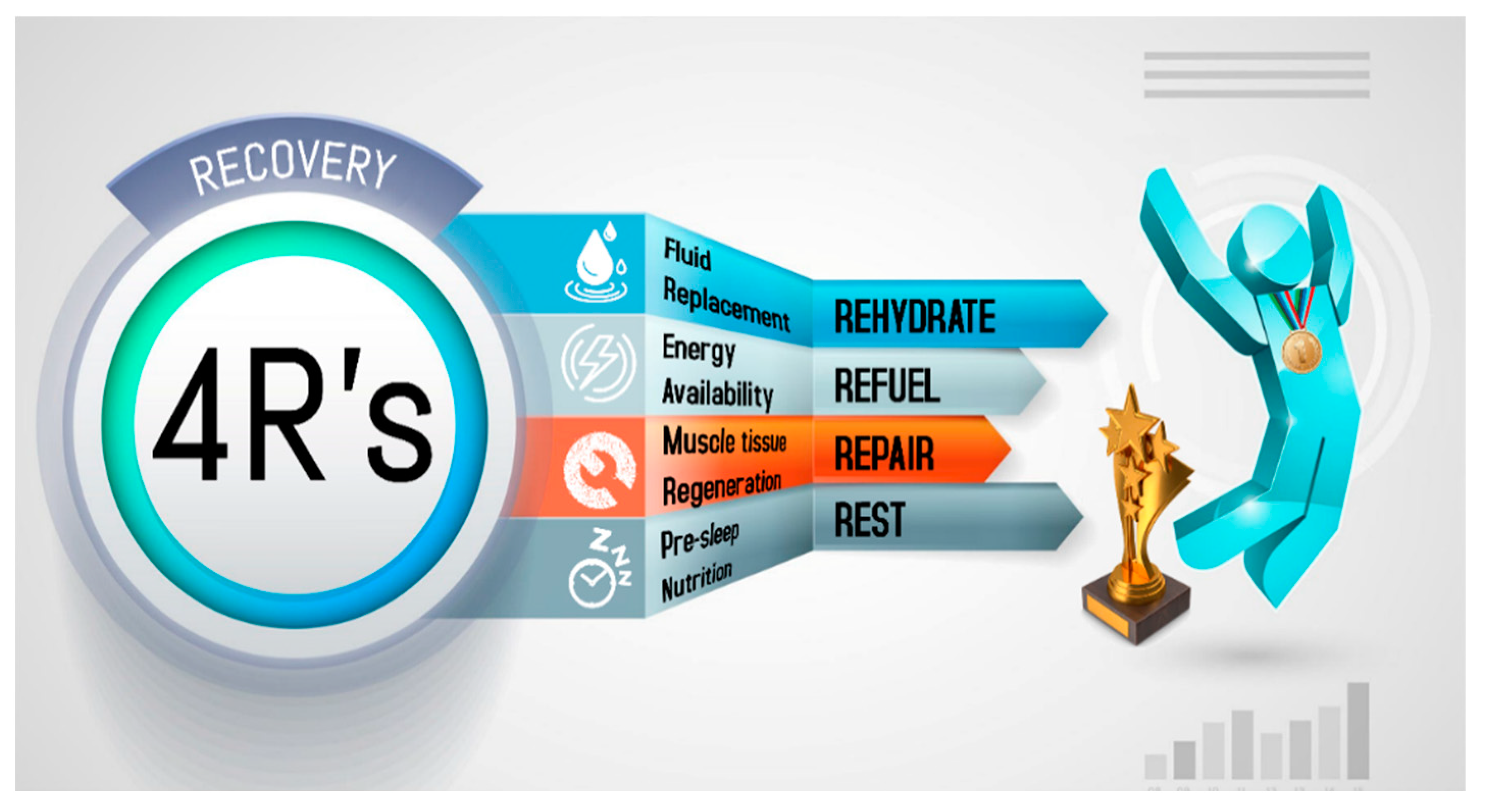
3. Results and Discussion
3.1. Rehydrate
3.2. Refuel
- Time to next training session or competition,
- Nutrition periodization to achieve adaptations,
- Need for muscle repair and growth,
- The amount consumed before and after as part of global requirements.
- Moderate duration/low-intensity training (e.g., 2–3 h per day of intense exercise performed 5–6 times per week): 5–8 g·kg−1 body mass·day−1
- Moderate to heavy endurance training (e.g., 3–6 h per day of intense training in 1–2 daily workouts for 5–6 days per week): 8–10 g·kg−1 body mass·day−1
- Extreme exercise programs or competition (+6 h per day or high competition frequency during the week): 10–12 + g·kg−1 body mass·day−1
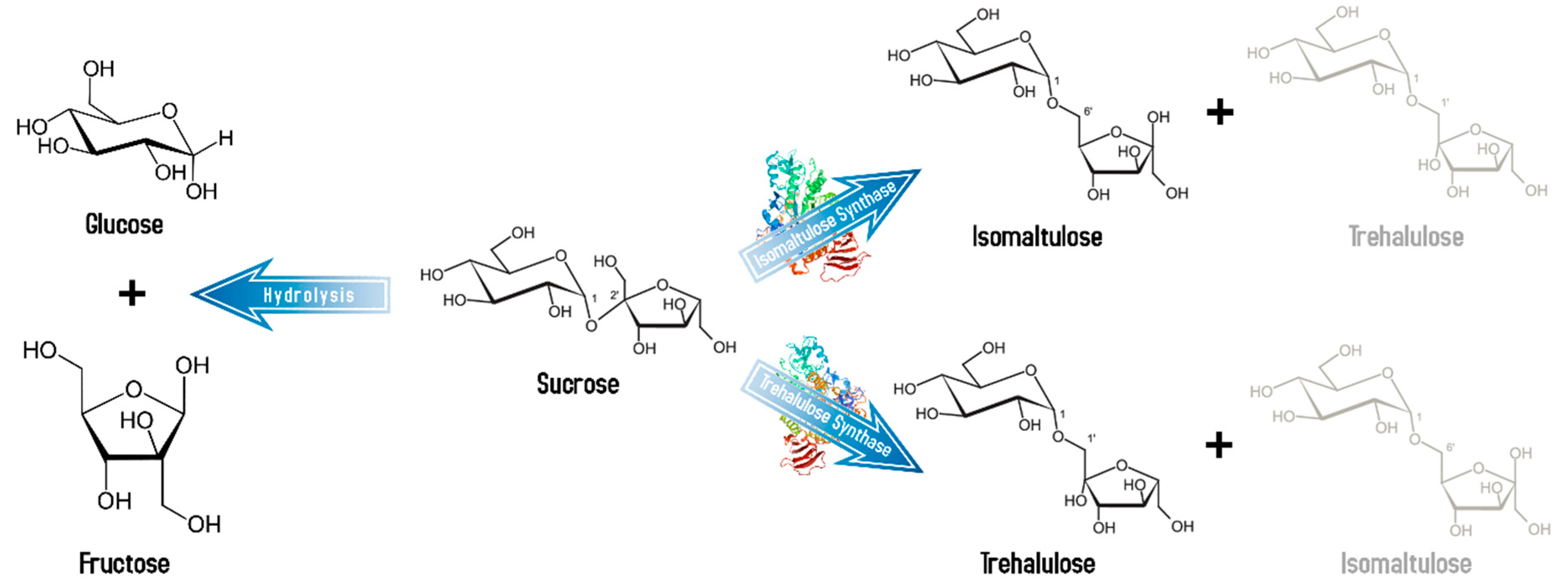
3.2.1. Isomaltulose (BCSDB ID: 111249; ChEBI 18394)
3.2.2. Trehalulose (BCSDB ID: 111199; ChEBI: 79284)
3.2.3. Modified Starches and High-Molecular Weight Carbohydrates
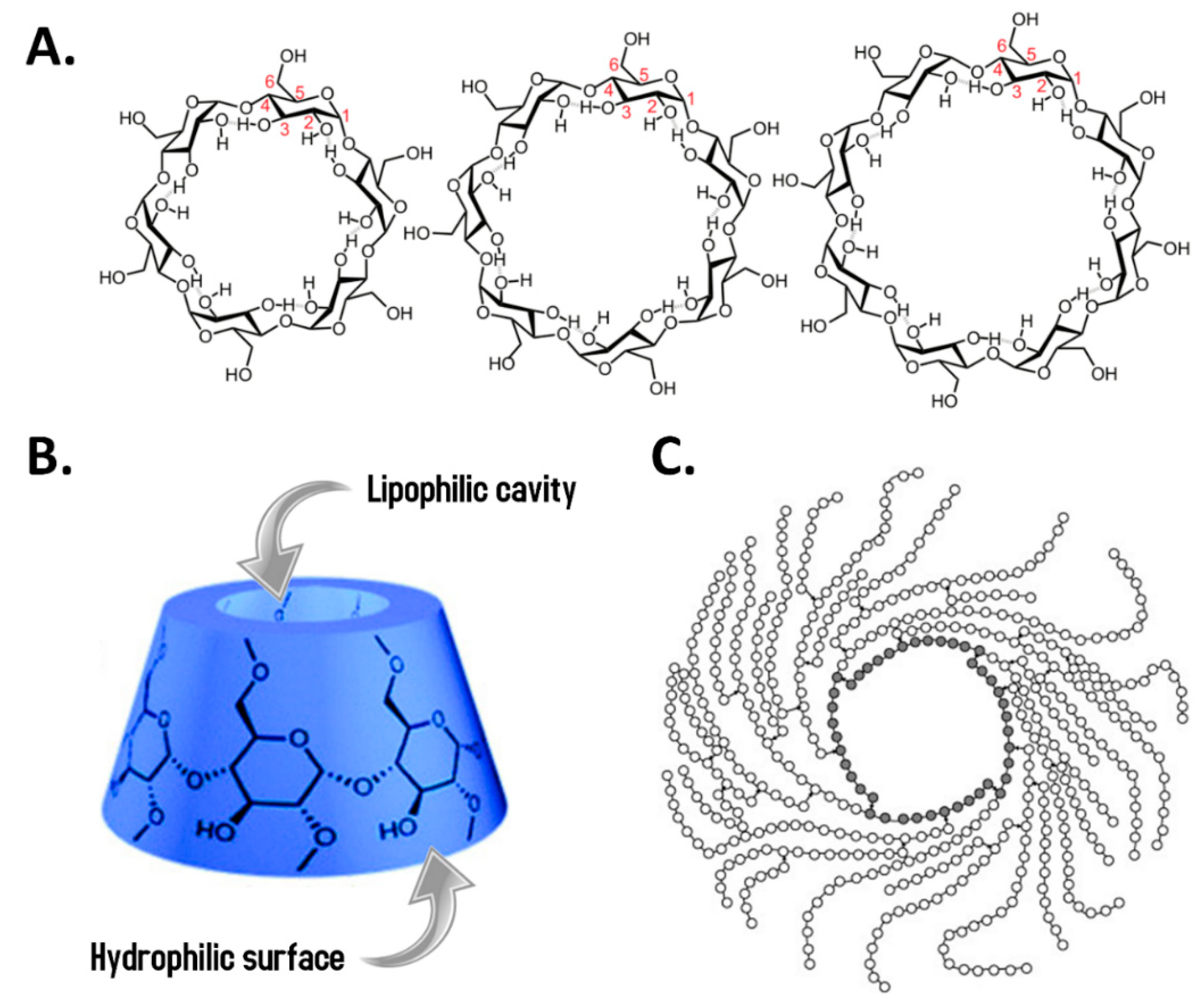
3.2.4. Cyclodextrins (ChEBI: 495055) and Derivates
3.3. Repair
- Optimal dose of protein for athletes to enhance MPS are dependent upon age, energy intake (higher amount is needed under energy restriction), and recent resistance exercise stimuli. Post-exercise recommendations are 0.5 g of a high-quality protein per kilogram of body mass, or an absolute dose of 40 g. Protein per meal should be between 0.25 and 0.40 g of protein per kg of body mass, or absolute values of 20 g.
- Given the observed benefits of pre- and post-exercise protein ingestion, athletes’ tolerance should be assessed to determine the optimal time period during which to ingest protein. Notwithstanding, in spite of the anabolic effect of exercise is long-lasting (at least 24 h), athletes can take advantage of the higher muscle sensitivity to nutrient uptake after exercise due to the likely diminishment over time.
3.4. Rest
- Ashwagandha supplementation (>150 mg aqueous root extract quaque hora somni) seems to be an effective nutritional strategy to improve sleep quality in healthy male and female subjects [156]; consequently, it should be also considered before sleep.
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Leveille, C.F.; Mikhaeil, J.S.; Turner, K.D.; Silvera, S.; Wilkinson, J.; Fajardo, V.A. Mitochondrial cristae density: A dynamic entity that is critical for energy production and metabolic power in skeletal muscle. J. Physiol. 2017, 595, 2779–2780. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.; Gejl, K.D.; Hey-Mogensen, M.; Holmberg, H.-C.; Suetta, C.; Krustrup, P.; Elemans, C.P.H.; Ørtenblad, N. Plasticity in mitochondrial cristae density allows metabolic capacity modulation in human skeletal muscle. J. Physiol. 2017, 595, 2839–2847. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.C.; Ellefsen, S.; Baar, K. Adaptations to Endurance and Strength Training. Cold Spring Harb. Perspect. Med. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Lambert, M.I. Training, Adaptations. In Encyclopedia of Exercise Medicine in Health and Disease; Mooren, F.C., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 854–856. [Google Scholar]
- Sundberg, C.W.; Fitts, R.H. Bioenergetic basis of skeletal muscle fatigue. Curr. Opin. Physiol. 2019, 10, 118–127. [Google Scholar] [CrossRef]
- Hunter, S.K. Performance Fatigability: Mechanisms and Task Specificity. Cold Spring Harb. Perspect. Med. 2018, 8. [Google Scholar] [CrossRef]
- Sundberg, C.W.; Hunter, S.K.; Trappe, S.W.; Smith, C.S.; Fitts, R.H. Effects of elevated H+ and Pi on the contractile mechanics of skeletal muscle fibres from young and old men: Implications for muscle fatigue in humans. J. Physiol. 2018, 596, 3993–4015. [Google Scholar] [CrossRef]
- Sterling, P. Allostasis: A model of predictive regulation. Physiol. Behav. 2012, 106, 5–15. [Google Scholar] [CrossRef]
- Burke, L. Nutricion en el Deporte/Nutrition in Sport: Un Enfoque Práctico/a Practical Approach; Editorial Médica Panamericana S.A.: Madrid, Spain, 2010. [Google Scholar]
- Zoorob, R.; Parrish, M.-E.E.; O’Hara, H.; Kalliny, M. Sports Nutrition Needs. Prim. Care 2013, 40, 475–486. [Google Scholar] [CrossRef]
- Ivy, J.L.; Ferguson-Stegall, L.M. Nutrient Timing. Am. J. Lifestyle Med. 2013, 8, 246–259. [Google Scholar] [CrossRef]
- Edes, A.N.; Crews, D.E. Allostatic load and biological anthropology. Am. J. Phys. Anthropol. 2017, 162, 44–70. [Google Scholar] [CrossRef]
- Egan, B.; Zierath, J.R. Exercise Metabolism and the Molecular Regulation of Skeletal Muscle Adaptation. Cell Metab. 2013, 17, 162–184. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.M.; Dasari, S.; Konopka, A.R.; Johnson, M.L.; Manjunatha, S.; Esponda, R.R.; Carter, R.E.; Lanza, I.R.; Nair, K.S. Enhanced Protein Translation Underlies Improved Metabolic and Physical Adaptations to Different Exercise Training Modes in Young and Old Humans. Cell Metab. 2017, 25, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Karagounis, L.G.; Hawley, J.A. Genes, Exercise, and Glucose and Insulin Metabolism. In Genetic and Molecular Aspects of Sport Performance; John Wiley & Sons Ltd.: Chichester, UK, 2010; pp. 240–248. [Google Scholar] [CrossRef]
- Hood, D.A. Invited Review: Contractile activity-induced mitochondrial biogenesis in skeletal muscle. J. Appl. Physiol. 2001, 90, 1137–1157. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A.E. Periodized Nutrition for Athletes. Sports Med. 2017, 47, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.T.; Erdman, K.A.; Burke, L.M. Position of the Academy of Nutrition and Dietetics, Dietitians of Canada, and the American College of Sports Medicine: Nutrition and Athletic Performance. J. Acad. Nutr. Diet. 2016, 116, 501–528. [Google Scholar] [CrossRef]
- Kreider, R.B. Essentials of Exercise & Sport Nutrition: Science to Practice; Lulu Press, Inc.: Morrisville, CA, USA, 2019. [Google Scholar]
- American College of Sports Medicine; Sawka, M.N.; Burke, L.M.; Eichner, E.R.; Maughan, R.J.; Montain, S.J.; Stachenfeld, N.S. Exercise and Fluid Replacement. Med. Sci. Sports Exerc. 2007, 39, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Kreider, R.B. Physiological Considerations of Ultraendurance Performance. Int. J. Sport Nutr. 1991, 1, 3–27. [Google Scholar] [CrossRef]
- McCubbin, A.J.; Allanson, B.A.; Caldwell Odgers, J.N.; Cort, M.M.; Costa, R.J.S.; Cox, G.R.; Crawshay, S.T.; Desbrow, B.; Freney, E.G.; Gaskell, S.K.; et al. Sports Dietitians Australia Position Statement: Nutrition for Exercise in Hot Environments. Int. J. Sport Nutr. Exerc. Metab. 2020, 30, 83–98. [Google Scholar] [CrossRef]
- Roberts, W.O.; O'Connor, F.G.; Kenney, W.L.; Cooper, L.; Cheuvront, S.N.; Casa, D.J.; Armstrong, L.E.; Anderson, S.A.; McDermott, B.P. National Athletic Trainers' Association Position Statement: Fluid Replacement for the Physically Active. J. Athl. Train. 2017, 52, 877–895. [Google Scholar] [CrossRef]
- Merson, S.J.; Maughan, R.J.; Shirreffs, S.M. Rehydration with drinks differing in sodium concentration and recovery from moderate exercise-induced hypohydration in man. Eur. J. Appl. Physiol. 2008, 103, 585–594. [Google Scholar] [CrossRef]
- Kavouras, S.A.; Armstrong, L.E.; Maresh, C.M.; Casa, D.J.; Herrera-Soto, J.A.; Scheett, T.P.; Stoppani, J.; Mack, G.W.; Kraemer, W. Rehydration with glycerol: endocrine, cardiovascular, and thermoregulatory responses during exercise in the heat. J. Appl. Physiol. 2005, 100, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Van Rosendal, S.P.; Coombes, J.S. Glycerol Use in Hyperhydration and Rehydration: Scientific Update. In Acute Topics in Sport Nutrition; Karger AG: Basel, Switzerland, 2012; pp. 104–112. [Google Scholar] [CrossRef]
- Perez-Idarraga, A.; Aragon-Vargas, L.F. Postexercise rehydration: Potassium-rich drinks versus water and a sports drink. Appl. Physiol. Nutr. Metab. = Physiol. Appl. Nutr. Metab. 2014, 39, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Aragón-Vargas, L.F.; Pérez Idárraga, A. Rehidratación posejercicio: La forma de distribuir la ingesta de un volumen constante de líquido no altera su conservación. Pensar En Mov. Rev. Cienc. Ejerc. Salud 2011, 9, 12–21. [Google Scholar] [CrossRef]
- Aragón-Vargas, L.F.; Pérez-Idárraga, A. Post-Exercise Rehydration: No Change in Diuresis from Water Ingested at Different Temperatures. Med. Sport. 2010, 14, 77–82. [Google Scholar] [CrossRef]
- Sandick, B.L.; Engell, D.B.; Maller, O. Perception of drinking water temperature and effects for humans after exercise. Physiol. Behav. 1984, 32, 851–855. [Google Scholar] [CrossRef]
- Karp, J.R.; Johnston, J.D.; Tecklenburg, S.; Mickleborough, T.D.; Fly, A.D.; Stager, J.M. Chocolate milk as a post-exercise recovery aid. Int. J. Sport Nutr. Exerc. Metab. 2006, 16, 78–91. [Google Scholar] [CrossRef]
- Roy, B.D. Milk: The new sports drink? A Review. J. Int. Soc. Sports Nutr. 2008, 5, 15. [Google Scholar] [CrossRef]
- Shirreffs, S.M.; Watson, P.; Maughan, R.J. Milk as an effective post-exercise rehydration drink. Br. J. Nutr. 2007, 98, 173–180. [Google Scholar] [CrossRef]
- Jentjens, R.; Jeukendrup, A. Determinants of post-exercise glycogen synthesis during short-term recovery. Sports Med. 2003, 33, 117–144. [Google Scholar] [CrossRef]
- Burke, L.M.; Loucks, A.B.; Broad, N. Energy and carbohydrate for training and recovery. J. Sports Sci. 2006, 24, 675–685. [Google Scholar] [CrossRef]
- Mujika, I.; Burke, L.M. Nutrition in Team Sports. Ann. Nutr. Metab. 2010, 57, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Kerksick, C.M.; Wilborn, C.D.; Roberts, M.D.; Smith-Ryan, A.; Kleiner, S.M.; Jäger, R.; Collins, R.; Cooke, M.; Davis, J.N.; Galvan, E.; et al. ISSN exercise & sports nutrition review update: Research & recommendations. J. Int. Soc. Sports Nutr. 2018, 15. [Google Scholar] [CrossRef]
- Burke, L.M.; Hawley, J.A.; Wong, S.H.; Jeukendrup, A.E. Carbohydrates for training and competition. J. Sports Sci. 2011, 29, S17–S27. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.; Fuchs, C.; Betts, J.; van Loon, L. Glucose Plus Fructose Ingestion for Post-Exercise Recovery—Greater than the Sum of Its Parts? Nutrients 2017, 9, 344. [Google Scholar] [CrossRef] [PubMed]
- Murray, B.; Rosenbloom, C. Fundamentals of glycogen metabolism for coaches and athletes. Nutr. Rev. 2018, 76, 243–259. [Google Scholar] [CrossRef]
- Varðardóttir, B.; Guðmundsdóttir, S.L.; Ólafsdóttir, A.S. Þegar orkuna skortir–áhrif hlutfallslegs orkuskorts í íþróttum (RED-s) á heilsu og árangur. Læknablaðið 2020, 106, 406–413. [Google Scholar] [CrossRef]
- Mountjoy, M.; Sundgot-Borgen, J.; Burke, L.; Ackerman, K.E.; Blauwet, C.; Constantini, N.; Lebrun, C.; Lundy, B.; Melin, A.; Meyer, N.; et al. International Olympic Committee (IOC) Consensus Statement on Relative Energy Deficiency in Sport (RED-S): 2018 Update. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 316–331. [Google Scholar] [CrossRef]
- Toukach, P.V.; Egorova, K.S. Carbohydrate structure database merged from bacterial, archaeal, plant and fungal parts. Nucleic Acids Res. 2016, 44, D1229–D1236. [Google Scholar] [CrossRef]
- Degtyarenko, K.; de Matos, P.; Ennis, M.; Hastings, J.; Zbinden, M.; McNaught, A.; Alcantara, R.; Darsow, M.; Guedj, M.; Ashburner, M. ChEBI: A database and ontology for chemical entities of biological interest. Nucleic Acids Res. 2007, 36, D344–D350. [Google Scholar] [CrossRef]
- Goulter, K.C.; Hashimi, S.M.; Birch, R.G. Microbial sucrose isomerases: Producing organisms, genes and enzymes. Enzym. Microb. Technol. 2012, 50, 57–64. [Google Scholar] [CrossRef]
- Zhang, G.; Ao, Z.; Hamaker, B.R. Controlling the delivery of glucose in foods. In Designing Functional Foods; Woodhead Publishing: Cambridge, UK, 2009; pp. 547–571. [Google Scholar]
- König, D.; Theis, S.; Kozianowski, G.; Berg, A. Postprandial substrate use in overweight subjects with the metabolic syndrome after isomaltulose (Palatinose™) ingestion. Nutrition 2012, 28, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Kemp, S.; Schweppes, C.; Lindley, M. Developments in sweeteners. In Modifying Flavour in Food; Woodhead Publishing: Cambridge, UK, 2007; pp. 185–201. [Google Scholar]
- Bracken, R.M.; Page, R.; Gray, B.; Kilduff, L.P.; West, D.J.; Stephens, J.W.; Bain, S.C. Isomaltulose Improves Glycemia and Maintains Run Performance in Type 1 Diabetes. Med. Sci. Sports Exerc. 2012, 44, 800–808. [Google Scholar] [CrossRef] [PubMed]
- König, D.; Zdzieblik, D.; Holz, A.; Theis, S.; Gollhofer, A. Substrate Utilization and Cycling Performance Following Palatinose™ Ingestion: A Randomized, Double-Blind, Controlled Trial. Nutrients 2016, 8, 390. [Google Scholar] [CrossRef]
- Hattori, S.; Noguchi, A.; Sasagawa, K.; Ogata, H.; Kobayashi, M.; Omi, N. Influence of Isomaltulose Ingestion on Fat Oxidation During Inclemental Exercise in Endurance Athletes. Am. J. Sports Sci. 2019, 7. [Google Scholar] [CrossRef]
- Stevenson, E.J.; Watson, A.; Theis, S.; Holz, A.; Harper, L.D.; Russell, M. A comparison of isomaltulose versus maltodextrin ingestion during soccer-specific exercise. Eur. J. Appl. Physiol. 2017, 117, 2321–2333. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, W.J.; Hooper, D.R.; Szivak, T.K.; Kupchak, B.R.; Dunn-Lewis, C.; Comstock, B.A.; Flanagan, S.D.; Looney, D.P.; Sterczala, A.J.; DuPont, W.H.; et al. The Addition of Beta-hydroxy-beta-methylbutyrate and Isomaltulose to Whey Protein Improves Recovery from Highly Demanding Resistance Exercise. J. Am. Coll. Nutr. 2015, 34, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Amano, T.; Sugiyama, Y.; Okumura, J.; Fujii, N.; Kenny, G.P.; Nishiyasu, T.; Inoue, Y.; Kondo, N.; Sasagawa, K.; Enoki, Y.; et al. Effects of isomaltulose ingestion on postexercise hydration state and heat loss responses in young men. Exp. Physiol. 2019, 104, 1494–1504. [Google Scholar] [CrossRef]
- Maresch, C.C.; Petry, S.F.; Theis, S.; Bosy-Westphal, A.; Linn, T. Low Glycemic Index Prototype Isomaltulose—Update of Clinical Trials. Nutrients 2017, 9, 381. [Google Scholar] [CrossRef]
- Ferrugem, L.; Martini, G.; de Souza, C. Influence of the Glycemic Index of Pre-exercise Meals in Sports Performance: A Systematic Review. Int. J. Med. Rev. 2018, 5, 151–158. [Google Scholar] [CrossRef]
- Surana, R.; Pyne, A.; Suryanarayanan, R. Effect of Preparation Method on Physical Properties of Amorphous Trehalose. Pharm. Res. 2004, 21, 1167–1176. [Google Scholar] [CrossRef]
- Côté, G.L. Flavorings and other value-added products from sucrose * *Mention of trade names or commercial products is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture. In Novel Enzyme Technology for Food Applications; Woodhead Publishing: Cambridge, UK, 2007; pp. 243–269. [Google Scholar]
- Arai, S.; Morinaga, Y.; Yoshikawa, T.; Ichiishi, E.; Kiso, Y.; Yamazaki, M.; Morotomi, M.; Shimizu, M.; Kuwata, T.; Kaminogawa, S. Recent Trends in Functional Food Science and the Industry in Japan. Biosci. Biotechnol. Biochem. 2014, 66, 2017–2029. [Google Scholar] [CrossRef] [PubMed]
- Lina, B.A.R.; Jonker, D.; Kozianowski, G. Isomaltulose (Palatinose®): A review of biological and toxicological studies. Food Chem. Toxicol. 2002, 40, 1375–1381. [Google Scholar] [CrossRef]
- Ngo, C. In Vitro Starch Digestion for Analysis of Healthy Carbohydrates. Master’s Thesis, Luleå University of Technology, Luleå, Sweden, 2018. [Google Scholar]
- Zia ud, D.; Xiong, H.; Fei, P. Physical and chemical modification of starches: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 2691–2705. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, A.; Schmidt, T.; Kaess, H. The role of osmolality in the absorption of a nutrient solution. Aliment. Pharmacol. Ther. 1998, 12, 281–286. [Google Scholar] [CrossRef]
- Piehl Aulin, K.; Söderlund, K.; Hultman, E. Muscle glycogen resynthesis rate in humans after supplementation of drinks containing carbohydrates with low and high molecular masses. Eur. J. Appl. Physiol. 2000, 81, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Stephens, F.B.; Roig, M.; Armstrong, G.; Greenhaff, P.L. Post-exercise ingestion of a unique, high molecular weight glucose polymer solution improves performance during a subsequent bout of cycling exercise. J. Sports Sci. 2008, 26, 149–154. [Google Scholar] [CrossRef]
- Oliver, J.M.; Almada, A.L.; Van Eck, L.E.; Shah, M.; Mitchell, J.B.; Jones, M.T.; Jagim, A.R.; Rowlands, D.S. Ingestion of High Molecular Weight Carbohydrate Enhances Subsequent Repeated Maximal Power: A Randomized Controlled Trial. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- McGlory, C.; Morton, J.P. The Effects of Postexercise Consumption of High-Molecular-Weight versus Low-Molecular-Weight Carbohydrate Solutions on Subsequent High-Intensity Interval-Running Capacity. Int. J. Sport Nutr. Exerc. Metab. 2010, 20, 361–369. [Google Scholar] [CrossRef]
- Anzalone, A.J.; Almada, A.L.; Van Eck, L.E.; Jones, M.T.; Jagim, A.R.; Mitchell, J.B.; Shah, M.; Oliver, J.M. Effect of post-exercise ingestion of different molecular weight carbohydrate solutions. Part II: The incretin response. J. Int. Soc. Sports Nutr. 2015, 12. [Google Scholar] [CrossRef]
- Mock, M.G.; Hirsch, K.R.; Blue, M.N.M.; Trexler, E.T.; Roelofs, E.J.; Smith-Ryan, A.E. Post-Exercise Ingestion of Low or High Molecular Weight Glucose Polymer Solution Does Not Improve Cycle Performance in Female Athletes. J. Strength Cond. Res. 2018. [Google Scholar] [CrossRef]
- Leemhuis, H.; Kelly, R.M.; Dijkhuizen, L. Engineering of cyclodextrin glucanotransferases and the impact for biotechnological applications. Appl. Microbiol. Biotechnol. 2009, 85, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Stick, R.V.; Williams, S.J. Disaccharides, Oligosaccharides and Polysaccharides. In Carbohydrates: The Essential Molecules of Life; Elsevier Science: Amsterdam, The Netherlands, 2009; pp. 321–341. [Google Scholar] [CrossRef]
- French, D.; Pulley, A.O.; Effenberger, J.A.; Rougvie, M.A.; Abdullah, M. Studies on the Schardinger dextrins. XII. The molecular size and structure of the delta-, epsilon-, zeta-, and eta-dextrins. Arch. Biochem. Biophys. 1965, 111, 153–160. [Google Scholar] [CrossRef]
- Avakyan, V.G.; Nazarov, V.B.; Alfimov, M.V.; Bagatur"yants, A.A.; Voronezheva, N.I. The role of intra- and intermolecular hydrogen bonds in the formation of β-cyclodextrin head-to-head and head-to-tail dimers. The results of ab initio and semiempirical quantum-chemical calculations. Russ. Chem. Bull. 2001, 50, 206–216. [Google Scholar] [CrossRef]
- Narayanaswamy, R.; Kanagesan, S.; Pandurangan, A.; Padmanabhan, P. Basics to different imaging techniques, different nanobiomaterials for image enhancement. In Nanobiomaterials in Medical Imaging; William Andrew: Norwich, NY, USA, 2016; pp. 101–129. [Google Scholar]
- Semalty, A. Cyclodextrin and phospholipid complexation in solubility and dissolution enhancement: A critical and meta-analysis. Expert Opin. Drug Deliv. 2014, 11, 1255–1272. [Google Scholar] [CrossRef]
- Dura, A.; Yokoyama, W.; Rosell, C.M. Glycemic Response to Corn Starch Modified with Cyclodextrin Glycosyltransferase and its Relationship to Physical Properties. Plant Foods Hum. Nutr. 2016, 71, 252–258. [Google Scholar] [CrossRef]
- Choi, S.S.H.; Danielewska-Nikiel, B.; Ohdan, K.; Kojima, I.; Takata, H.; Kuriki, T. Safety evaluation of highly-branched cyclic dextrin and a 1,4-α-glucan branching enzyme from Bacillus stearothermophilus. Regul. Toxicol. Pharmacol. 2009, 55, 281–290. [Google Scholar] [CrossRef]
- Takii, H.; Takii, Y.; Kometani, T.; Nishimura, T.; Nakae, T.; Kuriki, T.; Fushiki, T. Fluids Containing a Highly Branched Cyclic Dextrin Influence the Gastric Emptying Rate. Int. J. Sports Med. 2005, 26, 314–319. [Google Scholar] [CrossRef]
- Fenyvesi, É.; Vikmon, M.; Szente, L. Cyclodextrins in Food Technology and Human Nutrition: Benefits and Limitations. Crit. Rev. Food Sci. Nutr. 2015, 56, 1981–2004. [Google Scholar] [CrossRef]
- Takata, H.; Kojima, I.; Taji, N.; Suzuki, Y.; Yamamoto, M. Industrial production of branching enzyme, and its application to production of highly branched cyclic dextrin (Cluster DextrinTM). Seibutsu Kogaku Kaishi 2006, 84, 61–66. [Google Scholar]
- Kometani, T. Production of novel bioactive compounds by enzymes, and their application to food. Pure Appl. Chem. 2010, 82, 269–287. [Google Scholar] [CrossRef]
- Takii, H.; Kometani, T.; Nishimura, T.; Kuriki, T.; Fushiki, T. A Sports Drink Based on Highly Branched Cyclic Dextrin Generates Few Gastrointestinal Disorders in Untrained Men during Bicycle Exercise. Food Sci. Technol. Res. 2004, 10, 428–431. [Google Scholar] [CrossRef]
- Shiraki, T.; Kometani, T.; Yoshitani, K.; Takata, H.; Nomura, T. Evaluation of Exercise Performance with the Intake of Highly Branched Cyclic Dextrin in Athletes. Food Sci. Technol. Res. 2015, 21, 499–502. [Google Scholar] [CrossRef]
- Furuyashiki, T.; Tanimoto, H.; Yokoyama, Y.; Kitaura, Y.; Kuriki, T.; Shimomura, Y. Effects of ingesting highly branched cyclic dextrin during endurance exercise on rating of perceived exertion and blood components associated with energy metabolism. Biosci. Biotechnol. Biochem. 2014, 78, 2117–2119. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Shiraishi, K.; Yoshitani, K.; Sugama, K.; Kometani, T. Effect of a sports drink based on highly-branched cyclic dextrin on cytokine responses to exhaustive endurance exercise. J. Sports Med. Phys. Fit. 2014, 54, 622–630. [Google Scholar]
- Bruns, C.J. Exploring and Exploiting the Symmetry-Breaking Effect of Cyclodextrins in Mechanomolecules. Symmetry 2019, 11, 1249. [Google Scholar] [CrossRef]
- Rousseau, J.; Menuel, S.; Rousseau, C.; Hapiot, F.; Monflier, E. Cyclodextrins as Porous Material for Catalysis. In Organic Nanoreactors; Academic Press: London, UK, 2016; pp. 15–42. [Google Scholar]
- Baykal, A.; Bozkurt, A.; Jeremy, R.; Asiri, S.M.M.; Lima-Tenório, M.K.; Kaewsaneha, C.; Elaissari, A. Multistimuli-responsive magnetic assemblies. In Stimuli Responsive Polymeric Nanocarriers for Drug Delivery Applications; Woodhead Publishing: Cambridge, UK, 2019; pp. 155–193. [Google Scholar]
- Kadota, K.; Senda, A.; Ito, T.; Tozuka, Y. Feasibility of highly branched cyclic dextrin as an excipient matrix in dry powder inhalers. Eur. J. Pharm. Sci. 2015, 79, 79–86. [Google Scholar] [CrossRef]
- Fujii, K.; Takata, H.; Yanase, M.; Terada, Y.; Ohdan, K.; Takaha, T.; Okada, S.; Kuriki, T. Bioengineering and Application of Novel Glucose Polymers. Biocatal. Biotransform. 2010, 21, 167–172. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2013, 505, 559–563. [Google Scholar] [CrossRef]
- Huang, W.-C.; Pan, C.-H.; Wei, C.-C.; Huang, H.-Y. Lactobacillus plantarum PS128 Improves Physiological Adaptation and Performance in Triathletes through Gut Microbiota Modulation. Nutrients 2020, 12, 2315. [Google Scholar] [CrossRef]
- Jäger, R.; Mohr, A.E.; Carpenter, K.C.; Kerksick, C.M.; Purpura, M.; Moussa, A.; Townsend, J.R.; Lamprecht, M.; West, N.P.; Black, K.; et al. International Society of Sports Nutrition Position Stand: Probiotics. J. Int. Soc. Sports Nutr. 2019, 16. [Google Scholar] [CrossRef]
- Mohr, A.E.; Jäger, R.; Carpenter, K.C.; Kerksick, C.M.; Purpura, M.; Townsend, J.R.; West, N.P.; Black, K.; Gleeson, M.; Pyne, D.B.; et al. The athletic gut microbiota. J. Int. Soc. Sports Nutr. 2020, 17. [Google Scholar] [CrossRef] [PubMed]
- Jager, R.; Kerksick, C.M.; Campbell, B.I.; Cribb, P.J.; Wells, S.D.; Skwiat, T.M.; Purpura, M.; Ziegenfuss, T.N.; Ferrando, A.A.; Arent, S.M.; et al. International Society of Sports Nutrition Position Stand: Protein and exercise. J. Int. Soc. Sports Nutr. 2017, 14, 20. [Google Scholar] [CrossRef] [PubMed]
- Kerksick, C.M.; Arent, S.; Schoenfeld, B.J.; Stout, J.R.; Campbell, B.; Wilborn, C.D.; Taylor, L.; Kalman, D.; Smith-Ryan, A.E.; Kreider, R.B.; et al. International society of sports nutrition position stand: Nutrient timing. J. Int. Soc. Sports Nutr. 2017, 14. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.J.; Luden, N.D.; DeWitt, C.R.; Gross, M.C.; Dillon Rios, A. Protein Supplementation During or Following a Marathon Run Influences Post-Exercise Recovery. Nutrients 2018, 10, 333. [Google Scholar] [CrossRef] [PubMed]
- Churchward-Venne, T.A.; Pinckaers, P.J.M.; Smeets, J.S.J.; Betz, M.W.; Senden, J.M.; Goessens, J.P.B.; Gijsen, A.P.; Rollo, I.; Verdijk, L.B.; van Loon, L.J.C. Dose-response effects of dietary protein on muscle protein synthesis during recovery from endurance exercise in young men: A double-blind randomized trial. Am. J. Clin. Nutr. 2020, 112, 303–317. [Google Scholar] [CrossRef]
- Kreider, R.B.; Kalman, D.S.; Antonio, J.; Ziegenfuss, T.N.; Wildman, R.; Collins, R.; Candow, D.G.; Kleiner, S.M.; Almada, A.L.; Lopez, H.L. International Society of Sports Nutrition position stand: Safety and efficacy of creatine supplementation in exercise, sport, and medicine. J. Int. Soc. Sports Nutr. 2017, 14. [Google Scholar] [CrossRef]
- Bonilla, D.A.; Moreno, Y. Molecular and metabolic insights of creatine supplementation on resistance training. Rev. Colomb. Química 2015, 44, 11–18. [Google Scholar] [CrossRef]
- Roberts, P.A.; Fox, J.; Peirce, N.; Jones, S.W.; Casey, A.; Greenhaff, P.L. Creatine ingestion augments dietary carbohydrate mediated muscle glycogen supercompensation during the initial 24 h of recovery following prolonged exhaustive exercise in humans. Amino Acids 2016, 48, 1831–1842. [Google Scholar] [CrossRef]
- Myburgh, K.H. Polyphenol supplementation: Benefits for exercise performance or oxidative stress? Sports Med. 2014, 44, S57–S70. [Google Scholar] [CrossRef]
- Levers, K.; Dalton, R.; Galvan, E.; O'Connor, A.; Goodenough, C.; Simbo, S.; Mertens-Talcott, S.U.; Rasmussen, C.; Greenwood, M.; Riechman, S.; et al. Effects of powdered Montmorency tart cherry supplementation on acute endurance exercise performance in aerobically trained individuals. J. Int. Soc. Sports Nutr. 2015, 13, 22. [Google Scholar] [CrossRef]
- Brown, M.A.; Stevenson, E.J.; Howatson, G. Montmorency tart cherry (Prunus cerasus L.) supplementation accelerates recovery from exercise-induced muscle damage in females. Eur. J. Sport Sci. 2018, 19, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Somerville, V.; Bringans, C.; Braakhuis, A. Polyphenols and Performance: A Systematic Review and Meta-Analysis. Sports Med. 2017, 47, 1589–1599. [Google Scholar] [CrossRef] [PubMed]
- Bowtell, J.; Kelly, V. Fruit-Derived Polyphenol Supplementation for Athlete Recovery and Performance. Sports Med. 2019, 49, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Doma, K.; Gahreman, D.; Connor, J. Fruit supplementation reduces indices of exercise-induced muscle damage: A systematic review and meta-analysis. Eur. J. Sport Sci. 2020, 1–18. [Google Scholar] [CrossRef]
- Delecroix, B.; Abaidia, A.E.; Leduc, C.; Dawson, B.; Dupont, G. Curcumin and Piperine Supplementation and Recovery Following Exercise Induced Muscle Damage: A Randomized Controlled Trial. J. Sports Sci. Med. 2017, 16, 147–153. [Google Scholar]
- Bonilla, D.A.; Paipilla, A.F.; Marin, E.; Vargas-Molina, S.; Petro, J.L.; Perez-Idarraga, A. Dietary Nitrate from Beetroot Juice for Hypertension: A Systematic Review. Biomolecules 2018, 8, 134. [Google Scholar] [CrossRef]
- Siervo, M.; Lara, J.; Ogbonmwan, I.; Mathers, J.C. Inorganic nitrate and beetroot juice supplementation reduces blood pressure in adults: A systematic review and meta-analysis. J. Nutr. 2013, 143, 818–826. [Google Scholar] [CrossRef]
- Cuenca, E.; Jodra, P.; Perez-Lopez, A.; Gonzalez-Rodriguez, L.G.; Fernandes da Silva, S.; Veiga-Herreros, P.; Dominguez, R. Effects of Beetroot Juice Supplementation on Performance and Fatigue in a 30-s All-Out Sprint Exercise: A Randomized, Double-Blind Cross-Over Study. Nutrients 2018, 10, 1222. [Google Scholar] [CrossRef]
- Mosher, S.L.; Sparks, S.A.; Williams, E.L.; Bentley, D.J.; Mc Naughton, L.R. Ingestion of a Nitric Oxide Enhancing Supplement Improves Resistance Exercise Performance. J. Strength Cond. Res. 2016, 30, 3520–3524. [Google Scholar] [CrossRef]
- Clifford, T.; Berntzen, B.; Davison, G.W.; West, D.J.; Howatson, G.; Stevenson, E.J. Effects of Beetroot Juice on Recovery of Muscle Function and Performance between Bouts of Repeated Sprint Exercise. Nutrients 2016, 8, 506. [Google Scholar] [CrossRef]
- Clifford, T.; Bell, O.; West, D.J.; Howatson, G.; Stevenson, E.J. The effects of beetroot juice supplementation on indices of muscle damage following eccentric exercise. Eur. J. Appl. Physiol. 2015, 116, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, C.F.; Kwong, D.A.; Minow, Z.A.; Davis, B.A.; Lozada, C.F.; Casazza, G.A. Betalain-rich concentrate supplementation improves exercise performance and recovery in competitive triathletes. Appl. Physiol. Nutr. Metab. 2017, 42, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.B. A Randomized Double-Blind Trial of Ginger Root for Reducing Muscle Soreness and Improving Physical Performance Recovery among Experienced Recreational Distance Runners. J. Diet. Suppl. 2018, 17, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Balmaseda, D.; Diez-Vega, I.; Larrosa, M.; San Juan, A.F.; Issaly, N.; Moreno-Pérez, D.; Burgos, S.; Sillero-Quintana, M.; Gonzalez, C.; Bas, A.; et al. Effect of a Blend of Zingiber officinale Roscoe and Bixa orellana L. Herbal Supplement on the Recovery of Delayed-Onset Muscle Soreness Induced by Unaccustomed Eccentric Resistance Training: A Randomized, Triple-Blind, Placebo-Controlled Trial. Front. Physiol. 2020, 11. [Google Scholar] [CrossRef]
- Pu, W.-l.; Zhang, M.-y.; Bai, R.-y.; Sun, L.-k.; Li, W.-h.; Yu, Y.-l.; Zhang, Y.; Song, L.; Wang, Z.-x.; Peng, Y.-f.; et al. Anti-inflammatory effects of Rhodiola rosea L.: A review. Biomed. Pharmacother. 2020, 121. [Google Scholar] [CrossRef]
- Tan, L.; Song, X.; Ren, Y.; Wang, M.; Guo, C.; Guo, D.; Gu, Y.; Li, Y.; Cao, Z.; Deng, Y. Anti-inflammatory effects of cordycepin: A review. Phytother. Res. 2020. [Google Scholar] [CrossRef]
- Tsai, P.-H.; Lin, F.-C.; Huang, C.-C.; Hou, C.-W.; Cheng, I.-S. Effects of Rhodiola rosea-Cordyceps sinensis Supplementation on Glycogen Synthesis in Exercised Human Skeletal Muscle. Sports Exerc. Res. 2019, 21, 375–386. [Google Scholar]
- Choudhary, B.; Shetty, A.; Langade, D.G. Efficacy of Ashwagandha (Withania somnifera [L.] Dunal) in improving cardiorespiratory endurance in healthy athletic adults. Ayu (Int. Q. J. Res. Ayurveda) 2015, 36, 63. [Google Scholar] [CrossRef]
- Wankhede, S.; Langade, D.; Joshi, K.; Sinha, S.R.; Bhattacharyya, S. Examining the effect of Withania somnifera supplementation on muscle strength and recovery: A randomized controlled trial. J. Int. Soc. Sports Nutr. 2015, 12. [Google Scholar] [CrossRef]
- Ziegenfuss, T.; Kedia, A.; Sandrock, J.; Raub, B.; Kerksick, C.; Lopez, H. Effects of an Aqueous Extract of Withania somnifera on Strength Training Adaptations and Recovery: The STAR Trial. Nutrients 2018, 10, 1807. [Google Scholar] [CrossRef]
- Langade, D.; Kanchi, S.; Salve, J.; Debnath, K.; Ambegaokar, D. Efficacy and Safety of Ashwagandha (Withania somnifera) Root Extract in Insomnia and Anxiety: A Double-blind, Randomized, Placebo-controlled Study. Cureus 2019. [Google Scholar] [CrossRef] [PubMed]
- Salve, J.; Pate, S.; Debnath, K.; Langade, D. Adaptogenic and Anxiolytic Effects of Ashwagandha Root Extract in Healthy Adults: A Double-blind, Randomized, Placebo-controlled Clinical Study. Cureus 2019. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Bhalla, M.; De Jager, P.; Gilca, M. An Overview on Ashwagandha: A Rasayana (Rejuvenator) of Ayurveda. Afr. J. Tradit. Complementary Altern. Med. 2011, 8. [Google Scholar] [CrossRef]
- Kraemer, W.J.; Ratamess, N.A.; Volek, J.S.; Häkkinen, K.; Rubin, M.R.; French, D.N.; Gómez, A.L.; McGuigan, M.R.; Scheett, T.P.; Newton, R.U.; et al. The effects of amino acid supplementation on hormonal responses to resistance training overreaching. Metab. Clin. Exp. 2006, 55, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Fouré, A.; Bendahan, D. Is Branched-Chain Amino Acids Supplementation an Efficient Nutritional Strategy to Alleviate Skeletal Muscle Damage? A Systematic Review. Nutrients 2017, 9, 1047. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, M.H.; Shab-Bidar, S.; Mollahosseini, M.; Djafarian, K. Branched-chain amino acid supplementation and exercise-induced muscle damage in exercise recovery: A meta-analysis of randomized clinical trials. Nutrition 2017, 42, 30–36. [Google Scholar] [CrossRef]
- Hormoznejad, R.; Zare Javid, A.; Mansoori, A. Effect of BCAA supplementation on central fatigue, energy metabolism substrate and muscle damage to the exercise: A systematic review with meta-analysis. Sport Sci. Health 2019, 15, 265–279. [Google Scholar] [CrossRef]
- Fedewa, M.V.; Spencer, S.O.; Williams, T.D.; Becker, Z.E.; Fuqua, C.A. Effect of branched-Chain Amino Acid Supplementation on Muscle Soreness following Exercise: A Meta-Analysis. Int. J. Vitam. Nutr. Res. 2019, 89, 348–356. [Google Scholar] [CrossRef]
- VanDusseldorp, T.A.; Escobar, K.A.; Johnson, K.E.; Stratton, M.T.; Moriarty, T.; Cole, N.; McCormick, J.J.; Kerksick, C.M.; Vaughan, R.A.; Dokladny, K.; et al. Effect of Branched-Chain Amino Acid Supplementation on Recovery Following Acute Eccentric Exercise. Nutrients 2018, 10, 1389. [Google Scholar] [CrossRef]
- Correia, A.L.M.; de Lima, F.D.; Bottaro, M.; Vieira, A.; da Fonseca, A.C.; Lima, R.M. Pre-exercise β-hydroxy-β-methylbutyrate free-acid supplementation improves work capacity recovery: A randomized, double-blinded, placebo-controlled study. Appl. Physiol. Nutr. Metab. 2018, 43, 691–696. [Google Scholar] [CrossRef]
- Silva, V.R.; Belozo, F.L.; Micheletti, T.O.; Conrado, M.; Stout, J.R.; Pimentel, G.D.; Gonzalez, A.M. β-hydroxy-β-methylbutyrate free acid supplementation may improve recovery and muscle adaptations after resistance training: A systematic review. Nutr. Res. 2017, 45, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, M.H.; Mohammadi, H.; Eshaghi, H.; Askari, G.; Miraghajani, M. The Effects of Beta-Hydroxy-Beta-Methylbutyrate Supplementation on Recovery Following Exercise-Induced Muscle Damage: A Systematic Review and Meta-Analysis. J. Am. Coll. Nutr. 2018, 37, 640–649. [Google Scholar] [CrossRef] [PubMed]
- Rawson, E.S.; Miles, M.P.; Larson-Meyer, D.E. Dietary Supplements for Health, Adaptation, and Recovery in Athletes. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Ammar, A.; Bailey, S.J.; Chtourou, H.; Trabelsi, K.; Turki, M.; Hökelmann, A.; Souissi, N. Effects of pomegranate supplementation on exercise performance and post-exercise recovery in healthy adults: A systematic review. Br. J. Nutr. 2018, 120, 1201–1216. [Google Scholar] [CrossRef] [PubMed]
- Alcantara, J.M.A.; Sanchez-Delgado, G.; Martinez-Tellez, B.; Labayen, I.; Ruiz, J.R. Impact of cow’s milk intake on exercise performance and recovery of muscle function: A systematic review. J. Int. Soc. Sports Nutr. 2019, 16. [Google Scholar] [CrossRef]
- Amiri, M.; Ghiasvand, R.; Kaviani, M.; Forbes, S.C.; Salehi-Abargouei, A. Chocolate milk for recovery from exercise: A systematic review and meta-analysis of controlled clinical trials. Eur. J. Clin. Nutr. 2018, 73, 835–849. [Google Scholar] [CrossRef]
- Vitale, K.C.; Owens, R.; Hopkins, S.R.; Malhotra, A. Sleep Hygiene for Optimizing Recovery in Athletes: Review and Recommendations. Int. J. Sports Med. 2019, 40, 535–543. [Google Scholar] [CrossRef]
- Fullagar, H.H.K.; Duffield, R.; Skorski, S.; Coutts, A.J.; Julian, R.; Meyer, T. Sleep and Recovery in Team Sport: Current Sleep-Related Issues Facing Professional Team-Sport Athletes. Int. J. Sports Physiol. Perform. 2015, 10, 950–957. [Google Scholar] [CrossRef]
- von Rosen, P.; Frohm, A.; Kottorp, A.; Fridén, C.; Heijne, A. Too little sleep and an unhealthy diet could increase the risk of sustaining a new injury in adolescent elite athletes. Scand. J. Med. Sci. Sports 2017, 27, 1364–1371. [Google Scholar] [CrossRef]
- Silva, M.R.G.; Paiva, T. Poor precompetitive sleep habits, nutrients’ deficiencies, inappropriate body composition and athletic performance in elite gymnasts. Eur. J. Sport Sci. 2015, 16, 726–735. [Google Scholar] [CrossRef]
- Gupta, L.; Morgan, K.; Gilchrist, S. Does Elite Sport Degrade Sleep Quality? A Systematic Review. Sports Med. 2016, 47, 1317–1333. [Google Scholar] [CrossRef] [PubMed]
- Bonnar, D.; Bartel, K.; Kakoschke, N.; Lang, C. Sleep Interventions Designed to Improve Athletic Performance and Recovery: A Systematic Review of Current Approaches. Sports Med. 2018, 48, 683–703. [Google Scholar] [CrossRef] [PubMed]
- Peuhkuri, K.; Sihvola, N.; Korpela, R. Diet promotes sleep duration and quality. Nutr. Res. 2012, 32, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Doherty, R.; Madigan, S.; Warrington, G.; Ellis, J. Sleep and Nutrition Interactions: Implications for Athletes. Nutrients 2019, 11, 822. [Google Scholar] [CrossRef]
- Kim, J. Pre-sleep casein protein ingestion: New paradigm in post-exercise recovery nutrition. Phys. Act. Nutr. 2020, 24, 6–10. [Google Scholar] [CrossRef]
- Res, P.T.; Groen, B.; Pennings, B.; Beelen, M.; Wallis, G.A.; Gijsen, A.P.; Senden, J.M.G.; Van Loon, L.J.C. Protein Ingestion before Sleep Improves Postexercise Overnight Recovery. Med. Sci. Sports Exerc. 2012, 44, 1560–1569. [Google Scholar] [CrossRef]
- Snijders, T.; Res, P.T.; Smeets, J.S.J.; van Vliet, S.; van Kranenburg, J.; Maase, K.; Kies, A.K.; Verdijk, L.B.; van Loon, L.J.C. Protein Ingestion before Sleep Increases Muscle Mass and Strength Gains during Prolonged Resistance-Type Exercise Training in Healthy Young Men. J. Nutr. 2015, 145, 1178–1184. [Google Scholar] [CrossRef]
- Wall, B.T.; Burd, N.A.; Franssen, R.; Gorissen, S.H.M.; Snijders, T.; Senden, J.M.; Gijsen, A.P.; van Loon, L.J.C. Presleep protein ingestion does not compromise the muscle protein synthetic response to protein ingested the following morning. Am. J. Physiol.-Endocrinol. Metab. 2016, 311, E964–E973. [Google Scholar] [CrossRef]
- Trommelen, J.; Kouw, I.W.K.; Holwerda, A.M.; Snijders, T.; Halson, S.L.; Rollo, I.; Verdijk, L.B.; van Loon, L.J.C. Presleep dietary protein-derived amino acids are incorporated in myofibrillar protein during postexercise overnight recovery. Am. J. Physiol.-Endocrinol. Metab. 2018, 314, E457–E467. [Google Scholar] [CrossRef]
- Snijders, T.; Trommelen, J.; Kouw, I.W.K.; Holwerda, A.M.; Verdijk, L.B.; van Loon, L.J.C. The Impact of Pre-sleep Protein Ingestion on the Skeletal Muscle Adaptive Response to Exercise in Humans: An Update. Front. Nutr. 2019, 6. [Google Scholar] [CrossRef]
- Larsen, M.S.; Clausen, D.; Jørgensen, A.A.; Mikkelsen, U.R.; Hansen, M. Presleep Protein Supplementation Does Not Improve Recovery during Consecutive Days of Intense Endurance Training: A Randomized Controlled Trial. Int. J. Sport Nutr. Exerc. Metab. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Reis, C.E.G.; Loureiro, L.M.R.; Roschel, H.; da Costa, T.H.M. Effects of pre-sleep protein consumption on muscle-related outcomes—A systematic review. J. Sci. Med. Sport 2020. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.; Irani, N.; Balkrishnan, R.; Benny, I.R. A randomized, double blind, placebo controlled study to evaluate the effects of ashwagandha (Withania somnifera) extract on sleep quality in healthy adults. Sleep Med. 2020, 72, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Flynn, S.; Rosales, A.; Hailes, W.; Ruby, B. Males and females exhibit similar muscle glycogen recovery with varied recovery food sources. Eur. J. Appl. Physiol. 2020, 120, 1131–1142. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonilla, D.A.; Pérez-Idárraga, A.; Odriozola-Martínez, A.; Kreider, R.B. The 4R’s Framework of Nutritional Strategies for Post-Exercise Recovery: A Review with Emphasis on New Generation of Carbohydrates. Int. J. Environ. Res. Public Health 2021, 18, 103. https://doi.org/10.3390/ijerph18010103
Bonilla DA, Pérez-Idárraga A, Odriozola-Martínez A, Kreider RB. The 4R’s Framework of Nutritional Strategies for Post-Exercise Recovery: A Review with Emphasis on New Generation of Carbohydrates. International Journal of Environmental Research and Public Health. 2021; 18(1):103. https://doi.org/10.3390/ijerph18010103
Chicago/Turabian StyleBonilla, Diego A., Alexandra Pérez-Idárraga, Adrián Odriozola-Martínez, and Richard B. Kreider. 2021. "The 4R’s Framework of Nutritional Strategies for Post-Exercise Recovery: A Review with Emphasis on New Generation of Carbohydrates" International Journal of Environmental Research and Public Health 18, no. 1: 103. https://doi.org/10.3390/ijerph18010103
APA StyleBonilla, D. A., Pérez-Idárraga, A., Odriozola-Martínez, A., & Kreider, R. B. (2021). The 4R’s Framework of Nutritional Strategies for Post-Exercise Recovery: A Review with Emphasis on New Generation of Carbohydrates. International Journal of Environmental Research and Public Health, 18(1), 103. https://doi.org/10.3390/ijerph18010103