First Occurrence of Eustrongylides spp. (Nematoda: Dioctophymatidae) in a Subalpine Lake in Northwest Italy: New Data on Distribution and Host Range

 ,
, 
 ,
,  , ,
, , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Fish Collection and Anatomopathological and Parasitological Examination
2.3. Molecular Analysis
2.4. Statistical Analysis
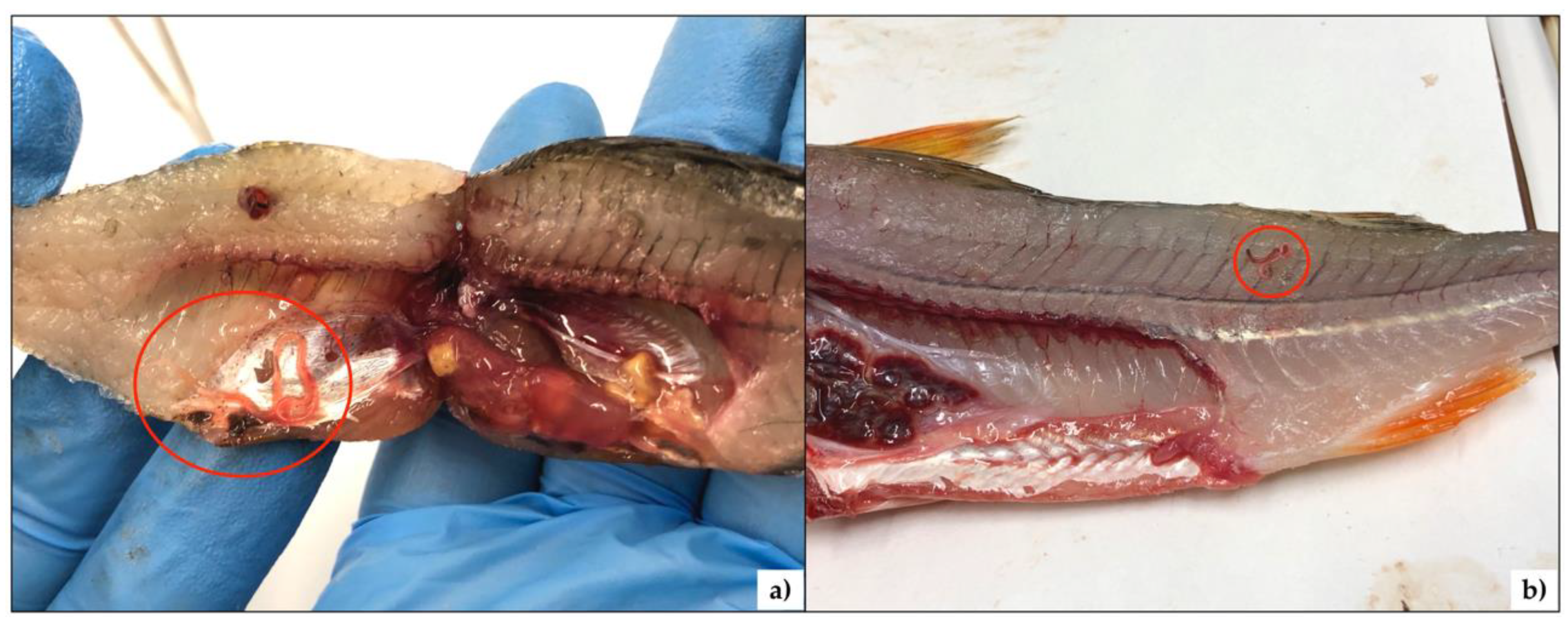
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- El-Dakhly, K.M.; El-Nahass, E.; Uni, S.; Tuji, H.; Sakai, H.; Yanai, T. Levels of infection of gastric nematodes in a flock of great cormorants (Phalacrocorax carbo) from Lake Biwa, Japan. J. Helminthol. 2012, 86, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Zhokhov, A.E.; Pugacheva, M.N. First Record of Eustrongylides excisus (Dorylaimea: Dioctophymatidae) in Fish of the Rybinsk Reservoir. Russ. J. Biol. Invasions 2019, 10, 325–327. [Google Scholar] [CrossRef]
- Measures, L.N. Revision of the genus Eustrongylides Jagerskiold, 1909 (Nematoda: Dioctophymatoidea) of piscivorous birds. Can. J. Zool. 1988, 66, 885–895. [Google Scholar] [CrossRef]
- Novakov, N.; Bjelic-Cabrilo, O.; Circovic, M.; Jubojevik, D.; Lujic, J. Eustrongylidosis of European Catfish (Siluris glaris). Bulg. J. Agric. Sci. 2013, 1, 72–76. [Google Scholar]
- Goncharov, S.L.; Soroka, N.M.; Pashkevich, I.Y.; Dubovy, A.I.; Bondar, A.O. Infection of predatory fish with larvae of Eustrongylides excisus (Nematoda, Dioctophymatidae) in the Delta of the Dnipro River and the Dnipro-Buh Estuary in Southern Ukraine. Vestn. Zool. 2018, 52, 137–144. [Google Scholar] [CrossRef]
- Guerin, P.F.; Marapendi, S.; MC Grail, L. Intestinal perforation caused by larval Eustrongylides. Morb. Mort. Week Rep. 1982, 31, 383–389. [Google Scholar]
- Paperna, I. Hosts distribution and pathology of infections with larvae of Eustrongylides (Dioctophymidae, Nematoda) in fish from East African lakes. J. Fish. Biol. 1974, 6, 67–76. [Google Scholar] [CrossRef]
- Lichtenfels, J.R.; Stroup, C.F. Eustrongylides ssp. (Nematoda: Dioctophymatoidea): First Report of an Invertebrate Host (Oligochaeta: Tubificidae) in North America. Proc. Helminthol. Soc. Wash. 1985, 52, 320–323. [Google Scholar]
- Pazouki, J.; Masoumian, M.; Yahyazadeh, M.; Abbasi, J. Metazoan Parasites from Freshwater Fishes of Northwest Iran. J. Agric. Sci. Technol. 2007, 9, 25–33. [Google Scholar]
- Soylu, E. Metazoan Parasites of Perch Perca fluviatilis L. from Lake Sığırcı, Ipsala, Turkey. Pak. J. Zool. 2013, 45, 47–52. [Google Scholar]
- Yesipova, N.B. The Spread of Parasitic Nematodes in Fish Eustrongylides Excisus Zaporozhye (Dnipro) Reservoir. 2013, pp. 86–88. Available online: https://www.degruyter.com/viewarticle/j$002fvzoo.2018.52.issue-2$002fvzoo-2018-0015$002fvzoo-2018-0015.xml (accessed on 9 May 2020). (In Russian).
- Fedorov, N.M.; Firsov, N.F.; Soloviev, N.A. Veterinary and sanitary examination in river perch with Eustrongylidosis. Vet. Pathol. 2014, 3, 68–73. [Google Scholar]
- Melo, F.T.; Melo, C.S.; Nascimento, L.C. Morphological characterization of Eustrongylides ssp. Larvae (Nematoda, Dioctophymatoidea) parasite of Rhinella marina (Amphibia: Bufonidae) from Eastern Amazonia. Braz. J. Vet. Parasitol. Jaboticabal. 2015, 7–12. [Google Scholar] [CrossRef]
- Noei, M.R.; Ibrahimov, S.; Sattari, M. Parasitic worms of the Persian sturgeon, Acipenser persicus Borodin, 1897 from the southwestern shores of the Caspian Sea. Iran. J. Ichthyol. 2015, 2, 287–295. [Google Scholar] [CrossRef]
- Branciari, R.; Ranucci, D.; Miraglia, D.; Valiani, A.; Veronesi, F.; Urbani, E.; Lo Vaglio, G.; Pascucci, L.; Franceschini, R. Occurrence of parasites of the genus Eustrongylides spp. (Nematoda: Dioctophymatidae) in fish caught in Trasimeno lake, Italy. Ital. J. Food Saf. 2016, 5, 6130. [Google Scholar] [CrossRef]
- Švažas, S.; Chukalova, N.; Grishanov, G.; Pūtys, Ž.; Sruoga, A.; Butkauskas, D.; Raudonikis, L.; Prakas, P. The role of great cormorant (Phalacrocorax carbo sinensis) for fish stock and dispersal of helminthes parasites in the Curonian lagoon area. Vet. Med. Zoot. 2011, 55, 79–85. [Google Scholar]
- Menconi, V.; Pastorino, P.; Cavazza, G.; Santi, M.; Mugetti, D.; Zuccaro, G.; Prearo, M. The role of live fish trade in the translocation of parasites: The case of Cystidicola farionis in farmed rainbow trout (Oncorhynchus mykiss). Aquac. Int. 2019, 27, 1667–1671. [Google Scholar] [CrossRef]
- Dezfuli, B.S.; Manera, M.; Lorenzoni, M.; Pironi, F.; Shinn, A.P.; Giari, L. Histopathology and the inflammatory response of European perch, Perca fluviatilis muscle infected with Eustrongylides sp. (Nematoda). Parasit. Vectors 2015, 8, 227. [Google Scholar] [CrossRef] [PubMed]
- Agnetti, F.; Sensidoni, L.; Di Raimo Marrocchi, E.; Vaglio, G.; Sgariglia, E.; Valentini, A.; Ghittino, C.; Caffara, M.; Gustinelli, A.; Fioravanti, M.L. Presence of Eustrongylides sp. (Nematoda: Dioctophymatidae) in fish species of the Trasimeno lake (Umbria): Preliminary data. In Proceedings of the XVII Congresso Nazionale, S.I.Di.L.V. Società Italiana di Diagnostica di Laboratorio Veterinaria: Pacengo di Lazise (VR), Lazise, Italy, 28–30 September 2016; pp. 253–254. [Google Scholar]
- Mazzone, A.; Caffara, M.; Gustinelli, A.; Agnetti, F.; Sgariglia, E.; Lo Vaglio, G.; Quaglio, F.; Fioravanti, M.L. Morphological and molecular characterization of larval and adult stages of Eustrongylides excisus (Nematoda: Dioctophymatoidea) with histopathological observations. J. Parasitol. 2019, 105, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, M.L.; Hurwitz, H.; Sun, A.M.; Coletta, D. Intestinal perforation caused by larval Eustrongylides (Nematoda: Dioctophymatoidae) in New Jersey. Am. J. Trop. Med. Hyg. 1989, 40, 648–650. [Google Scholar] [CrossRef]
- Caudill, G.; Wolf, D.; Caudill, D.; Brown, J.; Shearn-Bochsler, V.A. Juvenile wading-bird mortality event in urban Jacksonville, Florida, associated with the parasite Eustrongylides. Florida Field Nat. 2014, 42, 108–113. [Google Scholar]
- Cole, R.A. Eustrongylidosis. In Field Manual of Wildlife Diseases: General Field Procedures and Diseases of Birds; Friend, M., Franson, J.C., Eds.; U.S. Department of the Interior Fish and Wildlife Service: Washington, DC, USA, 1999; pp. 223–228. [Google Scholar]
- Moravec, F. Nematodes parasitic in fishes as larvae. In Parasitic Nematodes of Freshwater Fishes of Europe; Moravec, F., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; pp. 374–421. [Google Scholar]
- Bjeli-Cabrilo, O.; Novakov, N.; Irkovi, M.; Kosti, D.; Popovi, E.; Aleksi, N.; Luji, J. The first determination of Eustrongylides excisus Jägerskiöld, 1909: Larvae (Nematoda: Dioctophymatidae) in the pike-perch Sander lucioperca in Vojvodina (Serbia). Helminthologia 2013, 50, 291–294. [Google Scholar] [CrossRef]
- Wittner, M.; Turner, J.W.; Jacquette, G.; Ash, L.R.; Salgo, M.P.; Tanowitz, H.B. Eustrongylidiasis: A parasitic infection acquired by eating sushi. N. Engl. J. Med. 1989, 320, 1124–1126. [Google Scholar] [CrossRef] [PubMed]
- Gunby, P. One worm in the minnow equals too many in the gut. JAMA 1982, 248, 163. [Google Scholar] [CrossRef]
- Narr, L.L.; O’Donnell, J.G.; Libster, B.; Alessi, P.; Abraham, D. Eustrongylidiasis a parasitic infection acquired by eating live minnow. J. Am. Osteopath. Assoc. 1996, 96, 400–402. [Google Scholar] [CrossRef]
- Eberhard, M.L.; Ruiz-Tiben, E. Cutaneous emergence of Eustrongylides in two persons from South Sudan. Am. J. Trop. Med. Hyg. 2014, 90, 315–317. [Google Scholar] [CrossRef]
- Minuzzo, C.; Tisi, A.; Caramiello, R.; Siniscalco, C. Flora acquatica e palustre della zona dei “Cinque Laghi” di Ivrea. Riv. Piem. St. Nat. 2005, 26, 41–71. [Google Scholar]
- CEN. Water Quality-Sampling of Fish with Multi-Mesh Gillnets (EN 14757: 2005); CEN: Brussels, Belgium, 2005. [Google Scholar]
- Gustinelli, A.; Caffara, M.; Florio, D.; Elick, O.O.; Euty, M.W.; Fioravanti, M.L. First description of the adult stage of Clinostomum cutaneum Paperna, 1964 (Digenea: Clinostomidae) from grey herons Ardea cinerea L. and a redescription of the metacercaria from the Nile tilapia Oreochromis niloticus niloticus (L.) in Kenya. Syst. Parasitol. 2010, 76, 39–51. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, M.; Shostak, A.W. Parasitology meets ecology. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Chai, J.Y.; Murrell, K.D.; Lymbery, A.J. Fish-borne parasitic zoonoses: Status and issues. Int. J. Parasitol. 2005, 35, 1233–1254. [Google Scholar] [CrossRef]
- Sattari, M. The occurrence and intensity of Eustrongylides excisus (Nematoda: Dioctophymidae) in some bony fish species of Caspian Sea and its basin. Casp. J. Environ. Sci. 2004, 2, 9–12. [Google Scholar]
- Coyner, D.F.; Spalding, M.G.; Forrester, D.J. Epizootiology of Eustrongylides ignotus in Florida: Distribution, density, and natural infections in intermediate hosts. J. Wildl. Dis. 2002, 38, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Menconi, V.; Manfrin, C.; Pastorino, P.; Mugetti, D.; Cortinovis, L.; Pizzul, E.; Pallavicini, A.; Prearo, M. First Report of Clinostomum complanatum (Trematoda: Digenea) in European Perch (Perca fluviatilis) from an Italian Subalpine Lake: A Risk for Public Health? Int. J. Environ. Res. Public Health 2020, 17, 1389. [Google Scholar] [CrossRef] [PubMed]
- Gagliardi, A.; Preatoni, D.; Wauters, L.; Martinoli, A. Selective predators or choosy fishermen? Relation between fish harvest, prey availability and great cormorant (Phalacrocorax carbo sinensis) diet. Ital. J. Zool. 2015, 82, 544–555. [Google Scholar] [CrossRef]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Tottrup, A.P.; Thorup, K.; Rainio, K.; Yosef, R.; Lehikoinen, E.; Rahbek, C. Avian migrants adjust migration inresponse to environmental conditions en route. Biol. Lett. 2008, 4, 685–688. [Google Scholar] [CrossRef]
- Polley, L.; Thompson, R.C.A. Parasite zoonoses and climate change: Molecular tools for tracking shifting boundaries. Trends Parasitol. 2009, 25, 285–291. [Google Scholar] [CrossRef]
- Measures, L.N. Epizootiology, pathology, and description of Eustrongylides tubifex (Nematoda: Dioctophymatoidea) in fish. Can. J. Zool. 1988, 66, 2212–2222. [Google Scholar] [CrossRef]
- Persson, L.; Diehl, S.; Johansson, L.; Andersson, G.; Hamrin, S.F. Shifts in fish communities along the productivity gradient of temperate lakes—patterns and the importance of size-structured interactions. J. Fish Biol. 1991, 38, 281–293. [Google Scholar] [CrossRef]
- Hatzenbeler, G.R.; Bozek, M.A.; Jennings, M.J.; Emmons, E.E. Seasonal variation in fish assemblage structure and habitat structure in the nearshore littoral zone of Wisconsin lakes. N. Am. J. Fish. Manag. 2000, 20, 360–368. [Google Scholar] [CrossRef]
- Long, J.M.; Fisher, W.L. Inter-annual and size-related differences in the diets of three sympatric black bass in an Oklahoma reservoir. J. Freshw. Ecol. 2000, 15, 465–474. [Google Scholar] [CrossRef]
- Scheuerell, M.D.; Schindler, D.E. Changes in the spatial distribution of fishes in lakes along a residential development gradient. Ecosystems 2004, 7, 98–106. [Google Scholar] [CrossRef]
- Persson, L. Asymmetries in competitive and predatory interactions in fish populations. In Size-Structured Populations-Ecology and Evolution; Ebenman, B., Persson, L., Eds.; Springer: Berlin, Germany, 1988; pp. 203–218. [Google Scholar]
- García--Berthou, E. Ontogenetic diet shifts and interrupted piscivoryin introduced largemouth bass (Micropterus salmoides). Internat. Rev. Hydrobiol. 2002, 87, 353–363. [Google Scholar] [CrossRef]
- Wainwright, P.C.; Osenberg, C.W.; Mittelbach, G.G. Trophic polymorphism in the pumpkinseed sunfish (Lepomis gibbosus Linnaeus): Effects of environment on ontogeny. Funct. Ecol. 1991, 5, 40–55. [Google Scholar] [CrossRef]
- Azim, S.; Dojki, F.; Ahmad, S.S.; Beg, A.M. Role of Human Behaviour and Parasitic Diseases. IFDJP 2008, 17, 128–134. [Google Scholar]
- Volta, P.; Jeppesen, E.; Sala, P.; Galafassi, S.; Foglini, C.; Puzzi, C.; Winfield, I.J. Fish assemblages in deep talian subalpine lakes: History and present status with an emphasis on non-native species. Hydrobiologia 2018, 824, 255–270. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Fish Species | Captured Fish No. | Total Length (cm) | Total Weight (g) |
|---|---|---|---|
| Ameiurus melas | 30 | 20.1 ± 2.1 | 102.3 ± 35.3 |
| Ictalurus punctatus | 30 | 26.8 ± 15.1 | 58.2 ± 28.1 |
| Squalius cephalus | 30 | 37.7 ± 5.8 | 876.9 ± 352.1 |
| Carassius carassius | 30 | 34.8 ± 6.2 | 985 ± 623.7 |
| Lepomis gibbosus | 82 | 10.9 ± 2.4 | 32.6 ± 18.3 |
| Micropterus salmoides | 30 | 15.6 ± 3.7 | 62.2 ± 56.4 |
| Scardinius erythrophthalmus | 30 | 21.3 ± 2.1 | 165 ± 4.5 |
| Perca fluviatilis | 30 | 14.3 ± 2.1 | 42.4 ± 41.4 |
| Lepomis Gibbosus | Micropterus Salmoides | Perca Fluviatilis | |
|---|---|---|---|
| Mean intensity | 1.15 | 1 | 1 |
| Mean abundance | 0.28 | 0.16 | 0.1 |
| Number of larvae | 23 | 5 | 3 |
| Location of larvae | 2 PD; 5 PV; 16 VC | 3 AV; 2 VC | 2 AD; 1 VC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menconi, V.; Riina, M.V.; Pastorino, P.; Mugetti, D.; Canola, S.; Pizzul, E.; Bona, M.C.; Dondo, A.; Acutis, P.L.; Prearo, M. First Occurrence of Eustrongylides spp. (Nematoda: Dioctophymatidae) in a Subalpine Lake in Northwest Italy: New Data on Distribution and Host Range. Int. J. Environ. Res. Public Health 2020, 17, 4171. https://doi.org/10.3390/ijerph17114171
Menconi V, Riina MV, Pastorino P, Mugetti D, Canola S, Pizzul E, Bona MC, Dondo A, Acutis PL, Prearo M. First Occurrence of Eustrongylides spp. (Nematoda: Dioctophymatidae) in a Subalpine Lake in Northwest Italy: New Data on Distribution and Host Range. International Journal of Environmental Research and Public Health. 2020; 17(11):4171. https://doi.org/10.3390/ijerph17114171
Chicago/Turabian StyleMenconi, Vasco, Maria Vittoria Riina, Paolo Pastorino, Davide Mugetti, Serena Canola, Elisabetta Pizzul, Maria Cristina Bona, Alessandro Dondo, Pier Luigi Acutis, and Marino Prearo. 2020. "First Occurrence of Eustrongylides spp. (Nematoda: Dioctophymatidae) in a Subalpine Lake in Northwest Italy: New Data on Distribution and Host Range" International Journal of Environmental Research and Public Health 17, no. 11: 4171. https://doi.org/10.3390/ijerph17114171
APA StyleMenconi, V., Riina, M. V., Pastorino, P., Mugetti, D., Canola, S., Pizzul, E., Bona, M. C., Dondo, A., Acutis, P. L., & Prearo, M. (2020). First Occurrence of Eustrongylides spp. (Nematoda: Dioctophymatidae) in a Subalpine Lake in Northwest Italy: New Data on Distribution and Host Range. International Journal of Environmental Research and Public Health, 17(11), 4171. https://doi.org/10.3390/ijerph17114171