Bird Satellite Tracking Revealed Critical Protection Gaps in East Asian–Australasian Flyway


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. GPS Tracking
2.3. Delineating Migration Routes and Stopover Sites
2.4. Habitat Use at Stopover Sites and Protection Gap Analysis
- >75% Totally/satisfactorily protected;
- 25–75% Under-protected;
- <25% Critically under-protected; and
- =0 unprotected.
3. Results
3.1. General Description of Spring Migration of Eastern Geese
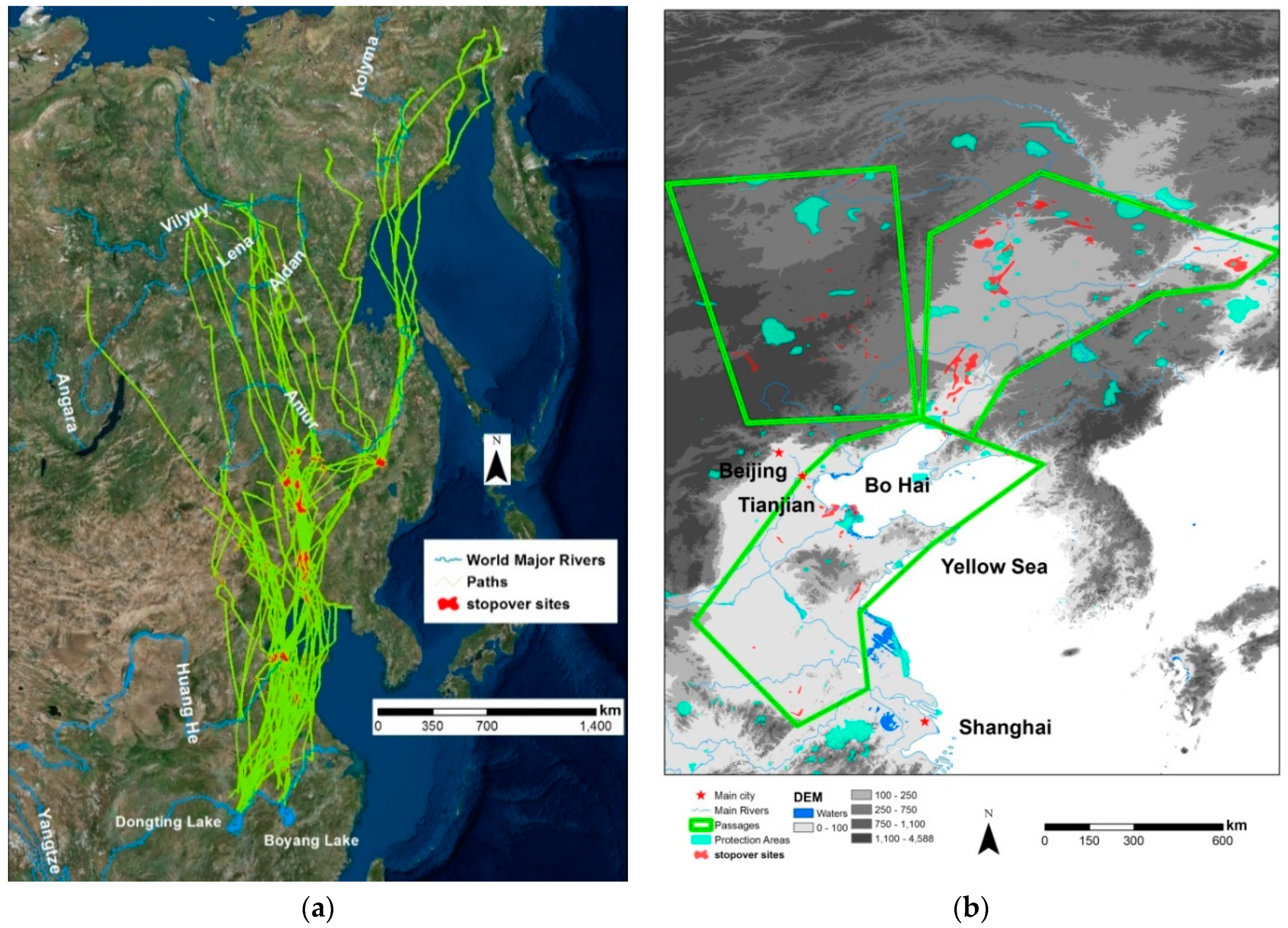
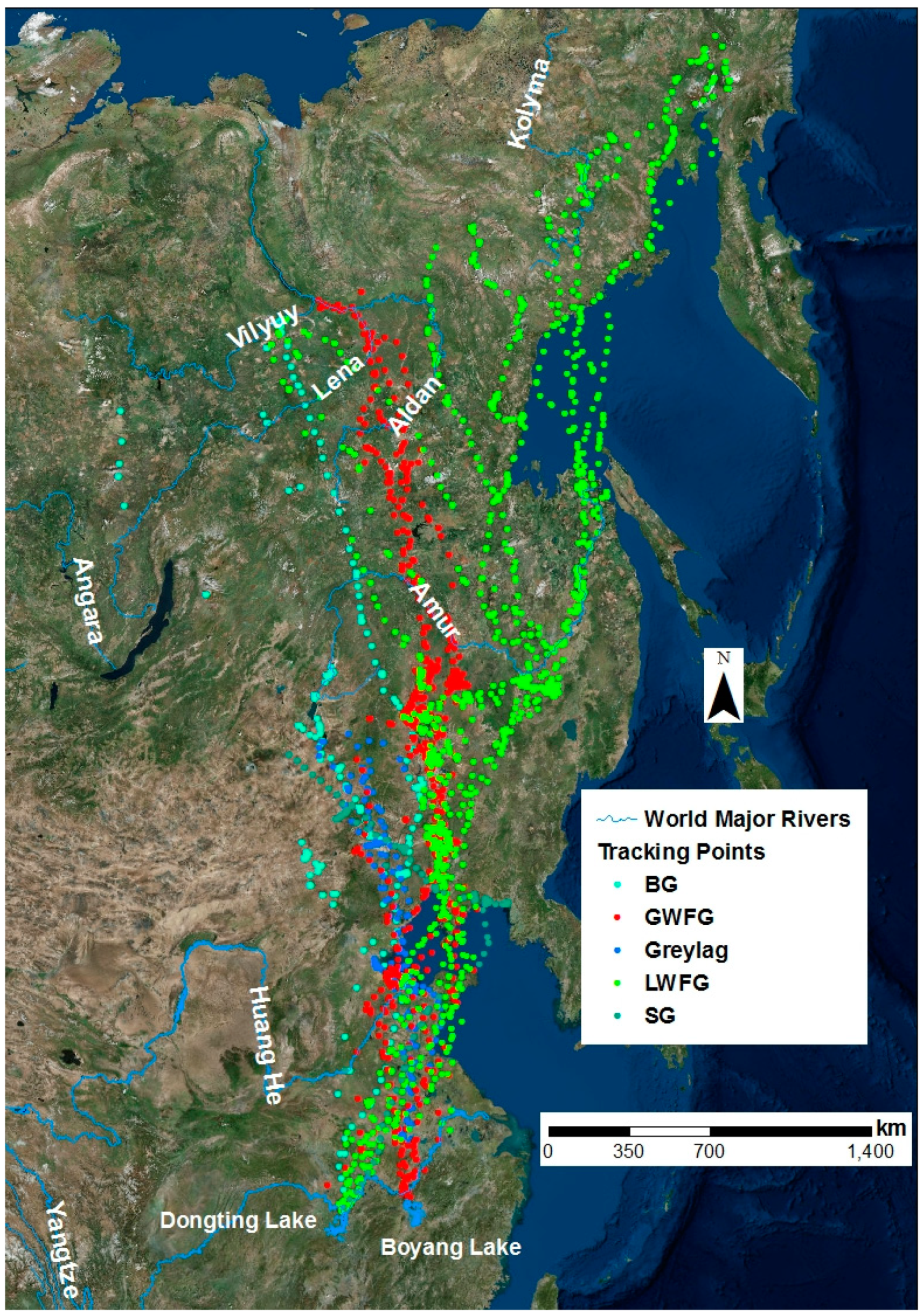
3.2. Migration Paths and Stopover Sites
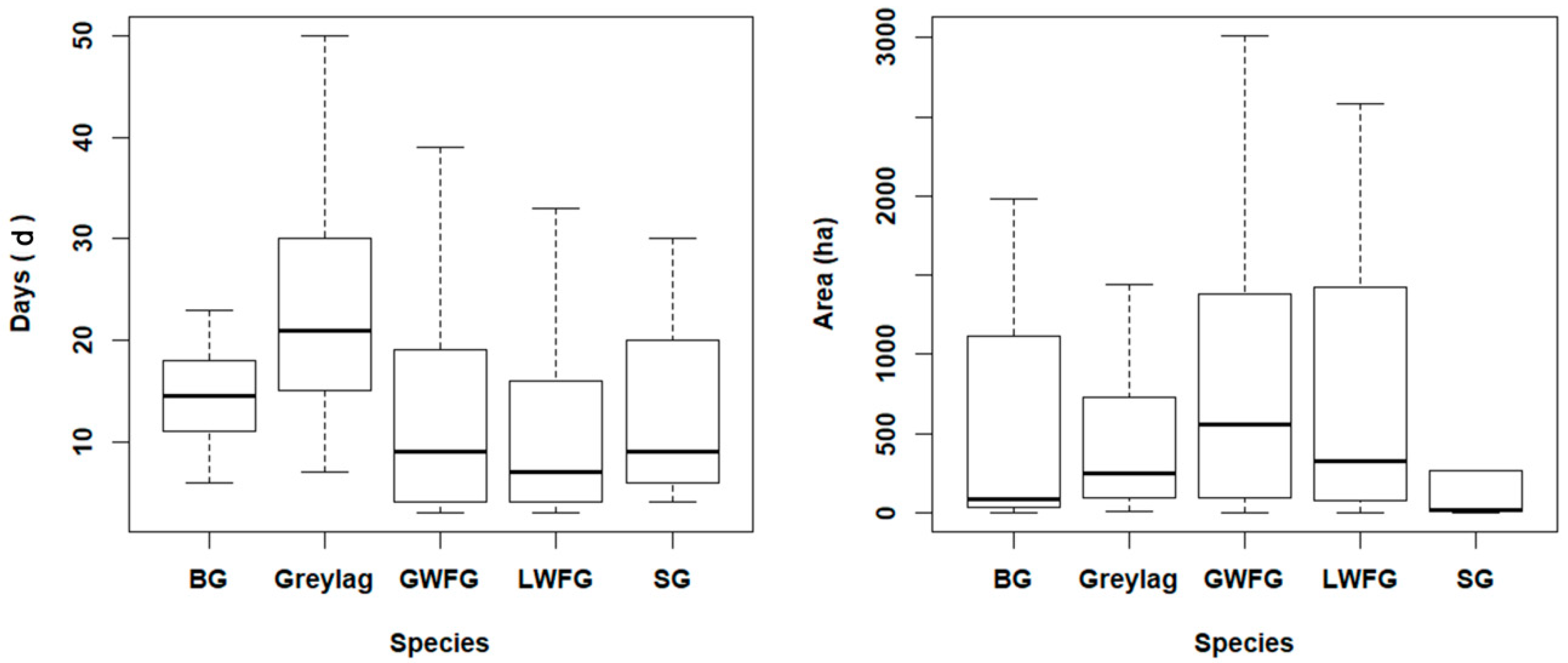
3.3. Habitat Use and Gaps in Protection
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Route ID 1 | Year | Start (Year/Month/Day) | End (Year/Month/Day) | Fixes | Interval 2 (hour) | Interval 3 (hour) | Distance 4 (km) | Airspeed 5 (km/h) |
|---|---|---|---|---|---|---|---|---|
| BG016.17 | 2017 | 22/03/2017 | 12/05/2017 | 733 | 2 | 1.7 (1.2) | 5944 | 4.82 (18.05) |
| BG05.16 | 2016 | 12/03/2016 | 13/05/2016 | 1155 | 1 | 7.9 (223.2) | 5643 | 3.81 (18.72) |
| BG09.17 | 2017 | 25/03/2017 | 24/05/2017 | 1410 | 1 | 1 (0.5) | 9175 | 6.40 (23.69) |
| BG22.17 | 2017 | 30/03/2017 | 3/06/2017 | 144 | 4 | 11 (18.4) | 5859 | 3.72 (8.75) |
| Greylag02.15 | 2015 | 12/03/2015 | 15/04/2015 | 681 | 1 | 12.5 (295.2) | 2458 | 3.08 (14.28) |
| Greylag02.16 | 2016 | 1/03/2016 | 13/04/2016 | 1717 | 1 | 4.9 (118.9) | 5040 | 4.62 (22.63) |
| Greylag02.17 | 2017 | 11/02/2017 | 10/04/2017 | 1369 | 1 | 1 (0.00) | 4505 | 3.29 (18.14) |
| Greylag04.15 | 2015 | 2/03/2015 | 14/04/2015 | 890 | 1 | 9.5 (249.2) | 3877 | 3.77 (20.07) |
| Greylag04.16 | 2016 | 18/02/2016 | 1/04/2016 | 1037 | 1 | 1 (0.1) | 4464 | 4.30 (21.54) |
| GWFG01.15 | 2015 | 22/03/2015 | 25/05/2015 | 1369 | 1 | 6.5 (198.4) | 10,074 | 6.57 (23.82) |
| GWFG01.16 | 2016 | 26/03/2016 | 20/05/2016 | 1271 | 1 | 1 (0.3) | 11,382 | 8.64 (27.84) |
| GWFG02.15 | 2015 | 26/03/2015 | 29/04/2015 | 708 | 1 | 12.2 (292.6) | 5009 | 6.12 (24.28) |
| GWFG03.16 | 2016 | 19/03/2016 | 12/05/2016 | 569 | 2 | 15.7 (319.3) | 10,424 | 7.98 (22.87) |
| GWFG03.17 | 2017 | 26/03/2017 | 8/05/2017 | 963 | 1 | 1.1 (0.8) | 6623 | 6.34 (22.12) |
| GWFG04.15 | 2015 | 22/03/2015 | 28/04/2015 | 398 | 1 | 1 (0.6) | 3512 | 8.52 (28.69) |
| GWFG05.15 | 2015 | 26/03/2015 | 19/05/2015 | 1070 | 1 | 8.3 (230.2) | 11,533 | 8.91 (29.1) |
| GWFG05.16 | 2016 | 28/03/2016 | 19/05/2016 | 1103 | 1 | 7.9 (225.2) | 10,425 | 8.39 (31.83) |
| GWFG05.17 | 2017 | 26/03/2017 | 14/05/2017 | 418 | 2 | 2.8 (1.7) | 7826 | 6.76 (19.41) |
| GWFG06.17 | 2017 | 23/03/2017 | 7/06/2017 | 1472 | 1 | 1.2 (5.4) | 9770 | 5.35 (21.43) |
| LWFG12.16 | 2016 | 4/03/2016 | 4/06/2016 | 1599 | 1 | 5.8 (177) | 15,536 | 7.00 (27.95) |
| LWFG12.17 | 2017 | 26/03/2017 | 25/05/2017 | 1436 | 1 | 1 (0.1) | 16,172 | 11.21 (33.14) |
| LWFG14.16 | 2016 | 26/03/2016 | 20/05/2016 | 1234 | 1 | 7.1 (211.8) | 11,027 | 8.46 (26.26) |
| LWFG14.17 | 2017 | 26/03/2017 | 28/05/2017 | 1474 | 1 | 1 (0.2) | 12,880 | 8.56 (25.62) |
| LWFG28.17 | 2017 | 26/03/2017 | 24/05/2017 | 1457 | 1 | 1 (0.1) | 12,575 | 8.82 (27.64) |
| LWFG29.17 | 2017 | 26/03/2017 | 14/05/2017 | 595 | 2 | 2 (0.9) | 13,665 | 11.66 (31.75) |
| LWFG38.17 | 2017 | 26/03/2017 | 11/06/2017 | 644 | 3 | 2.9 (1.2) | 8736 | 4.74 (18.72) |
| LWFG40.17 | 2017 | 23/03/2017 | 18/06/2017 | 1001 | 2 | 2.1 (1.2) | 15,152 | 7.22 (24.31) |
| LWFG42.17 | 2017 | 23/03/2017 | 18/06/2017 | 1994 | 1 | 1.1 (0.5) | 15,632 | 7.44 (25.75) |
| LWFG45.17 | 2017 | 25/03/2017 | 11/05/2017 | 854 | 1 | 1.3 (0.6) | 14,387 | 12.8 (31.41) |
| LWFG47.17 | 2017 | 25/03/2017 | 13/06/2017 | 1812 | 1 | 1.1 (0.4) | 11,163 | 5.86 (23.03) |
| LWFG62.17 | 2017 | 26/03/2017 | 14/05/2017 | 590 | 2 | 2 (0.4) | 14,515 | 12.41 (34.27) |
| SG04.17 | 2017 | 8/03/2017 | 10/04/2017 | 775 | 1 | 1.5 (12.5) | 5568 | 7.08 (22.47) |
| SG05.17 | 2017 | 24/04/2017 | 3/07/2017 | 157 | 4 | 10.8 (17.2) | 2951 | 1.76 (5.94) |
| Order | ID | Setup Time (Year/Month/Day) | Last Communication (Year/Month/Day) | Total Working Days | Device Status Description | Necrology | Status of Last Location | Cause of Death |
|---|---|---|---|---|---|---|---|---|
| 1 | LWFG01·BFUL005 | 2015/12/6 | 2015/12/19 | 13.00 | Lost | wintering | ||
| 2 | LWFG02·BFUL006 | 2015/12/6 | 2016/5/28 | 174.00 | Lost | migrating(China) | ||
| 3 | LWFG03·BFUL004 | 2015/12/6 | 2016/9/8 | 277.00 | death | migrating(China) | ||
| 4 | LWFG04·BFUL007 | 2015/12/6 | 2016/4/1 | 117.00 | Lost | migrating(China) | ||
| 5 | LWFG05·BFUL008 | 2015/12/11 | 2015/12/13 | 2.00 | Lost | wintering | ||
| 6 | LWFG06·BFUL009 | 2015/12/11 | 2016/4/24 | 135.00 | Lost | migrating(Boundary) | ||
| 7 | LWFG07·BFUL010 | 2015/12/31 | 2016/3/15 | 75.00 | Lost | wintering | ||
| 8 | LWFG08·BFUL014 | 2015/12/30 | 2016/3/31 | 92.00 | Lost | migrating(China) | ||
| 9 | LWFG09·BFUL005 | 2015/12/5 | 2016/5/10 | 157.00 | Lost | migrating(Boundary) | ||
| 10 | LWFG10·BFUL013 | 2016/1/14 | 2016/3/31 | 77.00 | Lost | migrating(China) | ||
| 11 | LWFG11·BFUL011 | 2016/1/14 | 2016/5/6 | 113.00 | Lost | migrating(Boundary) | ||
| 12 | LWFG12·BFUL24 | 2016/1/14 | 2018/5/13 | 850.00 | normal | |||
| 13 | LWFG14·BFUL003 | 2016/1/31 | 2018/5/7 | 827.00 | normal | |||
| 14 | LWFG22·BFUL040 | 2016/11/22 | 2016/11/26 | 4.00 | death | wintering | ||
| 15 | LWFG23·BFUL041 | 2016/11/22 | 2018/4/16 | 510.00 | Lost | migrating(Boundary) | ||
| 16 | LWFG24·BFUL042 | 2016/11/22 | 2016/12/3 | 11.00 | death | wintering | ||
| 17 | LWFG25·BFUL043 | 2016/11/22 | 2017/5/17 | 176.00 | Lost | migrating(Boundary) | ||
| 18 | LWFG26·BFUL040 | 2016/11/27 | 2017/6/1 | 186.00 | death | migrating(Boundary) | hunting | |
| 19 | LWFG27·BFUL049 | 2016/11/27 | 2016/12/3 | 6.00 | Lost | wintering | ||
| 20 | LWFG28·BFUL050 | 2016/11/27 | 2018/5/19 | 538.00 | normal | |||
| 21 | LWFG29·BFUL051 | 2016/11/27 | 2018/11/14 | 717.00 | normal | |||
| 22 | LWFG30·BFUL052 | 2016/11/27 | 2016/12/2 | 5.00 | Lost | wintering | ||
| 23 | LWFG31·BFUL053 | 2016/11/27 | 2016/12/21 | 24.00 | death | wintering | ||
| 24 | LWFG32·BFUL042 | 2016/12/2 | 2017/5/12 | 161.00 | death | migrating(China) | ||
| 25 | LWFG33·BFUL044 | 2016/12/2 | 2018/6/9 | 554.00 | Lost | migrating(Boundary) | ||
| 26 | LWFG34·BFUL052 | 2016/12/2 | 2017/6/14 | 194.00 | Lost | migrating(Boundary) | ||
| 27 | LWFG35·BFUL054 | 2016/12/2 | 2017/4/16 | 135.00 | death | migrating(China) | poison | |
| 28 | LWFG36·BFUL055 | 2016/12/2 | 2017/4/20 | 139.00 | Lost | migrating(Boundary) | ||
| 29 | LWFG38·BFUL057 | 2016/12/2 | 2017/7/17 | 227.00 | normal | |||
| 30 | LWFG39·BFUL058 | 2016/12/2 | 2017/4/7 | 126.00 | death | migrating(China) | poison | |
| 31 | LWFG40·BFUL059 | 2016/12/2 | 2017/12/29 | 392.00 | normal | wintering | ||
| 32 | LWFG41·BFUL060 | 2016/12/2 | 2017/6/22 | 202.00 | Lost | migrating(Boundary) | ||
| 33 | LWFG42·BFUL061 | 2016/12/2 | 2018/11/14 | 712.00 | normal | |||
| 34 | LWFG43·BFUL064 | 2016/12/7 | 2017/7/4 | 209.00 | Lost | migrating(Boundary) | ||
| 35 | LWFG44·BFUL065 | 2016/12/7 | 2017/9/5 | 272.00 | Lost | migrating(Boundary) | ||
| 36 | LWFG45·BFUL062 | 2016/12/7 | 2018/11/14 | 707.00 | normal | |||
| 37 | LWFG46·BFUL067 | 2016/12/16 | 2018/5/9 | 509.00 | Lost | migrating(Boundary) | ||
| 38 | LWFG47·BFUL068 | 2016/12/16 | 2018/5/28 | 528.00 | normal | |||
| 39 | LWFG48·BFUL086_HQP172 | 2016/12/26 | 2018/5/12 | 502.00 | Lost | migrating(Boundary) | ||
| 40 | LWFG49·BFUL089_HQP165 | 2016/12/26 | 2018/5/8 | 498.00 | Lost | migrating(Boundary) | ||
| 41 | LWFG50·BFUL085_HQP139 | 2016/12/26 | 2017/4/4 | 99.00 | Lost | migrating(China) | ||
| 42 | LWFG51·BFUL084_HQP174 | 2016/12/26 | 2017/1/26 | 31.00 | Lost | wintering | ||
| 43 | LWFG52·BFUL087_HQP170 | 2016/12/26 | 2017/4/29 | 124.00 | Lost | migrating(Boundary) | ||
| 44 | LWFG53·BFUL083_HQP168 | 2016/12/26 | 2017/8/12 | 229.00 | Lost | migrating(China) | ||
| 45 | LWFG54·BFUL092_HQP150 | 2016/12/27 | 2017/5/10 | 134.00 | Lost | migrating(Boundary) | ||
| 46 | LWFG55·BFUL091_HQP153 | 2016/12/27 | 2018/5/5 | 494.00 | Lost | migrating(China) | ||
| 47 | LWFG56·BFUL090_HQP166 | 2016/12/27 | 2017/3/11 | 74.00 | death | wintering | ||
| 48 | LWFG57·BFUL088_HQP175 | 2016/12/27 | 2017/5/9 | 133.00 | Lost | migrating(Boundary) | ||
| 49 | LWFG58·BFUL082 | 2016/12/27 | 2017/5/11 | 135.00 | Lost | wintering | ||
| 50 | LWFG59·BFUL063 | 2017/1/18 | 2017/3/27 | 68.00 | Lost | migrating(China) | ||
| 51 | LWFG60·BFUL071 | 2017/1/18 | 2017/5/17 | 119.00 | Lost | migrating(Boundary) | ||
| 52 | LWFG61·BFUL073 | 2017/1/18 | 2017/10/7 | 262.00 | Lost | migrating(Boundary) | ||
| 53 | LWFG62·BFUL074 | 2017/1/18 | 2017/4/26 | 98.00 | normal | migrating(Boundary) | ||
| 54 | LWFG63·BFUL095 | 2017/1/18 | 2017/5/25 | 127.00 | Lost | migrating(China) | ||
| 55 | LWFG64·BFUL070 | 2017/1/18 | 2017/10/1 | 256.00 | Lost | migrating(Boundary) | ||
| 56 | LWFG65·BFUL072 | 2017/2/14 | 2017/12/12 | 301.00 | death | migrating(Boundary) | hunting | |
| 57 | LWFG66·BFUL096 | 2017/2/14 | 2017/4/6 | 51.00 | Lost | migrating(China) | ||
| 58 | LWFG67·BFUL097 | 2017/2/14 | 2017/4/22 | 67.00 | death | migrating(China) | ||
| 59 | LWFG68·BFUL138 | 2017/2/18 | 2017/3/6 | 16.00 | Lost | wintering | ||
| 60 | LWFG69·BFUL113 | 2017/3/2 | 2017/3/15 | 13.00 | Lost | wintering | ||
| 61 | LWFG70·BFUL114 | 2017/3/2 | 2017/4/18 | 47.00 | death | migrating(China) | ||
| 62 | LWFG71·BFUL115 | 2017/3/2 | 2017/3/25 | 23.00 | Lost | wintering | ||
| 63 | LWFG72·BFUL116 | 2017/3/2 | 2017/5/15 | 74.00 | Lost | migrating(Boundary) | ||
| 64 | LWFG73·BFUL117 | 2017/3/2 | 2018/3/21 | 384.00 | Lost | wintering | ||
| 65 | LWFG74·BFUL119 | 2017/3/2 | 2017/3/5 | 3.00 | Lost | wintering | ||
| 66 | LWFG75·BFUL121 | 2017/3/2 | 2017/5/16 | 75.00 | Lost | migrating(Boundary) | ||
| 67 | LWFG76·BFUL122 | 2017/3/2 | 2017/5/16 | 75.00 | Lost | migrating(Boundary) | ||
| 68 | LWFG77·BFUL123 | 2017/3/2 | 2017/5/15 | 74.00 | Lost | migrating(Boundary) | ||
| 69 | LWFG78·BFUL128 | 2017/3/13 | 2017/3/29 | 16.00 | Lost | wintering | ||
| 70 | LWFG79·BFUL129 | 2017/3/13 | 2017/3/14 | 1.00 | Lost | wintering | ||
| 71 | LWFG80·BFUL130 | 2017/3/13 | 2017/3/27 | 14.00 | Lost | migrating(China) | ||
| 72 | LWFG81·BFUL131 | 2017/3/13 | 2017/3/13 | 0.00 | Lost | wintering | ||
| 73 | LWFG82·BFUL132 | 2017/3/13 | 2017/3/13 | 0.00 | Lost | wintering | ||
| 74 | LWFG83·BFUL151 | 2017/3/14 | 2017/5/15 | 62.00 | Lost | migrating(Boundary) | ||
| 75 | GG01·BFUL045 | 2016/11/22 | 2016/12/8 | 16.00 | death | wintering | poison | |
| 76 | GG02 ·CAS003 | 2014/12/29 | 2017/5/14 | 867.00 | normal | |||
| 77 | GG03·BFUL046 | 2014/12/29 | 2018/1/14 | 1112.00 | Lost | migrating(China) | ||
| 78 | GG04·CAS004 | 2014/12/29 | 2016/7/5 | 554.00 | normal | |||
| 79 | GG05·BYH020 | 2014/12/29 | 2014/12/31 | 2.00 | Lost | wintering | ||
| 80 | GG06·CAS010 | 2014/12/29 | 2015/1/9 | 11.00 | Lost | wintering | ||
| 81 | GG07·BYH013 | 2014/12/29 | 2015/3/4 | 65.00 | Lost | migrating(China) | ||
| 82 | GWFG01·BYH024 | 2014/12/29 | 2016/7/3 | 552.00 | normal | |||
| 83 | GWFG02·BYH005 | 2014/12/29 | 2015/9/9 | 254.00 | normal | |||
| 84 | GWFG03·BFUL012 | 2015/12/11 | 2018/5/15 | 886.00 | normal | migrating(Boundary) | ||
| 85 | GWFG04·BYH021 | 2014/12/30 | 2015/5/16 | 137.00 | normal | |||
| 86 | GWFG05·BYH027 | 2014/12/29 | 2016/8/9 | 589.00 | normal | |||
| 87 | GWFG06·BFUL094 | 2017/1/18 | 2017/6/22 | 155.00 | normal | migrating(Boundary) | ||
| 88 | GWFG07·BFUL102 | 2017/2/14 | 2017/5/19 | 94.00 | Lost | migrating(China) | ||
| 89 | GWFG08·BFUL105 | 2017/2/14 | 2017/7/22 | 158.00 | Lost | migrating(Boundary) | ||
| 90 | GWFG09·BFUL111 | 2017/2/18 | 2017/2/20 | 2.00 | Lost | wintering | ||
| 91 | GWFG10·BFUL153 | 2017/4/14 | 2017/6/16 | 63.00 | Lost | migrating(Boundary) | ||
| 92 | GWFG11·BFUL081 | 2016/12/26 | 2018/4/5 | 465.00 | Lost | migrating(China) | ||
| 93 | GWFG12·CAS008 | 2014/12/29 | 2015/4/9 | 101.00 | Lost | migrating(China) | ||
| 94 | GWFG13·BYH021 | 2014/12/29 | 2015/4/12 | 104.00 | Lost | migrating(China) | ||
| 95 | GWFG14·BFUL093 | 2017/1/18 | 2017/7/9 | 172.00 | Lost | migrating(Boundary) | ||
| 96 | GWFG15·BYH011 | 2014/12/29 | 2015/3/23 | 84.00 | Lost | wintering | ||
| 97 | GWFG16·BYH018 | 2014/12/30 | 2015/5/9 | 130.00 | Lost | migrating(China) | ||
| 98 | GWFG17·BYH019 | 2014/12/30 | 2015/4/10 | 101.00 | Lost | migrating(China) | ||
| 99 | GWFG18·BYH009 | 2014/12/29 | 2015/4/9 | 101.00 | Lost | migrating(China) | ||
| 100 | GWFG19·BYH009 | 2014/12/29 | 2015/1/9 | 11.00 | Lost | wintering | ||
| 101 | GWFG20·BYH017 | 2014/12/29 | 2015/3/2 | 63.00 | Lost | wintering | ||
| 102 | BG01·BYH025 | 2015/3/25 | 2015/3/28 | 3.00 | Lost | wintering | ||
| 103 | BG02·BYH026 | 2015/3/25 | 2015/5/11 | 47.00 | Lost | migrating(China) | ||
| 104 | BG03·CAS025 | 2015/1/26 | 2015/6/24 | 149.00 | Lost | migrating(Boundary) | ||
| 105 | BG04·4B04E8 | 2014/3/8 | 2014/3/15 | 7.00 | Lost | wintering | ||
| 106 | BG005·BFUL037 | 2016/1/4 | 2016/12/22 | 353.00 | normal | wintering | ||
| 107 | BG007·BFUL048 | 2016/11/22 | 2016/12/24 | 32.00 | Lost | wintering | ||
| 108 | BG008·BFUL23 | 2016/12/3 | 2017/3/6 | 93.00 | death | Yueyang Wintering site | ||
| 109 | BG009·BFUL27 | 2016/12/4 | 2017/4/21 | 138.00 | normal | |||
| 110 | BG010·BFUL047 | 2016/12/16 | 2017/5/5 | 140.00 | Lost | migrating(Boundary) | ||
| 111 | BG011·BFUL069 | 2016/12/26 | 2017/3/14 | 78.00 | Lost | wintering | ||
| 112 | BG012·BFUL077 | 2016/12/26 | 2017/6/7 | 163.00 | death | Hubei Probince | ||
| 113 | BG013·BFUL080 | 2016/12/26 | 2017/6/10 | 166.00 | Lost | migrating(Boundary) | ||
| 114 | BG014·BFUL078 | 2016/12/26 | 2016/12/26 | 0.00 | Lost | wintering | ||
| 115 | BG015·BFUL076 | 2016/12/26 | 2017/5/29 | 154.00 | Lost | migrating(Boundary) | ||
| 116 | BG016·BFUL075 | 2016/12/26 | 2017/12/24 | 363.00 | normal | wintering | ||
| 117 | BG017·BFUL079 | 2016/12/26 | 2017/5/10 | 135.00 | Lost | migrating(Boundary) | ||
| 118 | BG018·BFUL099 | 2017/1/18 | 2017/5/10 | 112.00 | Lost | migrating(China) | ||
| 119 | BG019·BFUL098 | 2017/2/14 | 2017/2/26 | 12.00 | Lost | wintering | ||
| 120 | BG020·BFUL100 | 2017/2/14 | 2017/3/27 | 41.00 | Lost | migrating(China) | ||
| 121 | BG021·BFUL103 | 2017/2/14 | 2017/5/14 | 89.00 | Lost | migrating(Boundary) | ||
| 122 | BG022·BFUL104 | 2017/2/14 | 2017/5/26 | 101.00 | normal | |||
| 123 | BG023·BFUL106 | 2017/2/14 | 2017/5/8 | 83.00 | Lost | migrating(Boundary) | ||
| 124 | BG024·BFUL108 | 2017/2/20 | 2017/2/25 | 5.00 | Lost | wintering | ||
| 125 | BG025·BFUL109 | 2017/2/20 | 2017/2/25 | 5.00 | Lost | wintering | ||
| 126 | BG026·BFUL110 | 2017/2/20 | 2017/2/20 | 0.00 | Lost | wintering | ||
| 127 | BG027·4AD550 | 2014/3/8 | 2014/3/16 | 8.00 | Lost | wintering | ||
| 128 | BG028·4AD553 | 2014/3/8 | 2014/4/28 | 51.00 | Lost | migrating(China) | ||
| 129 | BG029· 3864AEFE1 | 2014/2/24 | 2014/3/11 | 15.00 | Lost | wintering | ||
| 130 | BG030·3864B0378 | 2014/2/24 | 2014/3/27 | 31.00 | Lost | wintering | ||
| 131 | BG031·38D475FFE | 2015/9/8 | 2016/4/7 | 212.00 | Lost | migrating(China) | ||
| 132 | BG032·CAS001 | 2014/12/22 | 2014/12/23 | 1.00 | Lost | wintering | ||
| 133 | SG01·BFUL033 | 2016/1/4 | 2016/4/3 | 90.00 | Lost | migrating(China) | ||
| 134 | SG02·BFUL034 | 2016/1/4 | 2016/2/18 | 45.00 | Lost | wintering | ||
| 135 | SG003·BFUL004 | 2016/12/9 | 2017/3/10 | 91.00 | Lost | migrating(China) | ||
| 136 | SG004·BFUL045 | 2016/12/9 | 2018/11/13 | 704.00 | normal | |||
| 137 | SG005·BFUL101 | 2017/1/18 | 2018/1/30 | 377.00 | normal | |||
| 138 | SG006·BFUL107 | 2017/1/18 | 2017/3/25 | 66.00 | Lost | migrating(China) | ||
| 139 | SG007·BFUL146 | 2017/3/14 | 2017/4/16 | 33.00 | Lost | migrating(China) | ||
| 140 | SG008·BFUL147 | 2017/3/14 | 2017/5/20 | 67.00 | Lost | migrating(China) | ||
| 141 | SG009·CAS006 | 2014/7/21 | 2015/5/9 | 292.00 | Lost | migrating(China) |
| Route ID | No. of Stopovers | Total Days | Total Area (ha) | Percentage of Samples |
|---|---|---|---|---|
| BG016.17 | 3 | 48 | 74,607 | 92.63 |
| BG05.16 | 3 | 48 | 166,893 | 94.03 |
| BG09.17 | 4 | 56 | 425,560 | 90.57 |
| BG22.17 | 2 | 13 | 144,181 | 60.41 |
| Greylag02.15 | 1 | 22 | 144,490 | 71.22 |
| Greylag02.16 | 2 | 45 | 101,355 | 96.44 |
| Greylag02.17 | 2 | 58 | 27,975 | 98.25 |
| Greylag04.15 | 2 | 38 | 28,564 | 91.80 |
| Greylag04.16 | 1 | 41 | 176,753 | 92.48 |
| GWFG01.15 | 1 | 59 | 850,934 | 91.75 |
| GWFG01.16 | 3 | 51 | 409,786 | 91.82 |
| GWFG02.15 | 3 | 33 | 15,366 | 91.95 |
| GWFG03.16 | 3 | 51 | 592,593 | 92.27 |
| GWFG03.17 | 3 | 43 | 162,725 | 91.59 |
| GWFG04.15 | 1 | 15 | 5233 | 82.91 |
| GWFG05.15 | 3 | 52 | 990,029 | 93.18 |
| GWFG05.16 | 4 | 51 | 501,513 | 93.11 |
| GWFG05.17 | 5 | 42 | 572,606 | 91.87 |
| GWFG06.17 | 6 | 65 | 982,452 | 91.51 |
| LWFG12.16 | 7 | 65 | 648,292 | 87.12 |
| LWFG12.17 | 8 | 60 | 733,961 | 88.93 |
| LWFG14.16 | 2 | 50 | 478,995 | 92.87 |
| LWFG14.17 | 4 | 55 | 1,026,059 | 85.35 |
| LWFG28.17 | 4 | 56 | 485,033 | 88.54 |
| LWFG29.17 | 3 | 42 | 630,021 | 84.03 |
| LWFG38.17 | 8 | 70 | 120,140 | 80.28 |
| LWFG40.17 | 4 | 76 | 520,131 | 84.42 |
| LWFG42.17 | 5 | 82 | 674,976 | 89.82 |
| LWFG45.17 | 3 | 43 | 291,943 | 88.76 |
| LWFG47.17 | 6 | 77 | 308,214 | 91.94 |
| LWFG62.17 | 3 | 44 | 172,960 | 84.07 |
| SG04.17 | 1 | 30 | 302,917 | 87.10 |
| SG05.17 | 2 | 29 | 28,426 | 66.67 |
| Average | 3.39 | 48.79 | 387,748 | 87.87 |
| Standard deviation | 1.92 | 16.40 | 308,684 | 8.12 |

References
- Runge, C.A.; Watson, J.E.; Butchart, S.H.; Hanson, J.O.; Possingham, H.P.; Fuller, R.A. Protected areas and global conservation of migratory birds. Science 2015, 350, 1255–1258. [Google Scholar] [CrossRef] [PubMed]
- Bauer, S.; Hoye, B.J. Migratory animals couple biodiversity and ecosystem functioning worldwide. Science 2014, 344, 1242552. [Google Scholar] [CrossRef] [PubMed]
- Wilcove, D.S.; Wikelski, M. Going, going, gone: Is animal migration disappearing. PLoS Biol. 2008, 6, e188. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zeng, G.; Liang, J.; Chen, J.; Xu, J.; Dai, J.; Sang, L.; Li, X.; Ye, S. Responses of landscape pattern of China’s two largest freshwater lakes to early dry season after the impoundment of Three-Gorges Dam. Int. J. Appl. Earth Obs. Geoinf. 2017, 56, 36–43. [Google Scholar] [CrossRef]
- Klaassen, R.H.; Hake, M.; Strandberg, R.; Koks, B.J.; Trierweiler, C.; Exo, K.M.; Bairlein, F.; Alerstam, T. When and where does mortality occur in migratory birds? Direct evidence from long-term satellite tracking of raptors. J. Anim. Ecol. 2014, 83, 176–184. [Google Scholar] [CrossRef]
- Yang, H.; Chen, B.; Barter, M.; Piersma, T.; Zhou, C.; Li, F.; Zhang, Z. Impacts of tidal land reclamation in Bohai Bay, China: Ongoing losses of critical Yellow Sea waterbird staging and wintering sites. Bird Conserv. Int. 2011, 21, 241–259. [Google Scholar] [CrossRef]
- Møller, A.P.; Rubolini, D.; Lehikoinen, E. Populations of migratory bird species that did not show a phenological response to climate change are declining. Proc. Natl. Acad. Sci. USA 2008, 105, 16195–16200. [Google Scholar] [CrossRef]
- Crick, H.Q. The impact of climate change on birds. IBIS 2004, 146, 48–56. [Google Scholar] [CrossRef]
- Taylor, C.M.; Stutchbury, B.J. Effects of breeding versus winter habitat loss and fragmentation on the population dynamics of a migratory songbird. Ecol. Appl. 2016, 26, 424–437. [Google Scholar] [CrossRef]
- Kamp, J.; Oppel, S.; Ananin, A.A.; Durnev, Y.A.; Gashev, S.N.; Hölzel, N.; Mishchenko, A.L.; Pessa, J.; Smirenski, S.M.; Strelnikov, E.G. Global population collapse in a superabundant migratory bird and illegal trapping in China. Conserv. Biol. 2015, 29, 1684–1694. [Google Scholar] [CrossRef]
- Rocke, T.E. The global importance of avian botulism. In Waterbirds around the World; The Stationery Office: Edinburgh, UK, 2006; pp. 422–426. [Google Scholar]
- Bairlein, F. Migratory birds under threat. Science 2016, 354, 547–548. [Google Scholar] [CrossRef]
- Runge, C.A.; Martin, T.G.; Possingham, H.P.; Willis, S.G.; Fuller, R.A. Conserving mobile species. Front. Ecol. Environ. 2014, 12, 395–402. [Google Scholar] [CrossRef]
- Sawyer, H.; Kauffman, M.J. Stopover ecology of a migratory ungulate. J. Anim. Ecol. 2011, 80, 1078–1087. [Google Scholar] [CrossRef] [PubMed]
- Battley, P.F.; Warnock, N.; Tibbitts, T.L.; Gill, R.E.; Piersma, T.; Hassell, C.J.; Douglas, D.C.; Mulcahy, D.M.; Gartrell, B.D.; Schuckard, R. Contrasting extreme long-distance migration patterns in bar-tailed godwits Limosa lapponica. J. Avian Biol. 2012, 43, 21–32. [Google Scholar] [CrossRef]
- Sheehy, J.; Taylor, C.M.; Norris, D.R. The importance of stopover habitat for developing effective conservation strategies for migratory animals. J. Ornithol. 2011, 152, 161–168. [Google Scholar] [CrossRef]
- Warnock, N. Stopping vs. staging: The difference between a hop and a jump. J. Avian Biol. 2010, 41, 621–626. [Google Scholar] [CrossRef]
- Schaub, M.; Jenni, L.; Bairlein, F. Fuel stores, fuel accumulation, and the decision to depart from a migration stopover site. Behav. Ecol. 2008, 19, 657–666. [Google Scholar] [CrossRef]
- Trinder, M.N.; Hassell, D.; Votier, S. Reproductive performance in arctic-nesting geese is influenced by environmental conditions during the wintering, breeding and migration seasons. OIKOS 2009, 118, 1093–1101. [Google Scholar] [CrossRef]
- Newton, I. Can conditions experienced during migration limit the population levels of birds? J. Ornithol. 2006, 147, 146–166. [Google Scholar] [CrossRef]
- Halupka, L.; Wierucka, K.; Sztwiertnia, H.; Klimczuk, E. Conditions at autumn stopover sites affect survival of a migratory passerine. J. Ornithol. 2017, 158, 979–988. [Google Scholar] [CrossRef]
- McCabe, J.D.; Olsen, B.J. Landscape-scale habitat availability, and not local geography, predicts migratory landbird stopover across the Gulf of Maine. J. Avian Biol. 2015, 46, 395–405. [Google Scholar] [CrossRef]
- La Sorte, F.A.; Tingley, M.W.; Hurlbert, A.H. The role of urban and agricultural areas during avian migration: An assessment of within-year temporal turnover. Glob. Ecol. Biogeogr. 2014, 23, 1225–1234. [Google Scholar] [CrossRef]
- Warnock, N.; Bishop, M.A. Spring stopover ecology of migrant Western Sandpipers. Condor 1998, 100, 456–467. [Google Scholar] [CrossRef]
- Piersma, T.; Lok, T.; Chen, Y.; Hassell, C.J.; Yang, H.Y.; Boyle, A.; Slaymaker, M.; Chan, Y.C.; Melville, D.S.; Zhang, Z.W. Simultaneous declines in summer survival of three shorebird species signals a flyway at risk. J. Appl. Ecol. 2016, 53, 479–490. [Google Scholar] [CrossRef]
- Jia, Y.; Zhang, Y.; Lei, J.; Jiao, S.; Lei, G.; Yu, X.; Liu, G. Activity Patterns of four Cranes in Poyang Lake, China: Indication of Habitat Naturalness. Wetlands 2017, 1–9. [Google Scholar] [CrossRef]
- Fox, A.D.; Lei, C.; Barter, M.; Rees, E.C.; Hearn, R.D.; Hao, C.P.; Xin, W.; Yong, Z.; Tao, D.S.; Fang, S.X. The functional use of East Dongting Lake, China, by wintering geese. Wildfowl 2013, 58, 3–19. [Google Scholar]
- Webster, M.S.; Marra, P.P.; Haig, S.M.; Bensch, S.; Holmes, R.T. Links between worlds: Unraveling migratory connectivity. Trends Ecol. Evol. 2002, 17, 76–83. [Google Scholar] [CrossRef]
- Harris, J.; Liying, S.; Higuchi, H.; Ueta, M.; Zhengwang, Z.; Yanyun, Z.; Xijun, N. Migratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G. monacha, as determined by satellite tracking. Forktail 2000, 16, 93–100. [Google Scholar]
- Marra, P.P.; Cohen, E.B.; Loss, S.R.; Rutter, J.E.; Tonra, C.M. A call for full annual cycle research in animal ecology. Biol. Lett. 2015, 11, 20150552. [Google Scholar] [CrossRef]
- Bridge, E.S.; Kelly, J.F.; Contina, A.; Gabrielson, R.M.; MacCurdy, R.B.; Winkler, D.W. Advances in tracking small migratory birds: A technical review of light-level geolocation. J. Field Ornithol. 2013, 84, 121–137. [Google Scholar] [CrossRef]
- Wink, M. Use of DNA markers to study bird migration. J. Ornithol. 2006, 147, 234–244. [Google Scholar] [CrossRef]
- Rubenstein, D.R.; Hobson, K.A. From birds to butterflies: Animal movement patterns and stable isotopes. Trends Ecol. Evol. 2004, 19, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Trierweiler, C.; Klaassen, R.H.; Drent, R.H.; Exo, K.; Komdeur, J.; Bairlein, F.; Koks, B.J. Migratory connectivity and population-specific migration routes in a long-distance migratory bird. Proc. R. Soc. Lond. B 2014, 281, 20132897. [Google Scholar] [CrossRef]
- Worton, B.J. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 1989, 70, 164–168. [Google Scholar] [CrossRef]
- Benhamou, S. Dynamic approach to space and habitat use based on biased random bridges. PLoS ONE 2011, 6, e14592. [Google Scholar] [CrossRef] [PubMed]
- Delany, S.; Scott, D. Waterbird Population Estimates, 4th ed.; Wetlands International: Wageningen, The Netherlands, 2006; 239p. [Google Scholar]
- Cao, L.; Barter, M.; Lei, G. New Anatidae population estimates for eastern China: Implications for current flyway estimates. Biol. Conserv. 2008, 141, 2301–2309. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, L.; Barter, M.; Fox, A.D.; Zhao, M.; Meng, F.; Shi, H.; Jiang, Y.; Zhu, W. Changing distribution and abundance of Swan Goose Anser cygnoides in the Yangtze River floodplain: The likely loss of a very important wintering site. Bird Conserv. Int. 2011, 21, 36–48. [Google Scholar] [CrossRef]
- Guan, L.; Wen, L.; Feng, D.; Zhang, H.; Lei, G. Delayed flood recession in central Yangtze floodplains can cause significant food shortages for wintering geese: Results of inundation experiment. Environ. Manag. 2014, 54, 1331–1341. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, Z.; Song, K.; Zhang, B.; Zhang, S.; Liu, D.; Ren, C.; Yang, F. Land Use Changes in Northeast China Driven by Human Activities and Climatic Variation. Chinese Geogr. Sci. 2009, 19, 225–230. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, D.; Gao, J.; Deng, W. Land Use/Cover Changes, the Environment and Water Resources in Northeast China. Environ. Manage. 2005, 36, 691. [Google Scholar] [CrossRef] [PubMed]
- Drent, R.H.; Eichhorn, G.; Flagstad, A.; Van der Graaf, A.J.; Litvin, K.E.; Stahl, J. Migratory connectivity in Arctic geese: Spring stopovers are the weak links in meeting targets for breeding. J. Ornithol. 2007, 148, 501–514. [Google Scholar] [CrossRef]
- Wang, X.; Fox, A.D.; Cong, P.; Barter, M.; Cao, L. Changes in the distribution and abundance of wintering Lesser White-fronted Geese Anser erythropus in eastern China. Bird Conserv. Int. 2012, 22, 128–134. [Google Scholar] [CrossRef]
- Gang, L. Status of Lesser White-fronted Goose in China. In Fennoscandian Lesser White-Fronted Goose Conservation Project—Annual Report 1999; Bridlife: Cambridge, UK, 1999. [Google Scholar]
- Kenward, R.E. A Manual for Wildlife Radio Tagging; Academic Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Kölzsch, A.; Neefjes, M.; Barkway, J.; Müskens, G.J.; Langevelde, F.; Boer, W.F.; Prins, H.H.; Cresswell, B.H.; Nolet, B.A. Neckband or backpack? Differences in tag design and their effects on GPS/accelerometer tracking results in large waterbirds. Anim. Biotelem. 2016, 4, 13. [Google Scholar] [CrossRef]
- Tomkiewicz, S.M.; Fuller, M.R.; Kie, J.G.; Bates, K.K. Global positioning system and associated technologies in animal behaviour and ecological research. Philos. Trans. R. Soc. B 2010, 365, 2163–2176. [Google Scholar] [CrossRef] [PubMed]
- Horne, J.S.; Garton, E.O.; Krone, S.M.; Lewis, J.S. Analyzing animal movements using Brownian bridges. Ecology 2007, 88, 2354–2363. [Google Scholar] [CrossRef] [PubMed]
- Benhamou, S.; Cornélis, D. Incorporating movement behavior and barriers to improve kernel home range space use estimates. J. Wildlife Manag. 2010, 74, 1353–1360. [Google Scholar] [CrossRef]
- Calenge, C. Home Range Estimation in R: The Adehabitathr Package; Office National de la Classe et de la Faune Sauvage: Auffargis, France, 2011.
- Monsarrat, S.; Benhamou, S.; Sarrazin, F.; Bessa-Gomes, C.; Bouten, W.; Duriez, O. How predictability of feeding patches affects home range and foraging habitat selection in avian social scavengers? PLoS ONE 2013, 8, e53077. [Google Scholar] [CrossRef] [PubMed]
- Jenks, G.F. The data model concept in statistical mapping. In International Yearbook of Cartography; George Philip: London, UK, 1967; Volume 7, pp. 186–190. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Available online: http://www.iucnredlist.org (accessed on 5 July 2018).
- Arzel, C.; Elmberg, J.; Guillemain, M. Ecology of spring-migrating Anatidae: A review. J. Ornithol. 2006, 147, 167–184. [Google Scholar] [CrossRef]
- Wang, X.; Fox, A.D.; Cong, P.; Cao, L. Food constraints explain the restricted distribution of wintering Lesser White-fronted Geese Anser erythropus in China. IBIS 2013, 155, 576–592. [Google Scholar] [CrossRef]
- Fox, A.D.; Madsen, J.; Boyd, H.; Kuijken, E.; Norriss, D.W.; Tombre, I.M.; Stroud, D.A. Effects of agricultural change on abundance, fitness components and distribution of two arctic-nesting goose populations. Glob. Chang. Biol. 2005, 11, 881–893. [Google Scholar] [CrossRef]
- Jefferies, R.L.; Rockwell, R.F.; Abraham, K.F. Agricultural Food Subsidies, Migratory Connectivity and Large-Scale Disturbance in Arctic Coastal Systems: A Case Study1. Integr. Comp. Biol. 2004, 44, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Fox, A.D.; Abraham, K.F. Why geese benefit from the transition from natural vegetation to agriculture. Ambio 2017, 46, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Tinkler, E.; Montgomery, W.I.; Elwood, R.W. Foraging ecology, fluctuating food availability and energetics of wintering brent geese. J. Zool. 2009, 278, 313–323. [Google Scholar] [CrossRef]
- Koffijberg, K.; Schekkerman, H.; van der Jeugd, H.; Hornman, M.; van Winden, E. Responses of wintering geese to the designation of goose foraging areas in The Netherlands. Ambio 2017, 46, 241–250. [Google Scholar] [CrossRef]
- Wang, Z.; Song, K.; Ma, W.; Ren, C.; Zhang, B.; Liu, D.; Chen, J.M.; Song, C. Loss and fragmentation of marshes in the Sanjiang Plain, Northeast China, 1954–2005. Wetlands 2011, 31, 945. [Google Scholar] [CrossRef]
- Zhang, S.; Na, X.; Kong, B.; Wang, Z.; Jiang, H.; Yu, H.; Zhao, Z.; Li, X.; Liu, C.; Dale, P. Identifying wetland change in China’s Sanjiang Plain using remote sensing. Wetlands 2009, 29, 302–313. [Google Scholar] [CrossRef]
- Li, Z.; Tang, H.; Yang, P.; Wu, W.; Chen, Z.; Zhou, Q.; Zhang, L.; Zou, J. Spatio-temporal responses of cropland phenophases to climate change in Northeast China. J. Geogr. Sci. 2012, 22, 29–45. [Google Scholar] [CrossRef]
- Riddington, R.; Hassall, M.; Lane, S.J.; Turner, P.A.; Walters, R. The impact of disturbance on the behaviour and energy budgets of Brent Geese Branta b. bernicla. Bird Study 1996, 43, 269–279. [Google Scholar] [CrossRef]
- Owen, M.; Black, J.M. Factors affecting the survival of Barnacle Geese on migration from the breeding grounds. J. Anim. Ecol. 1989, 58, 603–617. [Google Scholar] [CrossRef]
- Liu, H.; Cai, X.; Geng, L.; Zhong, H. Restoration of pastureland ecosystems: Case study of Western Inner Mongolia. J. Water Resour. Plan. Mannag. 2005, 131, 420–430. [Google Scholar] [CrossRef]
- Luo, J.; Wang, Y.; Yang, F.; Liu, Z. Effects of human disturbance on the Hooded Crane (Grus monacha) at stopover sites in northeastern China. Chin. Birds 2012, 3, 206–216. [Google Scholar] [CrossRef]
- Li, D.; Chen, S.; Guan, L.; Lloyd, H.; Liu, Y.; Lv, J.; Zhang, Z. Patterns of waterbird community composition across a natural and restored wetland landscape mosaic, Yellow River Delta, China. Estuar. Coast. Shelf Sci. 2011, 91, 325–332. [Google Scholar] [CrossRef]
- Chudzińska, M.E.; Nabe Nielsen, J.; Nolet, B.A.; Madsen, J. Foraging behaviour and fuel accumulation of capital breeders during spring migration as derived from a combination of satellite-and ground-based observations. J. Avian Biol. 2016, 47, 563–574. [Google Scholar] [CrossRef]
- Kirby, J.S.; Stattersfield, A.J.; Butchart, S.H.; Evans, M.I.; Grimmett, R.F.; Jones, V.R.; O’Sullivan, J.; Tucker, G.M.; Newton, I. Key conservation issues for migratory land-and waterbird species on the world’s major flyways. Bird Conserv. Int. 2008, 18, S49–S73. [Google Scholar] [CrossRef]
- Kölzsch, A.; Müskens, G.J.; Kruckenberg, H.; Glazov, P.; Weinzierl, R.; Nolet, B.A.; Wikelski, M. Towards a new understanding of migration timing: Slower spring than autumn migration in geese reflects different decision rules for stopover use and departure. OIKOS 2016, 125, 1496–1507. [Google Scholar] [CrossRef]
- Pereira, H.M.; Ferrier, S.; Walters, M.; Geller, G.N.; Jongman, R.; Scholes, R.J.; Bruford, M.W.; Brummitt, N.; Butchart, S.; Cardoso, A.C. Essential biodiversity variables. Science 2013, 339, 277–278. [Google Scholar] [CrossRef]


| Passage | No. of Sites | Total Area (ha) | Mean Area (ha) | Species | Usage in 2017 (days) * |
|---|---|---|---|---|---|
| Southern | 32 | 462,254 | 14,445 | All | 142 |
| Northeast | 31 | 1,502,998 | 48,483 | GWFG, LWFG, BG | 649 |
| Northwest | 47 | 210,409 | 4476 | GreyLag, SG, BG | 191 |
| Total | 110 | 2,175,661 | 67,404 | All | 982 |
| Passage | A | B | C | D | E | F |
|---|---|---|---|---|---|---|
| Southern | 56.68 | 26.23 | 13.97 | 0.43 | 0.69 | 1.98 |
| Northeast | 72.00 | 14.83 | 9.05 | 2.32 | 0.48 | 1.32 |
| Northwest | 24.57 | 9.24 | 62.94 | 0.86 | 2.15 | 0.23 |
| Migration path average | 62.94 | 17.86 | 15.48 | 1.60 | 0.70 | 1.43 |
| No of Stopover Sites | Protected Area (ha) | Percentage (%) | |
|---|---|---|---|
| I | 1 | 3030 | 0.14 |
| II | 6 | 183,773 | 8.38 |
| III | 14 | 155,953 | 7.11 |
| IV | 89 | 1,849,641 | 84.37 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, J.; Jia, Y.; Zuo, A.; Zeng, Q.; Shi, L.; Zhou, Y.; Zhang, H.; Lu, C.; Lei, G.; Wen, L. Bird Satellite Tracking Revealed Critical Protection Gaps in East Asian–Australasian Flyway. Int. J. Environ. Res. Public Health 2019, 16, 1147. https://doi.org/10.3390/ijerph16071147
Lei J, Jia Y, Zuo A, Zeng Q, Shi L, Zhou Y, Zhang H, Lu C, Lei G, Wen L. Bird Satellite Tracking Revealed Critical Protection Gaps in East Asian–Australasian Flyway. International Journal of Environmental Research and Public Health. 2019; 16(7):1147. https://doi.org/10.3390/ijerph16071147
Chicago/Turabian StyleLei, Jialin, Yifei Jia, Aojie Zuo, Qing Zeng, Linlu Shi, Yan Zhou, Hong Zhang, Cai Lu, Guangchun Lei, and Li Wen. 2019. "Bird Satellite Tracking Revealed Critical Protection Gaps in East Asian–Australasian Flyway" International Journal of Environmental Research and Public Health 16, no. 7: 1147. https://doi.org/10.3390/ijerph16071147
APA StyleLei, J., Jia, Y., Zuo, A., Zeng, Q., Shi, L., Zhou, Y., Zhang, H., Lu, C., Lei, G., & Wen, L. (2019). Bird Satellite Tracking Revealed Critical Protection Gaps in East Asian–Australasian Flyway. International Journal of Environmental Research and Public Health, 16(7), 1147. https://doi.org/10.3390/ijerph16071147