
Genetic Aspects of Susceptibility to Mercury Toxicity: An Overview

Abstract
:1. Introduction
2. Mechanisms of Toxicity
3. Health Effects and Clinical Presentation
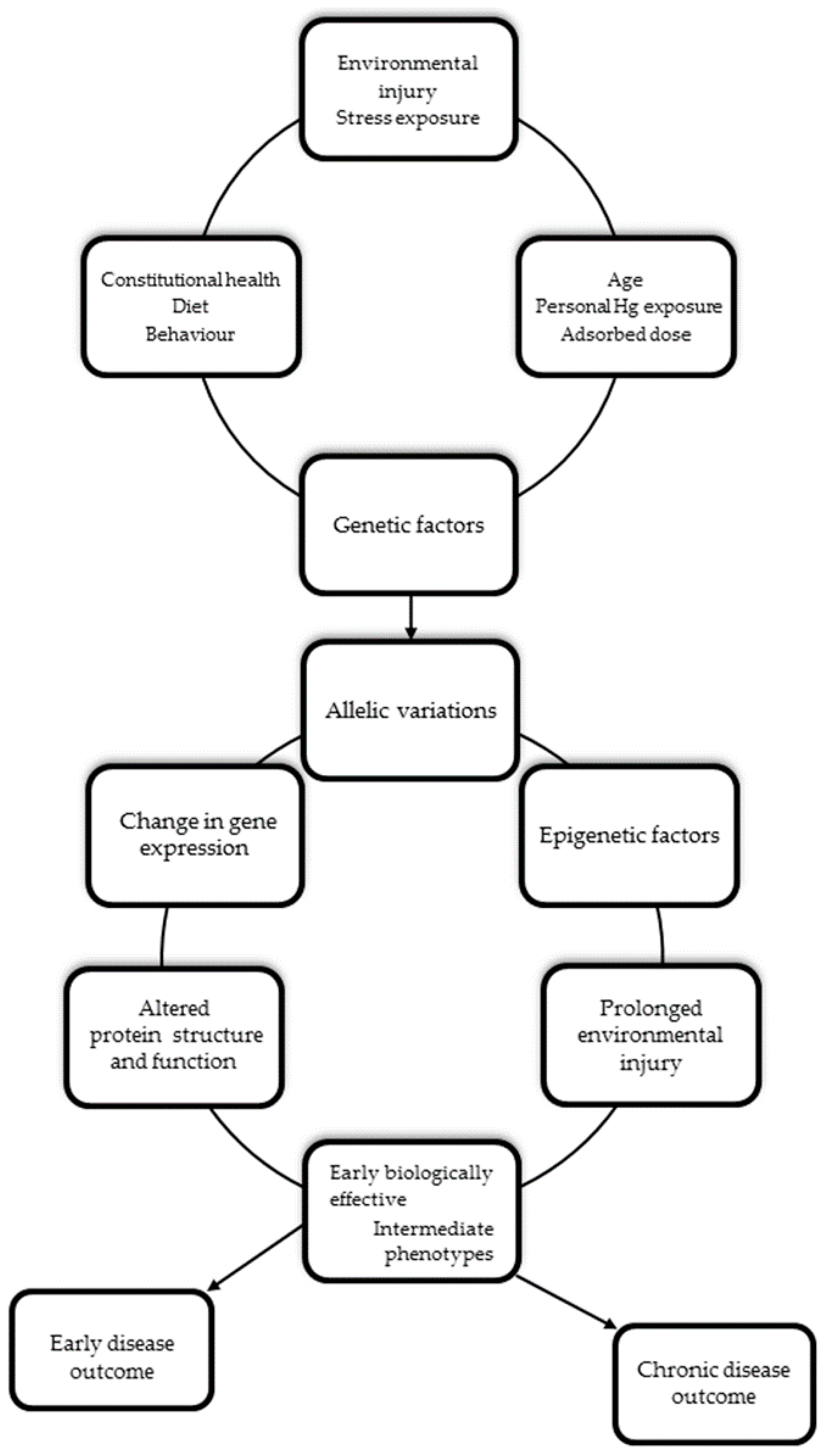
4. The Role of the Genetic Background in Mediating Individual Mercury Susceptibility
5. Search Strategy and Study Selection
6. Gene Classes of Interest and Related Polymorphisms
6.1. Inter-Individual Variability and Mercury Body Burden
6.1.1. The Glutathione System
6.1.2. The Metallothioneins Superfamily
6.1.3. The Selenoproteins Family
6.1.4. The Xenobiotic Transporter Proteins Superfamilies
6.2. Genetic Susceptibility and Neurodevelopmental Mercury Outcomes
6.3. Genetic Susceptibility and Cardiovascular Mercury Outcomes
7. Discussion
- Understanding of the underlying biological Hg mechanisms;
- Clarification of the variability and critical windows of susceptibility in the development of Hg-related health outcomes;
- Implementation potential preventive measures, and treatment;
- Improved Hg risk assessments and decision-making;
- Reduced global health disparities;
- Enhanced quality of life of all people.
8. Conclusions
Author Contributions
Conflicts of Interest
References
- United Nations Environment Programme. Global Mercury Assessment; United Nations Environment Programme, UNEP Chemicals: Geneva, Switzerland, 2002. [Google Scholar]
- Guidelines for Identifying Populations at Risk from Mercury Exposure. Available online: http://www.who.int/foodsafety/publications/risk-mercury-exposure/en/ (accessed on 22 August 2008).
- United Nations Environment Programme. Global Mercury Assessment 2013: Sources, Emissions, Releases and Environmental Transport; UNEP Chemicals Branch: Geneva, Switzerland, 2013. [Google Scholar]
- Poulin, J.; Gibb, H. Mercury: Assessing the Burden of Disease at National and Local Levels; World Health Organization: Geneva, Switzerland, 2008. [Google Scholar]
- Engström, K.; Ameer, S.; Bernaudat, L.; Drasch, G.; Baeuml, J.; Skerfving, S.; Bose-O’Reilly, S.; Broberg, K. Polymorphisms in genes encoding potential mercury transporters and urine mercury concentrations in populations exposed to mercury vapor from gold mining. Environ. Health Perspect. 2013, 121, 85–91. [Google Scholar] [PubMed]
- Goodrich, J.M.; Wang, Y.; Gillespie, B.; Werner, R.; Franzblau, A.; Basu, N. Glutathione enzyme and selenoprotein polymorphisms associate with mercury biomarker levels in Michigan dental professionals. Toxicol. Appl. Pharmacol. 2011, 25, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Harari, R.; Harari, F.; Gerhardsson, L.; Lundh, T.; Skerfving, S.; Strömberg, U.; Broberg, K. Exposure and toxic effects of elemental mercury in gold-mining activities in Ecuador. Toxicol. Lett. 2012, 21, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Goodrich, J.M.; Werner, R.; Gillespie, B.; Basu, N.; Franzblau, A. An investigation of modifying effects of single nucleotide polymorphisms in metabolism-related genes on the relationship between peripheral nerve function and mercury levels in urine and hair. Sci. Total Environ. 2012, 417–418, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Barcelos, G.R.; Grotto, D.; de Marco, K.C.; Valentini, J.; Lengert, A.V.; de Oliveira, A.Á.; Garcia, S.C.; Braga, G.Ú.; Schläwicke Engström, K.; Cólus, I.M.; et al. Polymorphisms in glutathione-related genes modify mercury concentrations and antioxidant status in subjects environmentally exposed to methylmercury. Sci. Total Environ. 2013, 463–464, 319–325. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.Á.; de Souza, M.F.; Lengert, A.V.; de Oliveira, M.T.; Camargo, R.B.; Braga, G.Ú.; Cólus, I.M.; Barbosa, F., Jr.; Barcelos, G.R. Genetic polymorphisms in glutathione (GSH-) related genes affect the plasmatic mercury/whole blood mercury partitioning and the distribution between inorganic and methylmercury levels in plasma collected from a fish-eating population. Biomed. Res. Int. 2014. [Google Scholar] [CrossRef] [PubMed]
- Basu, N.; Goodrich, J.M.; Head, J. Ecogenetics of mercury: From genetic polymorphisms and epigenetics to risk assessment and decision making. Environ. Toxicol. Chem. 2014, 33, 1248–1258. [Google Scholar] [CrossRef] [PubMed]
- Gundacker, C.; Gencik, M.; Hengstschläger, M. The relevance of the individual genetic background for the toxicokinetics of two significant neurodevelopmental toxicants: Mercury and lead. Mutat. Res. 2010, 705, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Rice, K.M.; Walker, E.M., Jr.; Wu, M.; Gillette, C.; Blough, E.R. Environmental mercury and its toxic effects. J. Prev. Med. Public Health 2014, 47, 74–83. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Elemental Mercury and Inorganic Mercury Compounds: Human Health Aspects. Concise International Chemical Assessment Document 50; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- World Health Organization. Methylmercury. Environmental Health Criteria 101; World Health Organization: Geneva, Switzerland, 1990. [Google Scholar]
- World Health Organization. Exposure to Mercury: A Major Public Health Concern; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Clarkson, T.W.; Magos, L.; Myers, G.J. The toxicology of mercury—Current exposures and clinical manifestations. N. Engl. J. Med. 2003, 349, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Syversen, T.; Kaur, P. The toxicology of mercury and its compounds. J. Trace Elem. Med. Biol. 2012, 26, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Ballatori, N.; Clarkson, T.W. Biliary secretion of glutathione and of glutathione-metal complexes. Fundam. Appl. Toxicol. 1985, 5, 816–831. [Google Scholar] [CrossRef]
- Björkman, L.; Lundekvam, B.F.; Lægreid, T.; Bertelsen, B.I.; Morild, I.; Lilleng, P.; Lind, B.; Palm, B.; Vahter, M. Mercury in human brain, blood, muscle and nails in relation to exposure: An autopsy study. Environ. Health 2007, 6, 30. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.M.; Choi, A.L.; Ha, E.H.; Pedersen, L.; Nielsen, F.; Weihe, P.; Hong, Y.C.; Budtz-Jørgensen, E.; Grandjean, P. Effect of hemoglobin adjustment on the precision of mercury concentrations in maternal and cord blood. Environ. Res. 2014, 132, 407–412. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Children’s Exposure to Mercury Compounds; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- National Research Council. Toxicological Effects of Methylmercury; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- Clarkson, T.W. The three modern faces of mercury. Environ. Health Perspect. 2002, 110, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Houston, M.C. Role of mercury toxicity in hypertension, cardiovascular disease, and stroke. J. Clin. Hypertens. 2011, 13, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Silbergeld, E.K.; Silva, I.A.; Nyland, J.F. Mercury and autoimmunity: Implications for occupational and environmental health. Toxicol. Appl. Pharmacol. 2005, 207, 282–292. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC Scientific Publications: Lyon, France, 1993. [Google Scholar]
- Karki, R.; Pandya, D.; Elston, R.C.; Ferlini, C. Defining “mutation” and “polymorphism” in the era of personal genomics. BMC Med. Genom. 2015, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Shastry, B.S. SNPs: Impact on gene function and phenotype. Methods Mol. Biol. 2009, 578, 3–22. [Google Scholar] [PubMed]
- Custodio, H.M.; Broberg, K.; Wennberg, M.; Jansson, J.H.; Vessby, B.; Hallmans, G.; Stegmayr, B.; Skerfving, S. Polymorphisms in glutathione-related genes affect methylmercury retention. Arch. Environ. Health 2004, 59, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Custodio, H.M.; Harari, R.; Gerhardsson, L.; Skerfving, S.; Broberg, K. Genetic influences on the retention of inorganic mercury. Arch. Environ. Occup. Health 2005, 60, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Engström, K.S.; Strömberg, U.; Lundh, T.; Johanson, I.; Vessby, B.; Hallmans, G.; Skerfving, S.; Broberg, K. Genetic variation in glutathione-related genes and body burden of methylmercury. Environ. Health Perspect. 2008, 116, 734–739. [Google Scholar] [CrossRef] [PubMed]
- Gundacker, C.; Komarnicki, G.; Jagiello, P.; Gencikova, A.; Dahmen, N.; Wittman, K.J.; Gencik, M. Glutathione-S-transferase polymorphism, metallothionein expression, and mercury levels among students in Austria. Sci. Total Environ. 2007, 385, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Gundacker, C.; Wittmann, K.J.; Kukuckova, M.; Komarnicki, G.; Hikkel, I.; Gencik, M. Genetic background of lead and mercury metabolism in a group of medical students in Austria. Environ. Res. 2009, 109, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Barcelos, G.R.; de Marco, K.C.; Grotto, D.; Valentini, J.; Garcia, S.C.; Leite Braga, G.Ú.; Barbosa, F., Jr. Evaluation of glutathione S-transferase GSTM1 and GSTT1polymorphisms and methylmercury metabolism in an exposed Amazon population. J. Toxicol. Environ. Health A 2012, 75, 960–970. [Google Scholar] [CrossRef] [PubMed]
- Parajuli, R.P.; Goodrich, J.M.; Chou, H.N.; Gruninger, S.E.; Dolinoy, D.C.; Franzblau, A.; Basu, N. Genetic polymorphisms are associated with hair, blood, and urine mercury levels in the American Dental Association (ADA) study participants. Environ. Res. 2016, 149, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Engström, K.S.; Wennberg, M.; Strömberg, U.; Bergdahl, I.A.; Hallmans, G.; Jansson, J.H.; Lundh, T.; Norberg, M.; Rentschler, G.; Vessby, B.; et al. Evaluation of the impact of genetic polymorphisms in glutathione-related genes on the association between methylmercury or N-3 polyunsaturated long chain fatty acids and risk of myocardial infarction: A case-control study. Environ. Health 2011, 19, 10–33. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Hong, Y.; Park, H.; Ha, M.; Koo, B.S.; Chang, N.; Roh, Y.M.; Kim, B.N.; Kim, Y.J.; Kim, B.M.; et al. Interaction between GSTM1/GSTT1 polymorphism and blood mercury on birth weight. Environ. Health Perspect. 2010, 118, 437–443. [Google Scholar] [PubMed]
- Julvez, J.; Smith, G.D.; Golding, J.; Ring, S.; Pourcain, B.S.; Gonzalez, J.R.; Grandjean, P. Prenatal methylmercury exposure and genetic predisposition to cognitive deficit at age 8 years. Epidemiology 2013, 24, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Goodrich, J.M.; Gillespie, B.; Werner, R.; Basu, N.; Franzblau, A. An investigation of modifying effects of metallothionein single-nucleotide polymorphisms on the association between mercury exposure and biomarker levels. Environ. Health Perspect. 2012, 120, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Engström, K.; Love, T.M.; Watson, G.E.; Zareba, G.; Yeates, A.; Wahlberg, K.; Alhamdow, A.; Thurston, S.W.; Mulhern, M.; McSorley, E.M.; et al. Polymorphisms in ATP-binding cassette transporters associated with maternal methylmercury disposition and infant neurodevelopment in mother-infant pairs in the Seychelles Child Development Study. Environ. Int. 2016, 94, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Llop, S.; Engström, K.; Ballester, F.; Franforte, E.; Alhamdow, A.; Pisa, F.; Tratnik, J.S.; Mazej, D.; Mario, M.; Marisa, R.; et al. Polymorphisms in ABC transporter genes and concentrations of mercury in newborns-Evidence from two Mediterranean birth cohorts. PLoS ONE 2014, 9, e97172. [Google Scholar] [CrossRef] [PubMed]
- Strange, R.C.; Spiteri, M.A.; Ramachandran, S.; Fryer, A.A. Glutathione-S-transferase family of enzymes. Mutat. Res. 2001, 482, 21–26. [Google Scholar] [CrossRef]
- Barcelos, G.R.; Souza, M.F.; Oliveira, A.Á.; Lengert, A.V.; Oliveira, M.T.; Camargo, R.B.; Grotto, D.; Valentini, J.; Garcia, S.C.; Braga, G.Ú. Effects of genetic polymorphisms on antioxidant status and concentrations of the metals in the blood of riverside Amazonian communities co-exposed to mercury and Pb. Environ. Res. 2015, 138, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, J.M.; Basu, N. Variants of glutathione s-transferase pi 1 exhibit differential enzymatic activity and inhibition by heavy metals. Toxicol. In Vitro 2012, 26, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Babula, P.; Masarik, M.; Adam, V.; Eckschlager, T.; Stiborova, M.; Trnkova, L.; Skutkova, H.; Provaznik, I.; Hubalek, J.; Kizek, R. Mammalian metallothioneins: Properties and functions. Metallomics 2012, 4, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Woods, J.S.; Heyer, N.J.; Russo, J.E.; Martin, M.D.; Pillai, P.B.; Farin, F.M. Modification of neurobehavioral effects of mercury by genetic polymorphisms of metallothionein in children. Neurotoxicol. Teratol. 2013, 39, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Ralston, N.V.C.; Ralston, C.R.; Raymond, L.J. Selenium Health Benefit Values: Updated Criteria for Mercury Risk Assessments. Biol. Trace Elem. Res. 2016, 171, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Penglase, S.; Hamre, H.; Ellingsen, S. Selenium prevents downregulation of antioxidant selenoprotein genes by methylmercury. Free Radic. Biol. Med. 2014, 75, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Yu, H.; Zhao, J.; Li, B.; Qu, L.; Liu, S.; Zhang, P.; Chai, Z. The roles of serum selenium and selenoproteins on mercury toxicity in environmental and occupational exposure. Environ. Health Perspect. 2006, 114, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.F.; Hill, K.E. Selenoprotein P-expression, functions, and roles in mammals. Biochim. Biophys. Acta 2009, 1790, 1441–1447. [Google Scholar] [CrossRef] [PubMed]
- Méplan, C.; Nicol, F.; Burtle, B.T.; Crosley, L.K.; Arthur, J.R.; Mathers, J.C.; Hesketh, J.E. Relative abundance of selenoprotein P isoforms in human plasma depends on genotype, Se intake, and cancer status. Antioxid. Redox Signal. 2009, 11, 2631–2640. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, J.M.; Wang, Y.; Gillespie, B.; Werner, R.; Franzblau, A.; Basu, N. Methylmercury and elemental mercury differentially associate with blood pressure among dental professionals. Int. J. Hyg. Environ. Health 2013, 216, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Vasiliou, V.; Vasiliou, K.; Nebert, D.W. Human ATP-binding cassette (ABC) transporter family. Hum. Genom. 2009, 3, 281–290. [Google Scholar] [CrossRef]
- Bridges, C.C.; Zalups, R.K. Molecular and ionic mimicry and the transport of toxic metals. Toxicol. Appl. Pharmacol. 2005, 204, 274–308. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, P.; Weihe, P.; White, R.F.; Debes, F.; Araki, S.; Yokoyama, K.; Murata, K.; Sørensen, N.; Dahl, R.; Jørgensenm, P.J. Cognitive deficit in 7-year-old children with prenatal exposure to methylmercury. Neurotoxicol. Teratol. 1997, 19, 417–428. [Google Scholar] [CrossRef]
- Grandjean, P.; Weihe, P.; White, R.F.; Debes, F. Cognitive performance of children prenatally exposed to “safe” levels of methylmercury. Environ. Res. 1998, 77, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Myers, G.J.; Davidson, P.W.; Cox, C.; Shamlaye, C.; Palumbo, D.; Cernichiari, E.; Sloane-Reeves, J.; Wilding, G.E.; Kost, J.; Huang, L.S.; et al. Prenatal methylmercury exposure from ocean fish consumption in the Seychelles Child Development Study. Lancet 2003, 361, 1686–1692. [Google Scholar] [CrossRef]
- Ng, S.; Lin, C.C.; Jeng, S.F.; Hwang, Y.H.; Hsieh, W.S.; Chen, P.C. Mercury, APOE, and child behavior. Chemosphere 2014, 120, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.; Lin, C.C.; Hwang, Y.H.; Hsieh, W.S.; Liao, H.F.; Chen, P.C. Mercury, APOE, and children’s neurodevelopment. Neurotoxicology 2013, 37, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Woods, J.S.; Heyer, N.J.; Russo, J.E.; Martin, M.D.; Farin, F.M. Genetic polymorphism affecting susceptibility to mercury neurotoxicity in children: Summary findings from the Casa Pia Children’s Amalgam clinical trial. Neurotoxicology 2014, 44, 288–302. [Google Scholar] [CrossRef] [PubMed]
- Echeverria, D.; Woods, J.S.; Heyer, N.J.; Rohlman, D.S.; Farin, F.M.; Bittner, A.C., Jr.; Li, T.; Garabedian, C. Chronic low-level mercury exposure, BDNF polymorphism, and associations with cognitive and motor functions. Neurotoxicol. Teratol. 2005, 27, 781–796. [Google Scholar] [CrossRef] [PubMed]
- Heyer, N.J.; Echeverria, D.; Bittner, A.C., Jr.; Farin, F.M.; Garabedian, C.C.; Woods, J.S. Chronic low-level mercury exposure BDNF polymorphism, and associations with self-reported symptoms and mood. Toxicol. Sci. 2004, 81, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Heyer, N.J.; Echeverria, D.; Martin, M.D.; Farin, F.M.; Woods, J.S. Catechol O methyl transferase (COMT) VAL158MET functional polymorphism, dental mercury exposure, and self-reported symptoms and mood. J. Toxicol. Environ. Health A 2009, 72, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Woods, J.S.; Echeverria, D.; Heyer, N.J.; Simmonds, P.L.; Wilkerson, J.; Farin, F.M. The association between genetic polymorphisms of coproporphyrinogen oxidase and an atypical porphryinogenic response to mercury exposure in humans. Toxicol. Appl. Pharmacol. 2005, 206, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Woods, J.S.; Heyer, N.J.; Echeverria, D.; Russo, J.E.; Martin, M.D.; Bernardo, M.F.; Luis, H.S.; Vaz, L.; Farin, F.M. Modification of neurobehavioral effects of mercury by a genetic polymorphism of coproporphyrinogen oxidase in children. Neurotoxicol. Teratol. 2012, 34, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Echeverria, D.; Woods, J.S.; Heyer, N.J.; Martin, M.D.; Rohlman, D.S.; Farin, F.M.; Li, T.; Garabedian, C.E. The association between a genetic polymorphism of coproporphyrinogen oxidase, dental mercury exposure and neurobehavioral response in humans. Neurotoxicol. Teratol. 2006, 28, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Woods, J.S. Cloning, expression, and biochemical properties of CPOX4, a genetic variant of coproporphyrinogen oxidase that affects susceptibility to mercury toxicity in humans. Toxicol. Sci. 2009, 109, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Echeverria, D.; Woods, J.S.; Heyer, N.J.; Martin, M.D.; Rohlman, D.S.; Farin, F.M.; Li, T. The association between serotonin transporter gene promoter polymorphism (5-HTTLPR) and elemental mercury exposure on mood and behavior in humans. J. Toxicol. Environ. Health A 2010, 73, 1003–1020. [Google Scholar] [CrossRef] [PubMed]
- Heyer, N.J.; Echeverria, D.; Farin, F.M.; Woods, J.S. The association between serotonin transporter gene promotor polymorphism (5-HTTLPR), self-reported symptoms, and dental mercury exposure. J. Toxicol. Environ. Health A 2008, 71, 1381–1386. [Google Scholar] [CrossRef] [PubMed]
- Woods, J.S.; Heyer, N.J.; Russo, J.E.; Martin, M.D.; Pillai, P.B.; Bammler, T.K.; Farin, F.M. Genetic polymorphisms of catechol-O-methyltransferase modify the neurobehavioral effects of mercury in children. J. Toxicol. Environ. Health A 2014, 77, 293–312. [Google Scholar] [CrossRef] [PubMed]
- Heyer, N.J.; Bittner, A.C., Jr.; Echeverria, D.; Woods, J.S. A cascade analysis of the interaction of mercury and coproporphyrinogen oxidase (CPOX) polymorphism on the heme biosynthetic pathway and porphyrin production. Toxicol. Lett. 2006, 161, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Strittmatter, W.J.; Saunders, A.M.; Schmechel, D.; Pericak-Vance, M.; Enghild, J.; Salvesen, G.S.; Roses, A.D. Apolipoprotein E: High-avidity binding to beta-amyloid and increased frequency of type 4 allele in late-onset familial Alzheimer disease. Proc. Natl. Acad. Sci. USA 1993, 90, 1977–1981. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, M.E.; Wojcik, D.P.; Krone, C.A. Apolipoprotein E genotyping as a potential biomarker for mercury neurotoxicity. J. Alzheimer’s Dis. 2003, 5, 189–195. [Google Scholar]
- Grandjean, P.; Murata, K.; Budtz-Jorgensen, E.; Weihe, P. Cardiac autonomic activity in methylmercury neurotoxicity: 14-year follow-up of a Faroese birth cohort. J. Pediatr. 2004, 144, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Yorifuji, T.; Tsuda, T.; Kashima, S.; Takao, S.; Harada, M. Long-term exposure to methylmercury and its effects on hypertension in Minamata. Environ. Res. 2010, 110, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Shi, P.; Morris, J.S.; Spiegelman, D.; Grandjean, P.; Siscovick, D.S.; Willett, W.C.; Rimm, E.B. Mercury exposure and risk of cardiovascular disease in two U.S. cohorts. N. Engl. J. Med. 2001, 364, 1116–1125. [Google Scholar] [CrossRef] [PubMed]
- Salonen, J.T.; Seppanen, K.; Lakka, T.A.; Salonen, R.; Kaplan, G.A. Mercury accumulation and accelerated progression of carotid atherosclerosis: A population based prospective 4-year follow-up study in men in eastern Finland. Atherosclerosis 2000, 148, 265–273. [Google Scholar] [CrossRef]
- Virtanen, J.K.; Voutilainen, S.; Rissanen, T.H.; Mursu, J.; Tuomainen, T.P.; Korhonen, M.J.; Valkonen, V.P.; Seppänen, K.; Laukkanen, J.A.; Salonen, J.T. Mercury, fish oils, and risk of acute coronary events and cardiovascular disease, coronary heart disease, and all-cause mortality in men in eastern Finland. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Roman, H.; Walsh, T.; Coull, B.; Dewailly, E.; Guallar, E.; Hattis, D.; Mariën, K.; Schwartz, J.; Stern, A.H.; Virtanen, J.K.; et al. Evaluation of the cardiovascular effects of methylmercury exposures: Current evidence supports development of a dose-response function for regulatory benefits analysis. Environ. Health Perspect. 2011, 119, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Jacob-Ferreira, A.L.; Passos, C.J.; Jordão, A.A.; Fillion, M.; Mergler, D.; Lemire, M.; Gerlach, R.F.; Barbosa, F., Jr.; Tanus-Santos, J.E. Mercury Exposure Increases Circulating Net Matrix Metalloproteinase (MMP)-2 and MMP-9 Activities. Basic Clin. Pharmacol. Toxicol. 2009, 105, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Jacob-Ferreira, A.L.; Passos, C.J.; Gerlach, R.F.; Barbosa, F., Jr.; Tanus-Santos, J.E. A functional matrix metalloproteinase (MMP)-9 polymorphism modifies plasma MMP-9 levels in subjects environmentally exposed to mercury. Sci. Total Environ. 2010, 408, 4085–4092. [Google Scholar] [CrossRef] [PubMed]
- Jacob-Ferreira, A.L.; Lacchini, R.; Gerlach, R.F.; Passos, C.J.; Barbosa, F., Jr.; Tanus-Santos, J.E. A common matrix metalloproteinase (MMP)-2 polymorphism affects plasma MMP-2 levels in subjects environmentally exposed to mercury. Sci. Total Environ. 2011, 409, 4242–4246. [Google Scholar] [CrossRef] [PubMed]
- Barcelos, G.R.; De Marco, K.C.; de Rezende, V.B.; Braga, G.Ú.; Antunes, L.M.; Tanus-Santos, J.E.; Barbosa, F., Jr. Genetic Effects of eNOS Polymorphisms on Biomarkers Related to Cardiovascular Status in a Population Coexposed to Methylmercury and Lead. Arch. Environ. Contam. Toxicol. 2015, 69, 173–180. [Google Scholar] [CrossRef] [PubMed]
- De Marco, K.C.; Antunes, L.M.; Tanus-Santos, J.E.; Barbosa, F., Jr. Intron 4 polymorphism of the endothelial nitric oxide synthase (eNOS) gene is associated with decreased NO production in a mercury-exposed population. Sci. Total Environ. 2012, 414, 708–712. [Google Scholar] [CrossRef] [PubMed]
- De Marco, K.C.; Braga, G.U.; Barbosa, F. Determination of the effects of eNOS gene polymorphisms (T-786C and Glu298Asp) on nitric oxide levels in a methylmercury-exposed population. J. Toxicol. Environ. Health A 2011, 74, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Julvez, J.; Grandjean, P. Genetic susceptibility to methylmercury developmental neurotoxicity matters. Front. Genet. 2013, 4, 278. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Human Organ System | Exposure Pathway | Marker | Associated Outcome |
|---|---|---|---|
| Nervous system | Transplacental for maternal occupational settings or dietary intake. Breast-feeding and ingestion of contaminated food. Inhalation, for occupational exposure. Use of thimerosal in vaccines. Cultural or religious practices. Dermal exposure. Chronic exposure from amalgam dental fillings. Inhalation, for occupational exposure or dental amalgams replacement. Ingestion of contaminated food, particularly in fish eating populations. | Cord blood. Maternal milk, hair and blood. Child blood, hair and urine. Adult urine, blood and hair. | Infants/Children: Deficit in language (late talking) and memory, deficit in attention, decrements in infant cognition and neurobehavioral deficits. Adults: Depression, paranoia, extreme irritability, hallucinations, inability to concentrate, memory loss, tremors of the hands, head, lips, tongue and eyelids, low body temperature, drowsiness, headaches, weight loss, insomnia, fatigue, blindness, optic neuropathy, retinopathy, hearing loss, sensory, neurological and behavioral dysfunctions. |
| Motor system | Pre and post-natal exposure. Ingestion, for fish eating populations. Inhalation, for gold mining activities. | Maternal hair. Adult blood, hair and urine | Motor dysfunctions, increased tiredness, reduction in muscle strength and twitching, late walking. |
| Cardiovascular system | Chronic exposure, attributed to fish consumption and gold mining activities. | Adult hair, toenail, plasma and urine | Cardiomyopathy, hypertension, coronary heart disease, myocardial infarction, cardiac arrhythmias, cerebro-vascular accident, ischemic heart disease, generalized atherosclerosis. |
| Pulmonary system | Inhalation, for chronic exposure of volatilized vapors. Inhalation, for burning of Hg-containing material. | Adult urine | Chemical pneumonitis, necrotizing bronchitis, pulmonary fibrosis, cough, dyspnea, chest tightness, asthmatic disorders. |
| Renal system | Occupational contact, for abuse of skin lightening cosmetics or Hg-containing compounds. Chronic exposure, related to the number of dental amalgam fillings. Chronic dietary exposure, for fish consumption. | Adult urine, hair and blood. | Glomerular disease whit oliguria or anuria, increased plasma creatinine level and proteinuria. Subacute-onset nephrotic syndrome, nephritic syndrome, tubular dysfunction, glomerulonephritis. |
| Endocrine/Reproductive system | Prenatal exposure for maternal amalgam fillings/replacement. Chronic exposure from amalgam dental fillings. Exposure to occupational routes | Child urine and blood. Adult urine, hair and blood. | Adrenal hyperplasia and atrophy. Hypothyroidism, thyroid inflammation, and depression. Pancreatic dysfunction. Decreases rate of fertility in both males and females. Birth of abnormal offsprings. |
| Immune/Hematological system | Chronic dietary exposure. Clinical hypersensitivity for Hg-containing amalgam. Prolonged exposure in clinically asymptomatic workers. | Adult urine. Lympho/monocyte stimulation tests. | Induction and exacerbation of autoimmune and allergic diseases in susceptible populations. Decreased immunity of the body. Hemolytic anemia, aplastic anemia. |
| Embrional system | Maternal occupational settings. Maternal dietary exposure. Maternal amalgam fillings/replacement. | Cord blood. Fetal blood. Maternal hair. | Hypoplasia of the cerebellum, decreased number of nerve cells in the cerebral cortex, decreased total brain weight, abnormal neuron migration. Spontaneous abortions, stillbirth, low birth weights. |
| Gene | Protein Function | Polymorphism | “In vitro” Functional Activity | Estimated Interactions | Mercury Metabolism |
|---|---|---|---|---|---|
| GSTM1 | Hg-binding modulation by GSH conjugation | + > – Deletion | Null gene expression and impaired catalytic activity | Urinary Hg levels | I-Hg 1 [31] |
| Hair+ Blood Hg levels | Me-Hg 2 [9] | ||||
| Hair Hg levels | Me-Hg [33,34] | ||||
| Maternal hair Hg levels | Me-Hg [38] | ||||
| Cord blood Hg levels | Me-Hg [38] | ||||
| Plasma/Blood Hg levels | Me-Hg [10] | ||||
| GSTT1 | Hg-binding modulation by GSH conjugation | + > – Deletion | Null gene expression and impaired catalytic activity | Urinary Hg levels | I-Hg [6,7] |
| Hair Hg levels | Me-Hg [33,34] | ||||
| Cord blood Hg levels | Me-Hg [38] | ||||
| GSTP1 | Hg-binding modulation by GSH conjugation | 105 Ile > Val (rs1695) | Decreased catalytic efficiency and protein activity | Urinary Hg levels | I-Hg [31,32,33,34,35,36] |
| Hair Hg levels | Me-Hg [6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34] | ||||
| 114Ala > Val (rs1138272) | Erythrocyte Hg levels | Me-Hg [30,31,32] | |||
| Plasma Hg levels | Me-Hg [10] | ||||
| GSTA1 | Hg-binding modulation by GSH conjugation | 3′UTR (rs3957356) | Lower transcriptional activation and decreased enzyme activity | Blood Hg levels? | Me-Hg [34] |
| GSS | Hg-binding modulation by GSH synthesis | 5’UTR (rs3761144) | Decreased gene expression | Hair Hg levels | Me-Hg [6] |
| GCLC | Hg-binding modulation by GSH synthesis | 5′ near gene (rs17883901) | Suppressed gene induction and promoter activity | Urinary Hg levels | I-Hg [31] |
| Hair Hg levels | Me-Hg [33,34] | ||||
| Intron 9 (rs1555903) * | Erythrocyte Hg levels | Me-Hg [30] | |||
| Blood Hg levels | Me-Hg [10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36] | ||||
| Cord blood Hg levels * | Me-Hg [39] | ||||
| GCLM | Hg-binding modulation by GSH synthesis | 5’ near gene (rs41303970) | Decrease promoter activity and gene expression | Urinary Hg levels | I-Hg [7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31] |
| Hair, Blood Hg levels | Me-Hg [9,30,37] | ||||
| Plasma, Blood Hg levels | Me-Hg [10] | ||||
| Erythrocyte Hg levels | Me-Hg [32] | ||||
| Plasma Hg levels | I-Hg [7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31] | ||||
| MT1M | Hg-binding and detoxifying capacity | 3′UTR (rs2270837) * | Altered transcriptional activity | Urinary Hg levels * | I-Hg [40] |
| 3′UTR (rs2270836) * | Hair, Blood Hg levels * | Me-Hg [36] | |||
| 3′UTR (rs9936471) * | Hair Hg levels * | Me-Hg [40] | |||
| MT2A | Hg-binding and detoxifying capacity | 3′UTR (rs10636) | Altered transcriptional activity | Urinary Hg levels | I-Hg [40] |
| MT1A | Hg-binding and detoxifying capacity | 51 Lys > Arg (rs8052394) | Altered transcriptional activity and protein structure | Hair Hg levels | Me-Hg [40] |
| 27 Thr > Asn (rs11640851) | |||||
| MT4 | Hg-binding and detoxifying capacity | 48 Gly > Asp (rs11643815) | Altered transcriptional activity and protein structure | Hair Hg levels | Me-Hg [34] |
| Hair, Blood Hg levels | Me-Hg [36] | ||||
| GPX1 GPX4 | Hg-detoxification and modulation by GSH metabolism | 200 Pro > Leu (rs1050450) | Defective gene expression and reduced protein activity | Undefined | None |
| Undefined | None | ||||
| 3′UTR (rs713041) | |||||
| SEPP1 | Hg-detoxification and distribution by Se-P synthesis | 3′UTR (rs7579) | Impaired gene expression and protein synthesis | Urinary Hg levels | I-Hg [6] |
| Hair Hg levels | Me-Hg [6] | ||||
| ABCC2 (MRP2) | Hg-transport and elimination | 5′UTR (rs1885301) * (rs717620) * | Defective promoter sites activity and altered protein | Urinary Hg levels * | I-Hg [5] |
| Maternal hair Hg levels * | Me-Hg [41] | ||||
| Cord blood Hglevels * | Me-Hg [42] | ||||
| 417 Val > Ile (rs2273697) * | |||||
| ABCB1 (MDR1) | Hg-transport and elimination | 893 Ala > Ser (rs2032582) * | Impaired protein structure and enzyme activity | Cord blood Hg levels | Me-Hg [42] |
| Maternal hair Hg levels * | Me-Hg [41] | ||||
| 3′UTR (rs12076499) * (rs1202169) * | |||||
| ABCC1 (MRP1) | Hg-transport and elimination | Intron 1 (rs11075290) * | Defective transcription factor binding sites | Cord blood Hg levels * | Me-Hg [42] |
| Maternal hair Hg levels * | Me-Hg [41] | ||||
| 5′UTR (rs212093) * (rs215088) * | |||||
| SLC7A5 (LAT1) | Hg-uptake and distribution | 5′UTR (rs33916661) | Altered transcriptional activity | Urinary Hg levels | I-Hg [5] |
| SLC22A6 (OAT1) | Hg-uptake and distribution | 5′UTR (rs4149170) | Altered transcriptional activity | Urinary Hg levels | I-Hg [5] |
| SLC22A8 (OAT3) | Hg-uptake and distribution | 5′UTR (rs4149182) | Altered transcriptional activity | Urinary Hg levels | I-Hg [5] |
| ATP7B | Hg-transport? | 832 Lys > Arg (rs1061472) | Altered mechanism of copper transport | Hair Hg levels | Me-Hg [36] |
| 952 Arg > Lys (rs732774) | |||||
| BDNF | Hg-neurotoxicity? | 66 Val > Met (rs6265) | Altered survival of striatal neurons in the brain | Hair Hg levels | Me-Hg [36] |
| Gene | SNP | Age Class/Place of Origin | Main Outcomes | Estimated Interactions | Exposure |
|---|---|---|---|---|---|
| APOE | * rs7412 | Children (2 years of age)/Taiwan | Adverse effects on cognition, behavior and whole neuro- development in pre-school children carrying the ε4 allele | Cord blood Hg levels | Prenatal MeHg exposure [59,60] |
| rs429358 | Children (8–12 years of age)/Portugal | Impaired neurobehavioral functions, related to working and learning memory, among boys with ε4 allele | Urinary Hg levels | Postnatal inorganic Hg exposure [61] | |
| BDNF | rs6265 rs2049046 | Children (8–12 years of age)/Portugal Adult (Dentists and dental assistants)/USA Children (8 years of age)/UK | Increased risk of neuro-behavioral deficits associated with learning &memory | Urinary Hg levels | Postnatal inorganic Hg exposure [61] |
| Altered cognitive flexibility, working & visual memory. Potential decline of cognitive & motor performance with increased neuro-behavioral symptoms, and mood | Urinary Hg levels | Occupational elemental Hg exposure [62,63] | |||
| Decreased performance IQ, and verbal scores with cognitive involvement in children at school age | Umbilical cord Hg levels | Prenatal MeHg exposure [39] | |||
| COMT | rs4680 rs4633 rs4618 rs6269 | Children (8–12 years of age)/Portugal Adult (Dental assistants)/USA | Impaired neurobehavioral test performance affecting attentional control, working and learning memory, visual spatial acuity (boys); attention control, learning memory, and executive functions (girls) | Urinary Hg levels | Postnatal inorganic Hg exposure [61] |
| Some mood states (tension, depression, fatigue, and confusion) among female assistants | Urinary Hg levels | Postnatal inorganic Hg exposure [64,65] | |||
| CPOX | rs1131857 (CPOX4) | Children (8–12 years of age)/Portugal | Altered performance on multiple neurobehavioral tests within neurological domains (attention, learning &memory, executive function, visual spatial acuity and motor function) (boys); impaired performance affecting learning & memory, executive function (girls) | Urinary Hg levels | Postnatal inorganic Hg exposure [61,66] |
| rs1729995 (CPOX5) | Adult (Dentists and dental assistants)/USA | Deficits in neuropsychological performance within neuro-behavioral (male dentists) and visuomotor (female dental assistants) domains, symptoms and mood | Urinary Hg levels | Occupational elemental Hg exposure [65,67,68] | |
| MTIM | rs2270837 | Children (8–12 years of age)/Portugal | Impaired behavioral performance involving the domains of visual spatial acuity and learning & memory, with some additional impacts on attention and motor function (boys); learning & memory (girls) | Urinary Hg level | Postnatal inorganic Hg exposure [47,61] |
| MT2A | rs10636 | Children (8–12 years of age)/Portugal | Modulation of adverse effects on neurobehavioral (attention, visual spatial acuity, learning & memory), and motor functions (boys) | Urinary Hg level | Postnatal inorganic Hg exposure [47,61] |
| Children (8 years of age)/UK | Increased cognitive consequences in children at school age | Umbilical cord Hg levels | Prenatal MeHg exposure [39] | ||
| PON1 | rs662 | Children (8 years of age)/UK | Cognitive deficit, associated with total and performance IQ, involving children at school age | Umbilical cord Hg levels | Prenatal MeHg exposure [39] |
| PGR | rs1042838 | Children (8 years of age)/UK | Cognitive deficit, associated with total and verbal IQ, involving children at school age | Umbilical cord Hg levels | Prenatal MeHg exposure [39] |
| SLC6A4 44bp (5-HTTLPR) Ins/Del | Children (8–12 years of age)/Portugal | Compromised neurobehavioral test performance affecting atten-tional control, and learning & memory (boys) | Urinary Hg levels | Postnatal inorganic Hg exposure [61] | |
| Adult (Dental assistants)/USA | Deteriorated cognitive skills for prolonged attention-memory, and psychomotor skills for cognitive flexibility, manual coordination (male dentists); attention, working memory, and manual coordination (female dental assistants). Increased mood scales between the two gender groups | Urinary Hg levels | Occupational elemental Hg exposure [69,70] | ||
| TF | rs3811647 | Children (8 years of age)/UK | Decreased Performance IQ among children at school age | Umbilical cord Hg levels | Prenatal MeHg exposure [39] |
| ABCC1 rs11075290 | Infants (20 months of age)/Republic of Seychelles | Compromised neuro-developmental test performance, affecting both mental and psychomotor development | Maternal hair Hg levels | Prenatal MeHg exposure [41] | |
| Gene | Variant | Study Population/Location | Probable Genetic Impact on Cardiovascular Status/Estimated Interactions | Mercury Form |
|---|---|---|---|---|
| GCLM | 5′near gene (* rs4130397) | Matched case-control subjects (age range: 30–77 years)/Northern Sweden | Increased risk of first ever-myocardial infarction, in people without cardiovascular disease/Erythrocyte Hg levels | MeHg [37] |
| MMP-2 | 5′near gene (rs243865) | Exposed subjects, through fish intake (15–87 years of age)/Brazilian Amazon | Potential risk of clinically relevant events, involving acute myocardial infarction, unstable or stable angina, and hypertension/Plasma Hg levels | MeHg [83] |
| MMP-9 | Promoter (CA)n repeats (rs3222264) | Exposed subjects, through fish intake (15–87 years of age)/Brazilian Amazon | Increased susceptibility to rapid coronary artery disease progression, fatal cardiovascular events or hypertension/Plasma Hg levels | MeHg [82] |
| NOS3 | Intron 4 a/b (27 bpVNTR) | Fish eating populations (>18 years old)/Amazon region, Brazil | Increased systolic and diastolic blood pressures, with probable predisposition to hypertension, thrombosis, vasospasm, and atherosclerosis/Plasma and blood nitrile levels Hg-related | MeHg [84] MeHg [85] |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andreoli, V.; Sprovieri, F. Genetic Aspects of Susceptibility to Mercury Toxicity: An Overview. Int. J. Environ. Res. Public Health 2017, 14, 93. https://doi.org/10.3390/ijerph14010093
Andreoli V, Sprovieri F. Genetic Aspects of Susceptibility to Mercury Toxicity: An Overview. International Journal of Environmental Research and Public Health. 2017; 14(1):93. https://doi.org/10.3390/ijerph14010093
Chicago/Turabian StyleAndreoli, Virginia, and Francesca Sprovieri. 2017. "Genetic Aspects of Susceptibility to Mercury Toxicity: An Overview" International Journal of Environmental Research and Public Health 14, no. 1: 93. https://doi.org/10.3390/ijerph14010093
APA StyleAndreoli, V., & Sprovieri, F. (2017). Genetic Aspects of Susceptibility to Mercury Toxicity: An Overview. International Journal of Environmental Research and Public Health, 14(1), 93. https://doi.org/10.3390/ijerph14010093