
Isolation and Characterization of Aquatic-Borne Klebsiella pneumoniae from Tropical Estuaries in Malaysia

 , ,
, ,
Abstract
:1. Introduction
2. Experimental Section
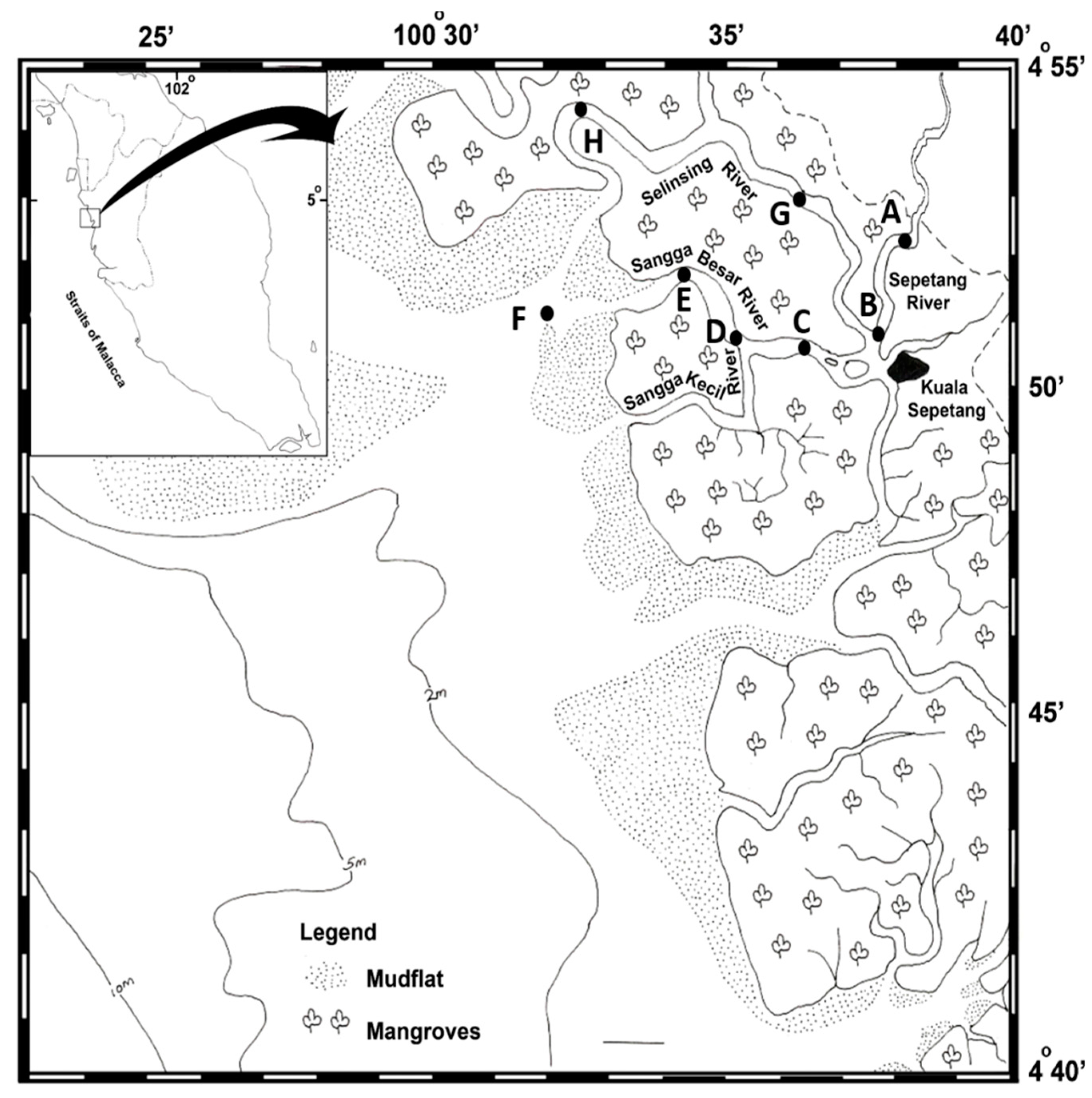
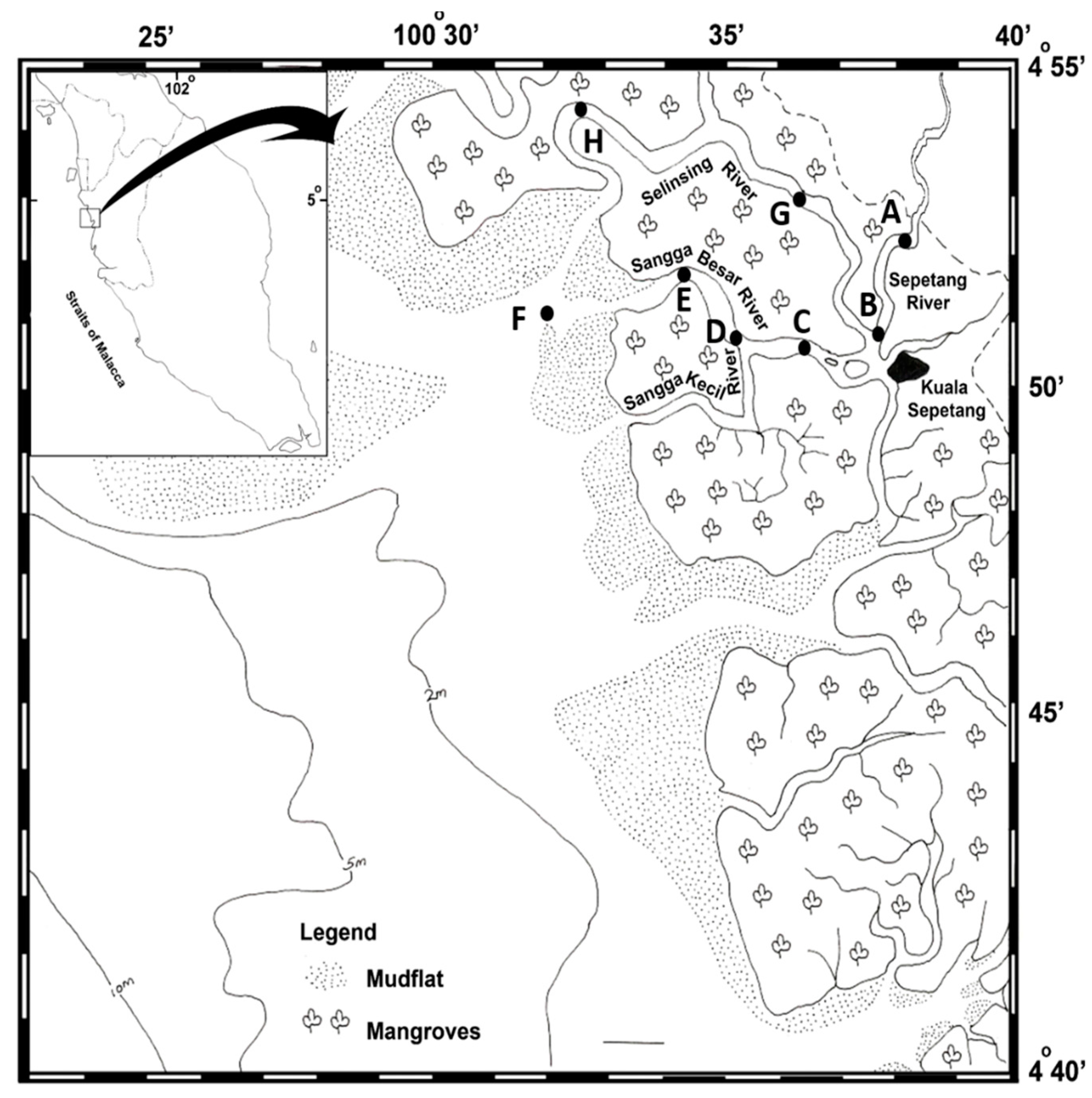
2.1. Sampling Location
2.2. Sample Collection
2.3. Detection and Isolation of K. pneumoniae
2.4. PCR Confirmation of K. pneumoniae
2.5. Detection of Virulence-Associated Genes
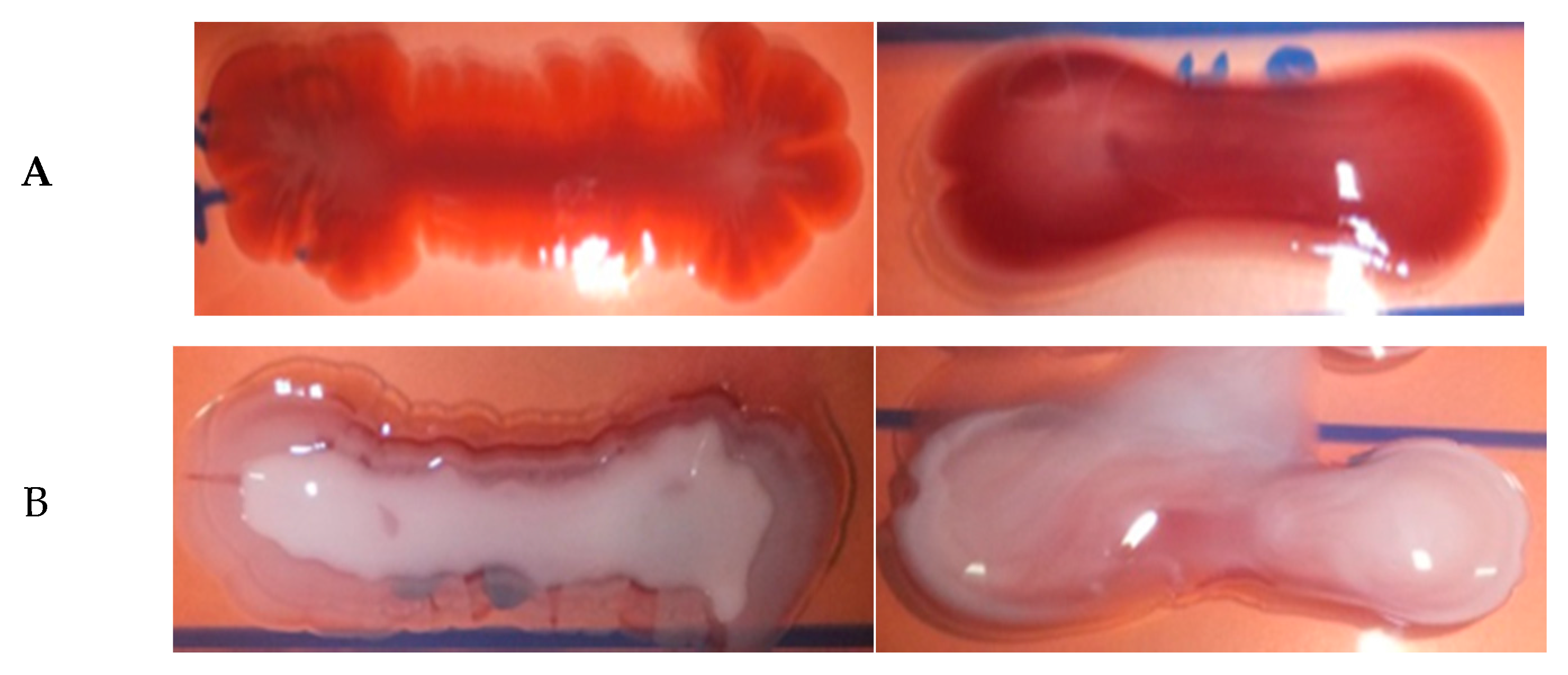
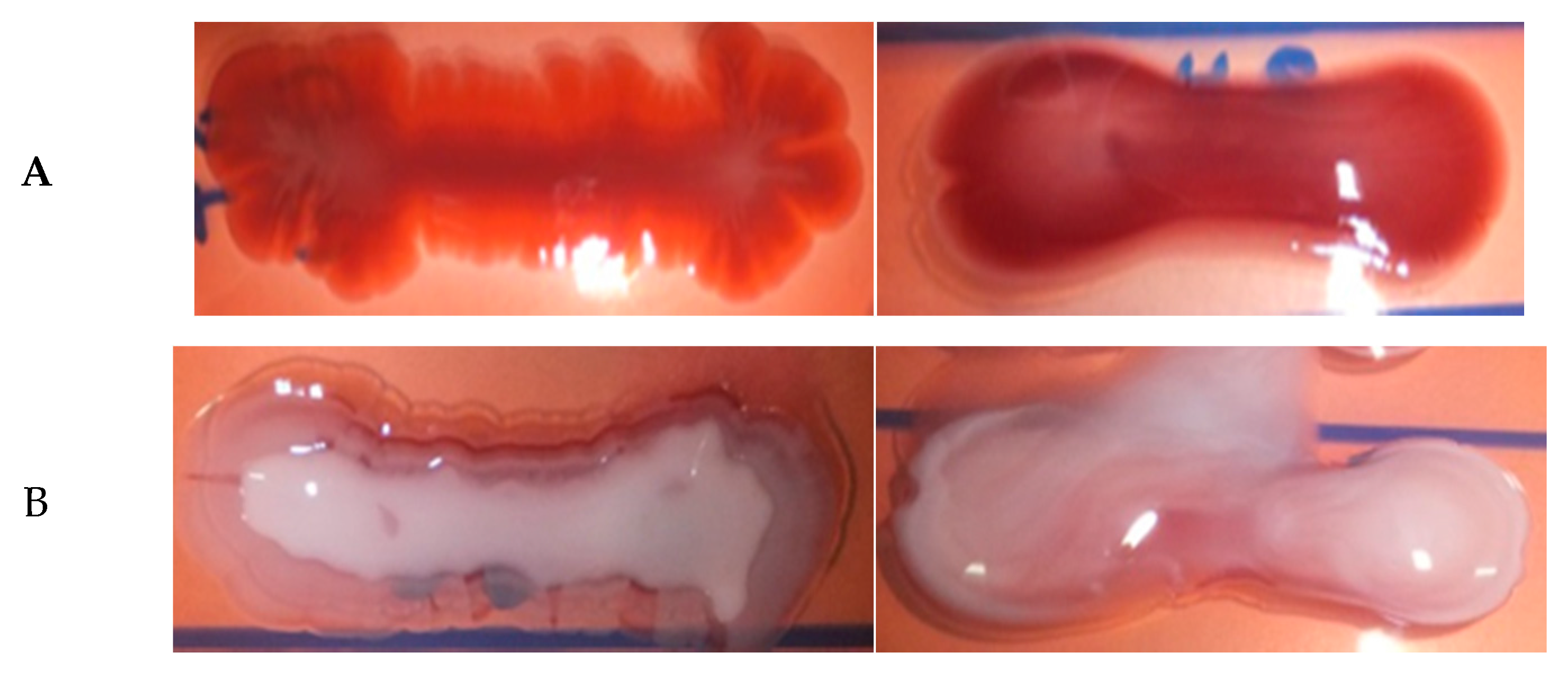
2.6. Hemolytic Assay, Hyperviscosity Test and Capsule Detection
2.7. Morphotypes on Congo Red Agar Plates
2.8. In Vitro Biofilm Formation Assay
2.9. Antibiotic Susceptibility Tests
2.10. Repetitive Extragenic Palindromic PCR (REP-PCR)
2.11. Statistical Analysis
3. Results and Discussion
3.1. Detection and Isolation of K. pneumoniae from Estuarine Water and Sediments
3.2. Virulence Factors and Phenotypic Characterization of Aquatic-Borne K. pneumoniae
3.3. Biofilm Production
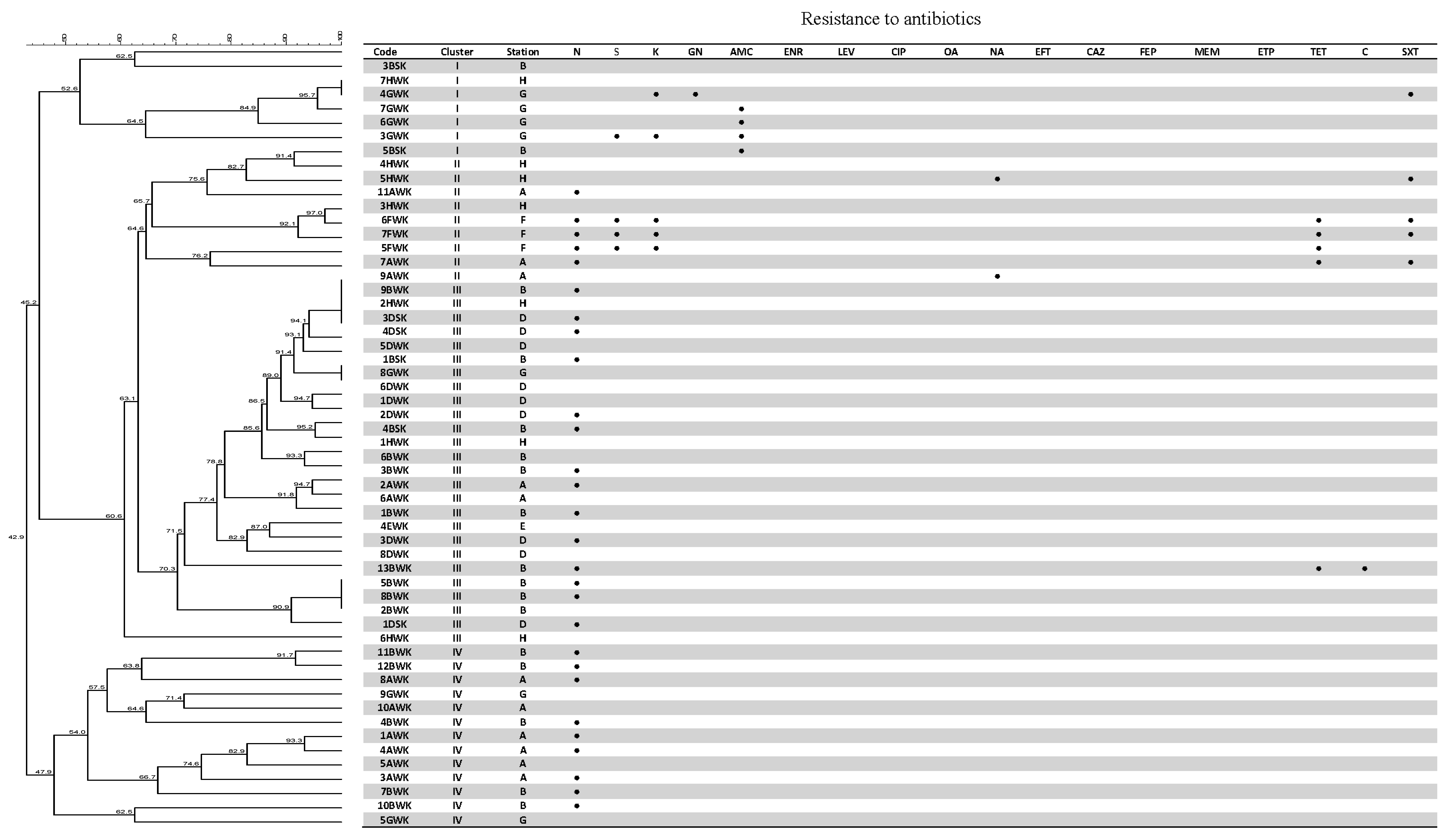
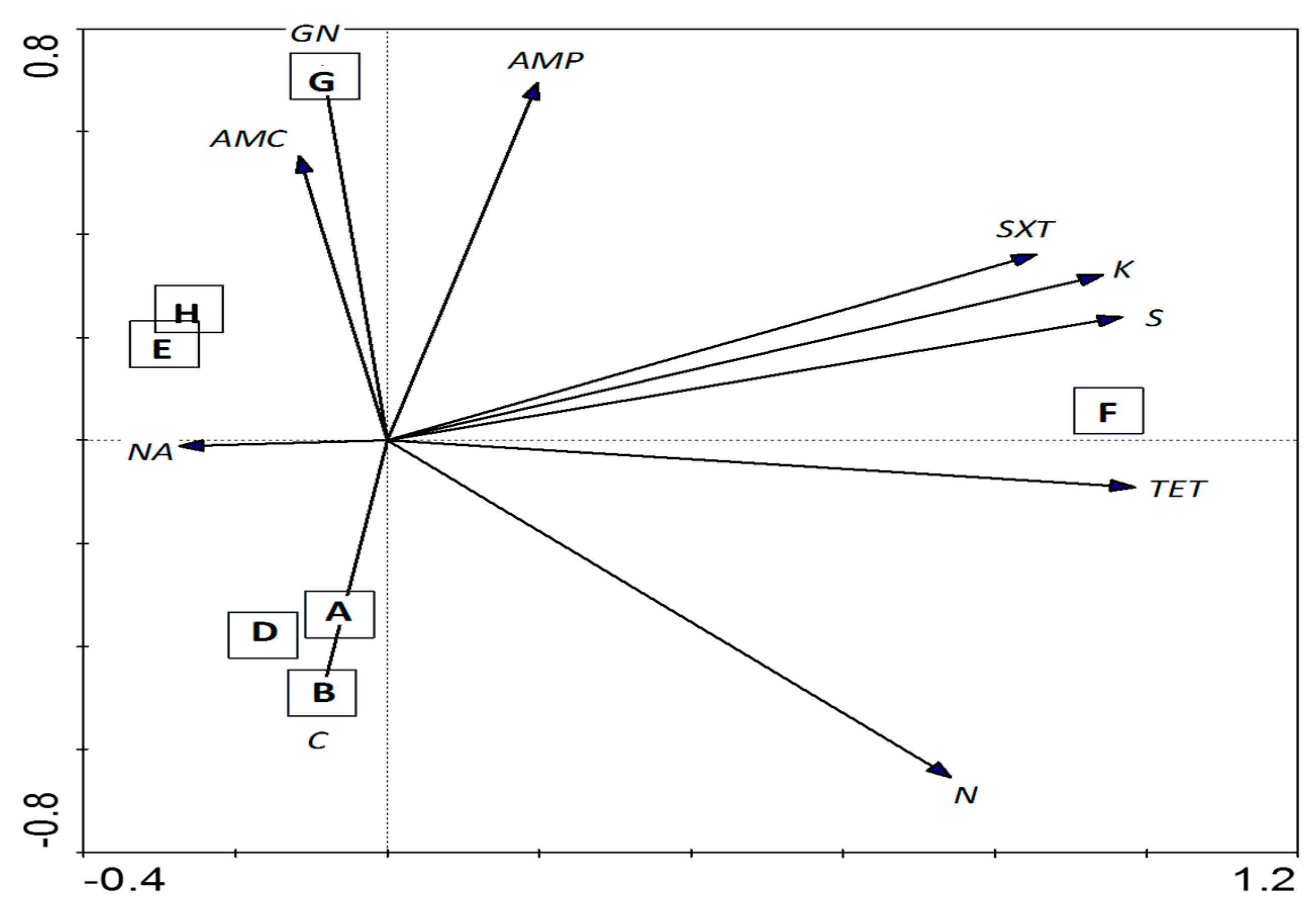
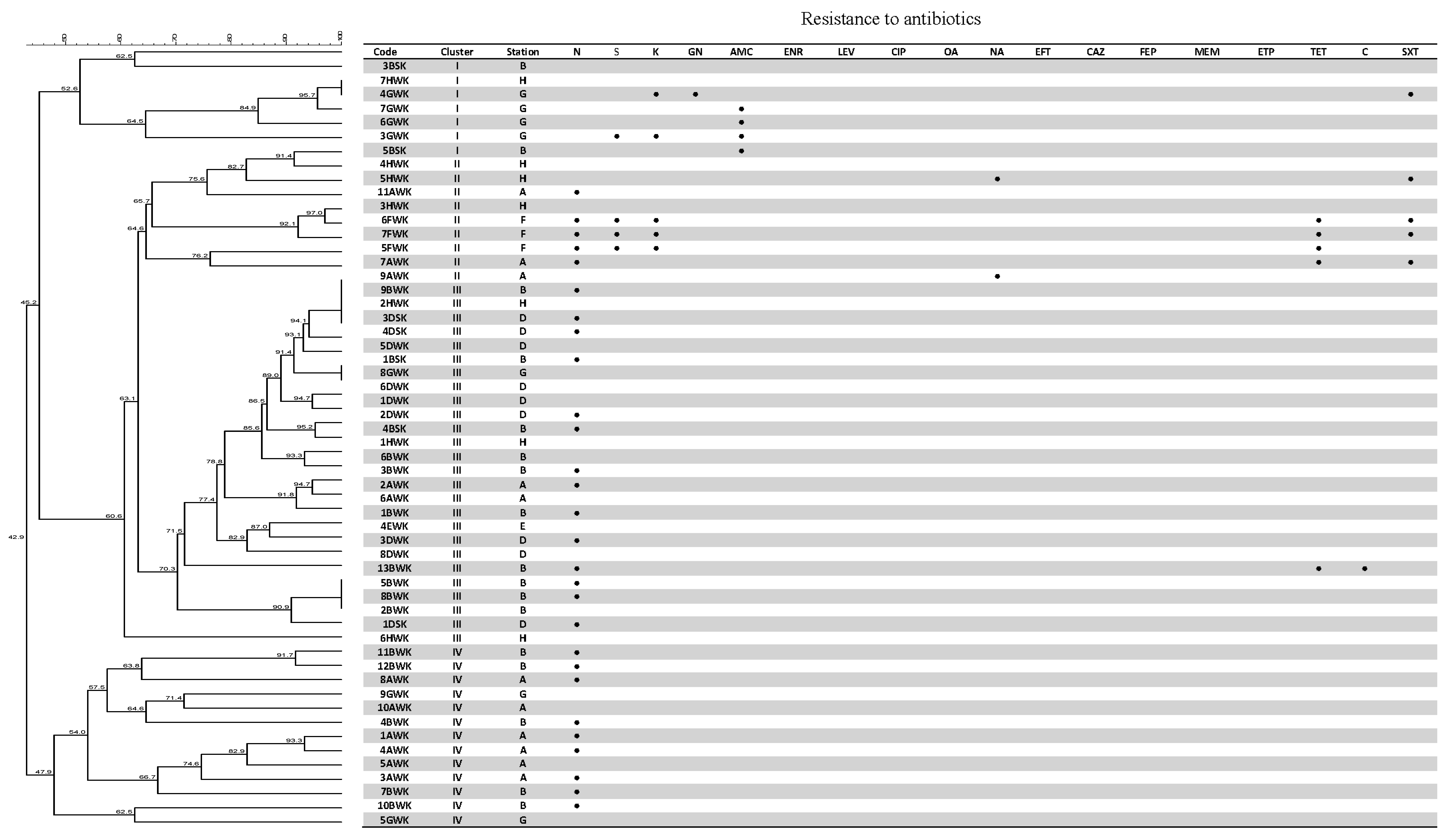
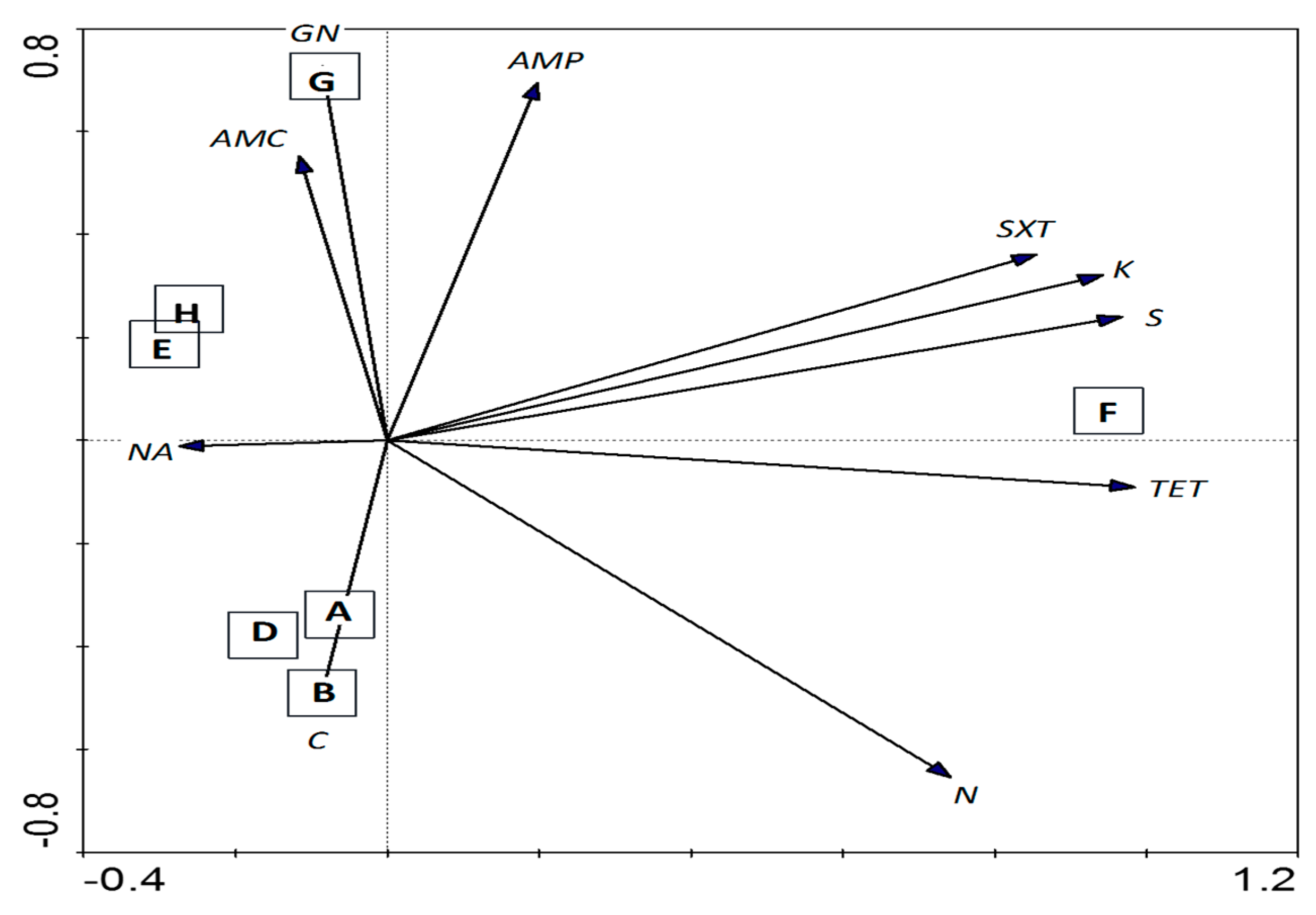
3.4. Antibiotic Susceptibility Test
3.5. Genetic Fingerprinting
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brisse, S.; Grimont, F.; Grimont, P.A. The genus Klebsiella. In The Prokaryotes; Springer: New York, NY, USA, 2006; pp. 159–196. [Google Scholar]
- Abbott, S.L. Klebsiella, Enterobacter, Citrobacter, Serratia, Plesiomonas, and other Enterobacteriaceae. Available online: http://www.asmscience.org/content/book/10.1128/9781555816728.chap37 (accessed on 26 July 2015).
- Bagley, S.T. Habitat association of Klebsiella species. Infect. Control 1985, 6, 52–58. [Google Scholar] [PubMed]
- Struve, C.; Krogfelt, K.A. Pathogenic potential of environmental Klebsiella pneumoniae isolates. Environ. Microbiol. 2004, 6, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Organization, W.H. Guidelines for Drinking-Water Quality: Recommendations; World Health Organization: Geneva, Switzerland, 2004; Volume 1. [Google Scholar]
- Podschun, R.; Pietsch, S.; Höller, C.; Ullmann, U. Incidence of Klebsiella species in surface waters and their expression of virulence factors. Appl. Environ. Microbiol. 2001, 67, 3325–3327. [Google Scholar] [CrossRef] [PubMed]
- Lightfoot, N. Bacteria of potential health concern. In Heterotrophic Plate Counts and Drinking-Water Safety; World Health Organization (WHO): London, UK, 2003; pp. 61–79. [Google Scholar]
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [PubMed]
- Ko, W.-C.; Paterson, D.L.; Sagnimeni, A.J.; Hansen, D.S.; Von Gottberg, A.; Mohapatra, S.; Casellas, J.M.; Goossens, H.; Mulazimoglu, L.; Trenholme, G.; et al. Community-acquired Klebsiella pneumoniae bacteremia: Global differences in clinical patterns. Emerg. Infect. Dis. 2002, 8, 160–166. [Google Scholar]
- Ronald, A. The etiology of urinary tract infection: Traditional and emerging pathogens. Dis. Mon. 2003, 49, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Keynan, Y.; Rubinstein, E. The changing face of Klebsiella pneumoniae infections in the community. Int. J. Antimicrob. Agents 2007, 30, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Muller, L.M.A.J.; Gorter, K.J.; Hak, E.; Goudzwaard, W.L.; Schellevis, F.G.; Hoepelman, A.I.M.; Rutten, G.E.H.M. Increased risk of common infections in patients with type 1 and type 2 diabetes mellitus. Clin. Infect. Dis. 2005, 41, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Lin, H.D.; Wang, L.M. Pyogenic liver abscess associated with septic pulmonary embolism. J. Chin. Med. Assoc. 2008, 71, 442–447. [Google Scholar] [CrossRef]
- Fang, F.C.; Sandler, N.; Libby, S.J. Liver abscess caused by magA+ Klebsiella pneumoniae in North America. J. Clin. Microbiol. 2005, 43, 991–992. [Google Scholar] [CrossRef] [PubMed]
- Rammaert, B.; Goyet, S.; Beauté, J.; Hem, S.; Te, V.; Try, P.L.; Mayaud, C.; Borand, L.; Buchy, P.; Guillard, B. Klebsiella pneumoniae related community-acquired acute lower respiratory infections in Cambodia: Clinical characteristics and treatment. BMC Infecti Dis. 2012, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.T.; Jeng, Y.Y.; Chen, T.L.; Fung, C.P. Bacteremic community-acquired pneumonia due to Klebsiella pneumoniae: Clinical and microbiological characteristics in Taiwan, 2001–2008. BMC Infecti. Dis. 2010, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Xi, C.; Zhang, Y.; Marrs, C.F.; Ye, W.; Simon, C.; Foxman, B.; Nriagu, J. Prevalence of antibiotic resistance in drinking water treatment and distribution systems. Appl. Environ. Microbiol. 2009, 75, 5714–5718. [Google Scholar] [CrossRef] [PubMed]
- Ariffin, R.; Nik Mustafa, N.M.S. A Working Plan for the Matang Mangrove Forest Reserve, 6th Revision ed; Perak State Forestry Department: Perak, Malaysia, 2013. [Google Scholar]
- Ghaderpour, A.; Nasori, K.N.M.; Chew, L.L.; Chong, V.C.; Thong, K.L.; Chai, L.C. Detection of multiple potentially pathogenic bacteria in Matang mangrove estuaries, Malaysia. Mar. Pollut. Bull. 2014, 83, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Cerwenka, H. Pyogenic liver abscess: Differences in etiology and treatment in Southeast Asia and Central Europe. World J. Gastroenterol. 2010, 16, 2458–2462. [Google Scholar] [CrossRef] [PubMed]
- Stiles, M.E.; Ng, L.K. Biochemical characteristics and identification of Enterobacteriaceae isolated from meats. Appl. Environ. Microbiol. 1981, 41, 639–645. [Google Scholar] [PubMed]
- Sun, Z.; Chen, Z.; Hou, X.; Li, S.; Zhu, H.; Qian, J.; Lu, D.; Liu, W. Locked nucleic acid pentamers as universal PCR primers for genomic DNA amplification. PLoS ONE 2008, 3, 3701. [Google Scholar] [CrossRef] [PubMed]
- Araújo, W.L.; de Angellis, D.A.; Azevedo, J.L. Direct RAPD evaluation of bacteria without conventional DNA extraction. Braz. Arch. Biol. Technol. 2004, 47, 375–380. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comp. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, L.; Coderch, N.; Piqué, N.; Bedini, E.; Corsaro, M.M.; Merino, S.; Fresno, S.; Tomás, J.M.; Regué, M. The Klebsiella pneumoniae wabG gene: Role in biosynthesis of the core lipopolysaccharide and virulence. J. Bacteriol. 2003, 185, 7213–7221. [Google Scholar] [CrossRef] [PubMed]
- Regué, M.; Hita, B.; Piqué, N.; Izquierdo, L.; Merino, S.; Fresno, S.; Benedí, V.J.; Tomás, J.M. A gene, uge, is essential for Klebsiella pneumoniae virulence. Infect. Immun. 2004, 72, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Schembri, M.A.; Blom, J.; Krogfelt, K.A.; Klemm, P. Capsule and fimbria interaction in Klebsiella pneumoniae. Infect. Immun. 2005, 73, 4626–4633. [Google Scholar] [CrossRef] [PubMed]
- Yeh, K.M.; Kurup, A.; Siu, L.; Koh, Y.; Fung, C.P.; Lin, J.C.; Koh, T.H. Capsular serotype K1 or K2, rather than magA and rmpA, is a major virulence determinant for Klebsiella pneumoniae liver abscess in Singapore and Taiwan. J. Clin. Microbiol. 2007, 45, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.T.; Lai, S.Y.; Yi, W.C.; Hsueh, P.R.; Liu, K.L.; Chang, S.C. Klebsiella pneumoniae genotype K1: An emerging pathogen that causes septic ocular or central nervous system complications from pyogenic liver abscess. Clin. Infect. Dis. 2007, 45, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Navia, M.M.; Capitano, L.; Ruiz, J.; Vargas, M.; Urassa, H.; Schellemberg, D.; Gascon, J.; Vila, J. Typing and Characterization of Mechanisms of Resistance of Shigella spp. Isolated from Feces of Children under 5 Years of Age from Ifakara, Tanzania. Clin. Microbiol. J. 1999, 37, 3113–3117. [Google Scholar]
- Gerhardt, P. Methods for general and molecular bacteriology. In American Society for Microbiology; Murry, R.G.E., Wood, W.A., Krieg, N.R., Eds.; ASM: Boston, MA, USA, 1994. [Google Scholar]
- Vila, A.; Cassata, A.; Pagella, H.; Amadio, C.; Yeh, K.M.; Chang, F.Y.; Siu, L.K. Appearance of Klebsiella pneumoniae liver abscess syndrome in Argentina: Case report and review of molecular mechanisms of pathogenesis. Open Microbiol. J. 2011, 5, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Corstvet, R.; Gentry, M.; Newman, P.; Rummage, J.; Confer, A. Demonstration of age-dependent capsular material on Pasteurella haemolytica serotype 1. J. Clin. Microbiol. 1982, 16, 1123–1126. [Google Scholar] [PubMed]
- Hammar, M.R.; Arnqvist, A.; Bian, Z.; Olsén, A.; Normark, S. Expression of two csg operons is required for production of fibronectin and Congo red-binding curli polymers in Escherichia coli K-12. Mol. Microbiol. 1995, 18, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Malcova, M.; Karasova, D.; Rychlik, I. aroA and aroD mutations influence biofilm formation in Salmonella Enteritidis. FEMS Microbiol. Lett. 2009, 291, 44–49. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Monteiro, C.; Fang, X.; Ahmad, I.; Gomelsky, M.; Römling, U. Regulation of biofilm components in Salmonella enterica serovar Typhimurium by lytic transglycosylases involved in cell wall turnover. J. Bacteriol. 2011, 193, 6443–6451. [Google Scholar] [CrossRef] [PubMed]
- Römling, U. Characterization of the rdar morphotype, a multicellular behaviour in Enterobacteriaceae. Cell. Mol. Life Sci. CMLS 2005, 62, 1234–1246. [Google Scholar] [CrossRef] [PubMed]
- Zogaj, X.; Bokranz, W.; Nimtz, M.; Römling, U. Production of cellulose and curli fimbriae by members of the family Enterobacteriaceae isolated from the human gastrointestinal tract. Infect. Immun. 2003, 71, 4151–4158. [Google Scholar] [CrossRef] [PubMed]
- Römling, U.; Bokranz, W.; Rabsch, W.; Zogaj, X.; Nimtz, M.; Tschape, H. Occurrence and regulation of the multicellular morphotype in Salmonella serovars important in human disease. Int. J. Med. Microbiol. 2003, 293, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, N.; de Silva Ruiz, C.; Cecilia, M.; Nader-Macias, M. A Simple Technique to Detect Klebsiella Biofilm-Forming-Strains. Inhibitory Potential of Lactobacillus fermentum CRL 1058 Whole Cells and Products. Available online: http://www.formatex.org/microbio/pdf/Pages52-59.pdf (accessed on 26 July 2015).
- Kwasny, S.M.; Opperman, T.J. Static Biofilm Cultures of Gram-Positive Pathogens Grown in a Microtiter Format Used for Anti-Biofilm Drug Discovery. Curr. Protoc. Pharmacol. 2010, 50. [Google Scholar] [CrossRef]
- Performance Standards for Antimicrobial Susceptibility Testing: Twenty-Second Informational Supplement. Available online: http://shop.clsi.org/site/Sample_pdf/M100S25_sample.pdf (accessed on 26 July 2015).
- Lim, K.T.; Yeo, C.C.; Yasin, R.M.; Balan, G.; Thong, K.L. Characterization of multidrug-resistant and extended-spectrum β-lactamase-producing Klebsiella pneumoniae strains from Malaysian hospitals. J. Med. Microbiol. 2009, 58, 1463–1469. [Google Scholar] [CrossRef] [PubMed]
- Ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows, User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2000. [Google Scholar]
- Merlino, J.; Siarakas, S.; Robertson, G.J.; Funnell, G.R.; Gottlieb, T.; Bradbury, R. Evaluation of CHROMagar orientation for differentiation and presumptive identification, gram-negative bacilli and Enterococcus species. J. Clin. Microbiol. 1996, 34, 1788–1793. [Google Scholar] [PubMed]
- Stahlhut, S.G.; Chattopadhyay, S.; Struve, C.; Weissman, S.J.; Aprikian, P.; Libby, S.J.; Fang, F.C.; Krogfelt, K.A.; Sokurenko, E.V. Population variability of the FimH type 1 fimbrial adhesin in Klebsiella pneumoniae. J. Bacteriol. 2009, 191, 1941–1950. [Google Scholar] [CrossRef] [PubMed]
- Szramka, B.; Kurlenda, J.; Bielawski, K. Hemolytic activity of Klebsiella pneumoniae and Klebsiella oxytoca. Med. Doswiadczalna I Mikrobiol. 1997, 50, 207–213. [Google Scholar]
- Struve, C.; Krogfelt, K.A. Role of capsule in Klebsiella pneumoniae virulence: Lack of correlation between in vitro and in vivo studies. FEMS Microbiol. Lett. 2003, 218, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Berkhoff, H.; Vinal, A. Congo red medium to distinguish between invasive and non-invasive Escherichia coli pathogenic for poultry. Avian Dis. 1986, 30, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Qadri, F.; Hossain, S.; Ciznár, I.; Haider, K.; Ljungh, Å.; Wadstrom, T.; Sack, D. Congo red binding and salt aggregation as indicators of virulence in Shigella species. J. Clin. Microbiol. 1988, 26, 1343–1348. [Google Scholar] [PubMed]
- Conjugative Plasmid Mediating Adhesive Pili in Virulent Klebsiella pneumoniae Isolates. Available online: http://www.acmicrob.com/microbiology/conjugative-plasmid-mediating-adhesive-pili-in-virulent-klebsiella-pneumoniae-isolates.php?aid=124 (accessed on 26 July 2015).
- Sharma, K.K.; Soni, S.S.; Meharchandani, S. Congo red dye agar test as an indicator test for detection of invasive bovine Escherichia coli. Vet. Arhiv. 2006, 76, 363–366. [Google Scholar]
- Zogaj, X.; Nimtz, M.; Rohde, M.; Bokranz, W.; Römling, U. The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol. Microbiol. 2001, 39, 1452–1463. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Mohan, H.; Sharma, S.; Chhibber, S. A comparative study of induction of pneumonia in mice with planktonic and biofilm cells of Klebsiella Pneumoniae. Microbiol. Immu. 2011, 55, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Stahlhut, S.G.; Struve, C.; Krogfelt, K.A.; Reisner, A. Biofilm formation of Klebsiella pneumoniae on urethral catheters requires either type 1 or type 3 fimbriae. FEMS Immunol. Med. Microbiol. 2012, 65, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Zhang, Z. Biofilm-forming Klebsiella pneumoniae strains have greater likelihood of producing extended-spectrum β-lactamases. J. Hosp. Infect. 2008, 68, 369–371. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar]
- Boddicker, J.D.; Anderson, R.A.; Jagnow, J.; Clegg, S. Signature-tagged mutagenesis of Klebsiella pneumoniae to identify genes that influence biofilm formation on extracellular matrix material. Infect. Immun. 2006, 74, 4590–4597. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.C.; Lin, T.L.; Hsieh, P.F.; Yang, H.C.; Wang, J.T. Isolation of genes involved in biofilm formation of a Klebsiella pneumoniae strain causing pyogenic liver abscess. PLoS ONE 2011, 6, e23500. [Google Scholar] [CrossRef] [PubMed]
- DeLeo, F.R.; Chen, L.; Porcella, S.F.; Martens, C.A.; Kobayashi, S.D.; Porter, A.R.; Chavda, K.D.; Jacobs, M.R.; Mathema, B.; Olsen, R.J.; et al. Molecular dissection of the evolution of carbapenem-resistant multilocus sequence type 258 Klebsiella pneumoniae. Proc. Natl. Acad. Sci. USA 2014, 111, 4988–4993. [Google Scholar] [CrossRef] [PubMed]
- Lestari, E.S.; Severin, J.A.; Verbrugh, H.A. Antimicrobial resistance among pathogenic bacteria in Southeast Asia. Southeast Asian J. Trop. Med. Public Health 2012, 43, 385–422. [Google Scholar] [PubMed]
- Saiful, A.A.; Mohd, Y.M.; Tay, S. Prevalence of plasmid-mediated qnr determinants and gyrase alteration in Klebsiella pneumoniae isolated from a university teaching hospital in Malaysia. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 1744–1747. [Google Scholar]
- Ahmad, N.; Hashim, R.; Shukor, S.; Khalid, K.N.M.; Shamsudin, F.; Hussin, H. Characterization of the first isolate of Klebsiella pneumoniae carrying New Delhi metallo-β-lactamase and other extended spectrum β-lactamase genes from Malaysia. Med. Microbiol. J. 2013, 62, 804–806. [Google Scholar] [CrossRef] [PubMed]
- Puspanadan, S.; Afsah-Hejri, L.; John, Y.H.T.; Rukayadi, Y.; Loo, Y.Y.; Nillian, E.; Kuan, C.H.; Goh, S.G.; Chang, W.S.; Lye, Y.L.; et al. Characterization of extended-spectrum β-lactamases (ESBLs) producers in Klebsiella pneumoniae by genotypic and phenotypic method. J. Inter. Food Res. 2013, 20, 1479–1483. [Google Scholar]
- Subramanian, P.; Shanmugam, N.; Sivaraman, U.; Kumar, S.; Selvaraj, S. Antiobiotic resistance pattern of biofilm-forming uropathogens isolated from catheterised patients in Pondicherry, India. Australas. Med. J. 2012, 7, 5344. [Google Scholar]
- Sánchez, C.J.; Mende, K.; Beckius, M.L.; Akers, K.S.; Romano, D.R.; Wenke, J.C.; Murray, C.K. Biofilm formation by clinical isolates and the implications in chronic infections. BMC Infect. Dis. 2013, 13, 1. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence | Bp | Annealing Temp | MgC2 | Ref |
|---|---|---|---|---|---|---|
| Mdh | Mdh-f | 5′-GCGTGGCGGTAGATCTAAGTCATA-3′ | 364 | 53 | 1 μL | [22] |
| Mdh-r | 5′-TTCAGCTCCGCCACAAAGGTA-3′ | |||||
| wabG | wabG-f | 5′-ACCATCGGCCATTTGATAGA-3′ | 683 | 55 | 0.5 μL | [25] |
| wabG-r | 5′-CGGACTGGCAGATCCATATC-3′ | |||||
| ugE | ugE-f | 5′-TCTTCACGCCTTCCTTCACT-3′ | 535 | 56 | 1.5 μL | [26] |
| ugE-r | 5′-GATCATCCGGTCTCCCTGTA-3′ | |||||
| FimH | fimH-f | 5′-TGCTGCTGGGCTGGTCGATG-3′ | 550 | 58 | 1.5 μL | [27] |
| fimH-r | 5′-GGGAGGGTGACGGTGACATC-3′ | |||||
| magA | magA-f | 5′-GGTGCTCTTTACATCATTGC-3′ | 1282 | 50 | 1 μL | [28] |
| magA-r | 5′-GCAATGGCCATTTGCGTTAG-3′ | |||||
| rmpA | rmpA-f | 5′-ACTGGGCTACCTCTGCTTCA-3′ | 535 | 50 | 1 μL | [28] |
| rmpA-r | 5′-CTTGCATGAGCCATCTTTCA-3′ | |||||
| K1 | K1-f | 5′-GTAGGTATTGCAAGCCATGC-3′ | 1046 | 56 | 3.0 μL | [29] |
| K2-r | 5′-GCCCAGGTTAATGAATCCGT-3′ | |||||
| K2 | K2-f | 5′-GGAGCCATTTGAATTCGGTG-3′ | 1121 | 56 | 3.0 μL | [29] |
| K2-r | 5′-TCCCTAGCACTGGCTTAAGT-3′ | |||||
| Rep | Rep | 5′-GCGCCGICATGCGGCATT-3′ | variable | 44 | 2.5 μL | [30] |
| Station | No. of Isolates | Capsule | K2 | Hyperviscosity | PDAR Morphotype | RDAR Morphotype | fimH | ugE | wabG |
|---|---|---|---|---|---|---|---|---|---|
| No. (%) | Serotype | No. (%) | No. (%) | No. (%) | No. (%) | No. (%) | No. (%) | ||
| A | 11 | 11 (100.0) | 0 | 0 | 2 (18.18) | 9 (81.82) | 7 (63.6) | 11 (100.0) | 11 (100.0) |
| B | 17 | 16 (94.1) | 6 (35.29) | 4 (23.5) | 5 (29.41) | 12 (70.59) | 12 (70.6) | 14 (82.3) | 15 (88.2) |
| C * | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| D | 9 | 9 (100.0) | 0 | 0 | 2 (22.22) | 7 (77.78) | 5 (55.5) | 4 (44.4) | 8 (88.8) |
| E | 1 | 1 (100.0) | 0 | 0 | 0 | 1 (100) | 1 (100.0) | 1 (100.0) | 1 (100.0) |
| F | 3 | 3 (100.0) | 0 | 0 | 2 (66.66) | 1 (33.34) | 3 (100.0) | 3 (100.0) | 3 (100.0) |
| G | 7 | 6 (85.7) | 0 | 0 | 2 (28.57) | 5 (71.43) | 5 (71.4) | 2 (28.6) | 4 (57.1) |
| H | 7 | 7 (100.0) | 0 | 1 (14.3) | 3 (42.85) | 4 (57.15) | 6 (85.7) | 4 (57.1) | 5 (71.4) |
| Total | 55 | 53 (96.36) | 6 (10.90) | 5 (9.1) | 16 (29.1) | 39 (70.9) | 39 (70.9) | 39 (70.9) | 47 (85.4) |
| Sampling Sites | Salinity | No. of Isolates (%) | No. of Non-Biofilm Formers (%) | No. of Weak Biofilm Formers (%) | No. of Moderate Biofilm Formers (%) | No. of Strong Biofilm Formers (%) |
|---|---|---|---|---|---|---|
| A | 18.5 | 11 | 1 (9.1) | 4 (36.4) | 1 (9.1) | 5 (45.4) |
| B | 19.2 | 17 | 7 (41.2) | 4 (23.5) | 3 (17.6) | 3 (17.6) |
| D | 20.1 | 9 | 1 (11.1) | 4 (44.4) | 4 (44.4) | 0 |
| E | 23.9 | 1 | 0 | 0 | 0 | 1 (100.0) |
| F | 23.5 | 3 | 0 | 0 | 3 (100.0) | 0 |
| G | 16.8 | 7 | 4 (57.1) | 3 (42.9) | 0 | 0 |
| H | 18.6 | 7 | 0 | 4 (57.1) | 0 | 3 (42.8) |
| TOTAL | 55 | 13 (23.6) | 19 (34.5) | 11 (20.0) | 12 (21.8) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barati, A.; Ghaderpour, A.; Chew, L.L.; Bong, C.W.; Thong, K.L.; Chong, V.C.; Chai, L.C. Isolation and Characterization of Aquatic-Borne Klebsiella pneumoniae from Tropical Estuaries in Malaysia. Int. J. Environ. Res. Public Health 2016, 13, 426. https://doi.org/10.3390/ijerph13040426
Barati A, Ghaderpour A, Chew LL, Bong CW, Thong KL, Chong VC, Chai LC. Isolation and Characterization of Aquatic-Borne Klebsiella pneumoniae from Tropical Estuaries in Malaysia. International Journal of Environmental Research and Public Health. 2016; 13(4):426. https://doi.org/10.3390/ijerph13040426
Chicago/Turabian StyleBarati, Anis, Aziz Ghaderpour, Li Lee Chew, Chui Wei Bong, Kwai Lin Thong, Ving Ching Chong, and Lay Ching Chai. 2016. "Isolation and Characterization of Aquatic-Borne Klebsiella pneumoniae from Tropical Estuaries in Malaysia" International Journal of Environmental Research and Public Health 13, no. 4: 426. https://doi.org/10.3390/ijerph13040426
APA StyleBarati, A., Ghaderpour, A., Chew, L. L., Bong, C. W., Thong, K. L., Chong, V. C., & Chai, L. C. (2016). Isolation and Characterization of Aquatic-Borne Klebsiella pneumoniae from Tropical Estuaries in Malaysia. International Journal of Environmental Research and Public Health, 13(4), 426. https://doi.org/10.3390/ijerph13040426