
Sex, Sport, IGF-1 and the Community Effect in Height Hypothesis


Abstract
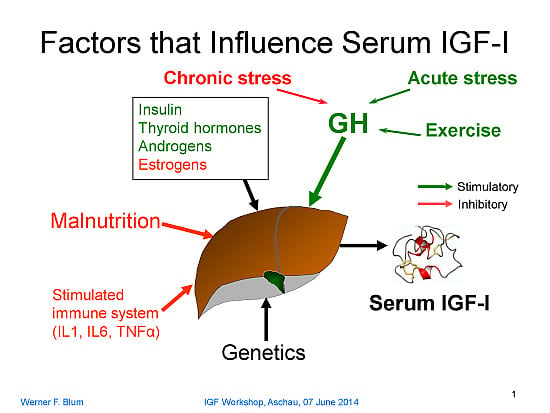
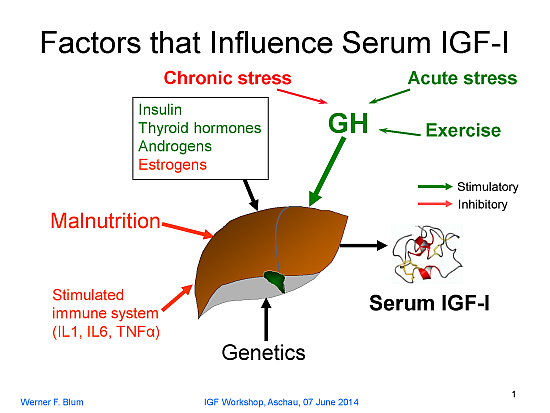
:
1. Introduction
2. Theoretical Background and Literature Review
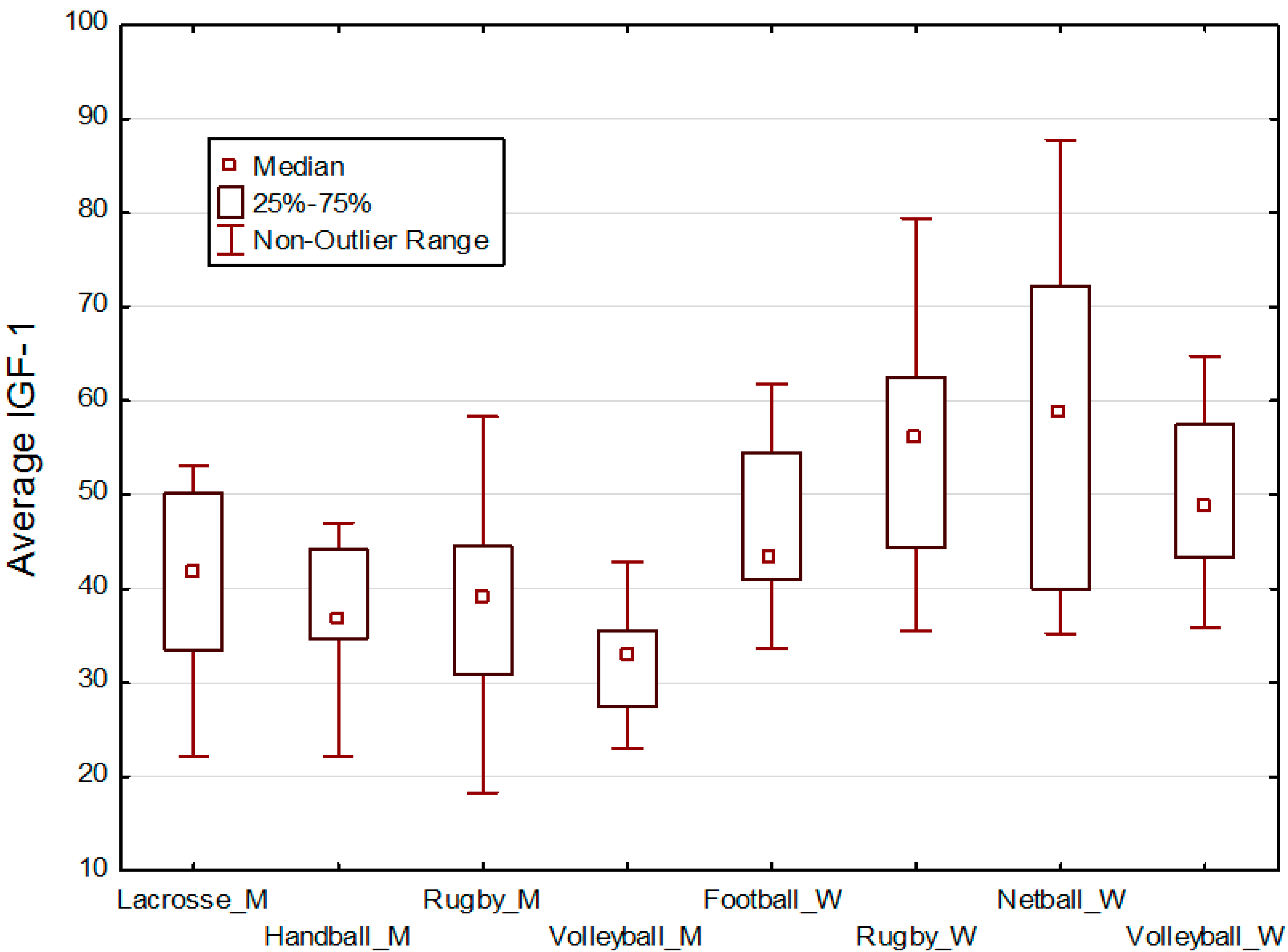
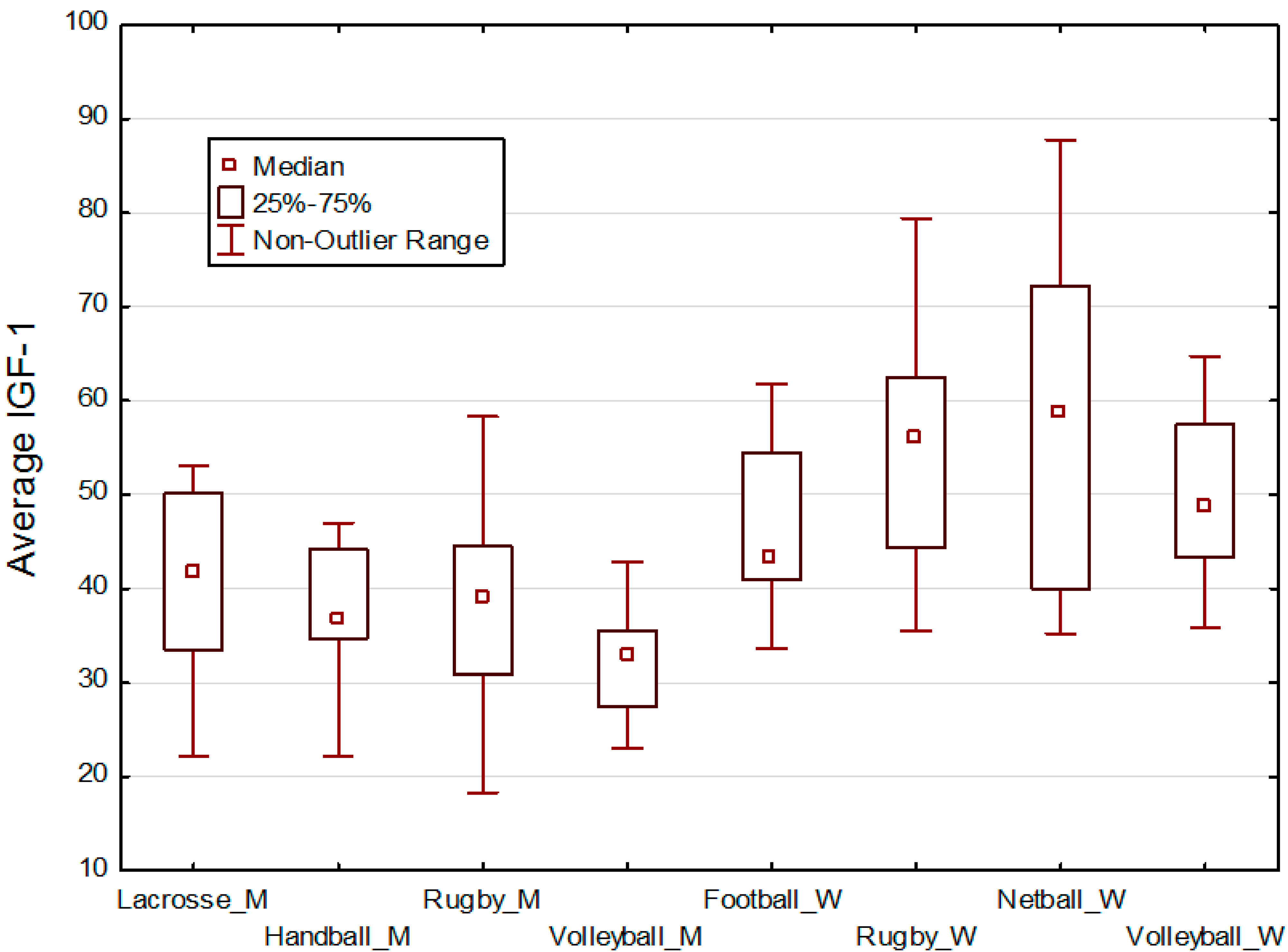
3. Hypotheses
4. Methods and Materials
4.1. Participants
4.2. Variables Measured
4.3. Ethical Clearance
4.4. IGF-1 Sampling and Analysis
5. Results

{kind=link}
{kind=link}
| Age, Years | Height, cm | Weight, kg | Pre-IGF-1 (µg/L) | Post-IGF-1 (µg/L) | |
|---|---|---|---|---|---|
| MEN (n = 71) | |||||
| Mean | 21.3 | 182.2 | 88.1 | 37.8 | 38.1 |
| SD | 1.5 | 6.4 | 10.6 | 9.9 | 10.7 |
| Min | 19 | 168.6 | 62.5 | 20.2 | 16.5 |
| Max | 25 | 195.0 | 109.0 | 66.6 | 60.0 |
| Age, Years | Height, cm | Weight, kg | Pre-IGF-1 (µg/L) | Post-IGF-1 (µg/L) | |
|---|---|---|---|---|---|
| WOMEN (n = 45) | |||||
| Mean | 20.1 | 171.0 | 69.2 | 53.0 | 54.3 |
| SD | 1.3 | 7.2 | 8.9 | 14.2 | 15.7 |
| Min | 18 | 157.0 | 56.8 | 27.5 | 33.0 |
| Max | 23 | 191.0 | 95.0 | 88.5 | 87.0 |
| Value | F; DFs | p | |
|---|---|---|---|
| Intercept | 0.89 | 2.95, 2, 50 | 0.06 |
| Age, years | 0.78 | 6.89 | 0.002 |
| SEX | 0.75 | 8.18 | 0.0008 |
| Height cm | 0.96 | 0.95 | 0.39 |
| Weight kg | 0.99 | 0.37 | 0.69 |
| SPORT | 0.91 | 2.53 | 0.09 |
6. Discussion
Limitations
7. Conclusions
GH secretion itself is regulated by endogenous signals coming from the central nervous system (e.g., the increase in GH during puberty) or by chronic psychosocial or physical distress (e.g., psychosocial growth retardation or extreme training load in high-performance athletes). In addition, the immune system may be involved and, when activated, it causes suppression of IGF-1… In the event that one of these major regulators is missing (GH or nutrients) or activated (immune system), there is resistance against the other factors with respect to IGF-1 production. That is, IGF-1 transmits integrated information at the cellular level on the nutritional status, the GH secretory status, and the immune status of the organism. Generally speaking, IGF-1 provides information to the cells on the well-being of the organism. Thus, the rate of cellular activities such as proliferation, differentiation, or the synthesis of cell-specific products is adapted to the situation. “Evidently, this kind of signal is of the utmost importance to the growing organism” (emphasis added).
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cameron, N.; Bogin, B. Human Growth and Development, 2nd ed.; Academic Press: London, UK, 2012. [Google Scholar]
- Bogin, B. Patterns of Human Growth, 2nd ed.; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Wijnhoven, T.; van Raaij, J.; Sjöberg, A.; Eldin, N.; Yngve, A.; Kunešová, M.; Starc, G.; Rito, A.; Duleva, V.; Hassapidou, M.; et al. WHO European childhood obesity surveillance initiative: School nutrition environment and body mass index in primary schools. Int. J. Environ. Res. Public Health 2014, 11, 11261–11285. [Google Scholar] [CrossRef] [PubMed]
- Bogin, B.; Varela-Silva, M.I. Leg length, body proportion, and health: A review with a note on beauty. Int. J. Environ. Res. Public Health 2010, 7, 1047–1075. [Google Scholar] [CrossRef] [PubMed]
- Hermanussen, M.; Bogin, B. Auxology–An editorial. Ital. J. Pediatr. 2014, 40. [Google Scholar] [CrossRef]
- Bogin, B.; Loucky, J. Plasticity, political economy, and physical growth status of Guatemala Maya children living in the United States. Am. J. Phys. Anthropol. 1997, 102, 17–32. [Google Scholar] [PubMed]
- Nazroo, J.Y. The structuring of ethnic inequalities in health: Economic position, racial discrimination, and racism. Am. J. Public Health 2003, 93, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Deaton, A. Height, health, and development. Proc. Natl. Acad. Sci. USA 2007, 104, 13232–13237. [Google Scholar] [CrossRef] [PubMed]
- López-Jaramillo, P.; López de Garcia, A.; Prevot, C.; Félix, C.; Sosa, C.; Romero, R.; Grijalva, Y.; Rappaport, R. Effect of social class and nutrient intake on height and plasma insulin-like growth factor in Andean Equadorian children. Eur. J. Clin. Nutr. 1992, 46, 137–142. [Google Scholar] [PubMed]
- Tanner, J.M. Growth as a mirror of the condition of society: Secular trends and class distinctions. Acta Paediatr. Jpn. 1987, 29, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Sikdar, M. Human Growth: The Mirror of the Society; B.R. Publishing Corporation: New Delhi, India, 2015. [Google Scholar]
- Blum, W.F.; Bottcher, C.; Wudy, S.A. Insulin-like growth factors and their binding proteins. In Diagnostics of Endocrine Function in Children and Adolescents; Ranke, M., Mullis, P., Eds.; Karger: Basel, Switzerland, 2011; pp. 157–182. [Google Scholar]
- Savage, M.O.; Burren, C.P.; Rosenfeld, R.G. The continuum of growth hormone-IGF-I axis defects causing short stature: Diagnostic and therapeutic challenges. Clin. Endocrinol. (Oxf.) 2010, 72, 721–728. [Google Scholar] [CrossRef]
- Hwa, V.; Fang, P.; Derr, M.A.; Fiegerlova, E.; Rosenfeld, R.G. IGF-I in human growth: Lessons from defects in the GH-IGF-I axis. Nestle Nutr. Inst. Workshop Ser. 2013, 71, 43–55. [Google Scholar] [PubMed]
- Leschek, E.W.; Rose, S.R.; Yanovski, J.A.; Troendle, J.F.; Quigley, C.A.; Chipman, J.J.; Crowe, B.J.; Ross, J.L.; Cassorla, F.G.; Blum, W.F.; et al. Effect of growth hormone treatment on adult height in peripubertal children with idiopathic short stature: A randomized, double-blind, placebo-controlled trial. J. Clin. Endocrinol. Metable 2004, 89, 3140–3148. [Google Scholar] [CrossRef]
- Kriström, B.; Lundberg, E.; Jonsson, B.; Albertsson-Wikland, K. IGF-1 and growth response to adult height in a randomized GH treatment trial in short non-GH-deficient children. J. Clin. Endocrinol. Metable 2014, 99, 2917–2924. [Google Scholar] [CrossRef]
- Albertsson-Wikland, K.; Aronson, A.S.; Gustafsson, J.; Hagenäs, L.; Ivarsson, S.A.; Jonsson, B.; Kriström, B.; Marcus, C.; Nilsson, K.O.; Ritzén, E.M.; et al. Dose-dependent effect of growth hormone on final height in children with short stature without growth hormone deficiency. J. Clin. Endocrinol. Metable 2008, 93, 4342–4350. [Google Scholar] [CrossRef]
- Alberti, C.; Chevenne, D.; Mercat, I.; Josserand, E.; Armoogum-Boizeau, P.; Tichet, J.; Léger, J. Serum concentrations of insulin-like growth factor (IGF)-1 and IGF binding protein-3 (IGFBP-3), IGF-1/IGFBP-3 ratio, and markers of bone turnover: Reference values for French children and adolescents and z-score comparability with other references. Clin. Chem. 2011, 57, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Rogers, I.; Metcalfe, C.; Gunnell, D.; Emmett, P.; Dunger, D.; Holly, J. Insulin-Like Growth Factor-I and Growth in Height , Leg Length , and Trunk Length between Ages 5 and 10 Years. J. Clin. Endocrinol. Metable 2006, 91, 2514–2519. [Google Scholar] [CrossRef]
- Mortier, G.R.; Vanden Berghe, W. Genomics, epigenetics and growth. In Human Growth and Development, 2nd ed.; Cameron, N., Bogin, B., Eds.; Academic Press: London, UK, 2012; pp. 153–171. [Google Scholar]
- Hendriks, A.E.J.; Brown, M.R.; Boot, A.M.; Oostra, B.A.; de Jong, F.H.; Drop, S.L.S.; Parks, J.S. Common polymorphisms in the GH/IGF-1 axis contribute to growth in extremely tall subjects. Growth Horm. IGF Res. 2011, 21, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Chrousos, G.P.; Gold, P.W. A healthy body in a healthy mind-and vice versa–The damaging power of “uncontrollable” stress. J. Clin. Endocrinol. Metable 1998, 83, 1842–1845. [Google Scholar]
- Batty, G.D.; Shipley, M.J.; Gunnell, D.; Huxley, R.; Kivimaki, M.; Woodward, M.; Lee, C.M.Y.; Smith, G.D. Height, wealth, and health: An overview with new data from three longitudinal studies. Econ. Hum. Biol. 2009, 7, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Bogin, B. Secular changes in childhood, adolescent and adult stature. Nestle Nutr. Inst. Workshop Ser. 2013, 71, 115–126. [Google Scholar] [PubMed]
- Christakis, N.A.; Fowler, J.H. Social contagion theory: Examining dynamic social networks and human behavior. Stat. Med. 2013, 32, 556–577. [Google Scholar] [CrossRef] [PubMed]
- Lyons, R. The Spread of Evidence-Poor Medicine via Flawed Social-Network Analysis. Stat. Polit. Policy 2011, 2, 1–22. [Google Scholar]
- Demongeot, J.; Taramasco, C. Evolution of social networks. Biogerontology 2014, 15, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Vanderweele, T.J.; Elizabeth, L.O.; Tchetgen, E.J.T. Why and When “Flawed” Social Network Analyses Still Yield Valid Tests of no Contagion. Stat. Polit. Policy 2012, 3, 2151–1050. [Google Scholar]
- Aßmann, C.; Hermanussen, M. Modeling determinants of growth: Evidence for a community-based target in height? Pediatr. Res. 2013, 74, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Hermanussen, M.; Alt, C.; Staub, K.; Groth, D. The impact of physical connectedness on body height in Swiss conscripts. Anthropol. Anzeiger 2014, 4, 313–327. [Google Scholar]
- Hermanussen, M.; Meitinger, T.; Veldhuis, J.D.; Low, M.J.; Pfäffle, R.; Staub, K.; Panczak, R.; Groth, D.; Brabec, M.; von Salisch, M.; et al. Adolescent Growth: Genes, Hormones and the Peer Group. In Proceedings of the 20th Aschauer Soiree, Glücksburg Castle, Germany, 15–17 November 2013.
- Tanner, J.M. Regulation of growth in size in mammals. Nature 1963, 199, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Rotoli, G.; Grignol, G.; Hu, W.; Merchenthaler, I.; Dudas, B. Catecholaminergic axonal varicosities appear to innervate growth hormone-releasing hormone-immunoreactive neurons in the human hypothalamus: The possible morphological substrate of the stress-suppressed growth. J. Clin. Endocrinol. Metable 2011, 96, 1606–1611. [Google Scholar] [CrossRef]
- Birmaher, B.; Dahl, R.E.; Williamson, D.E.; Perel, J.M.; Brent, D.A.; Axelson, D.A.; Kaufman, J.; Dorn, L.D.; Stull, S.; Rao, U.; et al. Growth hormone secretion in children and adolescents at high risk for major depressive disorder. Arch. Gen. Psychiatr. 2000, 57, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, M.; Bonafè, M.; Franceschi, C.; Paolisso, G. Insulin/IGF-I-signaling pathway: An evolutionarily conserved mechanism of longevity from yeast to humans. Am. J. Physiol. Endocrinol. Metable 2003, 285, E1064–E1071. [Google Scholar] [CrossRef]
- Stevens, A.; Hanson, D.; Whatmore, A.; Destenaves, B.; Chatelain, P.; Clayton, P. Human growth is associated with distinct patterns of gene expression in evolutionarily conserved networks. BMC Genomics 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Vera Cruz, E.M.; Brown, C.L. The influence of social status on the rate of growth, eye color pattern and Insulin-like Growth Factor-I gene expression in Nile tilapia, Oreochromis niloticus. Horm. Behav. 2007, 51, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Bartoš, L.; Reyes, E.; Schams, D.; Bubenik, G.; Lobos, A. Rank dependent seasonal levels of IGF-1, cortisol and reproductive hormones in male pudu (Pudu puda). Comp. Biochem. Physiol.-A Mol. Integr. Physiol. 1998, 120, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M.; Spencer, E.M. Insulin-like growth factor I is suppressed in socially subordinate male baboons. Am. J. Physiol. 1997, 273, R1346–R1351. [Google Scholar] [PubMed]
- Kumari, M.; Tabassum, F.; Clark, C.; Strachan, D.; Stansfeld, S.; Power, C. Social differences in insulin-like growth factor-1: Findings from a British birth cohort. Ann. Epidemiol. 2008, 18, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, R.; Farajis, H.; Sheikholeslami Vatani, D.; Qaderi, M. Creatine supplementation alters the hormonal response to resistance exercise. Kinesiology 2010, 42, 28–35. [Google Scholar]
- Schedel, J.M.; Tanaka, H.; Kiyonaga, A.; Shindo, M.; Schutz, Y. Acute creatine loading enhances human growth hormone secretion. J. Sports Med. Phys. Fitness 2000, 40, 336–342. [Google Scholar] [PubMed]
- Jørgensen, J.O.; Blum, W.F.; Møller, N.; Ranke, M.B.; Christiansen, J.S. Short-term changes in serum insulin-like growth factors (IGF) and IGF binding protein 3 after different modes of intravenous growth hormone (GH) exposure in GH-deficient patients. J. Clin. Endocrinol. Metab. 1991, 72, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Schütt, B.S.; Weber, K.; Elmlinger, M.W.; Ranke, M.B. Measuring IGF-I, IGFBP-2, and IGFBP-3 from dried blood spots on filter paper is not only practical but also reliable. Growth Horm. IGF Res. 2003, 13, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Blum, W.F.; Breier, B.H. Radioimmunoassays for IGFs and IGFBPs. Growth Regul. 1994, 4, 11–19. [Google Scholar] [PubMed]
- Blum, W.F.; Shavrikova, E.P.; Edwards, D.J.; Rosilio, M.; Hartman, M.L.; Marín, F.; Valle, D.; Van Der Lely, A.J.; Attanasio, A.F.; Strasburger, C.J.; et al. Decreased quality of life in adult patients with growth hormone deficiency compared with general populations using the new, validated, self-weighted questionnaire, Questions on Life Satisfaction Hypopituitarism Module. J. Clin. Endocrinol. Metable 2003, 88, 4158–4167. [Google Scholar] [CrossRef]
- Bray, I.; Gunnell, D.; Holly, J.M.P.; Middleton, N.; Smith, G.D.; Martin, R.M. Associations of childhood and adulthood height and the components of height with insulin-like growth factor levels in adulthood: A 65-year follow-up of the Boyd Orr cohort. J. Clin. Endocrinol. Metable 2006, 91, 1382–1389. [Google Scholar] [CrossRef]
- Gunnell, D.; Miller, L.L.; Rogers, I.; Holly, J.M.P. Association of insulin-like growth factor I and insulin-like growth factor-binding protein-3 with intelligence quotient among 8- to 9-year-old children in the Avon Longitudinal Study of Parents and Children. Pediatrics 2005, 116, 681–686. [Google Scholar] [CrossRef]
- Hardy, L.; Jones, G.; Gould, D. Understanding Psychological Preparation for Sport: Theory and Practice of Elite Performers; Wiley: Chichester, UK, 1996. [Google Scholar]
- Massuça, L.M.; Fragoso, I.; Teles, J. Attributes of top elite team-handball players. J. Strength Cond. Res. 2014, 28, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Young, M. Psychological Characteristics of Elite Athletes. Available online: http://www.elitetrack.com/article_files/sportpsych.pdf (accessed on 23 February 2015).
- Rowell, T.E. The concept of social dominance. Behav. Biol. 1974, 11, 131–154. [Google Scholar] [CrossRef] [PubMed]
- Hawley, P. The ontogenesis of social dominance: A strategy-based evolutionary perspective. Dev. Rev. 1999, 19, 97–132. [Google Scholar] [CrossRef]
- Creel, S. Social dominance and stress hormones. Trends Ecol. Evol. 2001, 16, 491–497. [Google Scholar] [CrossRef]
- Radley, J.J.; Kabbaj, M.; Jacobson, L.; Heydendael, W.; Yehuda, R.; Herman, J.P. Stress risk factors and stress-related pathology: Neuroplasticity, epigenetics and endophenotypes. Stress 2011, 14, 481–497. [Google Scholar] [CrossRef] [PubMed]
- Sävendahl, L. The effect of acute and chronic stress on growth. Sci. Signal. 2012, 5. [Google Scholar] [CrossRef] [PubMed]
- Stevens, A.; Clayton, P.; Tatò, L.; Yoo, H.W.; Rodriguez-Arnao, M.D.; Skorodok, J.; Ambler, G.R.; Zignani, M.; Zieschang, J.; Della Corte, G.; et al. Pharmacogenomics of insulin-like growth factor-I generation during GH treatment in children with GH deficiency or Turner syndrome. Pharmacogenomics J. 2014, 14, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Bogin, B. Childhood, adolescence, and longevity: A multilevel model of the evolution of reserve capacity in human life history. Am. J. Hum. Biol. 2009, 21, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Bogin, B.; Varela-Silva, M.I.; Rios, L. Life history trade-offs in human growth: Adaptation or pathology? Am. J. Hum. Biol. 2007, 19, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Komlos, J. Anthropometric history: An overview of a quarter century of research. Anthropol. Anz. 2009, 67, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.A. Evolution: A View from the 21st Century; FT Press Science/Pearson Education: Saddler River, NJ, USA, 2011. [Google Scholar]
- Widmann, P.; Reverter, A.; Fortes, M.R.S.; Weikard, R.; Suhre, K.; Hammon, H.; Albrecht, E.; Kuehn, C. A systems biology approach using metabolomic data reveals genes and pathways interacting to modulate divergent growth in cattle. BMC Genomics 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Deldicque, L.; Louis, M.; Theisen, D.; Nielens, H.; Dehoux, M.; Thissen, J.P.; Rennie, M.J.; Francaux, M. Increased IGF mRNA in human skeletal muscle after creatine supplementation. Med. Sci. Sports Exerc. 2005, 37, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Ronis, M.J.J.; Wands, J.R.; Badger, T.M.; de La Monte, S.M.; Lang, C.H.; Calissendorff, J. Alcohol-induced disruption of endocrine signaling. Alcohol. Clin. Exp. Res. 2007, 31, 1269–1285. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, K.; Lillycrop, K.; Burdge, G.; Gluckman, P.; Hanson, M. Non-imprinted epigenetics in fetal and postnatal development and growth. Nestle Nutr. Inst. Workshop Ser. 2013, 71, 115–126. [Google Scholar] [PubMed]
- Haig, D.; Wharton, R. Prader-Willi syndrome and the evolution of human childhood. Am. J. Hum. Biol. 2003, 15, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Thayer, Z.M.; Kuzawa, C.W. Biological memories of past environments: Epigenetic pathways to health disparities. Epigenetics 2011, 6, 798–803. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bogin, B.; Hermanussen, M.; Blum, W.F.; Aßmann, C. Sex, Sport, IGF-1 and the Community Effect in Height Hypothesis. Int. J. Environ. Res. Public Health 2015, 12, 4816-4832. https://doi.org/10.3390/ijerph120504816
Bogin B, Hermanussen M, Blum WF, Aßmann C. Sex, Sport, IGF-1 and the Community Effect in Height Hypothesis. International Journal of Environmental Research and Public Health. 2015; 12(5):4816-4832. https://doi.org/10.3390/ijerph120504816
Chicago/Turabian StyleBogin, Barry, Michael Hermanussen, Werner F. Blum, and Christian Aßmann. 2015. "Sex, Sport, IGF-1 and the Community Effect in Height Hypothesis" International Journal of Environmental Research and Public Health 12, no. 5: 4816-4832. https://doi.org/10.3390/ijerph120504816
APA StyleBogin, B., Hermanussen, M., Blum, W. F., & Aßmann, C. (2015). Sex, Sport, IGF-1 and the Community Effect in Height Hypothesis. International Journal of Environmental Research and Public Health, 12(5), 4816-4832. https://doi.org/10.3390/ijerph120504816